
Abstract
After being formed, plant cells increase in volume during the cell expansion process in which the cell wall plays a prominent role. Cell wall extensibility, a crucial characteristic that defines plant cell growth rate at the biophysical level, is currently estimated by the use of several indirect techniques. One of them, the creep method, measures cell wall extension under a constant load over time in the principal direction of cell and organ growth and mimics the action of turgor on growing cell walls better than other techniques. This review provides the last technical advances and novel metrics of the creep method predicting cell wall extensibility. The potential of the creep method to uncover new cell wall-loosening/tightening proteins is discussed.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
Cell Wall Extensibility and Its Estimation
Plants have evolved a unique mechanism of cell enlargement, the so-called expansive growth, which is largely mediated by water uptake. Water is absorbed by growing plant cells as a result of higher osmotic concentrations of their contents compared with the extracellular solution. The resulting increase in volume is restrained by stiff yet extensible cell walls that encase plant cells. The wall resistance to the osmotically mediated protoplast enlargement generates high turgor pressure, which in turn drives the irreversible cell wall expansion that accompanies growth. The involvement of cell walls in expansive growth was clarified by Lockhart (1965) and expressed in the following equation:
where (1/V)(dV/dt) is the relative cellular expansive growth rate expressed as a change in cell volume (V), Φ is the wall yielding coefficient characterizing the sensitivity of growth rate to changes in turgor, P is the turgor pressure and Y is the yield threshold, i.e., the minimal turgor required for growth (Fig. 1). The Lockhart Eq. (1) postulates that the growth rate is a linear function of turgor in excess of the yield threshold. The terms Φ and Y, both characterizing growth-relevant cell wall properties, determine its extensibility, i.e., the ability of the cell wall to irreversibly increase in surface area during growth (Cosgrove 2016a). Numerous studies conducted in the 1970–1980s convincingly demonstrated that cell wall extensibility is a limiting biophysical factor which defines the rate of cellular expansive growth (reviewed in Cosgrove 1986). It was cell wall extensibility that changed rapidly under many different conditions/treatments that induced alterations in expansive growth rate. Thus, multiple signal transduction pathways involved in the control of cell expansion seem to converge at the cell wall level with cell wall extensibility being their most downstream point, the last factor that changes before a cell attains a new growth rate. Cell wall extensibility depends on the biochemical composition of wall polymers, as well as on numerous cell wall-loosening and tightening proteins that break and form covalent and non-covalent bonds between wall constituents (Chebli and Geitmann 2017).

Graphic representation of the Lockhart equation and two hypothetical ways of plant cell growth stimulation. The wall yielding coefficient (Φ) characterizes the sensitivity of growth rate to changes in turgor and corresponds to the slopes of the lines. The yield threshold (Y) shows the minimal turgor at which cell expansive growth starts and corresponds to the intercepts of the lines with the x axis. The original moderate growth rate (thick line) is limited by Φ1 and Y1. Growth stimulation via increasing Φ (dashed line) where Φ2 > Φ1 and Y does not change. Growth stimulation via decreasing Y (thin line) where Y2 < Y1 and Φ does not change. As a matter of fact, independent changes in Φ and Y values can occur during growth stimulation or suppression in vivo
Given the recognized pivotal role of cell wall extensibility in the biophysical control of cellular expansive growth, measuring or estimating this characteristic is of utmost importance. Cell wall extensibility was quantified in vivo by several methods developed in the 1960–1980s, such as “growth rate versus turgor”, in vivo stress relaxation, and some others (reviewed in Cosgrove 1993). These techniques are not used at present anymore because they are laborious and incompatible with many experimental treatments required. Instead, cell wall extensibility is currently estimated indirectly by classical microtensile tests: the creep method (Cosgrove 1989; Thompson 2001; Suslov and Verbelen 2006), the stress/strain method (Ryden et al. 2003; Abasolo et al. 2009), the in vitro stress relaxation (Yamamoto 1996), as well as by more modern techniques based on nano- and microindentation of plant surfaces including atomic force microscopy (AFM) (Milani et al. 2013; Peaucelle et al. 2011, 2015) and cellular force microscopy (CFM) (Routier-Kierzkowska et al. 2012). All these techniques provide different metrics of cell wall biomechanics, and only some of them correlate with cell wall extensibility. Thus, the different biomechanical methods and their resulting metrics are not equivalent in their ability to explain growth rate in vivo.
This brief review is devoted to the creep (constant load) method and its use for the analysis of growth-relevant cell wall biomechanics in plants. We will compare it with alternative methods and consider practical aspects of its application and peculiarities of data interpretation, and discuss prospects for further development of this old, yet efficient technique.
We focus on the creep method because its outputs are easier to interpret and link with cell wall extensibility and in vivo growth rates when compared with the abovementioned alternative techniques of plant biomechanics. The creep method is based on measuring the wall deformation under a constant load over time. The stress [i.e., the force on a body divided by the cross-sectional area across which the force acts (Baskin 2005)] generated in cell walls during creep measurements resembles the in vivo stress established by turgor in that they both are essentially constant over time, as turgor does not vary appreciably during cell expansion under normal conditions (Taiz 1984; Cosgrove 1986). In contrast to the creep method, execution of all the alternative techniques causes rapid and large changes in wall stress, which is clearly inconsistent with the in vivo conditions. Another advantage of the creep method is that the rate of wall deformation under a constant load is generally slow and comparable to the rate of cellular expansive growth in vivo (Richmond et al. 1980; Suslov et al. 2015). As a result, endogenous cell wall-loosening/tightening proteins that are thought to be implicated in wall extensibility control in vivo have sufficient time to exert their effects under the conditions of creep measurements. On the contrary, the alternative techniques use very rapid cell wall deformations being ill-suited for studying the growth-relevant effects of cell wall proteins. Although the spatial resolution of the creep method is much lower than that of AFM and CFM techniques, the former has two major advantages over the latter: (i) in creep measurements cell walls are extended along the principal direction of growth in vivo, while in AFM and CFM the walls are deformed orthogonal to the growth vector (Milani et al. 2013; Cosgrove 2016a); (ii) creep tests mostly reveal irreversible (plastic) cell wall deformations (Suslov and Verbelen 2006), which can be directly linked with growth, an irreversible process, while AFM and CFM measure predominantly reversible (elastic) cell wall deformations having no clear link with growth itself (Peaucelle et al. 2011).
All the abovementioned advantages of the creep method predetermined its main application in modern plant (cell wall) biology. The technique has been successfully used to confirm the involvement of a certain cell wall protein of interest in the control of cell wall extensibility. If the purified protein as such increases or decreases cell wall deformation in the creep test, this finding is considered as a direct confirmation of its role as a cell wall-loosening or cell wall-tightening protein, respectively. Thus, the considered protein is highly probable to work as a natural regulator of cell wall extensibility. This approach has shed light on the functions of expansins (McQueen-Mason et al. 1992), yieldins (Okamoto-Nakazato et al. 2000), lipid transfer proteins (Nieuwland et al. 2005), and xyloglucan endotransglucosylase/hydrolases (XTHs) (Van Sandt et al. 2007; Maris et al. 2009) in the control of cell wall extensibility and plant growth.
Preparation of Plant Samples for Creep Tests
Creep tests have been carried out using either living turgid plant organs or isolated cell walls. The most widely used approach for cell wall isolation involves freezing and thawing of plant organs or their segments. This treatment destroys cell membranes, eliminates turgor and yields only relaxed cell walls (Cosgrove 1989; Thompson 2001; Suslov and Verbelen 2006). The samples prepared in this way are considered to retain a significant part of cell wall protein activities implicated in the control of cell wall extensibility, and subsequent constant load application partially mimics the action of turgor on cell walls. This approach is convenient as it allows rapid fixation of large batches of plant material that is uniform by age, which is critical for data analysis and interpretation. The frozen samples can then be stored and analyzed within up to two weeks without any appreciable loss in cell wall properties (Miedes et al. 2013). However, this approach provides only a static picture of cell wall properties at the moment of freezing without any contribution from cellular metabolism (the secretion of new cell wall-loosening/tightening proteins, in vivo changes in apoplastic pH and reactive oxygen species production). Additionally, some cell wall proteins do not survive the freezing/thawing cycle, although this problem can be circumvented by an alternative protocol involving cell wall glycerination (Okamoto-Nakazato 2002). At first glance, the use of living plant organs in creep tests can address the above problems with frozen/thawed samples. In this case, the wall creep will also be controlled by metabolic inputs from living protoplasts (Kutschera and Schopfer 1986b; Nonami and Boyer 1990). However, the retained metabolic activity may create different problems. First, physiological stress responses will be induced as a result of mechanical disturbances of living experimental samples before and during creep measurements. Some of them, e.g., reactive oxygen species (ROS) production, could rapidly modify cell wall properties (Monshausen et al. 2009). Second, an extra load applied to the fully turgid living organs is physiologically irrelevant as changes in their growth rate are not normally caused by variations in the force (turgor) extending the wall in vivo (Cosgrove 1986). Finally, it is not easy to obtain many uniform living samples with respect to their developmental stage, as creep measurements are time-consuming, while age-related changes in cell expansion rates and growth zone positions can be very rapid (Bastien et al. 2016). Due to all these reasons, cell walls isolated after a freeze/thaw cycle are currently used more frequently.
Plant samples have to be highly permeable for exogenous buffers and proteins in many applications of the creep method. Traditionally this is achieved by peeling or abrading the sample surfaces (Rayle and Cleland 1992). The peeling procedure involves the complete removal of the epidermis with the use of a fine forceps, thereby exposing the hydrophyllic surfaces of underlying cell walls that are stretched under a constant load. Although peeling can be highly reproducible, it eliminates the tissue that plays a critically important role in growth control (Kutschera and Niklas 2007). So, at present it is more common to use epidermal peels in creep tests (Thompson 2001; Suslov et al. 2009), i.e., isolated epidermal cell layers, which are permeable even for exogenous proteins (Van Sandt et al. 2007; Maris et al. 2009). During the abrasion procedure, the hydrophobic cuticle covering the outer epidermal cell wall is removed by gentle rubbing with fine abrasive materials (Kutschera and Schopfer 1986a; Park and Cosgrove 2012a). This treatment does not damage the growth-controlling outer epidermal cell wall. However, abrasion may be less reproducible than peeling, when performed by different people. Finally, some organs like roots are highly permeable for exogenous solutes by their nature and can be used in creep tests without any treatments increasing their permeability (Büntemeyer et al. 1998).
Another preliminary treatment of frozen/thawed samples before creep tests involves their flattening under a high load to remove excessive water (Durachko and Cosgrove 2009), since water expelling from plant samples may affect their extension in the first minutes of creep. The flattening procedure is normally not required for experimental samples with a low volume per length ratio like epidermal peels or miniature Arabidopsis hypocotyls.
Estimating the effects of exogenous proteins on cell wall extensibility with the creep method usually requires the inactivation of endogenous proteins in experimental samples. Traditionally they are heat-inactivated by boiling in water for 15 s (McQueen-Mason et al. 1992; Van Sandt et al. 2007). A more gentle treatment is used with more fragile Arabidopsis hypocotyls that are heated under sub-boiling temperatures followed by rehydration at 4 °C (Boron et al. 2015; Suslov et al. 2015).
Comparisons of the mechanics of plant samples differing in their cross-sectional area are often made with the creep method (Miedes et al. 2013). In these specific cases, the same constant load will generate a different stress in the cell walls that need to be compared. As a consequence, their comparison will only be relevant if these stress values are taken into account, as the rate of sample deformation is proportional to the stress in it. To calculate stress in the walls during creep tests, the force arising from a constant load should be divided by the wall cross-sectional area. This force is directly defined with a load cell or calculated based on the fact that 1 N (Newton) force is generated by 102 g load under normal gravity. The cross-sectional area can be measured using electron micrographs of freeze-fractured samples (Derbyshire et al. 2007) in a direct, but very time-consuming approach, or alternatively, it can be calculated indirectly by measuring the wall dry weight per sample unit length, taking into account its geometry (Cleland 1967) and assuming that the density of wall polymers is 1.5 g cm−3 (Gibson 2012). This approach combined with exhaustive extraction of low molecular weight compounds has recently been adapted for cell walls of Arabidopsis hypocotyls (Miedes et al. 2013; Suslov et al. 2015).
Practical Aspects of Cell Wall Creep Measurements
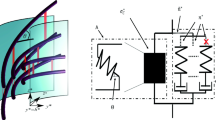
Plant samples are extended in creep tests using extensometers. The setup we use is shown in Fig. 2, whereas other designs of extensometers were described in Kutschera and Schopfer (1986a), Cosgrove (1989), Okamoto and Okamoto (1994), and Takahashi et al. (2006). Experimental samples are secured in the setup with instant glue or mechanical clamps and placed in a watertight chamber, where they will be extended during creep tests while being submerged in buffers. Further technical details of tensile testing are elaborated in Chap. 10. Keeping the samples in a well-hydrated state during measurements is critically important, as this significantly affects their extension (Evered et al. 2007). Buffers are selected such that their maximal buffering capacity corresponds to physiological pH values found in the plant apoplast (pH from 4.5 to 7.0) (Bibikova et al. 1998; Monshausen et al. 2007). Additionally, the buffers should not have any significant calcium-chelating capacity, since this could influence cell wall mechanics (Cosgrove 2011). These conditions are met with buffers based on MES, acetate, Hepes, and histidine.

Schematic drawing of the extensometer used in our onion epidermis and Arabidopsis hypocotyl work. The space where the hypocotyl segment resides is filled with buffers at different pH and can be supplemented with proteins of interest to reveal their effect on the measured extension of the segment
A constant load is applied to the sample secured in the extensometer using a load cell (Takahashi et al. 2006) or scale weights (Fig. 2; Durachko and Cosgrove 2009). The load chosen should ideally generate a wall stress that is comparable to those relevant in vivo. Excessive loads causing frequent cell wall failures have to be avoided. At the other extreme, too low loads resulting in cell wall deformation rates well below the in vivo growth rate are also irrelevant. Practically and as an example, in our work with Arabidopsis hypocotyls, we select loads such that the maximal load should induce no more than 30% of sample failures (Boron et al. 2015). The minimal load in our case is limited by the system resolution and normally produces creep rates that are four to eight times lower than the in vivo growth rate.
Standard creep tests give raw results in the form of biphasic extension curves including a high-amplitude initial deformation just after loading, followed by a slow time-dependent deformation referred to as “creep” (Fig. 3). Initial deformation is not a reliable predictor of cell wall extensibility, as it normally contains a very significant (Suslov and Verbelen 2006) or even predominant (Nolte and Schopfer 1997) elastic component. Still, initial deformation does relate to the rate of cell expansion under some conditions, e.g., in the case of auxin-induced growth (Kutschera and Schopfer 1986a) or age-related growth deceleration (Miedes et al. 2013). On the other hand, creep has long been considered as a process closely resembling in vivo extension of cell walls (Cosgrove 2005). So the majority of metrics obtained with the constant load method are currently based on the analysis of creep.

Typical rough deformation curve obtained using the creep method
Traditional and Novel Metrics of the Creep Method
Creep rate (expressed in μm h−1, % h−1 or h−1) is the simplest and most commonly used metric of the constant load method. It is reliable when control and test cell wall samples have similar cross-sectional areas, i.e., any given constant load will generate the same stress in them. As was mentioned above, creep rates should ideally be compared under constant loads producing wall stress comparable to that existing in vivo. However, the actual in vivo wall stress is difficult to measure because it is not defined solely by turgor but also depends on tissue tensions (Hejnowicz and Sievers 1995), cell geometry (Baskin 2005) and stress anisotropy within multicellular plant organs (Baskin and Jensen 2013). To cope with this problem, creep rates are often compared under many different loads with the assumption that one of them does generate the wall stress close to that in vivo (Thompson 2001; Suslov and Verbelen 2006; Vandenbussche et al. 2011).
When cross-sectional areas of cell walls are different, creep rates are poor predictors for in vivo growth rates (Miedes et al. 2013). The simplest solution, in this case, is to compare the ratios creep rate × stress−1. The fact that the ratios are defined by two independent variables (creep rate and stress) complicates their statistical comparisons. This problem has been recently solved by creating scripts that allow comparison of creep rate × stress−1 values using a novel algorithm (Suslov et al. 2015) based on bootstrapping.
Sometimes the creep rate depends on load (or wall stress) in a linear fashion (Okamoto and Okamoto 1994; Miedes et al. 2013; Suslov et al. 2015). In this particular case, two additional metrics of the creep method can be obtained: φ (in vitro cell wall extensibility) and y (in vitro cell wall yield threshold), i.e., in vitro analogs of the parameters Φ and Y from the Eq. (1). The value of φ characterizes the sensitivity of creep rate to changes in the load (or wall stress) and corresponds to the slope of the line in Fig. 4, while y shows the minimal load (or wall stress) at which creep starts and corresponds to the intercept of the line with the x axis (Fig. 4). The values of φ and y can be obtained based on serial loading (Okamoto and Okamoto 1994) or from independent creep measurements under different loads (Suslov et al. 2015). These alternative approaches give nonequivalent results (Takahashi et al. 2006; Suslov et al. 2015), and their critical comparison is given in Suslov et al. (2015). Although the latter approach (Suslov et al. 2015) is more laborious, its results are less affected by experimental conditions than in the former approach (Okamoto and Okamoto 1994; Takahashi et al. 2006). The advantage of φ and y over the other metrics of the creep method consists in their ability to give an insight into the mechanisms of changes in cell wall extension. For example, the expansin-mediated acid growth response is caused by increasing the φ values (Suslov et al. 2015), while XTH overexpression activates growth by lowering y (Miedes et al. 2013). Thus, φ and y determination may forecast that certain growth responses are induced by different cell wall proteins before they are revealed using transcriptomics or proteomics, purified, and their cell wall-loosening/tightening function is confirmed in the reconstitution creep assay.

Determination of φ (slopes of the lines) and y (intercepts with the x axis) values using independent creep measurements under different loads. Hypocotyls from frozen/thawed Columbia-0 Arabidopsis plants were extended at pH 5 or 6 under one out of four constant loads (400, 500, 600 and 700 mg). Average creep rate for each load was calculated. Then the loads were transformed into respective wall stress values based on cell wall cross-sectional area estimation. Average creep rates were plotted against average wall stress values generated under each constant load. The dependence of creep rate on wall stress was linear as determined by Deming regression taking into account variation in both creep rate and wall stress. The plot demonstrates that the acid-induced expansin-mediated cell wall extension in Arabidopsis hypocotyls results from an increase in φ
Cell wall-loosening/tightening proteins may affect not only the rate but also the duration of creep in plant samples. For example, expansins seem to increase both of them (Cosgrove 1989). The temporal aspect of cell wall creep is often not taken into account, in part because its straightforward estimation requires hours-long creep recordings, which is very inconvenient. The importance of the temporal aspect was underscored by demonstrating that XTH overexpression renders creep curves more linear in time (Miedes et al. 2013). This finding has an implication that XTH enzymes may prolong cell expansive growth. Thus, there should be metrics describing creep curve curvature, i.e., changes in cell wall creep rates in time. For example, one can measure the first derivative of creep rate with respect to time for different points along a creep curve. This approach was applied for treatments stimulating, inhibiting or having no effect on growth of Arabidopsis hypocotyls (Suslov et al. 2012). In the case of growth stimulation, the derivative increased to values approaching nil demonstrating the practically linear dependence of creep rate on time. The growth inhibition rendered the derivative more negative, consistent with a faster decay of creep rate over time. In the absence of growth effect, no significant differences in the derivative were found. Further studies are needed to confirm that this metric of creep curve curvature can be used as a universal predictor of in vivo growth rate.
Cell wall extension involves plastic (irreversible) and elastic (reversible) components, which both can be estimated with the creep method. This is achieved by using loading/unloading cycles wherein one or more loads are first sequentially added to the wall sample and then sequentially removed from it, whilst the resulting cell wall extension/contraction is continuously recorded (Kutschera and Schopfer 1986a; Hejnowicz and Sievers 1996; Suslov and Verbelen 2006). These experiments provided rather conflicting results. Some authors argued that the plastic component represents in fact retarded elasticity (Hohl and Schopfer 1992; Nolte and Schopfer 1997), while others found true plastic deformations (Thompson 2001; Suslov and Verbelen 2006). It is not clear whether this disagreement reflects natural variation between the plant objects studied, or it is imposed by the different protocols. However, knowing the exact relationship between plastic and elastic components of cell wall extension is critically important for correct data interpretation. All modern nano- and microindentation-based techniques only measure cell wall elasticity (Braybrook 2017). The creep method can answer the question whether the plastic and elastic components change in a concerted manner during variations in growth rate. If these components do positively correlate, then the metrics of AFM and CFM techniques provide growth-relevant information. In the absence of such correlation, the outputs of nano- and microindentation tests should be interpreted with extreme care when attempting to link them with in vivo growth rates. Interestingly, the dynamic interconversion between plastic and elastic components of wall deformation was found during cell expansive growth (Edelmann and Köhler 1995). It was concluded that growth may be regulated by wall-loosening mechanisms which initially result in elastic wall extension. Later, this extension becomes irreversible via a fixation process (Edelmann 1995), for which the mechanism was also proposed (Hohl et al. 1995). To sum up, no generally accepted metrics are currently available to describe plastic versus elastic components of cell wall extension. The new metrics to be developed have to be universal for plant samples of different origin. Establishing the objective criteria for their direct comparisons is a challenging task because there are apparently no natural equivalents to the wall perturbations occurring during loading/unloading cycles. So it is not clear to what extent cell walls should be loaded and unloaded, and how long each loading/unloading step should last to provide accurate estimations of cell wall plastic versus elastic properties with the creep method. If no objective criteria exist, there should be a kind of convention on the measurement conditions between specialists in the field.
Novel Trends and Improvements of the Creep Method
Although the wall stress generated in a standard uniaxial creep test and the in vivo stress are similar in that they are essentially constant over time, their nature is different. Turgor induces biaxial stress in the plane of the wall, which limits its polymer reorientations due to geometrical constraints (Chanliaud et al. 2002). On the contrary, the uniaxial stress generated by a constant load results in major polymer reorientations in the direction of cell wall deformations. Normally this leads to much higher cell wall extension compared to that caused by equal biaxial stress. However, the action of in vivo biaxial stress and in vitro uniaxial stress along the main axis of elongation in organs that consist of highly anisotropically growing cells may not be so dramatically different. This is because the polymer reorientations induced by a constant load are very similar to those occurring in these organs in vivo (Refrégier et al. 2004; Suslov et al. 2009; Crowell et al. 2011). On the other hand, isotropic growth is not normally accompanied by polymer realignment. In organs composed of isotropically growing cells, the mechanical effect of uniaxial stretching could differ dramatically from the effect of turgor. This problem has long been recognized, and it was solved by designing the biaxial stretching systems. The first successful biaxial creep measurements were performed using giant internodal cells of Nitella (Métraux et al. 1980; Richmond et al. 1980). The walls of cylindrical segments of these cells containing intact apical nodes were inflated with mercury, and their resulting extension in length and in girth was monitored microscopically. This biaxial stretching system cannot be universally used, as it is based on the extraordinarily large size and specific morphology of Nitella internodal cells. Chanliaud et al. (2002) proposed an approach with more universal application possibilities. They designed a bulge-testing device in which a flat sample of Acetobacter xylinus composite (a biomimetic of plant cell walls) was placed over an orifice of a cylindrical chamber. This chamber was pressurized and the resulting sample bulging was measured. Essentially the same approach was recently applied to the walls of cherry epidermal peels (Brüggenwirth et al. 2014), and their creep resulting from biaxial stretching was studied under different conditions (Brüggenwirth and Knoche 2016). Biaxial cell wall stress generated in bulge-testing devices or their analogs mimics the action of turgor on growing cell walls much better than the uniaxial stress generated in traditional uniaxial creep tests. Until now, the bulging test was only used with mechanically isotropic cell walls. So this efficient technique should be further adapted for anisotropic samples and this will require new algorithms to analyze complex surfaces. One can imagine that the biaxial stretching of anisotropic samples will produce symmetric distortions of their bulging surfaces that should be quantified. At the next step of improvement, the bulging test should allow reconstitution assays with purified cell wall proteins. This would help confirm their role in the control of anisotropic cell expansion. Until now, there have been very scarce data on proteins that can differentially affect cell wall extensibility along two perpendicular axes in the plane of the cell wall (Wiedemeier et al. 2002; Van Sandt et al. 2007).
Traditional creep tests have limited spatial resolution. The minimal length of sample segments to be stretched is about 2 mm. This means that the sample should be at least 4–6 mm long, as extra length is needed to secure it in the setup. If one wants to study creep in a number of model plant objects, such as root growth zones, light-grown Arabidopsis hypocotyls and etiolated hypocotyls from different Arabidopsis mutants, these are shorter than the abovementioned limit and can therefore not be stretched. Thus, an increase in the spatial resolution of the creep method is highly demanded. Fortunately, this seems to be technically feasible as judged from recent advances in the stress/strain method. Zamil et al. (2013) achieved subcellular resolution with a novel tensile testing device based on a microelectromechanical system. They have obtained stress/strain curves from as small as 15 × 5 μm pieces of onion epidermal cell walls. Original measurements were performed on dried samples, as cell wall displacement was monitored by scanning electron microscopy. Later, the technique was improved and allowed the use of hydrated cell walls (Zamil et al. 2014, 2015; Kim et al. 2015). The ideas and technical solutions of this group can serve as a basis for the development of a novel high-resolution creep-testing device.
Despite the central role of Arabidopsis thaliana in modern plant biology, the creep method was first used with this popular object only quite recently and later than other biomechanical techniques (Vandenbussche et al. 2011). This is because its growing organs are small and fragile, which makes their usage rather difficult. The majority of technical issues have been successfully solved since then and the method is now routinely used in studies of Arabidopsis mutants (Park and Cosgrove 2012a, b) and overexpression lines (Miedes et al. 2013; Boron et al. 2015). The full potential of the creep method can be realized with cell wall mutants of Arabidopsis and their analysis should give insights into the functions of certain cell wall polymers, their interactions with each other, and this will finally help to create a realistic model of the cell wall architecture. The first steps on this way have already been done casting doubt on the traditional view of cell wall organization (Park and Cosgrove 2012a, b). Further progress in the field will require to increase the method resolution and to use new metrics for comparisons. The former will make new cell wall mutants available for analysis, while the latter will help reveal very subtle differences between them. The creep method is very promising for a screen of new cell wall-loosening/tightening proteins in Arabidopsis, keeping in mind that a huge amount of omics data is available for this plant species. The only pitfall for this application is the lack of an efficient reconstitution assay based on Arabidopsis organs. Although the abrasion technique has been used with Arabidopsis hypocotyls (Park and Cosgrove 2012b) to increase their permeability for exogenous proteins, this treatment may potentially damage these tender organs. Thus, an alternative and more gentle protocol should be developed. After it is available, a technology for homologous reconstitution with endogenous Arabidopsis cell wall proteins can be created. In this case, a knockout Arabidopsis plant for a protein of interest is produced, after which the purified protein of interest is applied to this knockout in a creep test to verify whether biomechanical complementation takes place. Finally, it would also be very productive to study Arabidopsis hypocotyls or roots in different growth states by using the creep method in combination with modern AFM or CFM techniques. This combination approach could demonstrate which AFM or CFM metrics are the best indicators of growth-relevant cell wall properties.
To sum up, despite the very simple principle and long history of use, the creep method remains evergreen in the field of plant biomechanics. The advent of novel instruments that will increase the method’s spatial resolution along with better metrics for presenting its outputs will greatly improve our understanding of the mechanisms of cell expansive growth.
References
Abasolo W, Eder M, Yamauchi K et al (2009) Pectin may hinder the unfolding of xyloglucan chains during cell deformation: implications of the mechanical performance of Arabidopsis hypocotyls with pectin alterations. Mol Plant 2:990–999
Baskin TI (2005) Anisotropic expansion of the plant cell wall. Annu Rev Cell Dev Biol 21:203–222
Baskin TI, Jensen OE (2013) On the role of stress anisotropy in the growth of stems. J Exp Bot 64:4697–4707
Bastien R, Legland D, Martin M et al (2016) KymoRod: a method for automated kinematic analysis of rod-shaped plant organs. Plant J 88:468–475
Bibikova TN, Jacob T, Dahse I et al (1998) Localized changes in apoplastic and cytoplasmic pH are associated with root hair development in Arabidopsis thaliana. Development 125:2925–2934
Boron AK, Van Loock B, Suslov D et al (2015) Over-expression of AtEXLA2 alters etiolated arabidopsis hypocotyl growth. Ann Bot 115:67–80
Braybrook SA (2017) Analyzing cell wall elasticity after hormone treatment: an example using tobacco BY-2 cells and auxin. Methods Mol Biol 1497:125–133
Brüggenwirth M, Knoche M (2016) Factors affecting mechanical properties of the skin of sweet cherry fruit. J Am Soc Hortic Sci 141:45–53
Brüggenwirth M, Fricke H, Knoche M (2014) Biaxial tensile tests identify epidermis and hypodermis as the main structural elements of sweet cherry skin. AoB Plants 6:plu019
Büntemeyer K, Lüthen H, Böttger M (1998) Auxin-induced changes in cell wall extensibility of maize roots. Planta 204:515–519
Chanliaud E, Burrows KM, Jeronimidis G, Gidley MJ (2002) Mechanical properties of primary plant cell wall analogues. Planta 215:989–996
Chebli Y, Geitmann A (2017) Cellular growth in plants requires regulation of cell wall biochemistry. Curr Opin Cell Biol 44:28–35
Cleland R (1967) Extensibility of isolated cell walls: measurement and changes during cell elongation. Planta 74:197–209
Cosgrove D (1986) Biophysical control of plant cell growth. Ann Rev Plant Physiol 37:377–405
Cosgrove DJ (1989) Characterization of long-term extension of isolated cell walls from growing cucumber hypocotyls. Planta 177:121–130
Cosgrove DJ (1993) Wall extensibility: its nature, measurement and relationship to plant cell growth. New Phytol 124:1–23
Cosgrove DJ (2005) Growth of the plant cell wall. Nat Rev Mol Cell Bio 6:850–861
Cosgrove DJ (2011) Measuring in vitro extensibility of growing plant cell walls. Methods Mol Biol 715:291–303
Cosgrove DJ (2016) Plant cell wall extensibility: connecting plant cell growth with cell wall structure, mechanics, and the action of wall-modifying enzymes. J Exp Bot 67:463–476
Crowell EF, Timpano H, Desprez T, Franssen-Verheijen T, Emons AM, Höfte H, Vernhettes S (2011) Differential regulation of cellulose orientation at the inner and outer face of epidermal cells in the Arabidopsis hypocotyl. Plant Cell 23:2592–2605
Derbyshire P, Findlay K, McCann MC et al (2007) Cell elongation in Arabidopsis hypocotyls involves dynamic changes in cell wall thickness. J Exp Bot 58:2079–2089
Durachko DM, Cosgrove DJ (2009) Measuring plant cell wall extension creep induced by acidic pH and by alpha-expansin. JoVE 25:e1263
Edelmann HG (1995) Wall extensibility during hypocotyl growth: A hypothesis to explain elastic-induced wall loosening. Physiol Plant 95:296–303
Edelmann HG, Köhler K (1995) Auxin increases elastic wall-properties in rye coleoptiles: implications for the mechanism of wall loosening. Physiol Plant 93:85–92
Evered C, Majevadia B, Thompson DS (2007) Cell wall water content has a direct effect on extensibility in growing hypocotyls of sunflower (Helianthus annuus L.). J Exp Bot 58:3361–3371
Gibson LJ (2012) The hierarchical structure and mechanics of plant materials. J R Soc Interf 9:2749–2766
Hejnowicz Z, Sievers A (1995) Tissue stresses in organs of herbaceous plants II. Determination in three dimensions in the hypocotyl of sunflower. J Exp Bot 46:1045–1053
Hejnowicz Z, Sievers A (1996) Tissue stresses in organs of herbaceous plants—III. Elastic properties of the tissues of sunflower hypocotyl and origin of tissue stresses. J Exp Bot 47:519–528
Hohl M, Schopfer P (1992) Physical extensibility of maize coleoptile cell walls: apparent plastic extensibility is due to elastic hysteresis. Planta 187:498–504
Hohl M, Greiner H, Schopfer P (1995) The cryptic-growth response of maize coleoptiles and its relationship to H2O2-dependent cell wall stiffening. Physiol Plant 94:491–498
Kim K, Yi H, Zamil MS, Haque MA, Puri VM (2015) Multiscale stress-strain characterization of onion outer epidermal tissue in wet and dry states. Am J Bot 102:12–20
Kutschera U, Niklas KJ (2007) The epidermal-growth-control theory of stem elongation: an old and a new perspective. J Plant Physiol 164:1395–1409
Kutschera U, Schopfer P (1986a) Effect of auxin and abscisic acid on cell wall extensibility in maize coleoptiles. Planta 167:527–535
Kutschera U, Schopfer P (1986b) In vivo measurement of cell wall extensibility in maize coleoptiles: effects of auxin and abscisic acid. Planta 169:437–442
Lockhart JA (1965) An analysis of irreversible plant cell elongation. J Theor Biol 8:264–275
Maris A, Suslov D, Fry SC et al (2009) Enzymic characterization of two recombinant xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Arabidopsis and their effect on root growth and cell wall extension. J Exp Bot 60:3959–3972
McQueen-Mason S, Durachko DM, Cosgrove DJ (1992) Two endogenous proteins that induce cell wall extension in plants. Plant Cell 4:1425–1433
Métraux JP, Richmond PA, Taiz L (1980) Control of cell elongation in Nitella by endogenous cell wall pH gradients: multiaxial extensibility and growth studies. Plant Physiol 65:204–210
Miedes E, Suslov D, Vandenbussche F et al (2013) Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics in etiolated Arabidopsis hypocotyls. J Exp Bot 64:2481–2497
Milani P, Braybrook SA, Boudaoud A (2013) Shrinking the hammer: micromechanical approaches to morphogenesis. J Exp Bot 64:4651–4662
Monshausen GB, Bibikova TN, Messerli MA et al (2007) Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs. P Natl Acad Sci USA 104:20996–21001
Monshausen GB, Bibikova TN, Weisenseel MH et al (2009) Ca2 + regulates reactive oxygen species production and pH during mechanosensing in Arabidopsis roots. Plant Cell 21:2341–2356
Nieuwland J, Feron R, Huisman BAH et al (2005) Lipid transfer proteins enhance cell wall extension in tobacco. Plant Cell 17:2009–2019
Nolte T, Schopfer P (1997) Viscoelastic versus plastic cell wall extensibility in growing seedling organs: a contribution to avoid some misconceptions. J Exp Bot 48:2103–2107
Nonami H, Boyer JS (1990) Wall extensibility and cell hydraulic conductivity decrease in enlarging stem tissues at low water potentials. Plant Physiol 93:1610–1619
Okamoto H, Okamoto A (1994) The pH-dependent yield threshold of the cell wall in a glycerinated hollow cylinder (in vitro system) of cowpea hypocotyl. Plant Cell Environ 17:979–983
Okamoto-Nakazato A (2002) A brief note on the study of yieldin, a wall-bound protein that regulates the yield threshold of the cell wall. J Plant Res 115:309–313
Okamoto-Nakazato A, Nakamura T, Okamoto A (2000) The isolation of wall-bound proteins regulating yield threshold in glycerinated hollow cylinders of cowpea hypocotyls. Plant Cell Environ 23:145–154
Park YB, Cosgrove DJ (2012a) Changes in cell wall biomechanical properties in the xyloglucan-deficient xxt1/xxt2 mutant of Arabidopsis. Plant Physiol 158:465–475
Park YB, Cosgrove DJ (2012b) A revised architecture of primary cell walls based on biomechanical changes induced by substrate-specific endoglucanases. Plant Physiol 158:1933–1943
Peaucelle A, Braybrook SA, Le Guillou L et al (2011) Pectin-induced changes in cell wall mechanics underlie organ initiation in Arabidopsis. Curr Biol 21:1720–1726
Peaucelle A, Wightman R, Höfte H (2015) The control of growth symmetry breaking in the Arabidopsis hypocotyl. Curr Biol 25:1746–1752
Refrégier G, Pelletier S, Jaillard D, Höfte H (2004) Interaction between wall deposition and cell elongation in dark-grown hypocotyl cells in Arabidopsis. Plant Physiol 135:959–968
Richmond PA, Métraux J-P, Taiz L (1980) Cell expansion patterns and directionality of wall mechanical properties in Nitella. Plant Physiol 65:211–217
Routier-Kierzkowska AL, Weber A, Kochova P et al (2012) Cellular force microscopy for in vivo measurements of plant tissue mechanics. Plant Physiol 158:1514–1522
Rayle D, Cleland R (1992) The acid growth theory of auxin-induced cell elongation is alive and well. Plant Physiol 99:1271–1274
Ryden P, Sugimoto-Shirasu K, Smith AC et al (2003) Tensile properties of Arabidopsis cell walls depend on both a xyloglucan cross-linked microfibrillar network and rhamnogalacturonan II-borate complexes. Plant Physiol 132:1033–1040
Suslov D, Verbelen J-P (2006) Cellulose orientation determines mechanical anisotropy in onion epidermis cell walls. J Exp Bot 57:2183–2192
Suslov D, Verbelen JP, Vissenberg K (2009) Onion epidermis as a new model to study the control of growth anisotropy in higher plants. J Exp Bot 60:4175–4187
Suslov D et al (2012) Cell wall mechanical properties of Arabidopsis hypocotyls in connection with growth and gravitropism: effects of xyloglucan endotransglucosylase/hydrolases, expansin-like proteins and brassinosteroids. In: 7th plant biomechanics international conference, Clermont-Ferrand, 20–24 Aug 2012
Suslov D, Ivakov A, Boron AK et al (2015) In vitro cell wall extensibility controls age-related changes in the growth rate of etiolated Arabidopsis hypocotyls. Funct Plant Biol 42:1068–1079
Taiz L (1984) Plant cell expansion: regulation of cell wall mechanical properties. Ann Rev Plant Physiol 35:585–657
Takahashi K, Hirata S, Kido N et al (2006) Wall-yielding properties of cell walls from elongating cucumber hypocotyls in relation to the action of expansin. Plant Cell Physiol 47:1520–1529
Thompson DS (2001) Extensiometric determination of the rheological properties of the epidermis of growing tomato fruit. J Exp Bot 52:1291–1301
Vandenbussche F, Suslov D, de Grauwe L et al (2011) The role of brassinosteroids in shoot gravitropism. Plant Physiol 156:1331–1336
Van Sandt V, Suslov D, Verbelen J-P et al (2007) Xyloglucan endotransglucosylase activity loosens a plant cell wall. Ann Bot 100:1467–1473
Wiedemeier AM, Judy-March JE, Hocart CH, Wasteneys GO, Williamson RE, Baskin TI (2002) Mutant alleles of Arabidopsis RADIALLY SWOLLEN 4 and 7 reduce growth anisotropy without altering the transverse orientation of cortical microtubules or cellulose microfibrils. Development 129:4821–4830
Yamamoto R (1996) Stress relaxation property of the cell wall and auxin-induced cell elongation. J Plant Res 109:75–84
Zamil MS, Yi H, Haque MA, Puri VM (2013) Characterizing microscale biological samples under tensile loading: stress-strain behavior of cell wall fragment of onion outer epidermis. Am J Bot 100:1105–1115
Zamil MS, Yi H, Puri VM (2014) Mechanical characterization of outer epidermal middle lamella of onion under tensile loading. Am J Bot 101:778–787
Zamil MS, Yi H, Puri VM (2015) The mechanical properties of plant cell walls soft material at the subcellular scale: the implications of water and of the intercellular boundaries. J Mater Sci 50:6608–6623
Acknowledgements
The authors acknowledge Saint Petersburg State University for the research grant 1.40.492.2017, RFBR for the grant No 15-04-04075, the University of Antwerp, the Research Foundation-Flanders (FWO; grants G039815 N, G065613 N, G.0.602.11.N.10 and 1.5.091.11.N.00).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Suslov, D., Vissenberg, K. (2018). Cell Wall Expansion as Viewed by the Creep Method. In: Geitmann, A., Gril, J. (eds) Plant Biomechanics. Springer, Cham. https://doi.org/10.1007/978-3-319-79099-2_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-79099-2_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-79098-5
Online ISBN: 978-3-319-79099-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)