
Abstract
An ichnofabric includes all structure and textural changes of the sediment resulting from bioturbation (and bioerosion) at all scales. Abundance and distribution of burrows reflects the non-linear sorting effects of physical and biological parameters, resulting in a disturbance regime which generates patchiness. To analyze the patch dynamics of trace fossils as a result of environmental disturbance, this study quantifies bioturbation rates and spatial and temporal variation based on fractal geometry. Multifractal spectrum is used as a measure of spatial ichnofabric heterogeneities. The magnitude of the Daedalus ichnofabric fluctuations for the two stratigraphic sequences sampled shows that sandflat substrate colonization by the Daedalus halli worm producer after each storm event was opportunistic, mostly multigenerational, with an exclusive and significant occupation of emptied ecospace, for the purpose of meiofauna harvesting in clean sands.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others

Keywords
1 Introduction
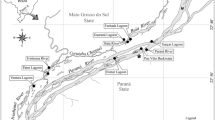
Behavior is the set of strategically and flexible responses of any phenotype for purposes of protection and transmission of its genetic legacy. This programmed responses allow the homeostatic development necessary to an organism exert some control over its ecosystem, which is intrinsically unpredictable. The fossilized remains of biological activity or ichnofossils are, in this context, the preserved solutions for the chaotic and aperiodic ecological constraints that define the evolution of behavior. The resulted programs were developed and modified by genetic pre-adaptations and by ecological parameters originally prevailing, that may have been preserved or even changed in the geological record by diagenesis imprint and tectonics. Ichnology, the discipline of paleontology focused on organism–substrate interaction dynamics (description, classification and interpretation), is a fundamental link between the evolutionary biological mechanisms and geological processes. In fact, ichnofossils represent the functional morphology and behavior of organism producers as well as the physical–chemical properties of the substrate where they were generated [1]. As ichnofossils are fossilized snapshots of behavioral and physiological functions, they translate organism producers condition to specific ecological situations. An ichnofabric includes all sediment structure and textural changes resulting from bioturbation and bioerosion at all scales [2]. Methods have been developed to improve quantification of past and present biological sedimentary reworking. Quantifying bioturbation has been a very useful tool for supporting evolutionary theories of ecological radiation, among many other applications (see for example [3]). Descriptive or semi-quantitative methods were developed for analysis of ecospace used by benthic communities in shifting substrate environments based on bioturbation percentual indices, e.g. Refs. [3–6]. Our research is based in new methods for quantifying bioturbation and ichnofabrics, based on fractal geometry, which permits, at the same time, to recognize with precision the ways in which benthic communities organize and explore their ecological niches and the ecological succession with the temporal evolution of environmental parameters. As application, we used fossil domiciliary/feeding behavior patterns classified as Daedalus halli, occasionally very abundant in siliciclastics deposited in sandflat subtidal marine bottoms almost 480 million years ago that are commonly present in the Floian-age Armorican Quartzite Formation from Portugal. For this study we quantified dense ichnofabrics of Daedalus in the sections of Serra de Barreiras Brancas [7], and Angueira valley, near Serapicos. Daedalus ichnofossils are burrows of a still unknown worm-shaped producer that reworked sands in the three spatial directions by the coiled, protrusive or retrusive displacement of a vertical-to-oblique J-tube [8, 9] intersecting bedding planes normal or at high-angle, and reaching 50 cm deep.
2 Fractal and Multifractal Estimation of Bioturbation Spatial and Temporal Heterogeneities
One of the primary goals in the use of fractals is that statistically descriptive methods were developed in order to better describe natural forms. In this way, the measure of fractal dimension of certain ichnofossils is of particularly interest in more quantitative approaches to ichnotaxonomy [10–12]. Meanders, spirals or branches in feeding biogenic structures are ways of food processing and foraging that increase extraordinarily the explored area in a restricted volume, as well as collecting information from environmental stimuli. Fractal dimension in the ichnological context determines the probability of sediment to be covered by the producer’ behavioral activity. Capacity fractal dimension estimation is particularly useful in the study of ichnofossils with complex patterns, which occur in sedimentary bedding planes. For fractal determination it is used the Box Counting theorem as described in Refs. [10, 11]. We used the free software FRACTALYSE 2.4.1 to estimate capacity dimension of monochrome sketches (.bmp) of Daedalus halli. Those images are covered with a grid of side a, the software counting the minimal number of squares N(a) which includes bioturbation. Counting should be repeated M times for different a sizes (\(\text {M}~>~2\) orders of magnitude). The absolute value of slope of the regression line adjusted to Richardson plot of logN(a) versus loga corresponds to Fractal Dimension by the relation
Standard deviation of error describes the adjustment quality of regression line. Approaching to 0, log\(N(a)\) and log\((1/a)\) both reach a very high value when compared with \(\log (const)\). In the limit when a tends to 0, it is obtained the exact definition of Hausdorff or Capacity Dimension
Multifractal describes several processes with fractal signatures (fractals or pseudofractals) when characterized at multiple levels of information. Multifractal spectra can be used as a measure of spatial or sequential ichnofabric heterogeneities. It is based on the box counting method thus enabling quantitative analysis of ecospace occupation strategies translated for behavioral forms preserved on the bedding planes. The heterogeneity measure is given by a probability distribution which can be estimated, according to [13], as
where \(N_v(a)\) is the number of identified bioturbations within the v-square grid and N is the total number of Daedalus in the digitalized image. This probabilityv fits in the equation that quantifies density distribution in Ref. [14],
where v identifies the square of side a and \({p_{v}}(a)\) registers the relative weight of v-square as expression of the total density. Generalized fractal dimensions spectra, D(q), quantify non-uniformity in ichnofossil distribution density, which is dependent of q momentum [13].
3 Discussion: Evaluating Community Strategies in Space and Time for the Daedalus Producer
Multifractal D(q) anisotropies along the \(40~\text {m}^{2 }\) bedding plane at Martim Preto shows spatial heterogeneity \((1,61~\pm ~0.07~\le ~D(q)~\le ~1.89~\pm ~0.14)\). A non-uniform distribution of ichnofossils may be consequence of intraspecific low competition for resources, which actually does not promote the regular distribution of organisms. Otherwise, discrimination of a faint clustering by multifractal analysis could be a sign of stable and favorable environmental conditions during Daedalus fabrication by producers’ population. This patchy distribution pattern opens the possibility for a model of meiofauna harvesting to explain Daedalus halli architecture based on the subtidal pump mechanism of interstitial water exchange by wave action.
Recurrence of tempestitic phenomena and its persistent effects in ecological succession can be evaluated from fractal data obtained in the Serapicos stratigraphic section. We measured bed-by-bed fractal frequence of Daedalus for 36 bedding planes. About 75.5 % of observed layers are bioturbated, with a Capacity Dimension mean value of \(\text {D}_{0}~=~1.62~\pm ~0.15\). Data shows that subtidal substrate settlement after each tempestitic event by Daedalus producers was intense, mostly multigenerational, with an exclusive and significant occupation of the emptied ecospace.
The presence of only one preserved behavioral strategy, the substrate depth affected by these structures, the high density of burrows and passive patchiness rates are characteristics of r-selected populations. Such large-scale and frequent disturbance events as storms, extreme on a gradient of disturbance intensities, were responsible for some of the earliest opportunistic behaviors in the fossil record. Daedalus was among the most resilient of them in the Early Paleozoic.
References
Pemberton, S. G., Frey, R. W., Saunders, T. D. A. (1990). Trace fossils. In D. E. G. Briggs, & P. R. Crowther (Eds.), Palaeobiology: a synthesis (pp. 355–361). Oxford : Blackwell Science.
Ekdale, A. A., & Bromley, R. G. (1983). Trace fossils and ichnofabrics in the Kjolby Gaard Marl, uppermost Cretaceous, Denmark. Bulletin of the Geological Society of Denmark, 31, 107–119.
Taylor, A., Goldring, R., & Gowland, S. (2003). Analysis and application of ichnofabrics. Earth-Science Reviews, 60, 227–259.
Reineck, H.-E. (1963). Sedimentgefuege im Bereich der suedlichen Nordsee. Abhandl. der Senckenb. Naturforsch. Gesellsh., 505, 1–138.
Droser, M. L., & Bottjer, D. J. (1986). A semiquantitative field classification of ichnofabric. Journal of Sedimentary Petrology, 56, 558–559.
Miller, M. F., & Smail, S. E. (1997). Multifractal characterization of microbially induced magnesian calcite formation in Recent tidal flat sediments. Palaios, 12, 391–396.
Sá, A. A., Meireles, C., & Coke, C. (2002). Concentração maciça de Daedalus labechei (Rouault) (Icnofóssil Ordovícico) no Alto do Martim Preto (Guadramil-Bragança): Património paleontológico a preservar e divulgar. In J. Civis , & J. A. González Delgado (Eds.), XVIII Jornadas de la Sociedad Española de Paleontología/II Congreso Ibérico de Paleontología (pp. 138–139). Salamanca: Libro de Resúmenes.
Durand, J. (1985). Le Grés Armoricain. Sédimentologie, Traces Fossiles, Milieux de dépôt. Mem. et Doc. du Centr. Armor. d’Etude Struct. des Socles 3, pp. 1–119.
Seilacher, A. (2000). Ordovician and Silurian arthrophycid ichnostratigraphy. In: M. A. Sola, & D. Worsley (Eds.), Geological exploration in Murzuq Basin (pp. 237–258). Amsterdam: Elseiver.
Gibert, J. M., Jeong, K., & Martinell, J. (1999). Ethologic and ontogenetic significance of the Pliocene trace fossil Sinusichnus sinuosus from the northwestern Mediterranean. Lethaia, 32, 31–40.
Neto de Carvalho, C. (2006). Roller coaster behaviour in the Cruziana rugosa group from Penha Garcia (Portugal): implications for the feeding program of Trilobites. Ichnos, 13(4), 255–265.
Baucon, A. (2010). Da Vinci’s Paleodictyon: the fractal beauty of traces. Acta Geologica Polonica, 60(1), 3–17.
Kropp, J., Block, A., Bloh, W., Klenke, T., & Schellnhuber, H. J. (1997). Multifractal characterization of microbially induced magnesian calcite formation in Recent tidal flat sediments. Sedimentary Geology, 109, 37–51.
Hentschel, H. G. E., & Procaccia, I. (1983). The infinite number of generalized dimensions of fractals and strange atractors. Physica D, 8, 4435–4444.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this paper
Cite this paper
Neto de Carvalho, C., Baucon, A. (2014). Multifractals and Capacity Dimension as Measures of Disturbance Patch Dynamics in Daedalus Ichnofabrics. In: Pardo-Igúzquiza, E., Guardiola-Albert, C., Heredia, J., Moreno-Merino, L., Durán, J., Vargas-Guzmán, J. (eds) Mathematics of Planet Earth. Lecture Notes in Earth System Sciences. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-32408-6_161
Download citation
DOI: https://doi.org/10.1007/978-3-642-32408-6_161
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-32407-9
Online ISBN: 978-3-642-32408-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)