
Abstract
Alpine and Apennine glaciers contain only a small part of the total ice mass existing on Earth and it is well known that these ice masses probably are going to disappear in the next future. The study of biodiversity and ecology of cold-adapted yeasts living in cold habitats of Alpine and Apennine glaciers (supraglacial and subglacial sediments, cryoconite, snow, ice, and melt water) represents a contribution to obtain a better defined picture of the microbial ecology of these peculiar ecosystems. Most frequently isolated yeast species belonged to the genera Cryptococcus (mainly of the order Filobasidiales) and Rhodotorula (mainly of the class Cystobasidiomycetes). The majority of strains exhibited psychrotolerant characteristics and some of them were able to secrete extracellular hydrolytic cold-active enzymes. From an ecological point of view, some yeasts species could be subjected to an in situ selective enrichment in both supraglacial and subglacial sediments and cryoconite. Due to their heterotrophic nature, the possible role of cold-adapted yeasts in biogeochemical cycling through the mineralization of organic matter in cold environments is discussed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Arctic and Antarctic regions have been studied since the 1950s as reservoir of psychrophilic and psychrotolerant microorganisms (Abyzov 1993; Skidmore et al. 2000; Deming 2002). Bacteria were firstly considered but eukaryotic populations were also investigated in depth more recently (Buzzini et al. 2012). Some decades later, Alpine cold regions attracted the early interest of a few research groups (Margesin et al. 2002; Margesin 2009; Lazzaro et al. 2012). The limited extension of the Alpine chain and its characteristic geo-morphological conformation confer to this area some peculiar ecological traits in comparison with the complex Arctic and Antarctic surface.
The Alps are one of the great mountain range systems of Europe. Their length is approximately 1,200 km, the highest mountain is Mont Blanc (4,810 m a.s.l.) which is placed on the Italian-French border, whereas the whole Alpine area contains many peaks higher than 4,000 m. Alpine glaciers contain only a small part of the total ice mass existing on Earth. A few authors calculated that they lost about 35 % of their total area from 1850 until the 1970s, and almost 50 % by 2000. A total glacier volume of more than 200 km3 was estimated around 1850, whereas now only about one-third of this value is left. It was predicted that in the Alps most small glaciers (80 % of total glacial coverage, which represents an important contribution to local water resources) could disappear in the next decades (Cannone et al. 2008). In this context, global warming has a strong impact on Alpine mountains and a dramatic situation for future glacier developments including complete deglaciation of entire European mountain ranges can be depicted (Zemp et al. 2006).
Two different geo-morphological typologies of glaciers can be found in the European Alps: (1) the first is characterized by two or more different accumulation basins generating two or more ice flows usually ending in a common terminal plateau; (2) the second is represented by circular-shape glaciers (occasionally covered totally or in part by debris), which are usually smaller than the former and are representative of the majority of European Alpine glaciers.
The Apennines are a mountain chain consisting of parallel smaller chains extending around 1,200 km along the length of peninsular Italy. Their highest peak, Corno Grande (2,912 m a.s.l.), is partially covered by the Calderone glacier. With the disappearance of the Corral de la Veleta Glacier (Sierra Nevada, Spain) in 1913, the Calderone glacier became the southernmost one of Europe. Although still classified as a glacier, Calderone can be actually labeled as a debris-covered glacieret: a small ice mass of indefinite shape, placed over a protected slope that originated from snow drifting and/or avalanching, and exhibiting no marked ice flow pattern (Pecci et al. 2008). Global warming could particularly impact for the weak geo-morphological conditions of this small glacier, which is subjected to harsh fluctuations of temperature, and can be considered the most reliable witness of global warming (Pecci et al. 2008).
Because of this catastrophic scenario, all the studies on the biodiversity and ecology of microbial populations living in cold ecosystems of the Alps and the Apennines assume an increasing relevance: a temperature increase may determine an abundant loss of macro- and microbial diversity, thus influencing biogeochemical cycles. Additionally, ex situ conservation of microbial biodiversity collected in those ecosystems may be particularly interesting and represents a way to preserve species that are going to disappear, for future studies and utilization. An interesting description of the ex situ preservation of cold-adapted yeasts in worldwide collections is reported in Chap. 2.
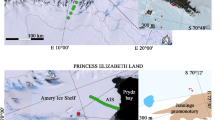
Yeast diversity of Alpine and Apennine habitats was studied only in a few sites (Table 5.1, Fig. 5.1). Within this general framework, the ecology of cold-adapted (psychrophilic and psychrotolerant) yeasts inhabiting various ecological niches, including supraglacial and subglacial sediments, ice, melting water, snow and cryoconite, has been considered. Additionally, some studies described yeast diversity and physiology of polluted cold soils, also in view of their ability for bioremediation purposes. In this Chapter, an overview of yeast biodiversity of Alpine and Apennine cold habitats is presented.

Geographic positions of the sample sites studied for yeast diversity in the Alps and Apennines
2 Cold-Adapted Yeasts in Alpine and Apennine Supraglacial and Subglacial Sediments and Soils
2.1 General Properties of Supraglacial and Subglacial Sediments
Glacial sediments are generally associated with both glacial surface and ice-bed interface (Hodson et al. 2008). Some glaciers exhibit a continuous debris mantle covering the surface of the lower part of their ablation zone (supraglacial sediments). Such glaciers are common in high-relief mountain environments where mass-wasting processes deliver large volumes of debris to glacial areas. Supraglacial sediments are generally composed of poorly sorted debris sized from both fine grain matrix to large rock boulders. The presence of thick supraglacial sediment accumulation may retard glacier ablation (Benn and Evans 1998). In contrast, subglacial sediments include rocks, sand, silt, and clay derived from glacier grinding up and mixing of rock and soil debris beneath its base. They are poorly sorted sediments exhibiting a little structure and often containing water micro-flow channels in pores within the matrix of the fine-grained material. Supraglacial and subglacial ecosystems may differ vastly in terms of their water activity (Aw), nutrient abundance, redox potential, rock-water contact, pressure, solar irradiation, and pH (Paterson 2002). Among them, the presence of a metabolically active microbial community in subglacial sediments is strictly dependent on the availability of water. Liquid water is vital for any ecosystem and the presence of water coming from rainfall and snowfall and ice melting may represent the source that provides adequate water activity for microbial metabolisms in glacial habitats (Hodson et al. 2008; Anesio et al. 2010; Anesio and Laybourn-Parry 2012). Liquid water can occur at the bed of temperate glaciers (characterized by ice at the pressure and temperature close to melting point, which is typical of the Italian Alps) that additionally show abundant rock debris derived from ice ablation (Anesio and Laybourn-Parry 2012).
Recent studies have shown that supraglacial and subglacial sediments can act as suitable substrate for microbial growth and metabolism; this is apparently confirmed also by the positive correlation found between microbial abundance and concentration of sediments in ice and melt water (Sharp et al. 1999). However, the role of the microbial community in these habitats is still partially unclear.
Microbial communities inhabiting surface and deep habitats of worldwide glaciers have been extensively studied: the presence of algae and cyanobacteria performing photosynthetic metabolism and of aerobic heterotrophic prokaryotic and eukaryotic populations contributing to the cycle of organic matter was observed in supraglacial sediments (Hodson et al. 2008, 2010). Microorganisms in supraglacial sediments could derive mainly from air movement, snowfall, and rainfall, although only a small percentage could be able to survive under such extreme conditions (Simon et al. 2009). In contrast, in subglacial ecosystems, the predominance of chemoautotrophic prokaryotes and heterotrophic prokaryotes and eukaryotes, that even survive under aerobic conditions, has been observed (Sharp et al. 1999; Tranter et al. 2002; Skidmore et al. 2000, 2005; Foght et al. 2004; Priscu et al. 1999; Christner et al. 2001, 2006; Miteva et al. 2004). Some interesting information on forefields of receding Alpine glaciers (i.e., new terrain that becomes exposed due to the glacial retreat) has been recently reported by Lazzaro et al. (2012) and Zumsteg et al. (2012). According to both authors, microorganisms colonizing such forefields represented an exclusive biome, and it was suggested that these microorganisms could be involved in the formation of new soil, with a definite microbial succession from archaea to prokaryotes to eukaryotes. Their ability to interact with minerals and organic compounds through physical and metabolic processes makes them contributors to rock weathering (Zumsteg et al. 2012).
In recent years, the yeast community of unfrozen subglacial sediments of the glaciers in the Ortles-Cevedale complex and the one of the supraglacial sediments of both the Alps and Apennines (Miage and Calderone glaciers, respectively) have been investigated (Turchetti et al. 2008; Branda et al. 2010; Turchetti et al. 2013). Besides, dirt cones from Calderone glacier (Apennines) were also studied: these peculiar sediments originate from the melt out of either endoglacial or subglacial debris bands (Branda et al. 2010). These phenomena are due to differential ablation which produces such upstanding morphologies on the glacial surface. Dirt cones can also derive from deep-piping sediments originating from intra-glacier high water pressure. In both cases, dirt cones are composed predominantly of ice, water, and debris (Benn and Evans 1998).
2.2 Abundance of Cold-Adapted Yeasts in Supraglacial and Subglacial Sediments of Alps and Apennines
Overall, the numbers of culturable yeasts in Alpine and Apennine supra- and subglacial sediments were lower by one to several orders of magnitude than those of bacteria and filamentous fungi. In particular, numbers of yeast cells ranging from 102 to 103 CFU g−1 dry weight (DW) were observed in supraglacial sediments and dirt cones of the Calderone glacier, whereas the presence of culturable yeasts in supraglacial sediments of the Ortles-Cevedale complex was less abundant (about a few tens CFU g−1 DW), thus two orders of magnitude lower than in the corresponding subglacial habitat (102–103 CFU g−1 DW). Interestingly, similar amounts of total organic carbon, nitrogen, and phosphorous were observed in supraglacial and subglacial sediments of the Ortles-Cevedale complex, while the content of organic compounds in supraglacial sediments and dirt cones of the Calderone glacier was 10-fold (carbon and nitrogen) or 1000-fold (phosphorous) more concentrated (Turchetti et al. 2008; Branda et al. 2010).
2.3 Diversity and Ecology of Cold-Adapted Yeasts in Supraglacial and Subglacial Sediments of Alps and Apennines
A total of fifty different yeast species were found in Alpine and Apennine sediments: almost all were basidiomycetes. Cryptococcus was the prevalent genus with species belonging to both Filobasidiales and Tremellales orders (Table 5.2) (Turchetti et al. 2008; Branda et al. 2010; Turchetti et al. 2013). The prevalence of Cryptococcus species is consistent with previous studies carried out in worldwide glacial areas (Butinar et al. 2007; de Garcia et al. 2007; Connell et al. 2008). The high percentage of Cryptococcus strains could be justified by their ability to produce an extracellular capsule, which could result in a protection from external extreme conditions (Vishniac 2006). Other adaptation mechanisms, which could confer to this genus a superior ability to thrive and even to grow in such ecosystems (e.g., a wider carbon and nitrogen assimilation pattern), were also recently hypothesized (Connell et al. 2008).
The most frequently isolated species were Cryptococcus gilvescens, Cryptococcus gastricus, Cryptococcus victoriae, Rhodotorula psychrophenolica (Fig. 5.2), Mrakia robertii, Cryptococcus terricola, and Cryptococcus antarcticus. Around 15–20 % of the species resulted psychrophilic, whereas the remaining ones were psychrotolerant (Turchetti et al. 2008, 2013; Branda et al. 2010). The large part of the species observed in sediments of Alps and Apennines have been previously regarded as exclusive inhabitants of cold environments (Butinar et al. 2007; de Garcia et al. 2007; Connell et al. 2008). Interestingly, a few cosmopolitan species, namely Cryptococcus fuscescens, Rhodotorula hordea, and Holtermanniella takashimae, never been isolated so far from glacial ecosystems, were found in Alpine and Apennine sediments.

Cells of Rhodotorula psychrophenolica DBVPG 4792 after 2 weeks of incubation at 10 °C on malt extract agar (MEA), showing budding cells and pseudohyphae. Bar, 5 μm
A heterogeneous distribution of the different yeast species was observed in sediments collected in different geographical areas. C. gilvescens was the dominant species (58 % of the total isolated strains) in sediments sampled in the Ortles-Cevedale complex, but this species was absent in sediments of the Calderone glacier. On the contrary, C. gastricus was the dominant species in sediments of the Calderone glacier (49 %), but it was lacking in sediments sampled in the glaciers of the Ortles-Cevedale complex (Turchetti et al. 2008; Branda et al. 2010). Furthermore, both species were found in sediments of Mont Blanc glaciers (Turchetti et al. 2013). C. gilvescens is phenotypically undistinguishable from C. gastricus: the analysis of the D1/D2 domain of 26S rRNA and ITS1&2 sequences showed that both species occupy distinct positions within the Filobasidiales lineage, but with a strong phylogenetic relation (they differ by only three nucleotide substitutions in both regions) (Fonseca et al. 2011). Both species appear to be regular inhabitants of cold environments (Fonseca et al. 2011). Their psychrotolerance, their ability to produce extracellular hydrolytic cold-active enzymes and their frequent occurrence in cold habitats could suggest their possible role in geochemical cycling of organic matter in supraglacial and subglacial sediments (Turchetti et al. 2008).
The species C. victoriae was isolated in the past from cold Arctic and Antarctic soils, but also from other temperate habitats (Herzberg et al. 2002; Gildemacher et al. 2004, 2006). Neighbor-joining analysis of D1/D2 sequences placed this species into the order Tremellales, particularly into the victoriae clade together with the species Cryptococcus carnescens, Cryptococcus heimaeyensis, and Cryptococcus tephrensis (Fonseca et al. 2011), which have been all frequently isolated from cold ecosystems.
A total of twenty-eight percentage of all strains isolated from Alpine and Apennine glaciers were found to belong to novel species: R. psychrophenolica and Rhodotorula glacialis (Margesin et al. 2007), Mrakiella cryoconiti (Margesin and Fell 2008), M. robertii, Mrakia blollopis and Mrakiella niccombsii (Thomas-Hall et al. 2010), Glaciozyma martinii and Glaciozyma watsonii (Fig. 5.3) (Turchetti et al. 2011).

Optical microscopy image of Glaciozyma watsonii DBVPG 4760 after 1 week of incubation at 10 °C on MEA, showing budding cells. Bar, 5 μm
Interestingly, the species M. niccombsii was recently found in supraglacial sediments of the Miage glacier (Turchetti et al. 2013). Since this species has so far been considered endemic of Antarctic soils (Vishniac 1985; Vishniac and Kurtzman 1992; Vishniac and Onofri 2002; Arenz et al. 2006; Vishniac 2006; Thomas-Hall et al. 2010), this evidence could lead to reconsider its status as endemic species.
Most of the yeasts isolated from supraglacial and subglacial sediments of Alpine and Apennine glaciers were common inhabitants of worldwide cold areas, such as Antarctica and the Arctic (Butinar et al. 2007; de Garcia et al. 2007; Connell et al. 2008; Libkind et al. 2009). Atmospheric circulation, via wind and dust particles, rainfall, and snowfall vectors, could be considered the most plausible way of their global dissemination (Xiang et al. 2009). Of course, the sole dissemination cannot be considered sufficient to justify the differential diversity of cold-adapted yeasts observed in geographically distinct cold habitats, because only some habitats could support their in situ survival and growth.
2.4 Diversity of Cold-Adapted Yeasts in Alpine Soils
Cold-adapted yeasts strains isolated from contaminated soils in the Kitzbühler Alps, Stubaier Alps, and at the Brenner pass were studied by Bergauer et al. (2005) and Margesin et al. (2005b). Eleven basidiomycetous species were identified; dominant genera were Cryptococcus and Rhodotorula. Some of them, namely R. glacialis, R. psychrophenolica, Cryptococcus terreus, and C. terricola, have been frequently found in Alpine environments. On the contrary, five other species were isolated from cold habitats for the first time, namely Leucosporidiella creatinivora (formerly Rhodotorula creatinivora), Rhodosporidium lusitaniae, Rhodotorula ingeniosa, Rhodotorula psychrophila, and Trichosporon dulcitum (Table 5.2). All the strains isolated from Alpine cold soils showed the ability to degrade phenolic compounds at low temperature (for more information, see Chap. 21).
3 Cold-Adapted Yeasts in Alpine and Apennine Ice Cores, Snow, and Melt Water
3.1 General Properties of Ice Cores, Snow, and Melt Water
Ice constituting glaciers is a substrate which can act as long-term, chronological repository of viable microorganisms. Due to harsh conditions (high pressure, darkness, acidity and low Aw), an englacial environment can be considered an extreme habitat (Price 2000, 2007). Some peculiar situations occurring within englacial ice can apparently simplify microbial life in this environment. For example, the part of englacial ice lying at the pressure of the melting point can determine the increase in the temperature and the Aw, thus increasing the possibility of microbial life and metabolic activity (Price 2007). Besides, the phenomenon labeled as “hydratation distance” at the surface of solid impurity or hydrophilic mineral substrates could determine the presence of an unfrozen layer of temperature-dependent thickness around particles, providing nutrients and water for microorganisms (Price 2007). In temperate glaciers, englacial habitats include crevasses and water channels (Fig. 5.4), which can act themselves as growing substrates for microbial communities and at the same time convey water, nutrients, atmospheric gases, and microbiota into the glacier (Price 2007).

Representation of a potential ecological model for the distribution of cold-adapted yeasts inside the ecosystem of a temperate glacier in the Alps and Apennines
Microbial biodiversity of worldwide icy habitats has attracted the interest of an increasing number of biologists. Early studies, conducted by Abyzov et al. (1982, 2001), found diverse microorganisms including cold-adapted yeasts (belonging to the species Cryptococcus albidus and Rhodotorula mucilaginosa) in the deep ice cores drilled from Lake Vostok, Antarctica. Likewise, Skidmore et al. (2000) demonstrated the presence of aerobic chemoheterotrophs, anaerobic nitrate- and sulfate-reducing bacteria and methanogens in the debris-rich basal ice layers and the surface of high Arctic glaciers. Later, englacial ecosystems were also studied in glacier ice cores extracted from different geographic areas: the presence of diverse bacterial communities including representatives of Proteobacteria, Bacteroidetes (formerly Cytophaga–Flavobacterium–Bacteroides) and Gram-positive bacteria was observed (Christner et al. 2003b). The increasing awareness that a large number of viable microorganisms, including yeasts, are entombed within glacial ice enables the hypothesis that englacial microbial populations could play an important role in nutrient cycling inside glaciers and ice sheets. However, despite the presence of viable microorganisms in ice cores (Castello and Rogers 2005; Price 2007), their in situ activity remains to be fully elucidated.
Snowpack covering ice surface is another interesting niche characteristic of glacial environments that could act as reservoir of different types of microorganisms. Snow accumulates on the surface of icy surface and contains an extremely low concentration of organic matter. The ecological role of snow is significant since it provides an important inoculation of microbial cells, nutrient, and water that enriches the glacial ecosystems beneath the snow superficial layer (Fig. 5.4). Microorganisms occurring on surface snow are often exposed to light, nutrients, and temperatures of around 0 °C in summer, which facilitates the growth of phototrophic cyanobacteria and algae and can result in the presence of colorful snow coverage (Stibal et al. 2007). Hodson (2006) found a high concentration of inorganic nitrogen and phosphate in such habitat due to the phototrophic activity of algae and cyanobacteria. By contrast, microbial communities in the deep snow are exposed to abrupt temperature drop and low light intensity, which reduces the growth rate of light-dependent microorganisms (Xiang et al. 2009). Besides, complex bacterial consortia were found in snowpack covering the surface of a few glaciers located in the Alps, Andes, and Svalbard archipelago (De Angelis and Gaudichet 1991; Amato et al. 2007).
Sun irradiation provokes snow and ice melting due to an increase in temperature, albedo effect, and strong pressure on the glacier bed. The different origin of melt water makes it difficult to define a general idea about the characteristics of this peculiar substrate (Paterson 2002):
-
1.
streaming of melt water on the surface of the glacier derived from ice and snow ablation during summer time;
-
2.
percolation of water from the surface into ice;
-
3.
ice melting induced by pressure change and streaming in subglacial sediments or creating channels within ice;
-
4.
water trapped inside pockets originated from ice formation;
-
5.
lakes located close to the margins of the glacier, or on the glacier, originating from the melt water above described.
The abundance of microorganisms in melt water of worldwide temperate glaciers has been sporadically described (Sharp et al. 1999; Butinar et al. 2007; de Garcia et al. 2007) because more emphasis has been devoted to sediments and ice. The currently accepted hypothesis considers melt water as an important connection between different niches within the same glacier (or even as a natural link between glacial habitat and glacier forefield). Variable microbial abundance has been detected in melt water collected in glaciers of the Svalbard archipelago: bacterial numbers were about 105 cells ml−1 (Sharp et al. 1999), whereas yeast numbers ranged from 10 to 103 CFU ml−1 (Butinar et al. 2007).
3.2 Abundance of Cold-Adapted Yeasts in Ice Cores, Snow, and Melt Water of Alps and Apennines
According to recent studies, ice, snow, and melt water can be considered as extremely oligotrophic habitats in comparison with both supraglacial and subglacial sediments. This could apparently justify the lower yeast cells abundance (10–102 CFU L−1), if compared with the corresponding sediments (Turchetti et al. 2008; Branda et al. 2010).
These results are consistent with the data observed in melt water sampled from Patagonian glaciers (de Garcia et al. 2007; Libkind et al. 2009). Surprisingly, ice cores and snow sampled from Alpine and Apennine glaciers showed a yeast abundance that was 10-fold to 100-fold lower than that observed in corresponding sediments, in contrast with the results reported for Svalbard ice cores (10–103 CFU ml−1) (Butinar et al. 2007).
3.3 Diversity and Ecology of Cold-Adapted Yeasts in Ice Cores, Snow, and Melt Water of Alps and Apennines
The yeast diversity observed in ice cores collected from Alpine and Apennine glaciers partially overlapped with that observed in the corresponding supraglacial and subglacial sediments, i.e., C. gilvescens and C. gastricus, respectively, were the dominant species, although their relative abundance was 10-fold to 100-fold lower (Turchetti et al. 2008; Branda et al. 2010).
In contrast, snowpack covering the surface of the glacier du Geant (Mont Blanc) showed a wider yeast biodiversity spectrum: 38 different species were identified (Turchetti et al. 2013). C. victoriae (28 % of all isolated strains), R. psychrophenolica (24 %), the yeast-like species Aureobasidium pullulans (7 %), and C. gilvescens (6 %) were the most frequently found species. Some of them are regularly observed in worldwide cold environments, while some other are regarded as ubiquitous species (e.g., A. pullulans, Cryptococcus chernovii, Cryptococcus diffluens, and R. laryngis) (Turchetti et al. 2013). Their deposition on snow surface by air current, snowfalls, and rainfalls and/or their possible variable ability to survive (or even to grow) under these conditions could be the basis justifying their differential occurrence.
A large number of yeast species, already described as regular inhabitants of both supraglacial and sub-glacial sediments, were found in melt water sampled in Alpine and Apennine glaciers. The species R. glacialis and C. gilvescens represented over 50 and 20 %, respectively, of the yeasts isolated from meltwater collected in the Ortles-Cevedale complex (Turchetti et al. 2008). In contrast, Cystofilobasidium macerans and Cystofilobasidium capitatum, which were also found to be ubiquitous in water streams or glacial lakes (Sampaio 2011), were the predominant species (19 % and 17 %, respectively) in meltwater collected from the Apennine Calderone glacier (Branda et al. 2010).
4 Cold-Adapted Yeasts in Alpine Glacier Cryoconite
The term “cryoconite” (ice dust) was used for the first time by Nordenskjöld during his 1870 Greenland expedition and refers to small (0.1–3 mm in diameter) dark-colored wind-borne particles at the bottom of cryoconite holes. These particles consist of inorganic (e.g., mineral components of soils and sediments) and organic (e.g., litter, pollen, Bacteria, Eukarya) materials. Cryoconite holes are water-filled depressions (up to 50 cm deep and from a few centimeters to >1 m in diameter) on the glacial surface and form when the dark particles deposited on the glacier surface are warmed by solar radiation (black body effect) and melt into the underlying ice because of the lower albedo of cryoconite compared to the surrounding ice (Wharton et al. 1985; Takeuchi et al. 2001; Cameron et al. 2012). The reduced surface albedo may play an important role in the acceleration of glacier melting. Cryoconite holes are found worldwide in high-latitude and high-alpine glaciers (MacDonell and Fitzsimons 2008) as well in glaciers of temperate areas; they can cover 0.1–10 % of the glacier surface (Anesio et al. 2009). Most cryoconite holes are open to the environment, while those in Antarctic Dry Valley glaciers are covered by an ice lid and thus isolated from the atmosphere (Miteva 2008).
4.1 Properties of Alpine Cryoconite
The temperature in Alpine cryoconite ranges from about ≤0 °C to about +3 °C in summer (July to September), as it is permanently saturated with cold meltwater from ablating snow patches. During the residual time of the year, cryoconite is snow-covered and permanently frozen. Freezing also can occur every night during summer (Margesin et al. 2002). Cryoconite holes could serve as biological refuges during extended periods of subzero temperatures; members of cryoconite communities that survive these conditions might ensure the reseeding of the surrounding environment during warmer periods (Christner et al. 2003a).
Cryoconite contains substantial amounts of nutrients, which supports microbial growth and activity. Three Alpine cryoconite samples collected from an Austrian glacier (Margesin et al. 2002) were characterized by a pH value of 5, a temperature of 1.5–3 °C at the time of sampling (September), and a water content of 50 %, and contained 0.8–1.8 % (related to cryoconite dry mass) total carbon and 0.02–0.09 % total nitrogen. The C:N ratio ranged from 40:1 to 20:1. The CaCO3 content was in the range of 1.9–2.1 % (corresponding to 0.2–0.4 % inorganic carbon). Alpine cryoconites of an Austrian glacier contained high amounts of anthropogenic radionuclides from nuclear weapons and Chernobyl fallouts (Tieber et al. 2009). In comparison, Arctic cryoconite contained 1.1–1.9 % organic C and had a water content of 67–79 % (Singh and Singh 2012). Most of the organic carbon in Arctic cryoconite holes originates from allochthonous rather than indigenous primary production (Stibal et al. 2008).
In some areas (e.g., glacier-skiing areas), the composition of cryoconite is strongly influenced by human presence. Studies on the influence of human impact on microbial community composition of Alpine cryoconite samples collected from six Austrian glaciers indicated the introduction of allochthonous bacterial species, as shown by the presence of isolates with higher maximum growth temperatures on sites with strong human impact compared to sites without or with low human impact (Lee et al. 2011). Such data have not yet been gathered for yeast population, however, it is likely that a similar result can be obtained.
4.2 Diversity and Ecology of Cold-Adapted Yeasts in Alpine Cryoconite
Since cryoconite holes contain abundant populations of actively living organisms, Wharton et al. (1981, 1985) pointed already 30 years ago to the important role of cryoconite holes in glacier ecosystems. During the last decade, cryoconite holes have been recognized as an important microbial ecosystem (Hodson et al. 2008; Anesio and Laybourn-Parry 2012). Every cryoconite hole is unique and therefore may support a novel and discrete ecosystem (Christner et al. 2003a). These supraglacial mini-ecosystems are considered “hot spots” of microbial metabolic activity, which explains the strong interest in their biogeochemical processes, chemical gradients, and nutrient cycling (Margesin and Miteva 2011; Singh and Singh 2012). Diversity studies of microbial communities in cryoconite holes demonstrated the presence of photosynthetic cyanobacteria and algae, heterotrophic bacteria, yeasts and filamentous fungi, metazoa and viruses (Wharton et al. 1985; Christner et al. 2003a; Anesio et al. 2009; Stibal et al. 2012). Archaeal communities have so far only been detected in Antarctic samples (Cameron et al. 2012).
Members of individual cryoconite communities were often found to be specific to individual locations, suggesting that they may be seeded primarily via localized wind transportation and/or during glacial melt from adjacent surrounding environments (Christner et al. 2003a; Cameron et al. 2012). Long-distance wind transport may provide a mechanism for immigration into cryoconite holes. Cryoconite microorganisms may also play a role in the colonizing of newly exposed areas after glacier retreats (Wharton et al. 1985).
There is little information on the abundance and role of yeasts in cryoconite (Margesin et al. 2002; Singh and Singh 2012; Uetake et al. 2012). Three Alpine cryoconite samples collected from an Austrian glacier contained predominantly culturable aerobic heterotrophic bacteria, but culturable yeasts were also present and were identified as representatives of the genera Cryptococcus, Candida, and Rhodotorula (Margesin et al. 2002). Cryptococcus gilvescens dominated among Cryptococcus species (Margesin, unpublished data). The description of novel cold-adapted bacterial and yeast (Rhodotorula glacialis, R. psychrophenolica, and Mrakiella cryoconiti) taxa from Alpine glacier cryoconite (Margesin et al. 2007; Margesin and Fell 2008) indicates the typical community composition of cryoconite (Margesin and Miteva 2011). The difference between the abundance of bacteria and yeasts was influenced by temperature: significantly higher numbers of yeasts were able to grow at 2 °C (2 x 10³ to 2 × 105 CFU g−1 cryoconite dry mass) than at 20 °C (5 × 10² to 3 × 104 CFU g−1 dry mass) in all three cryoconite samples. Yeast populations were about 5–10-fold lower than bacterial numbers at 2 °C, but more than 100-fold lower at 20 °C (Margesin et al. 2002).
Many cryoconite microorganisms show remarkably good cold adaptation with regard to growth characteristics and metabolic activities (Margesin and Miteva 2011). The maximum temperatures for growth of yeast strains from Alpine cryoconite is usually 15 °C or 20 °C, while only few can grow at 25 °C (Margesin et al. 2002, 2007; Margesin and Fell 2008). These data and further studies led to the assumption that cold-adapted yeasts have a more restricted growth temperature range than cold-adapted bacteria (Margesin 2009).
The ability of cold-adapted yeasts to grow even below the freezing point of water (Panikov and Sizova 2007) indicates their capability to degrade organic compounds at low temperatures (Buzzini et al. 2012). Cryoconite microorganisms convert organic matter into dark-colored humic substances, which results in increased light absorbance (Takeuchi et al. 2001). Yeasts from Alpine glacier cryoconite utilize various organic compounds (cellulose, starch, tributyrin, polygalacturonic acid, diesel oil, and phenol) as the sole carbon and energy source (Margesin et al. 2002, 2005a; Bergauer et al. 2005).
Substrate utilization was more influenced by the cultivation temperature than by the composition of the substrate. All of 14 yeast strains investigated utilized the tested substrates when cultured at 5, 10, or 15 °C. The relative substrate utilization was higher at 0 °C than at temperatures above 15 °C, at which significantly reduced substrate utilization was noticed. The optimum temperature for biomass production and substrate hydrolysis is usually lower than the optimum growth temperature of cold-adapted bacteria and yeasts (Feller et al. 1996; Margesin 2009). This could also be demonstrated with a cold-active alkaline pectinase (pectate lyase) produced by Mrakia frigida from Alpine glacier cryoconite (Margesin et al. 2005a), later identified as a representative of a novel genus (Mrakiella cryoconiti; Margesin and Fell 2008).
5 Contribution of Yeast Populations to the Ecology of Alpine and Apennine Cold Habitats
Cold-adapted microorganisms colonizing glacier ecosystems are subjected to extreme low temperatures and Aw, and often to nutrient limitation. Such organisms are adapted to these extreme environments through some modifications of their metabolic processes, such as production of cold-active enzymes, anti-freezing proteins, and exopolymeric substances that provide cell protection against the potentially damaging effects of low temperatures. Besides, they maintain the fluidity of cell membranes until sub-zero temperatures by an increase in the proportion of unsaturated fatty acids and a decrease in the average fatty acid chain length (Siddiqui and Cavicchioli 2006; Buzzini et al. 2012). An exhaustive overview of the strategies adopted by cold-adapted yeasts to overcome the negative effect of cold is reported in Chaps. 9, 10, 11, 12, 13, 14, 15.
In glacier ecosystems, the nutrient input (especially in terms of carbon and nitrogen) derives principally from wind and alluvial deposition of allochtonous particulate material, which can release both inorganic and organic compounds, the last due to decomposing processes of organic materials catalyzed by microbial biomass (Anesio and Laybourn-Parry 2012). Both supraglacial and subglacial sediments are actively colonized by chemoautotrophic and heterotrophic microorgansims including cold-adapted yeasts that are able to make use of recalcitrant substrates more efficiently than bacteria. There is additional evidence that organic compounds in glacial sediments can be easily hydrolyzed aerobically bringing subglacial sediment to anoxia and guiding to the development of methanogens (Wadham et al. 2008). Microbial communities sharing glacial habitats could be responsible of some biogeochemical transformations, with some important implications at both local and global scales (Anesio and Laybourn-Parry 2012).
Chemical analysis of sediments and cryoconite (and to a lesser extent of ice, snow, and melt water) collected from Alpine and Apennine glaciers demonstrated the presence of organic carbon, nitrogen, and phosphorus in amounts that support survival and even growth of psychrophilic and psychrotolerant heterotrophic microorganisms including yeasts (Margesin et al. 2002; Turchetti et al. 2008; Branda et al. 2010). The majority of yeasts isolated from Alpine and Apennine glaciers exhibited the ability to secrete cold-active hydrolytic enzymes (Margesin et al. 2005a; Turchetti et al. 2008; Branda et al. 2010). These enzymes could be involved in the hydrolysis of organic macromolecules naturally present in these harsh environments (Anesio and Laybourn-Parry 2012).
Considering the above general findings and under the light of the studies on cold-adapted yeasts in Alpine and Apennine ecosystems (Sharp et al. 1999; Turchetti et al. 2008; Branda et al. 2010), it is possible to postulate that glacial habitats constitute an important reservoir of metabolically active heterotrophic prokaryotic and eukaryotic microbial communities. Although some important differences on abundance and biodiversity of yeast populations between eutrophic (i.e., sediments and cryoconite) and oligotrophic (ice, snow, melt water) habitats have been observed (Turchetti et al. 2008; Branda et al. 2010), it can be postulated that cold-adapted yeasts are able to survive (and even to grow) under such extreme conditions. In this framework, studies reporting their ability to perform some important biogeochemical activities (such as organic matter decomposition and nutrient cycling) at low temperature could be considered an important milestone in the knowledge of the in situ ecological role of cold-adapted yeasts, with special emphasis on more eutrophic habitats, specifically supraglacial and subglacial sediments, and cryoconites (Turchetti et al. 2008; Branda et al. 2010; Buzzini et al. 2012).
Accordingly, the following models of the ecological significance of cold-adapted yeasts in Alpine and Apennine ecosystems could be postulated:
-
1.
A few species (i.e., those occurring at a higher frequency, namely C. gilvescens and C. gastricus) could have probably a superior fitness with regard to the eutrophic conditions present in sediment and cryoconite ecosystems, which allows them not only to survive, but even to grow under such extreme conditions. Accordingly, an in situ selective enrichment of yeast populations could be postulated for these species. This hypothesis could be apparently supported by the results reported by Turchetti et al. (2008), who demonstrated that C. gilvescens is able to exponentially grow under laboratory-simulated in situ conditions.
-
2.
Due to their substantial oligotrophy, ice, snow, and melt water associated with glacier ecosystems could result lesser suitable to support active growth of cold-adapted yeasts. This could suggest that only limited (if any) in situ enrichment could occur in these niches. In particular, ice could represent only a reservoir of surviving cells. Price (2000, 2007) demonstrated that biological activity may occur inside ice, but microorganisms inhabiting this environment are characterized by very low metabolic rates because most of the nutrients and energy that become available could be likely expected to serve just for the repair of cellular and molecular damages. Accordingly, it is possible to deduce that even if this strategy could permit cold-adapted yeasts to remain viable for months and years (thousands or even millions of years, considering Antarctic ice) (Abyzov et al. 1982, 2001), these microorganisms could play only a minor significant role inside the ice mass of Alpine and Apennine glaciers.
-
3.
In this context, melt water could only act by transferring viable yeast cells from supraglacial to subglacial sediments. In fact, temperate glaciers of the Alps and Apennines exhibit crevassing and water channels (Fig. 5.4), which enhance ice melting and water movement from supraglacial to subglacial sides (passing by englacial ecosystems). Accordingly, yeast cells could be supposed to move from superficial snow, supraglacial sediments, water, and englacial ice to reach subglacial sediments.
Considering all above hypotheses, a possible pathway of cold-adapted yeasts inside the glacier ecosystem could be postulated (Fig. 5.4):
-
1.
Yeasts may initially fall on supraglacial debris, cryoconite, snow, and ice surface by wind, rainfall, and snowfall dissemination. The combination of these components could determine the amount and composition of microbial species in the glacier surface (Xiang et al. 2009). After this first step, an in situ post-depositional selection could occur, thus resulting in varying structures of the yeast community. Cryoconite and supraglacial sediments with sufficient concentration of organic compounds may support the selective in situ enrichment of yeasts in favor of the species more adaptable to the cold habitat and able to use complex organic matter (e.g., C. gastricus in supraglacial sediment of the Calderone glacier) (Branda et al. 2010). Other species exhibiting a lower fitness for such habitat could remain in very low cell concentration (or even not quantifiable by using classical, i.e., culture-dependent, methods of analysis).
-
2.
Temperate glaciers are characterized by hydrological transfers from surface to glacier bed due to firn burial and glacier ice flow, but also due to crevassing enhancing flow of melt water, nutrients, and organisms inside the ice body from surface to the glacier bed, where the presence of pressure-melting-point ice at the glacier bed provides not only a basis for nutrient achievement following rock–water contact (Tranter 2005), but also an increased likelihood of crevassing due to enhanced basal sliding and subglacial till deformation (Paterson 1994). All these effects could result in the coupling of supraglacial and subglacial ecosystems allowing yeast cells to percolate from snow and supraglacial sediment coverage through ice, arriving to unfrozen subglacial sediments beneath the glacier bed or remaining within melt water, which could be regarded (together with ice) only a transition step (Fig. 5.4).
-
3.
In subglacial sediments, a second in situ enrichment of yeast populations could be supposed (e.g., C. gilvescens in subglacial sediment in the Ortles-Cevedale complex) (Turchetti et al. 2008). Yeast cells growing in such an ecosystem could also be washed-out by the melt water flow running off from the glacier bed (Fig. 5.4).
6 Conclusions
Investigations on the ecology of glacial habitats have revealed that microbial community is active in biogeochemical transformations such as carbon fixation and respiration, iron cycling and methanogenesis (Anesio and Laybourn-Parry 2012). Cold-adapted yeasts can be considered as an active part of this community, because they have developed a complex suite of physiological and metabolic adaptations, which enhance their possibility to survive and, in some cases, to grow in glacial habitats. Therefore, they play a fundamental role in cold ecosystems, especially in relation to the hydrolysis of complex organic macromolecules connected with the mineralization of organic matter. Regarding their possible role in weathering of surfaces in forefields, their effective in situ quantitative contribution is a still open question, although a recent study hypothesized that cold-adapted yeasts may be the active part of this biogeochemical process (Brunner et al. 2011).
References
Abyzov SS (1993) Microorganisms in the Antarctic ice. In: Friedmann (ed) Antarctic microbiology. Wiley, New York, pp 265–295
Abyzov SS, Bobin NE, Koudryashov BB (1982) Quantitative assessment of microorganisms in microbiological studies of Antarctic glaciers. Biol Bull Acad Sci USSR 9:558–564
Abyzov SS, Mitskevich IN, Poglazova MN, Barkov NI, Lipenkov VY, Bobin NE, Koudryashov BB, Pashkevich VM, Ivanov MV (2001) Microflora of the basal strata at Antarctic ice core above the Vostok Lake. Adv Space Sci 28:701–706
Amato P, Hennebelle R, Magand O, Sancelme M, Delort AM, Barbante C, Boutron C, Ferrari C (2007) Bacterial characterisation of the snow cover at Spitsbergen, Svalbard. FEMS Microbiol Ecol 59:255–264
Anesio AM, Laybourn-Parry J (2012) Glaciers and ice sheets as a biome. Trends Ecol Evol 27:219–225
Anesio AM, Hodson AJ, Fritz A, Psenner R, Sattler B (2009) High microbial activity on glaciers: importance to the global carbon cycle. Global Change Biol 15:955–960
Anesio AM, Sattler B, Foreman C, Telling J, Hodson A, Trantner M, Psenner R (2010) Carbon fluxes through bacterial communities on glacier surface. Ann Glaciol 51:32–40
Arenz BE, Held BW, Jurgens JA, Farrell RL, Blanchette RA (2006) Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol Biochem 38:3057–3064
Benn DI, Evans DJA (1998) Glacier and glaciation. Arnold, London, pp 212–239
Bergauer P, Fonteyne PA, Nolard N, Schinner F, Margesin R (2005) Biodegradation of phenol and phenol-related compounds by psychrophilic and cold-tolerant alpine yeasts. Chemosphere 59:909–918
Branda E, Turchetti B, Diolaiuti G, Pecci M, Smiraglia C, Buzzini P (2010) Yeast and yeast-like diversity in the southernmost glacier of Europe (Calderone glacier, Apennines, Italy). FEMS Microbiol Ecol 72:354–369
Brunner I, Plotze M, Rieder S, Zumsteg A, Furrer G, Frey B (2011) Pioneering fungi from the Damma glacier forefield in the Swiss Alps can promote granite weathering. Geobiology 9:266–279
Butinar L, Spencer-Martins I, Gunde-Cimerman N (2007) Yeasts in high Arctic glaciers: the discovery of a new habitat for eukaryotic microorganisms. A van Leeuw J Microbiol 91:277–289
Buzzini P, Branda E, Goretti M, Turchetti B (2012) Psychrophilic yeasts from worldwide glacial habitats: diversity, daptation strategies and biotechnological potential. FEMS Microbiol Ecol 82:217–241
Cameron KA, Hodson AJ, Osborn AM (2012) Structure and diversity of bacterial, eukaryotic and archaeal communities in glacial cryoconite holes from the Arctic and the Antarctic. FEMS Microbiol Lett 82:254–267
Cannone N, Diolaiuti G, Guglielmin M, Smiraglia C (2008) Accelerating climate change impacts on alpine glacier forefield ecosystems in the European Alps. Ecol Appl 18:637–648
Castello JD, Rogers SO (2005) Life in ancient ice. Princeton University Press, Princeton
Christner BC, Mosley-Thompson E, Thompson LG, Reeve JN (2001) Isolation of bacteria and 16 S rDNAs from Lake Vostok accretion ice. Environ Microbiol 3:570–577
Christner BC, Krivko BH II, Reeve JN (2003a) Molecular identification of bacteria and eukarya inhabiting an Antarctic cryoconite hole. Extremophiles 7:177–183
Christner BC, Mosley-Thompson E, Thompson LG, Reeve JN (2003b) Bacterial recovery from ancient glacial ice. Environ Microbiol 5:433–436
Christner BC, Royston-Bishop G, Foreman CM, Arnold BR, Tranter M, Welch K, Lyons WB, Tsapin AI, Priscu J (2006) Limnological conditions in subglacial Lake Vostok. Limnol Oceanogr 51:2485–2501
Connell LB, Redman R, Craig S, Scorzetti G, Iszard M, Rodriguez R (2008) Diversity of soil yeasts isolated from South Victoria Land, Antarctica. Microb Ecol 56:448–459
De Angelis M, Gaudichet A (1991) Saharan dust deposition over Mont Blanc (French Alps) during the last 30 years. Tellus 43B:61–75
de Garcia V, Brizzio S, Libkind D, Buzzini P, van Broock M (2007) Biodiversity of cold-adapted yeasts from glacial meltwater rivers in Patagonia. Argentina FEMS Microbiol Ecol 59:331–341
Deming WJ (2002) Psychrophiles and Polar regions. Curr Opin Microbiol 5:301–309
Feller G, Narinx E, Arpigny JL, Aittaleb M, Baise E, Genicot S, Gerday C (1996) Enzymes from psychrophilic organisms. FEMS Microbiol Rev 18:189–202
Foght J, Aislabie J, Turner S, Brown CE, Ryburn J, Saul DJ, Lawson W (2004) Culturable bacteria in subglacial sediments and ice from two southern hemisphere glaciers. Microbial Ecol 47:329–340
Fonseca A, Boekhout T, Fell JW (2011) Cryptococcus Vuillemin (1901). In: Kurtzman CP, Fell JW, Boekhout T (eds) The yeasts, a taxonomic study. Elsevier, London, pp 1661–1738
Gildemacher P, Heijne B, Silvestri M, Houbraken J, Hoekstra E, Boekhout T (2006) Interactions between yeasts, fungicides and apple fruit russeting. FEMS Yeast Res 6:1149–1156
Gildemacher PR, Heijne B, Houbraken J, Vromans T, Hoekstra ES, Boekhout T (2004) Can phyllosphere yeasts explain the effect of scab fungicides on russeting of Elstar apples? Eur J Plant Pathol 110:929–937
Herzberg M, Fischer R, Titze A (2002) Conflicting results obtained by RAPD-PCR and large-subunit rDNA sequences in determining and comparing yeast strains isolated from flowers: a comparison of two methods. Int J Syst Evol Microbiol 52:1423–1433
Hodson A (2006) Biogeochemistry of snowmelt in an Antarctic glacial ecosystem. Water Resources Res 42 (11) doi: 10.1029/2005WR004311
Hodson A, Anesio AM, Trantner M, Fountain A, Osborn M, Priscu J, Laybourn-Parry J, Sattler B (2008) Glacial ecosystems. Ecol Monograph 78:41–67
Hodson A, Cameron K, Bøggild C, Irvine-Fynn T, Langford H, Pearce D, Banwart S (2010) The structure, biological activity and biogeochemistry of cryoconite aggregates upon an Arctic valley glacier: Longyearbreen, Svalbard. J Glaciol 56:349–362
Lazzaro A, Brankatschk R, Zeyer J (2012) Seasonal dynamics of nutrients and bacterial communities in unvegetated alpine glacier forefields. Appl Soil Ecol 53:10–22
Lee YM, Kim S-Y, Jung J, Kim EH, Cho KH, Schinner F, Margesin R, Hong SG, Lee HK (2011) Cultured bacterial diversity and human impact on alpine glacier cryoconite. J Microbiol 49:355–362
Libkind D, Moline M, Sampaio JP, van Broock M (2009) Yeasts from high-altitude lakes: infuence of UV radiation. FEMS Microbiol Ecol 69:353–362
MacDonell S, Fitzsimons S (2008) The formation and hydrological significance of cryoconite holes. Prog Phys Geogr 32:595–610
Margesin R (2009) Effect of temperature on growth parameters of psychrophilic bacteria and yeasts. Extremophiles 13:257–262
Margesin R, Miteva V (2011) Diversity and ecology of psychrophilic microorganisms. Res Microbiol 162:346–361
Margesin R, Fauster V, Fonteyne PA (2005a) Characterization of cold-active pectate lyases from psychrophilic Mrakia frigida. Lett Appl Microbiol 40:453–459
Margesin R, Fonteyne PA, Redl B (2005b) Low-temperature biodegradation oh high amounts of phenol by Rhodococcus spp. and basidiomycetous yeasts. Res Microbiol 156:68–75
Margesin R, Fonteyne PA, Schinner F, Sampaio JP (2007) Rhodotorula psychrophila sp. nov., Rhodotorula psychrophenolica sp. nov. and Rhodotorula glacialis sp. nov., novel psychrophilic basidiomycetous yeast species from alpine environments. Int J Syst Evol Microbiol 57:2179–2184
Margesin R, Fell JF (2008) Mrakiella cryoconiti gen. nov., sp. nov., a psychrophilic, anamorphic, basidiomycetous yeast from alpine and arctic habitats. Int J Syst Evol Microbiol 58:2977–2982
Margesin R, Zacke G, Schinner F (2002) Characterization of heterotrophic microorganisms in alpine glacier cryoconite. Arct Antarct Alp Res 34:88–93
Miteva V (2008) Bacteria in snow and glacier ice. In: Margesin R, Schinner F, Marx JC, Gerday C (eds) Psychrophiles: from biodiversity to biotechnology. Springer, Berlin, pp 31–50
Miteva VI, Sheridan PP, Brenchley JE (2004) Phylogenetic and physiological diversity of microorganisms isolated from a deep Greenland glacier ice core. Appl Environ Microbiol 70:202–213
Panikov NS, Sizova MV (2007) Growth kinetics of microorganisms isolated from Alaskan soil and permafrost in solid media frozen down to −35°C. FEMS Microbiol Ecol 59:500–512
Paterson WSB (1994) The physics of glaciers, 3rd edn. Pergamon, Oxford
Paterson WSB (2002) The physics of glaciers. Butterworth-Heinemann, Elsevier Science, Burlington, pp 159–172
Pecci M, D’Agata C, Smiraglia C (2008) Ghiacciaio del Calderone (Apennines, Italy): the mass balance of a shrinking Mediterranean glacier. Geogr Fis Din Quat 31:55–62
Price PB (2000) A habitat for psychrophiles in deep, Antarctic ice. Proc Natl Acad Sci USA 97:1247–1251
Price PB (2007) Microbial life in glacial ice and implications for a cold origin of life. FEMS Microbiol Ecol 59:217–231
Priscu JC, Adams EE, Lyons WB, Voytek MA, Mogk DW, Brown RL, McKay CP, Takacs CD, Welch KA, Wolf CF, Kirshtein CD, Avci R (1999) Geomicrobiology of subglacial ice above Lake Vostok, Antarctica. Science 286:2141–2144
Sampaio JP (2011) Cystofilobasidium Oberwinkler & Bandoni (1983). In: Kurtzman CP, Fell JW, Boekhout T (eds) The yeasts, a taxonomic study. Elsevier, London, pp 1423–1432
Sharp M, Parkes J, Cragg B, Fairchild IJ, Lamb H, Tranter M (1999) Widespread bacterial populations at glacier beds and their relationship to rock weathering and carbon cycling. Geology 27:107–110
Siddiqui KS, Cavicchioli R (2006) Cold-adapted enzymes. Annu Rev Biochem 75:403–433
Simon C, Wiezer A, Strittmatter AW, Daniel R (2009) Phylogenetic diversity and metabolic potential revealed in a glacier ice metagenome. Appl Environ Microb 75:7519–7526
Singh P, Singh SM (2012) Characterization of yeast and filamentous fungi isolated from cryoconite holes of Svalbard, Arctic. Polar Biol 35:575–583
Skidmore M, Foght JM, Sharp MJ (2000) Microbial life beneath a high arctic glacier. Appl Environ Microbiol 66:3214–3220
Skidmore ML, Anderson SP, Sharp M, Foght J, Lanoil BD (2005) Comparison of microbial community compositions of two subglacial environments reveals a possible role for microorganisms in chemical weathering processes. Appl Environ Microbiol 71:6986–6997
Stibal M, Trantner M, Benning LG, Rehak J (2008) Microbial primary production on an Arctic glacier is insignificant in comparison with allochtonous organic carbon input. Environ Microbiol 10:2172–2178
Stibal M, Elster J, Sabacka M, Kastovska K (2007) Seasonal and diel changes in photosynthetic activity of the snow alga Chlamydomonas nivalis (Chlorophyceae) from Svalbard determined by pulse amplitude modulation fluorometry. FEMS Microbiol Ecol 59:265–273
Stibal M, Sabacka M, Zarsky J (2012) Biological processes on glacier and ice sheet surfaces. Nat Geosci 5:771–774
Takeuchi N, Kohshima S, Seko K (2001) Structure, formation and darkening process of albedo-reducing material (cryoconite) on a Himalayan glacier: a granular algal mat growing on the glacier. Arct Antarct Alp Res 33:115–122
Thomas-Hall SR, Turchetti B, Buzzini P, Branda E, Boekhout T, Theelen B, Watson K (2010) Cold adapted yeasts from Antarctica and the Italian Alps. Description of three novel species: Mrakia robertii sp. nov., Mrakia blollopis sp. nov. and Mrakiella niccombsii sp. nov. Extremophiles 14:47–59
Tieber A, Lettner H, Bossew P, Hubmer A, Sattler B, Hofmann W (2009) Accumulation of anthropogenic radionuclides in cryoconite on alpine glaciers. J Environ Radioact 100:590–598
Tranter M (2005) Geochemical weathering in glacial and proglacial environments. In: Holland HD, Turekian KK (eds) Treatise on geochemistry, vol 5. Elsevier, London, pp 189–205
Tranter M, Sharp MJ, Lamb H, Brown GH, Hubbard BP, Willis IC (2002) Geochemical weathering at the bed of Haut Glacier d’Arolla, Switzerland: a new model. Hydrol Process 16:959–993
Turchetti B, Buzzini P, Goretti M, Branda E, Diolaiuti G, D’Agata C, Smiraglia C, Vaughan-Martini A (2008) Psychrophilic yeasts in glacial environments of Alpine glaciers. FEMS Microbiol Ecol 63:73–83
Turchetti B, Thomas-Hall SR, Connell LB, Branda E, Buzzini P, Theelen B, Muller WH, Boekhout T (2011) Psychrophilic yeasts from Antarctica and European glaciers: description of Glaciozyma gen. nov., Glaciozyma martinii sp. nov. and Glaciozyma watsonii sp. nov. Extremophiles 15:573–586
Turchetti B, Goretti M, Branda E, Diolaiuti G, D’Agata C, Smiraglia C, Onofri A, Buzzini P (2013) Influence of abiotic variables on culturable yeast diversity in two distinct Alpine glaciers. FEMS Microbiol Ecol (in press) doi: 10.1111/1574-6941.12164
Uetake J, Yoshimura Y, Nagatsuka N, Kanda H (2012) Isolation of oligotrophic yeasts from supraglacial environments of different altitude on the Gulkana Glacier (Alaska). FEMS Microbiol Ecol 82:1–8
Vishniac HS (1985) Cryptococcus friedmannii, a new species of yeast from the Antarctic. Mycologia 77:149–153
Vishniac HS (2006) A multivariate analysis of soil yeasts isolated from a latitudinal gradient. Microb Ecol 52:90–103
Vishniac HS, Kurtzman CP (1992) Cryptococcus antarcticus sp. nov, and Cryptococcus albidosimilis sp. nov., basidioblastomycetes from antarctic soils. Int J Syst Bacteriol 42:547–553
Vishniac HS, Onofri S (2002) Cryptococcus antarcticus var. circumpolaris var. nov., a basidiomycetous yeast from Antarctica. A van Leeuwenhoek 83:231–233
Wadham JL, Tranter M, Tulaczyk S, Sharp M (2008) Subglacial methanogenesis: a potential climatic amplifier? Global Biogeochem Cycles 22, GB2021
Wharton RA Jr, Vinyard WC, Parker BC, Simmons GM Jr, Seaburg KG (1981) Algae in cryoconit holes on Canada glacier in southern Victoria Land, Antarctica. Phycologia 20:208–211
Wharton JRA, McKay CP, Simmons GM, Parker BC (1985) Cryoconite holes on glaciers. Bioscience 8:499–503
Xiang SR, Shang TC, Chen Y, Yao TD (2009) Deposition and post deposition mechanisms as possible drivers of microbial population variability in glacier ice. FEMS Microbiol Ecol 70:165–176
Zemp M, Haeberli W, Hoelzle M, Paul F (2006) Alpine glaciers to disappear within decades? Geophys Res Lett 33:L13504
Zumsteg A, Luster K, Göransson H, Smittenberg RH, Brunner I, Bernasconi SM, Zeyer J, Frey B (2012) Bacterial, archaeal and fungal succession in the forefield of a receding glacier. Microb Ecol 63:552–564
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Turchetti, B., Goretti, M., Buzzini, P., Margesin, R. (2014). Cold-Adapted Yeasts in Alpine and Apennine Glaciers. In: Buzzini, P., Margesin, R. (eds) Cold-adapted Yeasts. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-39681-6_5
Download citation
DOI: https://doi.org/10.1007/978-3-642-39681-6_5
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-39680-9
Online ISBN: 978-3-642-39681-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)