
Abstract
Research on framing effects has been one of few multidisciplinary endeavors joined by psychologists, economists, political scientists, and management and marketing researchers. Framing effects epitomize the power of linguistic subtlety in regulating decision-making, showing that different ways of framing, phrasing, or presenting virtually identical choice options systematically affect risk preference, evaluation of experience, and persuasiveness of messages. Given its central role in the studies of decision biases, the framing effect has been used as an experimental probe for understanding general mechanisms of human judgment and decision-making. Researchers have proposed various models explaining the framing effect. However, it was not until recently that research of framing effects started to focus more on psychological mechanisms above and beyond phenomenology. We conducted a meta-analysis of neural correlates of framing effects. The topographic convergences from a total of 26 foci found in the fMRI studies of framing effects revealed two key brain areas underlying framing effects: the left anterior cingulate cortex (ACC) and the right inferior frontal gyrus (IFG). Together with behavioral findings, these results suggest that valence framing as a secondary cue becomes most salient and effective when primary contextual or social cues are absent or incongruent. The processing of choice problems under these conditions call for an ambiguity-reducing and conflict-monitoring function, which would result in the ACC activation. Second, the right IFG activation suggests that the nature of valence framing is both semantic and hedonic, involving not only verbatim linguistic analysis, but also interpretation of its affective tones and metaphorical implications.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Behavioral Studies of Framing Effects
1.1 Framing and Framing Effects: Definition and Typology
Since the seminal work by Kahneman and Tversky (1979), Tversky and Kahneman (1981), framing effects have been widely studied by researchers from across the social sciences for over 30 years. Research on framing effects has been one of few multidisciplinary endeavors joined by psychologists, economists, political scientists, and management and marketing researchers. Framing effects epitomize the power of linguistic subtlety in regulating decision-making, oftentimes without awareness of the decision-maker. Different ways of framing, phrasing or presenting virtually identical choice options systematically affect risk preference, evaluation of experience, products or job candidates, and persuasiveness of arguments in negotiation and communication. A meta-analysis has shown that among some primary predictors of risky choice, framing condition was the top predictor (partial r = 0.44) followed by the value of risky payoff (partial r = 0.14), and type of good at stake (partial r = 0.13) while gain or loss condition and probability of payoff were not significant in predicting choice preference (Kühberger et al. 1999, p. 213).
Tversky and Kahneman (1981) used the term ‘decision frame’ broadly to refer to “the decision-maker’s conception of the acts, outcomes, and contingencies associated with a particular choice” (p. 453). Later definitions of framing effects, despite the differences in their connotations and coverage, pinpoint two typical features: equivalency in choice outcomes and opposing valences in presentations of the choice outcomes. That is, framing effects occur when frames that cast “the same critical information in either a positive or a negative light” cause individuals to have different choice preferences (Chong and Druckman 2007; Druckman 2001; Levin et al. 1998, p. 150).
Consider the well-known Asian disease problem demonstrated first by Tversky and Kahneman (1981). In the cover story of the problem, the respondents were asked to imagine that “the US is preparing for the outbreak of an unusual Asian disease, which is expected to kill 600 people. Two alternative programs to combat the disease have been proposed.” The outcomes of the disease-combating plans were then framed (phrased) differently. In the positive frame the respondents were told: “If Plan A is adopted, 200 people will be saved. If Plan B is adopted, there is a one-third probability that all 600 people will be saved, and two-thirds probability that none of them will be saved.” Given a binary choice between the two alternative plans, the majority of the respondents (72 %) were risk averse, preferring the sure option (Plan A) over its gamble equivalent (Plan B). However, when the same outcomes were ‘negatively framed’ in terms of lives lost (“If Plan A is adopted, 400 people will die. If Plan B is adopted, there is a one-third probability that none of them will die, and two-thirds probability that all 600 people will die.”), the majority of the respondents (78 %) were risk-taking, preferring the gamble option (Plan B) over its sure-thing equivalent. This classic framing effect has been reliably replicated with different samples and across cultures and disciplines (e.g., see Kühberger 1998; Kühberger et al. 1999).
Such risk preference reversal due to valence framing of virtually equivalent choice outcomes raises radical doubts about basic assumptions in economic theories of rationality. How can decision agents make consistent and rational choices when empirical evidence of choice preference appears to be so malleable, so vulnerable to framing effects? The classic framing effect is thus viewed as an irrational decision bias and a cognitive illusion because it violates the invariance axiom of expected utility theory, which requires a rational decision-maker to have a consistent preference order among identical choice prospects independent of the way the prospects are presented or framed.
As illustrated in the Asian disease problem, equivalent choice outcomes can be framed with either positive valence or negative valence (e.g., lives saved vs. lives killed, survival rate vs. mortality rate, gain vs. loss, opportunity vs. threat, success vs. failure, benefits vs. costs). A cumulative body of evidence has shown a wide range of behavioral consequences due to framing of decision problems (Chong and Druckman 2007; Kühberger 1998; Levin et al. 1998). Based on their meta-analysis of different kinds of framing effects, Levin et al. (1998) proposed a well-adopted typology. They proposed three types of framing effects: (1) Risky choice framing, as illustrated by the Asian disease problem where framing of choice outcomes affects risk preference (risk averse or risk seeking) of the decision-maker; (2) Attribute framing, where framing of attributes or characteristics of an object or event affects the evaluation of the object (e.g., a sample of ground beef was rated as better tasting when it was labeled as ‘75 % lean’ rather than ‘25 % fat’, see Levin and Gaeth 1988); and (3) Goal framing, where framing of either the positive consequences of performing an act or the negative consequences of not performing the act affects implicit goals an individual adopts and persuasiveness of a message. A well-known example of goal framing effects has been documented by Meyerowitz and Chaiken (1987), showing that women were more apt to engage in breast self-examination when presented with information stressing the negative consequences of not engaging in this action than when presented with information stressing the positive consequences of engaging in this action.
As stated in the title of the paper by Levin et al. (1998), all frames are not created equally. Similarly, not all framing effects are the same. Some involve irrational preference reversals (e.g., the opposite choice preferences under different framings) while others involve a shift in choice preference (e.g., making one option even more attractive than another). A choice shift differs from a choice reversal in that the proportion of risky choices differs across framing conditions but is not significantly greater than 0.5 under one framing condition and significantly less than 0.5 in an alternative framing condition. Wang (1996a) makes the distinction between bidirectional framing effects which involve a reversal in risk preference and unidirectional framing effects which involve a preference shift (e.g., from risk seeking to even more risk seeking under negative framing, or from risk averse to even more risk averse under positive framing).
1.2 Theories of Framing Effects
Given its central role in the studies of decision biases, the framing effect has been used as an experimental probe for understanding general mechanisms of human judgment and decision-making. Researchers have proposed various models explaining framing effects.
-
1.
Prospect Theory
An initial and prevalent explanation of framing effects is based on the prospect theory’s (Kahneman and Tversky 1979) S-shaped value function which is separated by a zero point (status quo) with a gain region above the status quo and a loss region below the status quo. Tversky and Kahneman (1981) theorized that framing manipulation determines whether outcomes are encoded as gains or losses and that this encoding determines which portion of the S-shaped value function would contribute to the risk preference of the decision-maker. For gains implicated in the positive framing condition, the subjective value function is concave and promotes risk aversion, whereas for losses implicated in the negative framing condition, the value function is convex and promotes risk seeking. However, this prospect of theoretical account of framing effects is limited to risky choice framing. In case of attribute framing where the presence of risk is not essential, valence of framing (e.g., success rate vs. failure rate) may either evoke favorable or unfavorable associations in memory (Levin and Gaeth 1988) or evoke different reference points (goal or minimum requirement rather than status quo, Wang and Johnson 2012).
-
2.
Fuzzy-Trace Theory
Another famous account of framing effects is derived from fuzzy-trace theory proposed by Reyna and Brainerd (1991, 1995, 2011). The theory posits that people form two types of mental representations about a past event, called verbatim and gist traces. Gist traces are fuzzy representations of a past event (e.g., its bottom-line meaning), whereas verbatim traces are detailed representations of a past event. Although people (adults) are capable of processing both verbatim and gist information, they prefer to reason with gist traces rather than verbatim traces. In the case of encoding choice outcomes of a sure thing and a gamble of equal expected value in the Asian disease problem, the gist translations boil down to choosing between (a) saving some people, (b) saving some people or saving no one under the positive (gain) frame, and choosing between (c) some people die, and (d) some people die or none die under the negative (loss) framing. Reyna and Brainerd (1991, 1995) showed that relational gist of quantities (e.g., some, all, none) was sufficient to replicate the classic framing effect. Thus, framing effects can be seen as a result of translating expected choice outcomes to gist representations of quantities with positive or negative connotations. These nonnumerical framing effects demonstrated that numbers, which are essential to predictions of prospect theory and all other utility theories, were not necessary to observe the framing effect. Moreover, according to prospect theory or other expected utility theories, the outcome of zero in the gamble option literally contributes nothing to predictions of choice behavior because the value of zero, based on the S-shaped value function, is also set at zero. However, when the information about the zero outcomes was removed from the classic Asian disease problem, framing effects disappeared (see Reyna and Brainerd 2011).
-
3.
Asymmetry in Dual Processing
Another approach to theorizing framing effects emphasizes the interplay between two distinct processing systems or asymmetry in affective or cognitive processing. Dunegan (1993), for example, proposed that framing may act as a catalyst for different modes of cognitive processing. Characteristics of controlled cognitive modes were found when information was negatively framed; characteristics of more automatic processing were found when information was positively framed, suggesting that positive framing may be perceived as a default with a lower cognitive processing load. Kuo et al. (2009) employed eye tracking to measure cognitive effort in a framing study. Their results suggest an asymmetry in cognitive effort, as indicated by eye fixation and eye-movement time per word, due to positive and negative frames. More effort was observed in the negative framing condition than in the positive framing condition. However, a study by Whitney et al. (2008) found that working memory load slightly reduced risk-seeking tendency but not framing effects, suggesting that valence framings did not have differential effects on cognitive processing effort.
Another account of framing effects proposed by McElroy and Seta (2003, 2004) focuses on the asymmetry between analytic processing and holistic processing. This model assumes that analytic processing is insensitive to the influence of framing, whereas holistic processing is more susceptible to framing. By behaviorally inducing selective hemispheric activation, framing effects were found when the right hemisphere was selectively activated, whereas they were not observed when the left hemisphere was selectively activated.
-
4.
Ambiguity-Ambivalence Hypothesis
There are other limits and constraints to framing effects in addition to hemispheric dominance. Meta-analyses (Levin et al. 1998; Kühberger 1998) show that the effect of framing is overall significant but not as ‘pervasive’ or ‘robust’ as previously believed. Many researchers have explored the premise and moderators of framing effects. Some examples of the moderators are sex of the decision-maker (Fagley and Miller 1990, 1997), cognitive ability (Stanovich and West 1998), personal involvement (Levin et al. 1998, p. 160), reflection on and rationale for the decision (Takemura 1994; Fagley and Miller 1987; Miller and Fagley 1991; Sieck and Yates 1997), personal knowledge about risks involved (e.g., Bohm and Lind 1992; Levin and Chapman 1990), perceived ambiguity of the values presented in the problem descriptions (Kühberger 1995), task context (e.g., medical vs. statistical, Bless et al. 1998), need for cognition (LeBoeuf and Shafir 2003), perceived interdependence between individuals at risk (Bloomfield et al. 2006; Wang et al. 2001), and social group size and composition (Bloomfield 2006; Shimizu and Udagawa 2011a; Wang 1996a, b; Zhang and Miao 2008).
The aforementioned studies focusing on the variables that can enhance, limit, or even eliminate a framing effect have taken into consideration individual as well as situational factors that determine the magnitude as well as process of framing effects. This line of research calls for an overarching theory of framing effects that addresses these ecological, social, and dispositional constraints on and premises of framing effects. This led to the development of an ambiguity-ambivalence (AA) hypothesis of framing effects and decision biases (Wang 2008). The AA hypothesis assumes that (1) decision cues are selected and used in accordance to their priorities. (2) Cue priority reflects ecological and social validity of a cue in predicting specific risks, beyond the stated values and probabilities of outcomes in a choice task. Primary cues are valid ecological, social, and life-history variables (such as kith-and-kin relations, social group size, sex, age, health, socioeconomic status, and mating/reproductive cues, etc.). Secondary cues in decision-making are mainly communicational, such as verbal framing, facial expression, tone of voice, etc. (3) Primary cues determine the settings of decision reference points (e.g., goals and bottom lines) and anchor decision preference while secondary cues (and individual dispositional factors) fine tune the settings of reference points and choice preference. (4) Decision biases, such as framing effects, tend to occur as a result of secondary cue use when primary cues are either absent in risk communication (i.e., an ambiguity condition) or when primary cues elicit conflicting preferences (i.e., an ambivalence condition).
From this perspective, framing effects would occur as a result of ambiguity and ambivalence in decision cues and preferences. Similar ideas can be seen in fuzzy-trace theory (Reyna and Brainerd 1991) and the discussion on the use of probabilistic mental models (Gigerenzer et al. 1991) to deal with informational ambiguity as a premise of framing effects (Chang et al. 2002; Kühberger 1995).
As predicted from the AA hypothesis, framing effects occurred when making life–death decisions in evolutionarily novel and socially unfamiliar large group contexts but disappeared when the same problems were scaled down to a small group context with a handful of people (Bloomfield 2006; Shimizu and Udagawa 2011a; Zhang and Miao 2008; Wang and Johnston 1995; Wang 1996a, b). Moreover, work experience in large organizations reduced sensitivity to framing manipulation in large group contexts (Shimizu and Udagawa 2011b). Interestingly, framing effects in a small group reappeared when the small group included both strangers and kin relatives, thus creating ambivalence in risk preference between a “we all live or die together” risk-seeking preference for kith-and-kin and a more risk-averse preference for strangers (Wang et al. 2001).
2 A Neuroscience Approach to Understanding Framing Effects
The discussion so far summarizes behavioral studies of framing effects. These studies leave open the possibility for more theoretical development and better understanding of underlying processes that produce or inhibit framing effects. It was not until recently that research on framing effects started to focus more on the underlying psychological mechanisms above and beyond phenomenology, owing to rapid development in neuroimaging technology. Neuroscience and neuroimaging can help forward our understanding of framing effects in at least four different yet coherent ways.
First, the neuroscience approach would allow us to evaluate framing theories by mapping the framing-related brain activations against the key brain areas implicated by different theories of framing effects. Second, many hypothetical mechanisms and post-hoc explanations based on behavioral effects can be better understood, verified, or disproved by using the brain imaging technique. Third, neuroimaging studies often shed light into puzzling or conflicting findings from behavioral studies of framing effects. Fourth, brain imaging studies capture online neural activities of the entire brain during the decision-making. This brain map of activation or deactivation may highlight uncharted brain areas and provide insights and new leads for future investigations.
In the following sections of the chapter, we illustrate the use and usefulness of the neuroscience approach to framing effects by generating predictions about neural correlates of framing effects based on different theoretical viewpoints, and by conducting a meta-analysis of the fMRI studies of framing effects. We end the chapter with a summary and conclusions regarding the status quo of behavioral and neuroimaging studies of framing effects.
2.1 Contrasting and Evaluating Theories of Framing Effects
As illustrated in Table 9.1, theories of framing effects make overlapping but distinctive predictions about brain regions underpinning the proposed functions. The predictions presented in Table 9.1 are by no means meant to be thorough or systematic.
Table 9.1 is only a sketchy outline to exemplify how theories of framing effects can be tested with neuroimaging studies. For a more specific example, consider an fMRI study of framing effects by Gonzalez et al. (2005). The authors proposed a cognitive–affective tradeoff model which can be classified as one of the ‘asymmetry in dual processing models’ as listed in Table 9.1.
The cognitive–affective tradeoff model assumes that the framing effect occurs due to a tradeoff between the cognitive effort required to calculate expected values of an alternative and the affective value of the alternative. The model predicts that decision-makers prefer choice options that are cognitively less effortful to process or/and affectively more pleasant. In a positive frame, the compromise between maximizing hedonic feeling and minimizing cognitive effort is easier to achieve. For instance, selecting the option in which “200 people will be saved” feels more positive in an emotional sense and is less effortful (i.e., no calculations are necessary). In contrast, such a compromise is more difficult to attain in the negative frame. Although the option in which “400 people will die” requires little calculation, the relatively bad outcome makes it less attractive due to a stronger feeling of displeasure. Thus when selecting among options presented in a negative frame, individuals are more willing to undertake the cognitive effort demanded to assess the more risky option in order to get the hedonically less unpleasant outcome.
Previous studies have shown that individuals take longer to make decisions when the options are framed as losses rather than gains (Payne et al. 1993). However, it is still unknown whether it is a result of a greater cognitive effort in the negative than in the positive frame or a result of a larger affective cost. To answer these questions derived from the cognitive-affective tradeoff model, the authors conducted an fMRI study which revealed that the cognitive effort required to select a sure gain was considerably lower than the cognitive effort required to choose a risky gain. The fMRI results, although not directly correlated with the behavioral framing effects, showed significantly higher activation levels in the frontal and parietal lobes when making risky rather than certain choices under positive frames. This finding is consistent with their theoretical view that the sure gain is a default choice over its gamble equivalent when considering cognitive processing effort.
2.2 Neuroimaging Tests of Alternative Behavioral Accounts of Framing Effects
Behavioral studies of framing effects have yielded interesting results that are often open to alternative explanations. These alternative accounts of behavioral findings can be further evaluated in neuroimaging studies. A recent fMRI study by Zheng et al. (2010) on group- size-dependent framing effects illustrates such a value of the neuroimaging approach.
In a series of studies (Wang 1996a, b; Wang et al. 2001), we examined the appearance and disappearance of framing effects when the size of the group (the total number of lives at stake) was systematically manipulated. The same life–death problem was framed either in terms of lives saved or in terms of lives lost. The framing effect was evident, but it occurred only when the problem was presented in a large, anonymous, and thus ambiguous group context involving 600 lives or more. The framing effect disappeared when the size of the endangered group was within a two-digit number (<100), and the majority of the participants unambiguously preferred the gamble option under both the saving lives and losing lives framing conditions.
These findings suggest that the small size of a social group signals a higher interdependence between group members and evokes a kith-and-kin rationality that guides a live-or-die together risk preference. In contrast, risk preference of a decision-maker becomes erratic when prioritized group cues are absent in a large anonymous group context. When risk preference is ambiguous, secondary cues such as verbal framing are attended and used to direct choices. However, an alternative hypothesis of group size-dependent framing effects based on a utility theory would posit that people are more competent in calculating small numbers than large numbers when evaluating risky outcomes, and thus are less ambiguous in their choice preference and less susceptible to framing manipulation.
To evaluate these rival accounts of group size-dependent framing effects, Zheng et al. (2010) conducted an fMRI study. The results of this study, as shown in Table 9.2, help to evaluate the two alternative hypotheses.
Group size difference was captured by activation in the middle frontal gyrus. Verbal framing in the large group context was associated with activation of the right inferior frontal gyrus while the same valence framing in the small group context was associated with activation of different brain structures, including the insula and an area in the parietal lobe. These differential activations were all located in the right hemisphere. Note that the right inferior frontal gyrus includes the homologue of the Broca’s area.
These results support the group size account rather than the numerical size account of framing effects. First, behavioral framing effects only occurred in the large group context and were associated with the right IFG activation. In contrast, the disappearance of the framing effect in the small group context was associated with activation of different brain structures (e.g., the right insular). Second, the group size-dependent framing effect was restricted in the right hemisphere, thus was unlikely an explicit numerical processing effect. If numerical processing played a major role in determining appearance or disappearance of framing effects, the left hemisphere and the brain structures related to numerical processing should be differentially activated (e.g., Piazza et al. 2004; Pinel et al. 2004). Additional behavioral results further exclude the ‘large-number’ account of framing effects. Wang (1996a) demonstrated that framing effects occurred in large groups of 6000 as well as 600 people, and disappeared in small groups of 60 as well as 6 people. The tenfold difference between the two large groups and between the two small groups did not make a difference in choice preference. Further evidence comes from the Wang, et al. (2001) study, where the classic framing effect occurred in the context of 6 billion human lives but disappeared in the context of 6 billion ET (extraterrestrial) lives. Thus, the framing effect is not likely a large number effect, but is human group size sensitive.
Our findings overall are consistent with the predictions of the AA hypothesis: distinct brain areas are recruited for solving ambiguity or ambivalence caused by the lack of primary social or relational cues in a large anonymous group context. In contrast, framing effects diminished in a small group context while the insula and parietal lobe in the right hemisphere were distinctively activated, suggesting an important role of emotion in switching choice preference from an indecisive mode to a more consistent risk-taking inclination.
The brain imaging findings are interesting in that they suggest that the framing effect is both linguistic and implicit. The affective component of valence framing may direct a holistic, right hemispheric process while the cognitive connotation of framing activates implicit linguistic processing in the right hemisphere in addition to activation of the common linguistic processing regions in the left hemisphere. Previous studies show that the right IFG is involved in response inhibition and impulse control (Aron et al. 2003, 2004; Asahi et al. 2004). Thus, the higher activation found in the right IFG under positive framing suggests a greater control effort induced by using positive framing than negative framing in making hypothetical life–death choices.
2.3 Gaining New Insights from Brain Imaging Studies of Framing Effects
The findings from neuroimaging studies of framing effects often provide new insights and new research directions. In this section, we report several new findings derived from a current analysis of an fMRI study of framing effects, in which we examined brain activations when making hypothetical risky choices.
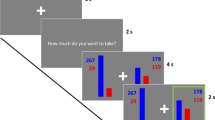
The risky choice problems, adopted from those used in the study by Wang (1996a), involved monetary investment (either 600 or 60,000 Chinese Yuan) and property (either 6 or 600 precious oil paintings) at stake. The structure of the choice problems used in this study was identical to that of the Asian disease problem. The participants were asked to make a binary choice between a sure outcome (framed either as saving one-third of a total amount or losing two-thirds of the total amount) and a gamble of equal expected value. Twenty-one participants (8 males) took part in this study. The procedure was the same as that used in Zheng et al. (2010).
The behavioral effects of valence framing were significant in the painting scenario (risk-seeking choices increased from 47.8 % under positive framing to 57.6 % under negative framing), χ2(1) = 7.617, p = 0.0058. The framing effect was not significant in the money investment scenario (the risk-seeking choice = 55.7 % under positive framing and 62.2 % under negative framing; χ2(1) = 0.246, p = 0.620).
The results of the fMRI data analysis are shown in Table 9.3 and Fig. 9.1. The left inferior frontal gyrus and the left superior frontal gyrus were identified to have a higher activation level under negative framing than positive framing while the left medial temporal cortex showed a higher activation under the positive framing than negative framing. These results are consistent with some previous findings. The inferior frontal gyrus is also found to be associated with framing effects in our previous study (Zheng et al. 2010). In addition, the left medial temporal cortex and the left superior frontal gyrus identified in the present analysis are geographically close to two of the activation regions reported by De Martino et al.’ (2006): the left medial temporal gyrus and the left superior frontal sulcus, respectively.

Brain regions showing the effect of frame. Abbreviation IFG Inferior frontal gyrus; MTC Medial temporal cortex; SFG Superior frontal gyrus. Error bars denote standard errors. The statistical significance was determined by Monte Carlo simulations to obtain a P-corrected value <0.05, after correcting for whole brain comparisons. The corrected threshold corresponds to a P-uncorrected value <0.005 with a minimum cluster size of 1080 mm3 (a gray mask with 55,342 voxels was used)
Human neuropsychological studies have highlighted the importance of both frontal cortex and the medial temporal cortex, including the hippocampus in memory encoding and retrieval (e.g., Cohen and Eichenbaum 1993; Tulving et al. 1994). Previous studies also suggest that effortful and strategic memory processes are mainly mediated by the frontal lobes, whereas automatic associative memory processes are mainly mediated by the medial temporal lobes and hippocampus (MTL/H) (e.g., Moscovitch 1994). Thus, our results suggest a differential activation pattern by framing where the negative framing involves more elaborative encoding in the frontal lobe and the positive framing elicits more holistic encoding in the medial temporal cortex.
2.4 A Meta-Analysis of Neuroimaging Studies of Framing Effects—Toward a Better Understanding of Framing Effects
In the field of neuroscience, there have not been many neuroimaging studies on framing effect so far. We carried out a systematic search using the keywords ‘framing effect’ and ‘MRI’ to identify relevant studies included in the PubMed and PsycINFO databases; the search was conducted in December 2012, and no time span was specified for date of publication. Our inclusion criteria were that: (1) the studies presented coordinate-based analyses of the data; (2) all or most of the brain was imaged; and (3) participants were asked to choose in different frames. The fMRI studies obtained from this search are summarized below in Table 9.4.
As shown in Table 9.4, the choice tasks involved both life–death and monetary and other types of problems. The behavioral framing effects were all significant, either bidirectional or unidirectional. Non-verbal valence framing had similar behavioral effects. Guitart-Masip et al. (2010) adopted a unique valence framing by associating visual stimuli (different texture patterns printed on a card) with gains or losses before using them as valence frames for framing choice outcomes. When the sure-thing outcome of a choice is either presented on a ‘gain’ card (positive frame) or on a ‘loss’ card (negative frame), classic monetary framing effects occurred.
Brain regions which were reported to contribute to the appearance of framing effects include the amygdala (De Martino et al. 2006; Guitart-Masip et al. 2010), the insula, and the right IFG (Zheng et al. 2010).
Some brain regions inhibit or reduce framing effects. De Martino et al. (2006) found that greater activity in the orbital and medial prefrontal cortex (OMPFC) predicted a reduced susceptibility to the framing effect across participants. The anterior cingulate cortex (ACC) was also implicated in reduced framing effects (De Martino et al. 2006; Roiser et al. 2009).
The ACC is thought to reduce framing effects by acting as a teaching signal (Botvinick 2007). Because ACC activation was greater when participants’ choices were incongruent with frame effects (De Martino et al. 2006), it may modulate the motivational influence of the amygdala on choice. This possibility is supported by the greater coupling between the ACC and the amygdala in participants who were less susceptible to the frame (Roiser et al. 2009). A similar role might be attributed to the orbital frontal cortex (OFC), a region which correlates with resistance to monetary framing effects (De Martino et al. 2006), and with which the ACC has strong reciprocal connectivity (Kringelbach and Rolls 2004).
Interestingly, researchers discovered that susceptibility to framing is related to genetic variation in the serotonin transporter gene (the 5-HTTLPR). Individuals homozygous for the short allele at the 5-HTTLPR rather than individuals homozygous for the long allele were found to be more susceptible to valence framing when making hypothetical monetary risky choices. This susceptibility to framing effects is associated with altered amygdala activity and lack of prefrontal regulatory control (Roiser et al. 2009). However, this finding has been challenged by a follow-up study. Talmi, Hurlemann, Patin, and Dolan (2010) reported that two patients with Urbach-Wiethe disease, a rare condition associated with congenital, complete bilateral amygdala degeneration, exhibited an intact framing effect. However, choice preference in these patients did show a qualitatively distinct pattern compared to controls, as evidenced by a significantly increased risk-seeking preference. These findings suggest that the amygdala does exert an overall influence on risk-taking but may not play a causal role in framing effects.
We then conducted our meta-analysis by entering the coordinates of all the foci that were reported in the above fMRI studies of framing effects to get their topographic convergences. The goal of coordinate-based meta-analysis of neuroimaging data is to identify brain areas where the reported foci of activation converge across published experiments. In this meta-analysis of neural correlates of framing effects, we adopted a widely used technique for coordinate-based meta-analyses of neuroimaging data, called activation likelihood estimation (ALE). ALE assesses the overlap between foci based on modeling them as probability distributions centered at their respective coordinates. ALE maps are then obtained by computing the union of activation probabilities for each voxel. To differentiate true convergence of foci from random clustering (i.e., noise), a permutation test is applied: to obtain an ALE null distribution, the same number of foci as in the real analysis are randomly redistributed throughout the brain (see Eickhoff et al. 2009; Turkeltaub et al. 2002). In other words, the meta-analysis determines if the clustering is significantly higher than expected under the null distribution of a random spatial association of results from the considered experiments.
Our analysis was implemented using GingerALE Version 2.1.1 (available at http://brainmap.org/ale). The meta-analysis was performed using the Montreal Neurological Institute (MNI) stereotactic coordinates derived from the studies listed in Table 9.4. Coordinates published in Talairach space were transformed to the Montreal Neurological Institute (MNI) template according to the Lancaster transform (icbm2tal) in GingerALE. Statistical significance was determined using a permutation test of randomly generated foci. No assumptions were made concerning the distribution or spatial separation of these random foci; however, clusters of activity were required to exceed 200 mm3 in volume. The test was corrected for multiple comparisons using the false discovery rate (FDR) method (Genovese et al. 2002). Anatomical labels of final cluster locations are provided by the Talairach Daemon.
Initially, a total of 23 foci were analyzed. The ALE meta-analysis showed that high ALE values were observed in the left ACC and the right IFG (see Table 9.5; Fig. 9.2), indicating these two brain regions are the key neural correlates of framing effects.

We then added into the ALE meta-analysis the coordinates of the three foci associated with framing effects as reported in the previous section of this chapter. The brain localization results for framing effects based on the 26 foci remained the same.
Neurobiological studies of the right IFG have shown some converging evidence for its unique role in ‘semantic selection’ by which competing activated concepts are sorted out, inhibiting competing concepts while selecting one concept for action, particularly when the concept to be selected involves atypical usage. For instance, when people are given a common noun (e.g., cake) and asked to produce a typical use (‘bake’ or ‘eat’), the left IFG is strongly active; but when asked to produce an unusual use (e.g. ‘sell’) of such nouns, the right IFG is more strongly active (see Jung-Beeman 2005 for a review). Both the left and the right IFG play a unique role in understanding the figurative meaning of a metaphor beyond semantic analysis (Rapp et al. 2004). In addition, the right IFG is active in response inhibition and selection. The go/no-go task, which taps the ability to inhibit prepotent response tendency (for instance, stop pressing a button when a red signal appears), has consistently activated the lateral prefrontal cortex, particularly the right IFG (Aron et al. 2003; Asahi et al. 2004; Chikazoe et al. 2007). Some researchers consider the right IFG the most important prefrontal structure that exerts inhibition and cognitive control over subcortical structures of the brain (Aron et al. 2004). The same area is also implicated in risk aversion: higher risk aversion is correlated with higher activity at the right IFG (Christopoulos et al. 2009).
Coupled with the results of our meta-analysis, these findings suggest that the right IFG is a unique neural correlate of framing effects. It plays an integrative role in evaluating risk-related cues and in regulating risk preference and choice selection based on both semantic and affective meanings imbedded in choice problems.
Researchers have suggested a variety of ways to interpret the neuroimaging findings of the framing effect. De Martino et al. (2006) suggested that the framing effect is driven by an affect heuristic underwritten by an emotional system. From the perspective of a dual-system framework, Kahneman and Frederick (2006) interpreted these findings as evidence that different frames evoke distinct emotional responses that different individuals can suppress to various degrees. Similarly, Gonzalez et al. (2005) proposed a model which tries to incorporate a tradeoff between the cognitive effort required to calculate expected values of an alternative and the affective value of the alternative to explain the choice process underlying the framing effect.
Based on our meta-analysis and the AA hypothesis of framing effects, we propose a new view on neural mechanisms of framing effects. First, valence framing as a secondary choice cue becomes most salient and effective when primary contextual or social cues are absent or incongruent. The processing of choice problems under these conditions calls for an ambiguity-reducing and conflict-monitoring function, which would result in the ACC recruitment and activation. Second, the right IFG activation suggests that the nature of valence framing is both semantic and hedonic, involving not only verbatim linguistic analysis but also interpretation of its affective tones and metaphorical implications.
References
Aron AR, Fletcher PC, Bullmore ET, Sahakian BJ, Robbins TW (2003) Stop-signal inhibition disrupted by damage to right inferior frontal gyrus in humans. Nat Neurosci 6:115–116
Aron AR, Robbins T, Poldrack R (2004) Inhibition and the right inferior frontal cortex. Trends Cogn Sci 8:170–177
Asahi S, Okamoto Y, Okada G, Yamawaki S, Yokota N (2004) Negative correlation between right prefrontal activity during response inhibition and impulsiveness: an fMRI study. Eur Arch Psychiatry Clin Neurosci 254:245–251
Bless H, Betsch T, Franzen A (1998) Framing the framing effect: the impact of context cues on solutions to the “Asian disease” problem. Euro J Soc Psychol 28:287–291
Bloomfield AN (2006) Group size and the framing effect: treats to human beings and animals. Mem Cogn 34:929–937
Bloomfield AN, Sager JA, Bartels DM, Medin DL (2006) Caring about framing effects. Mind Soc 5:123–138
Bohm P, Lind H (1992) A note on the robustness of a classical framing result. J Econ Psychol 13:355–361
Botvinick MM (2007) Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cogn Affect Behav Neurosci 7:356–366
Botvinick MM, Cohen JD, Carter CS (2004) Conflict monitoring and anterior cingulate cortex: an update. Trends in Cognitive Sciences 8:539–546
Breiter HC, Aharon H, Kahneman D, Dale A, Shizgal P (2001) Functional imaging of neural responses to expectancy and experience of monetary gains and losses. Neuron 30:619–639
Cendri A, Hutcherson CA, Plassmann H, Gross JJ, Rangel A (2012) Cognitive regulation during decision making shifts behavioral control between ventromedial and dorsolateral prefrontal value systems. J Neurosci 32:13543–13554
Chang CJ, Yen SH, Duh RR (2002) An empirical investigation of competing theories to explain the framing effect in accounting-related decision. Behav Res Acc 14:35–64
Chikazoe J, Konishi S, Asari T, Jimura K, Miyashita Y (2007) Activation of right inferior frontal gyrus during response inhibition across response modalities. J Cogn Neurosci Arch 19:69–80
Chong D, Druckman J (2007) Framing theory. Annu Rev Polit Sci 10:103–126
Christopoulos GI, Tobler PN, Bossaerts P, Dolan RJ, Schultz W (2009) Neural correlates of value, risk, and risk aversion contributing to decision making under risk. J Neurosci 29:12574–12583
Cohen NJ, Eichenbaum HE (1993) Memory, amnesia, and the hippocampal system. MIT Press, Cambridge, MA
De Martino B, Kumaran D, Seymour B, Dolan RJ (2006) Frames, biases, and rational decision-making in the human brain. Science 313:684–687
Druckman JN (2001) On the limits of framing effects: Who can frame? J Polit 63:1041–1066
Dunegan KJ (1993) Framing, cognitive modes, and image theory: toward an understanding of a glass half full. J Appl Psychol 78:491–503
Eickhoff SB, Laird AR, Grefkes C, Wang LE, Zilles K, Fox PT (2009) Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp 30:2907–2926
Fagley NS, Miller PM (1987) The effects of decision framing on choice of risky vs. certain options. Organ Behav Hum Decis Process 39:264–277
Fagley NS, Miller PM (1990) The effect of framing on choice: interactions with risk-taking propensity, cognitive style, and sex. Pers Soc Psychol Bull 16:496–510
Fagley NS, Miller PM (1997) Framing effects and arenas of choice: your money or your life? Organ Behav Hum Decis Process 71:355–373
Genovese CR, Lazar NA, Nichols TE (2002) Thresholding of statistical maps in functional neuroimaging using the false discovery rate. NeuroImage 15:870–878
Gigerenzer G, Hoffrage U, Kleinbölting H (1991) Probabilistic mental models: a Brunswikian theory of confidence. Psychol Rev 98:506–528
Gonzalez C, Dana J, Koshino H, Just M (2005) The framing effect and risky decisions: examining cognitive functions with fMRI. J Econ Psychol 26:1–20
Guitart-Masip M, Talmi D, Dolan R (2010) Conditioned associations and economic decision biases. NeuroImage 53:206–214
Holroyd CB, Nieuwenhuis S, Mars RB, Coles MGH (2004) Anterior cingulate cortex, selection for action, and error processing. In Posner MI (ed) Cognitive neuroscience of attention. Guilford Press, New York, pp 219–231
Honey GD, Fu CH, Kim J et al (2002) Effects of verbal working memory load on corticocortical connectivity modeled by path analysis of functional magnetic resonance imaging data. NeuroImage 17:573–582
Jung-Beeman M (2005) Bilateral brain processes for comprehending natural language. Trends Cogn Sci 9:512–518
Kahneman D, Frederick S (2006) Frames and brains: elicitation and control of response tendencies. Trends Cogn Sci 11:45–46
Kahneman D, Tversky A (1979) Prospect theory. Econometrica 47:263–292
Kringelbach ML, Rolls ET (2004) The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog Neurobiol 72:341–372
Kühberger A (1995) The framing of decisions: a new look at old problems. Organ Behav Hum Decis Process 62:230–240
Kühberger A (1998) The influence of framing on risky decisions: a meta-analysis. Organ Behav Hum Decis Process 75:23–55
Kühberger A, Schulte-Mecklenbeck M, Perner J (1999) The effects of framing, reflection, probability, and payoff on risk preference in choice tasks. Organ Behav Hum Decis Process 78:204–231
Kuo FY, Hsu CW, Day RF (2009) An exploratory study of cognitive effort involved in decision under Framing-an application of the eye-tracking technology. Decis Support Syst 48:81–91
LeBoeuf RA, Shafir E (2003) Deep thoughts and shallow frames: on the susceptibility to framing effects. J Behav Decis Mak 16:77–92
Levin IP, Chapman DP (1990) Risk taking, frame of reference, and characterization of victim groups in AIDS treatment decisions. J Exp Soc Psychol 26:421–434
Levin IP, Gaeth GJ (1988) Framing of attribute information before and after consuming the product. J Consum Res 15:374–378
Levin IP, Schneider S, Gaeth GJ (1998) All frames are not created equal: a typology and critical analysis of framing effects. Organ Behav Hum Decis Process 76:149–188
McElroy T, Seta J (2003) Framing effects: an analytic-holistic perspective. J Exp Soc Psychol 39:610–617
McElroy T, Seta JJ (2004) On the other hand Am I rational? Hemispheric activation and the farming effect. Brain Cogn 55:572–580
Meyerowitz BE, Chaiken S (1987) The effect of message framing on breast self-examination attitudes, intentions, and behavior. J Pers Soc Psychol 52:500–510
Miller PM, Fagley NS (1991) The effects of framing, problem variations, and providing rationale on choice. Pers Soc Psychol Bull 17:517–522
Montague PR, Berns GS (2002) Neural economics and the biological substrates of valuation. Neuron 36:265–284
Moscovitch M (1994) Cognitive resources and dual-task interference effects at retrieval in normal people: the role of the frontal lobes and medial temporal cortex. Neuropsychology 8:524
O’Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C (2001) Abstract reward and punishment representations in the human orbitofrontal cortex. Nat Neurosci 4:95–102
Owen AM (1997) The functional organization of working memory processes within human lateral frontal cortex: the contribution of functional neuroimaging. Euro J Neurosci 9:1329–1339
Payne JW, Bettman JR, Johnson EJ (1993) The adaptive decision maker. Cambridge University Press, New York
Piazza M, Izard V, Pinel P, Le Bihan D, Dehaene S (2004) Tuning curves for approximate numerosity in the human intraparietal sulcus. Neuron 44:547–555
Pinel P, Piazza M, Le Bihan D, Dehaene S (2004) Distributed and overlapping cerebral representations of number, size, and luminance during comparative judgments. Neuron 41:983–993
Rapp AM, Leube DT, Erb M, Grodd W, Kircher TTJ (2004) Neural correlates of metaphor processing. Cogn Brain Res 20:395–402
Reyna VF, Brainerd CJ (1991) Fuzzy-trace theory and framing effects in choice: Gist extraction, truncation, and conversion. J Behav Decis Mak 4:249–262
Reyna VF, Brainerd CJ (1995) Fuzzy-trace theory: an interim synthesis. Learn Individ Differ 7:1–75
Reyna VF, Brainerd CJ (2011) Dual processes in decision making and developmental neuroscience: a fuzzy-trace model. Dev Rev 31:180–206
Roiser JP, De Martino B, Tan GCY, Kumaran D, Seymour B, Wood NW, Dolan R (2009) A genetically mediated bias in decision making driven by failure of amygdala control. J Neurosci 29:5985–5991
Shimizu K, Udagawa D (2011a) A re-examination of the effect of contextual group size on people’s attitude to risk. Judgment and Decision Making 6:156–162
Shimizu K, Udagawa D (2011b) How can group experience influence the cue priority? A re-examination of the ambiguity-ambivalence hypothesis. Front Evol Psychol 2:1–9
Sieck W, Yates JF (1997) Exposition effects on decision making: choice and confidence in choice. Organ Behav Hum Decis Process 70:207–219
Sokol-Hessner P, Hutcherson C, Hare T, Rangel A (2012) Decision value computations adjust to the available decision time. Eur J Neurosci 35:1065–1074
Stanovich KE, West RF (1998) Individual differences in framing and conjunction effects. Think Reason 4:289–317
Takemura K (1994) Influence of elaboration on the framing of decision. J Psychol 128:33–39
Talmi D, Hurlemann R, Patin A, Dolan R (2010) Framing effect following bilateral amygdala lesion. Neuropsychologia 48:1823–1827
Tom SM, Fox CR, Trepel C, Poldrack RA (2007) The neural basis of loss aversion in decision making under risk. Science 315:515–518
Tulving E, Kapur S, Craik FIM, Markowitsch HJ, Houle S (1994) Hemispheric encoding/retrieval asymmetry in episodic memory: positron emission tomography findings. Proc Natl Acad Sci USA 91:2016–2020
Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA (2002) Meta-analysis of the functional neuroanatomy of single-word reading: method and validation. Neuroimage 16:765–780
Tversky A, Kahneman D (1981) The framing of decisions and the psychology of choice. Science 211:453–458
Wang XT (1996a) Framing effects: Dynamics and task domains. Organ Behav Hum Decis Process 68:145–157
Wang XT (1996b) Domain-specific rationality in human choices: violations of utility axioms and social contexts. Cognition 60:31–63
Wang XT (2008) Risk communication and risky choice in context: ambiguity and ambivalence hypothesis. Ann N Y Acad Sci 1128:78–89
Wang XT, Johnson GJ (2012) A tri-reference point theory of decision making under risk. J Exp Psychol Gen 141:743–756
Wang XT, Johnston V (1995) Perceived social context and risk preference: a re-examination of framing effects in a life-death decision problem. Journal of Behavioral Decision Making 8:279–293
Wang XT, Simons F, Brédart S (2001) Social cues and verbal framing in risky choice. J Behav Decis Mak 14:1–15
Whitney P, Rinehart CA, Hinson JM (2008) Framing effects under cognitive load: The role of working memory in risky decisions. Psychon Bull Rev 15:1179–1184
Zhang Y, Miao D (2008) Social cues and framing effects in risky decisions among Chinese military students. Asian J Soc Psychol 11:241–246
Zheng H, Wang XT, Zhu L (2010) Framing effects: behavioral dynamics and neural basis. Neuropsychologia 48:3198–3204
Acknowledgments
This work was partially supported by National Science Foundation (NSF) Grant SES-1123341, and a research fund from the Key Laboratory of Behavioral Science, Institute of Psychology of the Chinese Academy of Sciences to the first author, and a research fund (No. Y2CQ043005) from the Institute of Psychology of the Chinese Academy of Sciences to the second author. We thank Steven Garelik for his helpful comments on an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Wang, XT., Rao, L., Zheng, H. (2016). Framing Effects: Behavioral Dynamics and Neural Basis. In: Reuter, M., Montag, C. (eds) Neuroeconomics. Studies in Neuroscience, Psychology and Behavioral Economics. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-35923-1_9
Download citation
DOI: https://doi.org/10.1007/978-3-642-35923-1_9
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-35922-4
Online ISBN: 978-3-642-35923-1
eBook Packages: Behavioral Science and PsychologyBehavioral Science and Psychology (R0)