
Abstract
The trigeminal nerve is the largest of the 12 cranial nerves. It divides into three branches and provides primarily sensory input into the central nervous system (CNS) from the head and face. The three divisions of the trigeminal nerve run in close proximity to other cranial nerves. The ophthalmic nerve (V1) and maxillary nerve (V2) are purely sensory divisions. However, the mandibular nerve (V3) contains a large sensory and a small motor root. The auriculotemporal nerve (AT), a subdivision of V3, innervates the temporomandibular joint (TMJ), while the motor branch innervates the muscles of mastication.It is very important to understand the neuroanatomy of the trigeminal nerve and its central connections to other portions of the CNS. Trigeminal drive can cause a peripheral overload into the CNS resulting in central sensitization and neuro-inflammation. An understanding of the relevant anatomy and neurology will help clinicians to better understand and treat patients with temporomandibular joint dysfunction and correlated neurological disorders. This chapter covers the neuroanatomy and physiology of the trigeminal nerve system and its central connections.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The temporomandibular joint (TMJ) is the most superior joint in the body. The TMJ moves similarly to a ball and socket joint; however, a fibrocartilage articular disc in the TMJ separates the bones of the mandible from the temporal bone of the skull. The TMJ has an upper compartment that is translational, whereas the lower compartment is rotational.
Branches of the mandibular nerve innervate the TMJ. While a person is speaking, or chewing, a mal-relationship within this joint can cause afferent nociceptive and proprioceptive signals to be sent through the trigeminal network system into the central nervous system (CNS). Sharp neurologic pain is seen clinically when (1) a patient has pain on opening or closing the TMJ or (2) when the joint makes a clicking sound, causing a disc displacement with reduction. These signals can also be aberrant subthreshold, without the generation of pain.
It is very important to understand the neuroanatomy of the trigeminal nerve and its central connections to other portions of the CNS. Trigeminal drive can cause a peripheral overload into the CNS causing central sensitization and neuro-inflammation, which can evoke systemic effects in patients such as neurological disorders. Oral orthotics suppress certain neurological conditions by decompressing irritated afferent fibers of the auriculotemporal nerve in the TMJ. The reducing aberrant excitatory input to the reticular formation (RF), cerebellum, and thalamus will cause a reduction or suppression of neurologic symptoms.
1.1 Neuroanatomy of the Temporomandibular Joint
The temporomandibular joint (TMJ) is where the mandibular condyle and the temporal bone of the cranium articulate. The TMJ has two distinct compartments, created by the articular disc. The lower compartment of the TMJ is a hinge-type movement (relationship of the mandibular condyle to the articular disc). The upper compartment is a translational-type movement of the disc (formed by the gliding of the disc against the surface of the glenoid fossa). The TMJ is innervated by three branches of the mandibular branch of the trigeminal nerve. The auriculotemporal (AT) nerve provides sensory fibers to the surrounding region of the TMJ from the posterior extending medially and laterally. The masseteric nerve supplies sensory fibers to the capsule on the anteromedial region, while the posterior deep temporal nerve supplies the anterior lateral [1]. The neuroanatomy of the trigeminal nerve will be discussed in detail in this chapter.
1.1.1 Temporomandibular Joint and Trigeminal Nerve Connections
The three divisions of the trigeminal nerve run in close proximity to other cranial nerves. Any sensory dysfunction in the head and neck may be a symptom in patients with neurological dysfunction and movement disorders. The trigeminal spinal nucleus is located at all levels of the brainstem down to the level of C-3, and the central pathways connect this nucleus with the ascending up to the thalamus and sensory cortices [2]. Therefore, abnormalities or injuries of the TMJ can often accompany and cause neurologic syndromes in the central nervous system. An understanding of the relevant anatomy and neurology will help clinicians to better understand and treat patients with temporomandibular joint dysfunction and correlated neurological disorders. This chapter covers trigeminal system anatomy and physiology of the trigeminal nerve system and how the TMJ may affect the trigeminal nerve and its central connections.
1.2 Anatomy and Physiology of the Trigeminal Nerve and Its Central Connections
The trigeminal nerve exits the pons and travels anterior to the Gasserian ganglion (GG). The Gasserian ganglion is anatomically similar to spinal root ganglia [3,4,5]. The site of formation of the three divisions is separate but runs closely together within the ganglion. The GG contains the cell bodies of all the trigeminal sensory neurons, whereas the motor root does not enter the ganglion.
Somatic sensory impulses converge to the GG from the deep and superficial structures of the head and face via three major divisions (ophthalmic division, maxillary division, mandibular division) of the trigeminal nerve [2]. These three divisions represent an afferent system, which will be described anatomically, from the GG to the peripheral branches and their connections in the periphery. The mandibular nerve is the only division of the trigeminal nerve that contains a motor root in the trigeminal system. The trigeminal nerve also contains some peripheral branches of sympathetic and parasympathetic fibers that supply the salivary, sweat, or other glands of the face, eyes, and mouth [6].
1.3 Ophthalmic Nerve
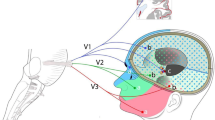
The ophthalmic nerve (V1) is the first division of the trigeminal nerve and is purely sensor. It is the smallest of the three divisions and supplies sensation to the forehead, eyeball, lacrimal gland, and lacrimal sac. It also supplies the upper eyelids, the frontal sinuses, and the side of the nose. As V1 arises from the Gasserian ganglion [7], it immediately enters the cavernous sinus inferiorly, where it lies below the trochlear nerve [8]. V1 gives off connections within the cavernous sinus to the oculomotor, trochlear, and abducens nerves, thereby supplying sensation to the muscles innervated by these nerves [9]. At this level, it also gives off recurrent branches that cross, adhering to the trochlear nerve, and are distributed to the tentorium cerebelli and dura. As V1 leaves the cavernous sinus, it divides into three branches, which are the lacrimal nerve, frontal nerve, and nasociliary nerve. All three branches enter the orbit through the superior orbital fissure [10].
Branches of V1 are as follows (Fig. 1.1): (a) tentorial nerve innervates the dura of the anterior fossa, tentorium cerebelli, falx cerebri, and the superior sagittal sinuses. In animal studies, the ophthalmic division has minor contribution from the maxillary division and innervates the vessels in the circle of Willis [11, 12]. (b) Frontal nerve enters the orbit from above the superior rectus and the levator palpebrae superioris muscles. Around the middle of the orbit, the supratrochlear nerve branches off, and at this point it becomes the supraorbital nerve. The supraorbital nerve exits the orbit through the supraorbital notch (with the supraorbital artery) and proceeds superiorly giving minor branches to the frontal sinus and eyelids [13]. It innervates the skin near the midline of the forehead, the upper eyelid (skin and conjunctiva), frontal sinuses, and the skin on the side of the nose. (c) The lacrimal nerve enters the orbit through the superior orbital fissure and divides into two divisions. The superior division provides sensory innervation to the lacrimal gland, the conjunctiva, and the upper eyelid. The lower division of the lacrimal nerve anastomoses with the maxillary division. (d) Nasociliary nerve is the sensory innervation to the eye and the nose as it passes through the cranial cavity and enters the orbit through the annulus of Zinn (common tendinous ring) between the two divisions of the oculomotor nerve and courses under the superior rectus muscle. It exits the orbit via the anterior ethmoidal foramen and reenters the cranial vault. It enters the nasal cavity through the nasal fissure, which is located on the side of the crista galli. At this point, it divides into three terminal branches. Two of the branches terminate in the anterior nasal cavity (medial and lateral internal nasal nerves), and the branch continues anterior to the end of the nose (external nasal nerve) [13].

Ophthalmic nerve. A diagram of the branches, pathways, and areas of innervation of the ophthalmic division of the trigeminal nerve. Supplied by Ms. Rachel-Marie Demerjian
1.4 Maxillary Nerve
The maxillary nerve (V2) of the trigeminal nerve is also a pure sensory division. It innervates the skin of the midface (small part of the temporal area, lower eyelid, cheek, upper lip, side of the nose, part of the mucous membrane of the nose, nasopharynx, maxillary teeth, maxillary sinus, soft palate, tonsil, and palate of the mouth).
V2 arises from the GG and enters the cavernous sinus beneath V1. The middle meningeal nerve is an intracranial branch of V2 that innervates the dura mater of the middle cranial fossa. V2 passes through the foramen rotundum (in the medial aspect of the greater wing of the sphenoid bone) [14] to the pterygopalatine fossa [15]. At this point, V2 gives off three branches: (A) pterygopalatine nerves, which enter the pterygopalatine ganglion and then divides into branches that supply portions of the nasal cavity and nasopharynx (posterior superior nasal, posterior inferior nasal) and hard and soft palate (nasopalatine nerve, greater palatine nerve, lesser palatine nerve, pharyngeal branch (Fig. 1.2). The pterygopalatine ganglion receives parasympathetic impulses from the genticulate ganglion of the facial nerve and sympathetic impulses from the superior cervical ganglion via the deep petrosal nerve (internal carotid artery). (B) Posterior superior alveolar nerves pass through the pterygomaxillary fissure to provide sensation to the maxillary gingiva and molar teeth; and (C) zygomatic nerve passes through the inferior orbital fissure into the orbit. When in the orbit, zygomatic nerve has connections with the lacrimal nerve. This zygomatic nerve then divides into two branches: (a) zygomaticofacial nerve that passes through the zygomatic foramen innervates the skin of the face, over the zygomatic bone, and (b) zygomaticotemporal nerve innervates the skin on the temporal side of the zygomatic bone.

Maxillary nerve. The maxillary nerve supplies innervation to the midface. This figure demonstrates the pathways, branches, and arears of innervation such as the teeth. Supplied by Ms. Rachel-Marie Demerjian
As the V2 nerve passes through the inferior orbital fissure and becomes the infraorbital nerve as it enters the orbit. The infraorbital nerve is the terminal branch of the V2. It passes through the orbital floor into the infraorbital canal. While in the infraorbital canal, it innervates the maxillary bicuspids and anterior teeth. The infraorbital nerve innervates the skin of the face from an area below the eye to the upper lip adjacent to the nose (Figs. 1.2). [16,17,18].
1.5 Mandibular Nerve
The mandibular nerve (V3) contains a large sensory and small motor root. The sensory root from the Gasserian ganglion passes inferiorly through the motor root. The motor root supplies muscles that derive from the first branchial arch of mammalian embryos [6]. The two roots are in close proximity in the middle cranial fossa and travel through the foramen ovale to the infratemporal fossa. There they become one trunk. The foramen ovale sits in the posteromedial aspect of the greater wing of the sphenoid bone [14]. At the infratemporal fossa, V3 gives off two branches: (a) nervus spinosus nerve which accompanies the middle meningeal artery through the foramen spinosum and innervates the dura mater on the temporal side of the cranium [19] and (b) medial pterygoid nerve, which passes through or by the otic ganglion to the tensor veli tympani and tensor veli palatini muscles [6]. V3 then divides into anterior (primarily motor) and posterior (primarily sensory) divisions.
1.6 Otic Ganglion
The otic ganglion is located medial to V3, as it exits the skull through the foramen ovale from the skull. [20]. The preganglionic parasympathetic fibers arise from the glossopharyngeal nerve (via tympanic plexus and the lesser petrosal nerve) and the facial nerve (via chorda tympani nerve) [20, 21]. Preganglionic sympathetic fibers from the superior cervical ganglion via the middle meningeal artery pass through the otic ganglion. Postganglionic fibers exit the otic ganglion via the auriculotemporal nerve to innervate the parotid gland [22].
1.6.1 Anterior Division of V3
The anterior division of V3, being mostly motor root, innervates the muscles of mastication. It travels downward and forward, medial to the lateral pterygoid muscle, and separates into three branches: (a) lateral pterygoid nerve (innervates lateral pterygoid muscle); (b) masseteric nerve (innervates master muscle) passes superior border to the lateral pterygoid muscle and over the mandibular notch and supplies a fiber to the retrodiscal tissue of the temporomandibular joint; (c) temporal nerve (anterior temporal nerve and posterior temporal nerve) passes to the temporalis muscle; and (d) the buccinator nerve passes between the two heads of the lateral pterygoid muscle to reach the masseter and buccinator muscle [2] (Fig. 1.3).

Mandibular nerve. The mandibular nerve has sensory and motor branches. This figure demonstrates the pathways and branches of the mandibular nerve. Supplied by Ms. Rachel-Marie Demerjian
1.6.2 Posterior Division of V3
The posterior division of V3 has three major subdivisions, all with dual functions [6]: (a) the auriculotemporal (AT) nerve can have 1–5 root variations and can start from the mandibular nerve or the inferior alveolar nerve. When a bifurcation of the inferior alveolar nerve occurs, the AT nerve roots could start from the anterior or posterior rami of the inferior alveolar nerve trunk. Regardless of the number of primary roots, the AT nerve trunk is finally formed by the merging of the roots. Over 80% of the dissections, the medial meningeal artery passed between the first and second branches of the auriculotemporal nerve, which are then combined into a single nerve [23].
This nerve supplies the external auditory meatus (ear canal), auricle (ear), external part of the tympanic membrane (ear drum), temporal skin, and parotid gland (secretomotor fibers) to the sides of the head. Several articular branches are also carried with the nerve, which supply blood to the temporomandibular joints [6, 24, 25]. The auriculotemporal nerve is the primary nerve to supply the TMJ, together with the masseteric nerve branches and the deep temporal. It is the primary sensory supply to the TMJ with contributions also from the masseteric (anterior lateral) and deep temporal nerves (anterior medial) [Fig. 1.4]. The auriculotemporal nerve has the majority of nerve endings that are located in the vascular connective tissue, on the posterior aspect of the disc called the retrodiscal tissue (bilaminar zone) [26, 27]. There are three types of nerve receptors in the TMJ: Ruffini corpuscles (unencapsulated), Pacinian corpuscles (encapsulated), and free nerve endings. The function of these receptors is perception of pain and to relay the position of the joint. There are no receptors on the articular surfaces of the disc [1, 26]. (b) The lingual nerve supplies innervation to the mandibular gingiva and anterior portion of the tongue. The chorda tympani of the facial nerve joins the lingual nerve, to supply taste fibers for the anterior two-thirds of the tongue. The lingual nerve sends parasympathetic fibers to the submandibular ganglion; then postganglionic fibers exit the submandibular ganglion and return to the lingual nerve before entering the salivary glands. (c) The inferior alveolar nerve gives off branch, mylohyoid nerve (a motor branch that innervates the mylohyoid muscle and the anterior belly of the digastric muscle) before entering the mandible through the mandibular foramen. In the mandible, it sends branches to the inferior dental plexus, which innervate the mandibular teeth and gingiva. The lingual never exits the mandible through the mental canal and becomes the mental nerve and innervates the skin and mucous membrane of the lower lip and gums and the chin (Fig. 1.4).

Afferent nerve innervations of the TMJ. As the auriculotemporal nerve passes by the TMJ from the posterior, it supplies branches innervating the surrounding tissues as it wraps around the mandibular condyle from medial to lateral (yellow). The black arrow is pointing to the retrodiscal tissue. The posterior deep temporal nerve (green) supplies a branch to the anterior lateral section of the condyle and the masseteric nerve (blue) innervated the anterior medial portion. Adapted from Saleem N et al.
1.7 Trigeminal Afferent Fibers
The Gasserian ganglion contains the cell bodies of the afferent sensory trigeminal fibers, which enter the lateral pons. There they split into a short ascending and a long descending branch. The descending branch forms the trigeminal spinal tract, at the medulla oblongata, which continues medially into the trigeminal spinal nucleus. Investigations of the trigeminal spinal nucleus show that the afferent fibers of all divisions extend and diminish caudally at the level of C3 [4]. The most caudal portion of the trigeminal spinal nucleus is involved in the transmission of pain perception as it contains the fibers from V3.
As the motor root of V3 enters the pons, it is composed of axons with terminals in the muscles of mastication and oral cavity. They bypass the trigeminal motor nucleus and end in the midbrain. The cells of origin of the motor root of V3 and the mesencephalic nucleus are found throughout the mesencephalic tegmentum and rostral pontine.
1.8 Gasserian (Trigeminal, Semilunar) Ganglion
The motor root of the trigeminal nerve originates from the mesencephalic nucleus and accompanies the sensory root. The sensory and motor divisions exist the pons as two separate roots, where the motor root exits just superior to the point of entrance of the sensory root. The trigeminal nerve exits the pons and travels anterior to the Gasserian ganglion . The motor root is underneath the Gasserian ganglion in Meckel’s cave. It is surrounded by arachnoid and dura, located in the middle cranial fossa [28]. The Gasserian ganglion is similar to a spinal root ganglion [3,4,5]. Formation sites for the three divisions of the trigeminal nerve are separate. However, all three divisions of the nerve run closely together. The Gasserian ganglion contains the cell bodies of all the trigeminal sensory axons. The motor root of the trigeminal nerve does not enter the ganglion.
1.9 Trigeminal Nerve Nuclei
The trigeminal system has four nuclei: three sensory nuclei, which are the trigeminal main sensory nucleus, trigeminal mesencephalic nucleus, and the trigeminal spinal nucleus, and one motor nucleus, the trigeminal motor nucleus [9].
1.9.1 Main Sensory Nucleus
The trigeminal main sensory nucleus is located in the midpons [29]. The main sensory nucleus provides tactile and pressure sensation from the head to the brainstem. Most of its neurons project to the contralateral ventralis posteromedialis (VPM) nucleus and ascend to the thalamus [30].
1.9.2 Mesencephalic Nucleus
Mesencephalic nucleus consists of unipolar, primary afferent cells similar to sensory ganglion cells [31]. Their cell bodies remain within the central nervous system and are derived from neural crest cells. Most of these neurons within the mesencephalic nucleus are proprioceptive in function [32, 33], with receptor terminals in the muscles of mastication, which respond to stretch. Neurons of this nucleus are unipolar cells that receive proprioceptive information input to the main sensory and trigeminal motor nucleus and reticular formation and send projections to the trigeminal motor nucleus to mediate a reflex response.
1.9.3 Spinal Trigeminal Nucleus
The spinal trigeminal nucleus extends down the spinal cord as far as the second cervical root and merges with the substantia gelatinosa [29]. The spinal trigeminal nucleus is divisible into three sections: (a) the spinal trigeminal nucleus oralis, (b) the spinal trigeminal nucleus interpolaris, and (c) spinal trigeminal nucleus caudalis [31]. The ophthalmic division is located most ventrally and those from the mandibular located most dorsally. The nucleus of the spinal trigeminal nucleus is primarily involved in the transmission of pain and temperature impulses [34].
1.9.4 Trigeminal Motor Nucleus
The trigeminal motor nucleus is located medial to the main sensory nucleus at the level of the pons. It contains interneurons and the cell bodies of alpha and gamma motor/branchiomotor neurons. The motor fibers join the mandibular division of the trigeminal nerve and are distributed to the muscles (masseter, temporalis, medial pterygoid, lateral pterygoid, mylohyoid, anterior belly of the digastric, tensor veli tympani, and tensor veli palatini muscles) [9].
1.10 Spinal Trigeminal Nuclei Connections
Many neurons of the spinal trigeminal nucleus send axons into the longitudinal axon plexus, which consists of bundles of interconnected myelinated and unmyelinated axons. These bundles run through the entire length of the nucleus within the spinal tract, containing ascending and descending axons. The axons give off collateral fibers that effectively connect the different levels of the nucleus [35, 36].
The trigeminal sensory nucleus is connected to the motor nuclei (ocular, trigeminal, facial, vestibular, glossopharyngeal, vagal, and hypoglossal) in the brainstem via short neurons in the tegmentum. Ascending impulses are conducted by multi-synaptic pathways via the dorsal section of the hypothalamus to the ventromedial nuclei of the thalamus. Then the thalamus will distribute the signals to cerebral cortex. The trigeminal nucleus also has collateral axons that send impulses to the superior colliculus, cerebellar cortex, and deep cerebellar nuclei [37, 38].
The ascending pathway of the spinal trigeminal nucleus and main sensory nucleus forms the trigeminothalamic tract also known as the ventral trigeminal tract. This tract transmits impulses of pain and temperature, arising from all levels of the spinal trigeminal nucleus.
Incoming pain impulses from the face are relayed to the thalamus via the trigeminothalamic tract. The trigeminal spinal nucleus and the spinal trigeminal tract extend to the level of the second cervical vertebrae. The pain impulses from the neck enter the spinal cord at the level of C1 and C2. Second-order neurons in the spinal trigeminal nucleus relay the pain impulses to the thalamus via the lateral spinothalamic tract. Investigators have followed the overlapping zones of the descending trigeminal system and cervical root fibers of C1, C2, and C3 to cervical levels down to the level of C6 [39]. A microelectrode study showed that there were some convergent fiber neurons from the trigeminal and cervical roots. As stimuli were delivered to the peripheral branches of the trigeminal and cervical roots, the same neurons were triggered. Kerr stated that stimulation of the dorsal root of the first dorsal cervical segment produces referred pain to the back of the eye, to the forehead, and, occasionally, to the vertex; rarely is pain evoked in the back of the head (ref). Therefore, he estimated that 25–30% of the neurons responded to the stimuli from either the face or neck regions [9].
1.11 Reticular Formation
The reticular formation (RF) forms the central core of the brainstem and is described as a diffuse structure that has no distinct cytoarchitectural boundaries, which houses over 100 identified nuclei. The RF forms the central gray matter of the midbrain, pons, and medulla. The spinal cord contains an analogue of the RF throughout its entire length, known as the intermediate zone of gray matter. The RF contains fibers oriented in all planes and appears as an interlacing structure that fills the area among various ascending and descending pathways (cranial nerve nuclei and gray matter) in histological sections [40]. It is continually receiving information of activity occurring in the nervous system and responding by influencing the skeletal muscle activity (motor), sensations (somatic and visceral), activity of the autonomic nervous system, endocrine function, reciprocal hypothalamus connections, consciousness levels, and biological rhythm [2].
RF neurons have elaborate dendritic trees, which branch out and radiate in all directions. They give rise to an ascending or descending axons and numerous collateral branches. The dendrites are in perpendicular orientation to the long axis of the brainstem. The axons, radiating dendritic trees, and collateral branches facilitate the ability of the neurons to collect and transmit information to and from various nuclei (e.g., oculomotor, trigeminal, facial, glossopharyngeal nuclei) via ascending and descending fibers along the brainstem. The RF integrates incoming information and then influences outgoing information to nerve cell activity at all levels of the CNS [41, 42].
The RF is divided into four longitudinal zones (columns) on the basis of their mediolateral locations in the brainstem: median zone (midline raphe), paramedian zone (located lateral to the midline), medial zone, and lateral zone [2, 43].
The medial zone (motor zone, efferent zone) consists of large neurons. The medial zone nuclei contain neurons with bifurcate axons that give rise to long ascending and descending branches, each with collateral branches. The ascending fibers course in the central tegmental tract to terminate in the hypothalamus (controls the autonomic nervous system) and thalamus (function in arousal). The descending fibers extend inferiorly to the spinal cord by joining the pontine and medullary reticulospinal tracts. The pontine and rostral medullary RF of the medial zone give rise to the reticulobulbar tract (motor control and modulation of sensory information transmission), which terminates in the motor and sensory cranial nerve nuclei [2].
The lateral zone (sensory zone, afferent zone) consists of small-sized interneurons, which are the most numerous type of cells in the RF . These small-sized interneurons contain short ascending and descending branches that are localized in the medial zone. Some interneurons terminate in the cranial nerve motor nuclei. It receives sensory information from the cerebrum, cranial nerves, cerebellum, and spinal cord via collateral branches of various somatosensory (touch, pressure, pain, temperature, and general proprioception) pathways. The lateral zone receives sensory information, integrates, and then relays the information to the medial zones [2, 44, 45].
1.12 Thalamus
The thalamus , a large sensory nucleus, is found between the cerebral cortex and the brainstem. It is located between the corpus striatum from above and the midbrain and hypothalamus from below. It is completely covered by the cerebral hemispheres. The posterior ventral nucleus of the thalamus sends impulses to the cerebral cortex and has connections to the basal ganglia. These projections to the cerebral cortex lead to the paracentral and postcentral gyri. It relays all sensory information from the external environment, except olfaction to the cerebral hemispheres for processing [9].
1.13 Auriculotemporal Nerve Connection to Systemic Neuropathology
The neural innervation of the TMJ was discussed previously by the auriculotemporal nerve , masseteric nerve , and the posterior deep temporal nerve [1, 46]. Tissue damage or inflammation can produce an excitability of the nociceptors at the site nerve injury. This is called peripheral sensitization [46,47,48,49,50]. Prolonged excitation can cause changes in the CNS termed neural plasticity resulting in the release of neuropeptides such as calcitonin gene-related peptide (CGRP) and substance P (SP) into the synapse from the primary afferent neurons. These neuropeptides act on the macrophages, mast cells, and platelets causing inflammatory mediators (cytokines, histamine, serotonin and bradykinin). These inflammatory mediators can cause an increase in excitability while acting on the nociceptive afferent nerve endings. An increase excitability of the nociceptors causes spontaneous activity, lower threshold activation, and an increased response to subsequent stimuli. These neuropeptides are transported in the trigeminal spinal nucleus [46, 51,52,53,54]. An increase in the nociception activity can lead to increase afferent signals into the CNS causing central sensitization. Sensory neurons at all three levels of the trigeminal spinal nucleus have relay neurons to the thalamus either directly or indirectly via the reticular formation [46, 49, 53, 55, 56].
Nerve damage or injury leads to an increase activity and repair, causing neuronal regeneration and sprouting [46]. Neuromas are sensitive to chemical, mechanical, and thermal stimulation. Neuromas transmit spontaneous stimulation into the CNS [56]. This spontaneous neural activity originates from the cell body in the Gasserian ganglion. The increase of neural activation arising from the neuroma and the Gasserian ganglion results in hypersensitivity and hyperexcitability of the CNS. Prolonged tissue and neural innervation can cause changes in the CNS leading to neural plasticity [46].
The trigeminal nerve has a tonic regulator (inhibitory), called the reticular formation . The reticular formation exerts control over the sensorimotor circuits within the brainstem [57]. The specific network for locomotor and postural control in humans is transmitted via the ponto-medullary reticular formation and integrated through multisensory input at different levels within the midbrain [58].
Research has shown that stimulation along the trigeminal nerve causes a motor activity in the sternocleidomastoid and splenius muscles of the neck [58, 59]. Therefore, stimulation of the primary sensory afferent fibers of the auriculotemporal nerve traveling via the trigeminal nerve to the brainstem activates the reticular formation [60, 61]. Siegel found movements of the head and neck were caused by neural stimulation of the reticular formation on the ipsilateral side [62]. The nucleus raphe of the reticular formation gets direct input from the trigeminal spinal tract when trigeminal nerves are excited. Excessive neuronal stimulation can cause an interference with impulse conduction from the cerebral cortex via the reticular formation and produce stimulation causing involuntary movements such as balance disorders [63]. The cerebellum gets afferent impulses from the vestibular nuclei, cerebral cortex, spinal cord, reticular formation, and trigeminal nuclei via the trigeminocerebellar tract. Damage to, or a lesion within, any of these pathways primarily produces a change of muscle tone or postural reflexes [57].
Conclusion
The TMJ is a complex joint, where the relations can be affected by several ways, such as macro-trauma (direct injury of the joint) or micro-trauma (clenching or bruxism). This complex joint is the only joint in the body that has a hard end point (dentition) that dictates the relationship of the joint. Furthermore, if one of the temporomandibular joints does not function properly such as an internal derangement, it will affect the TMJ on the other side causing a compensation of the opposing TMJ. Any mis-relationship that affects either TMJ or the dental relationship can start a chain of events due to the afferent signal conducted through the trigeminal network system into the CNS. Oral orthotics suppresses certain neurological conditions by reducing the aberrant excitatory input into the RF. Therefore, signals to the thalamus and cerebellum are reduced. The type of orthotic or splint is not relevant; however the proper mechanical force being put on TMJ and relieving the excitatory input into the CNS is most relevant.
Abbreviations
- AT:
-
Auriculotemporal
- CNS:
-
Central nervous system
- RF:
-
Reticular formation
- TMJ:
-
Temporomandibular joint
- VPM:
-
Ventralis posteromedialis
References
Saleem N. Loney E, Chuah JH. The auriculotemporal nerve—where is it, what does it do and why should we know about it? Eur Soc Radiol. ECR 2014/C-2122. doi: 10.1594/ecr2014/C-2122.
Patestas MA, Gartner LP. A textbook of neuroanatomy. Malden: Blackwell; 2006.
Dixon AD. Fine structure of nerve-cell bodies and satellite cells in the trigeminal ganglion. J Dent Res. 1963;42:990–9.
Kerr FWL. The divisional organization of afferent fibres of the trigeminal nerve. Brain. 1963;86:721–32.
Moses HL. Comparative fine structure of the trigeminal ganglion, including autopsy studies. J Neurosurg. 1967;26:112–26.
Romanes GJ. The peripheral nervous system: trigeminal nerve. In: Romanes GJ, editor. Cunningham’s textbook of anatomy. Ed 12 ed. Oxford: Oxford University Press; 1981. p. 748–56.
Hardy DG, Rhoton AL Jr. Microsurgical relationships of the superior cerebellar artery and the trigeminal nerve. J Neurosurg. 1978;49:669–78.
Umansky F, Nathan H. The lateral wall of the cavernous sinus: with special reference to the nerves related to it. J Neurosurg. 1982;56:228–34.
Lui GT. The trigeminal nerve and its central connections (Chapter 25). In: Miller NR, Newman NJ, editors. Walsh & Hoyt’s Clinical neuro-ophthalmology. 6th ed; 2005. p. 1233–74.
Harris FS, Rhoton AL. Anatomy of the cavernous sinus: a microsurgical study. J Neurosurg. 1976;45(2):169–80.
Ruskell GL. Ocular fibers of the maxillary nerves in monkeys. J Anat. 1974;118:195–203.
Simons T, Ruskell GL. Distribution and termination of trigeminal nerves to the cerebral arteries in monkeys. J Anat. 1988;159:57–71.
Whitnall SE. The anatomy of the human orbit and accessory organ of vision. 2nd ed. New York: Oxford University Press; 1932. p. 347–72.
Ginsberg LE, Pruett SW, Chen MYM, et al. Skull-base foramina of the middle cranial fossa: reassessment of normal variation with high-resolution CT. Am J Neuroradiol. 1994;15:283–91.
Moiseiwitsch J, Irvine T. Clinical significance of the length of the pterygopalat ine fissue in dental anesthesia. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2001;92:325–8.
Marfurt CF, Kingsley RE, Echtenkamp SE. Sensory and sympathetic innervation of the mammalian cornea: a retrograde tracing study. Invest Ophthalmol Vis Sci. 1989;30:461–72.
Morgan C, Jannetta PJ, deGroat WC. Organization of corneal afferent axons in the trigeminal nerve root entry zone in the cat. Exp Brain Res. 1987;68:411–6.
Ruskell GL, Simons T. Trigeminal nerve pathways to the cerebral arteries in monkeys. J Anat. 1987;155:23–37.
Penfield W, McNaughton F. Dural headache and innervation of the dura mater. Arch Neurol Psychiatry. 1940;44:43–75.
Kuntz A. Anatomic and physiologic properties of cutaneo-visceral vasomotr reflex ARCS. J Neurophysiol. 1945;8(6):421–9.
Kuchiiwa S, Kuchiiwa T, Nonaka S, Nakagawa S. Facial nerve parasympathetic preganglionic afferents to the accessory otic ganglia by way of the chorda tympani nerve in the cat. Anat Embryol. 1998;197:377–82.
Kuntz A, Richins CA. Components and distribution of the nerves of the parotid and submandibular glands. J Comp Neurol. 1946;85(1):21–32.
Komarnitki I, Andrzejczak-Sobocińska A, Tomczyk J, Deszczyńska K, Ciszek B. Clinical anatomy of the auriculotemporal nerve in the area of the infratemporal fossa. Folia Morphol. 2012;71(3):187–93.
Schmalfuss IM, Tart RP, Mukherji S, et al. Perineural tumor spread along the auriculotemporal nerve. Am J Neuroradiol. 2002;23:303–11.
Schmidt BL, Pogrel MA, Necoechea M, et al. The distribution of the auriculo-temporal nerve around the temporomandibular joint. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 1998;86:165–8.
Isberg A. Temporomandibular joint dysfunction: a practitioner’s guide (Chapter 2). Pain from the temporomandibular joint region. London: Isis Medical Media; 2001. p. 13–4.
Klineberg I, Lillie J. Regional nerve block of the temporomandibular joint capsule: a technique for clinical research and differential diagnosis. J Dent Res. 1980;59:111930–5. https://doi.org/10.1177/00220345800590110101.
Jannetta PJ. Gross (mesoscopic) description of the human trigeminal ganglion. J Neurosurg. 1967;26:109–15.
Afshar F, Dykes E, Watkins ES. Three-dimensional stereotatic anatomy of the human trigeminal nerve nuclear complex. Appl Neurophysiol. 1983;46:1470153.
Smith RL. Axonal projections and connections of the principal sensory trigeminal nucleus in the monkey. J Comp Neurol. 1975;163:347–76.
Darian-Smith I. The trigeminal system. In: Iggo A, editor. Handbook of sensory physiology, vol. 2. Berlin: Springer; 1973. p. 271–314.
Corbin KB, Harrison F. Function of mesencephalic root of the fifth cranial nerve. J Neurophysiol. 1940;3(5):423–35.
Jerge CR. Organization and function of the trigeminal mesencephalic nucleus. J Neurophysiol. 1963;26(3):379–92.
Dallel R, Raboisson P, Woda A. The rostal part of the trigeminal sensory complex is involved in orofacial nociception. Brain Res. 1988;448(1):7–19.
Gobel S, Purvis MB. Anatomical studies of the organization of the spinal V nucleus: the deep bundles and the spinal V tract. Brain Res. 1972;48:27–44.
Ikeda M, Tanami T, Matsushita M. Ascending and descending internuclear connections of the trigeminal sensory nuclei in the cat. A study with the retrograde and anterograde horseradish peroxidase technique. Neuroscience. 1984;12(4):1243–60.
Silverman JD, Kruger L. Projections of the rat trigeminal sensory nuclear complex demonstrated by multiple fluorescent dye retrograde transport. Brain Res. 1985;361(1-2):383–8.
Steindler DA. Trigeminocerebellar, trigemintectal, and trigeminothalamic projections: a double retrograde axonal tracing study in the mouse. J Comp Neurol. 1985;237(2):155–75.
Taren JA, Kahn EA. Anatomic pathways related to pain in the face and neck. J Neurosurg. 1962;19(2):116–21.
Olszewski J, Baxter D. Cytoarchitecture of the human brainstem. Neuroepidemiology. 2014;42(2):69–138. (3rd, revised and extended edition).
Jenkinsson N, Nandi D, Muthusamy K, Ray NJ, Gregory R, Stein JF, Aziz TZ. Anatomy, physiology, and pathophysiology of the pedunculopontine nucleus. Mov Disord. 2009;24(3):319–28.
Scheibel ME, Scheibel AB. Structural substrates for integrative patterns in the brainstem reticular core. In: Jasper HH, Proctor LD, Knighton RS, Noshay WC, Costello RT, editors. Reticular formation of the brain. Boston: Little & Brown; 1958. p. 31–55.
Nieuwenhuys R, Voogd J, Van Huijzen C. The reticular formation and the monoaminergic and cholinergic cell groups. In: Nieuwenhuys R, Voogd J, van Huijzen C, editors. The human central nervous system. Berlin: Springer; 2008. p. 889–916.
Holstege G. Descending motor pathways and spinal motor system: limbic and non-limbic components. Prog Brain Res. 1991;87:307–421.
Holstege G, Mouton LJ. Central nervous system control of micturition. Int Rev Neurobiol. 2003;56:123–45.
Sessle BJ. Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive transmission and neuroplasticity, and their clinical correlates. Crit Rev Oral Biol Med. 2000;11:57–91.
Hargreaves KM. Neurochemical factors in injury and inflammation of orofacial tissue. In: Lund JP, Lavigne GJ, Dunbar R, Sessle BJ, editors. Orofaical pian. Basic science to clinical management. Chicago: Quintessence; 2001. p. 59–66.
Lund JP, Sessle BL. Neurophysiological mechanisms related to chronic pain disorders of the temporomandibular joint and masticatory muscles. In: Zarb G, Carlsson G, Sessle BJ, Mohl N, editors. Temporomandibular joint and masticatory muscle disorders. Copenhagen: Munksgaard; 1994. p. 188–207.
Sessle BJ. The Neural basis of temporomandibular joint and masticatory muscles pain. J Orofac Pain. 1999;13:238–45.
Svensson P, Sessle BJ. Orofacial pain. In: Miles TS, Nauntofte B, Svensson P, editors. Clinical oral physiology. Copenhagen: Quintessence; 2004. p. 930139.
Cairns BE. Nociceptors in the orofacial region—TMJ and masseter muscle. In: Schmidt RF, Willis WD, editors. Encyclodepia of pain. Heidelberg: Springer; 2007. p. 1410–27. https://doi.org/10.1007/978-3-540-29805-2_2793.
Capra NF, Dessem D. Central connections of the trigeminal primary afferent neurons: Topographical and functional considerations. Crit Rev Oral Biol Med. 1992;4:1–52.
Dunbar R, Sessle BJ, Storey AT. The neural basis of oral and facial function. New York: Plenum; 1978.
Shigenaga Y, Sera M, Nishimori T. The central projections of masticatory afferent fibers to the trigeminal sensory nuclear complex and upper cervical spinal cord. J Comp Neurol. 1988;268:489–507.
Craig AD, Dostrovsky JO. Processing of nociceptive information at supraspinal levels. In: Yaksh TL, Lynch III C, Zapol WM, editors. Anesthesia: biologic foundations. Philadelphia: Lippincott-Raven; 1997. p. 625–42.
Hannan AG, Sessle BJ. Temporomandibular neurosurgery and neuromuscular physiology. In: Zarb G, Carlsson G, Sessle BJ, Mohl N, editors. Temporomandibular joint and masticatory muscle disorders. Copenhagen: Munksgaard; 1994. p. 67–100.
Brodal PR. The central nervous system: structure and function. 3rd ed. New York: Oxford University Press; 2004.
Jahn K, Zwergal A. Imaging supraspinal locomotor control in balance disorders. Restor Neurol Neurosci. 2010;28:105–14.
Tecco S, Tetè S, D’Attilio M, Perillo L, Festa F. Surface electromyographic patterns of masticatory, neck, and trunk muscles in temporomandibular joint dysfunction patients undergoing anterior repositioning splint therapy. Eur J Orthod. 2008;30:592–7.
Mainero C, Zhang WT, Kumar A, Rosen BR, Sorensen AG. Mapping the spinal and supraspinal pathways of dynamic mechanical allodynia in the human trigeminal system using cardiac-gated fMRI. Neuroimaging. 2007;35:1201–10.
Pinto ML, Olyntho-Tokunaga HH, Souccar C, Schoorlemmer GH, Lapa Rde C. The interstitial system of the trigeminal spinal tract projects to the red nucleus in mice. Somatosens Mot Res. 2007;24:221–5.
Siegel JM, Tomaszewski KS. Behavioral organization of reticular formation: studies in the unrestrained cat. I. Cells related to axial, limb, eye, and other movements. J Neurophysiol. 1983;50:696–716.
Buisseret-Delmas C, Compoint C, Delfini C, Buisseret P. Organization of reciprocal connections between trigeminal and vestibular nuclei in the rat. J Comp Neurol. 1999;409(1):153–68.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Demerjian, G.G., Barkhordarian, A., Chiappelli, F. (2018). Neuroanatomy of the Trigeminal Nerve and Proximal Innervation of the TMJ. In: Demerjian, G., Barkhordarian, A., Chiappelli, F. (eds) Temporomandibular Joint and Airway Disorders. Springer, Cham. https://doi.org/10.1007/978-3-319-76367-5_1
Download citation
DOI: https://doi.org/10.1007/978-3-319-76367-5_1
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-76365-1
Online ISBN: 978-3-319-76367-5
eBook Packages: MedicineMedicine (R0)