
Abstract
Background and Significance of the topic: This chapter contributes to the documentation of novel network-based resilience concepts to socio-ecological systems. Although the resilience concept has been studied in depth in ecological systems, it surely has relevance outside this area and in recent years has been a main domain of study for socioeconomic systems. This chapter provides an overview of the application of resilience concepts in ecology, with a particular focus on the application of two methods developed using ecological network analysis. Methodology: The first method uses information-theory based network analysis to ascertain the trade-off between efficiency and redundancy in networks (in terms of the structure and flows). The second method uses an energy-flow based method to assess keystoneness and the direct and indirect relations in the networks. Application/Relevance to systems analysis: Earlier work using information-theory based network analysis has shown that ecological systems display a robust balance between efficiency and redundancy in networks (in terms of the structure and flows) thereby bestowing them with robust and resilient features. Results indicate that a dam ecosystem in southwest China falls just short of the optimum but suffers substantial loss of robustness when the phytoplankton community is perturbed. Application to a virtual water network shows the system is not near the robustness peak. Using the energy-flow based method, a South African estuary showed alteration of the keystone species depending on the seasonality; a land use change model of Beijing showed a decrease in mutualism due to urban expansion. Policy and/or practice implications: The case studies presented illustrate the application of ecological network analysis. Positive and negative relations between sectors of ecosystems or economic systems highlight the influence of various species and economies on one another, resulting in a comprehensive picture of relations, impacts and therefore management options to achieve balance between sectors. Discussion and conclusion: Overall, networks provided a useful model to illustrate system resilience measures, and other system analysis methods of direct and indirect impacts of system components on each other.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

1 Introduction
In recent decades, resilience has become an important concept as ecosystems and socioeconomic systems are adversely impacted following chronic and acute resource exploitation stemming from economic policy based on growth rather than on environmental sustainability. Early warnings on the consequences of unlimited economic growth were already issued in the early 1970s by the Club of Rome (Meadows et al. 1972) on the limits that the Earth sets in the sustenance of an exponentially growing human population and its resource demands. These limits are tracked through measures such as ecological footprint in terms of human resource use, the rate of which has surpassed that of their renewal in the 1980s (Wackernagel et al. 2002). Recent investigations have looked at how human society might flourish within these biophysical limits by following general patterns and principles of ecological systems (Jørgensen et al. 2015).
The continued growth of the human population depends on the provision of many ecosystem services, mostly at no cost, but nevertheless vital for the survival of humankind. These include, for instance, pollination, availability of water and its purification, climate regulation, nutrient cycling, as well as direct provision of water, air, food, and building materials (MEA 2005). As such, continuation of ecosystem services is dependent on the sustained functioning of the ecosystems and the continued existence of its habitats and species. In this context, it is highly important to have a means to identify the space within which ecosystems function. How successful ecosystems are in remaining in this space depends on their degree of resilience, which is therefore an important trait to define.
Resilience of systems relies on the interrelationship of processes and interactions between groups, and is therefore, by definition, a system-level concept. The focus of this chapter is to review definitions of resilience on the systems-level and its related concepts, including a brief historical background over the past decades. Within this framework, we discuss various methodologies to analyse resilience, with a more detailed account of system analyses methodologies of networks for different types of systems (e.g., ecological, economic, social). We furthermore present several examples of their application to ecological and Socioeconomic systems, and discuss gaps in methodology and drawbacks of current methods, as well as future research and applications.
2 Resilience Concepts (Ecological and Socioeconomic)
Folke (2006) illustrates several concepts of resilience and portrays each as a stage in the development towards a more comprehensive understanding of the term within the context of large systems. Starting as a concept in ecology during the 1970s, resilience encompassed mainly the stability of smaller systems with limited players. This early engineering resilience (Folke 2006) mainly focussed on the return to a certain stable and constant state following a disturbance. Early work in ecology has intensely studied the latter, and used the term resilience and stability interchangeable. Beginning with population studies, often of simple predator-prey systems oscillating through cycles of high and low prey and predator abundances in a typical Lotka-Volterra (LV) fashion, stability was investigated in terms of maintaining the interplay between predators and preys by avoiding extinctions of either. Such studies gained wide recognition especially among mathematical ecologists, developing further on including important feedbacks with the environment, multiple food sources and predators (e.g., Chen and Cohen 2001). Later on, such studies were expanded towards communities and food webs, where stability has often been interpreted as the ability to return to an equilibrium starting point after impacts on the number of species, function and population sizes in the systems investigated (Moore and de Ruiter 2012). When such systems are viewed as interaction matrices between the components of the system, the interaction values at equilibrium can be disturbed and their return to equilibrium calculated. Within the LV framework, systems of equations representing predators and prey were established to investigate this return to equilibrium; namely, if the eigenvalues of its Jacobian matrix are negative for its real parts, then they describe the weakening of the disturbance over time which then brings the system back to equilibrium. Such return times to equilibrium were for instance investigated for three and four species systems, and it was apparent that return to equilibrium was faster in highly productive systems compared to those featuring lower productivity (Moore et al. 1993).
Later research was seeking to connect an ecosystem’s resilience and stability to its biodiversity, thereby also connecting human impacts on species loss and invasive species to resilience (e.g., Chapin III et al. 2000; Dudgeon et al. 2006; May 1972). Work on the influence of species diversity on the resilience and stability of ecosystems has a rich history. In the 20th century, species diversity was believed to have a positive impact on stability (e.g. Elton 1958; MacArthur 1955), and a diversity-stability debate over the past decades has hardly changed this view, although further details on various forms of diversity, and natural variability in ecosystems has become available (e.g., McCann 2000; Naeem et al. 1999; Tilman 1999). Growing concerns about species and habitat loss, and the introduction of invasive species continuously energise the debate by giving rise not only to a myriad of theoretical studies, but also to experimental studies explicitly connecting species diversity and functional diversity to resilience, in short and long term laboratory and field experiments (e.g., Gamfeldt and Hillebrand 2008; Hughes et al. 2003; Loreau 2000; Müller et al. 2016; Tilman 1999).
Changes on the ecosystem level following species loss may be averted by a rich taxonomic diversity, if taxonomic diversity is reflected in a system’s functional diversity (McNaughton 1977). On the other hand, if only rare species fulfil certain functions, their loss may be detrimental to system function (Chapin III et al. 2000). Hampering the application of such deliberations is of course the lack of comprehensive a priori knowledge on the consequences of the removal of a taxonomic species. Assessing species’ functions and their contribution to ecosystem resilience and human livelihoods may therefore be a more effective application of conservation efforts (Dudgeon et al. 2006). Or, as Mori et al. (2013) point out, assessing responses to anthropogenic impacts by evaluating the response diversity of an ecosystem may be a better gauge of its resilience. Species that may respond differently to disturbances, besides having the same functional diversity in the ecosystem, may thus be important factors of determining an ecosystem’s resilience and its adaptive capacity (Elmqvist et al. 2003). Lower response diversity may diminish a system’s resilience, whereby the relationship between the response diversity and increasing impact is of higher importance than a certain response diversity calculated for instance at the start of an impact (Mori et al. 2013).
The resilience of ecosystems has been depicted as a ball residing in a basin (Scheffer et al. 2001), where the shape of the basin discerns the extent of resilience. A flat basin indicates low resilience, whereas a cup-shaped basin indicates higher resilience. A system with high stability would reside at the lowest point of the basin, whereas a system with high resilience may occupy various points of the basin. Using a ball and basin imagery, it is easy to envision a landscape that has multiple basins each one representing different stability regimes.
The seminal work of Holling (1973) recognised already that the concept of resilience had more to offer in addition to the concept of stability, in that it provided a way to describe persistence under conditions of disturbances and variability. The focus shifted from perceiving a certain state with little fluctuation as desirable to the ability of withstanding perturbations and persisting over time (Holling 1973). Expanding this concept to multiple equilibria allowed the exploration of a wider range of functioning and stable states. This is termed ecological or social resilience, depending on the type of system explored and opens the possibility for regime shift from one stable state to another. Disturbances such as reductions in functional and response diversities may move the ecosystem towards the edge, or outside of its basin, inducing the so-called regime shift (e.g., Folke et al. 2004). Also, the alteration of the duration and frequency of existing natural variability through human activities may reduce the resilience and induce regime shifts. Following a regime shift, ecosystems have been shown to operate in spaces of stable states, and to switch between certain alternative states following disruption of the current state (Scheffer et al. 2001). The new states are deemed stable when emergent negative feedbacks operate to maintain the system in this new state. In this manner, a hysteresis may develop such that the same shock that brought the system to the new state might not be sufficient to dislodge it back again.
Following a reduction in resilience leading to a regime shift, a reconfiguration of the original energy flow pathways to a different configuration (e.g. species composition) may occur. Classic examples are of coral reefs, switching to algae dominated reefs, following overfishing of herbivorous fish and nutrient input. In shallow lakes, a switch from a clear water and macrophyte dominated state to a turbid, phytoplankton dominated state occurs after increased anthropogenic nutrient input (Scheffer et al. 2001). Once an alternative state is reached, considerable effort is needed to create conditions that occurred long before the switch occurred, or may not be possible at all, due to hysteresis effects. In such alternative states the ecosystem may not be able to provide the same ecosystem services any longer to the same degree, on which humans depend upon.
Not only did these studies elucidate the concepts of stability and resilience, they also fostered an in depth understanding of different kinds of diversities—species, functional, trophic—and their role in the resilience and resistance of ecosystems. Nowadays information is available on specific invasive species, or on the consequences of a species lost, and this knowledge on the complex interactions and on the consequences if they disappear has dramatically increased the understanding of the functioning of real world systems, and additionally of theoretical and applied models. In addition to species, community, and ecosystem considerations, the impact of humans, directly, and on their economy and society (Chapin III et al. 2000) has moved to the center of this debate.
In this wider context, social-ecological resilience includes additional concepts such as feedbacks within a system, and across various spatial and temporal scales (Folke 2006). Such scales are interlinked in systems with a panarchic setup, cycling repeatedly through exploitation, growth, collapse, and reorganisation phases across scales (Holling 1986; Gunderson and Holling 2002). The changes a system undergoes in this arrangement are part of its resilience as it can self-organise, can stay in one state depending on the amount of disturbance, and most importantly, provides opportunity for adaptation to different influences (Carpenter et al. 2001). Resilience has been proposed as the capacity of a system to navigate all stages of the adaptive cycle (Fath et al. 2015). This model was applied to the survival of firms in a socioeconomic system, with indication of how preparedness for each stage must be cultivated in each of the other preceding stages continuously, not simply the stage immediately prior. For example, to manage the collapse phase, one should already reduce possible fault cascades during the growth phase to prevent crises from spreading throughout the system; enhance cohesive leadership during the conservation stage; identify and maintain vital functions during the collapse itself; and, learn improvisation during the reorganization stage (Fath et al. 2015). Resilience has thus developed into a concept that encompasses systems in their entirety throughout time and space, rather than being restricted to understanding the stable states which a system should return to after perturbation (Kharrazi et al. 2016).
Judging the state of resilience of a system, characterising the space within which it operates, and defining the borders of this state (ranges of variability) is thus of importance when certain ecosystem services produced by certain states are desired not to be lost. It is equally important for systems heavily influenced by anthropogenic designs, such as production ecosystems, to maintain resilience in order to ensure food security. However the continued functioning of such systems usually depends on considerable external influences that uphold its production value, including the use of fertilisers, pesticides, water, or fossil fuels. Due to this, the resilience of such a system is upheld by anthropogenic influence, and is termed ‘coerced’ resilience (Rist et al. 2014). The production systems are held in a certain basin of attraction, and therefore the state of resilience, by the anthropogenic activities, often in otherwise unstable states, and bringing forth questions on their sustainability (Rist et al. 2014). The value of resilience of production and other ecosystems has found its way into the economic valuation of ecosystems, planning land-use according to bundles of ecosystem services, rather than too few, in order to maintain resilience of the ecosystem (Admiraal et al. 2013). The optimisation of an ecosystem’s services, and thus its value, may, according to Admiraal et al. (2013), be informed by resilience theory and incorporate functional diversity.
3 Resilience and Networks
Resilience on the systems-level can be studied by depicting the system as a network in which all actors in the system are nodes, and their direct interactions are links. When links have quantifiable attributes (e.g., biomass transfer, goods or money exchanges, or interactions between people), so called weighted networks describe the system (Fath et al. 2007). Resilience in networks has been studied at the systems-level in the form of system indicators, or at the level of how indirect effects are propagated through the system. The importance of nodes (e.g. keystone species) and links (e.g. indirect propagation, redundant pathways) within the system are then quantified (see methodology and examples below and in Sect. 11.4). A related series of talks on resilience and networks, which is found here: www.fas-research.com/resilience/, may be of interest.
In the field of ecology, certain network configurations have been put forward as providing stability in networks (and therefore increasing its resilience). These include for instance the relation of weak links (low weight) to strong links (large weight) in a network, and especially when weak links are configured into large cycles a stabilising effect can be apparent (Neutel et al. 2002). Stability has also been investigated in densely connected sub-sections, or small modules of networks, and found that small predator-prey modules stabilise networks (Allesina and Pascual 2008). With this knowledge, the necessity of long cycles for stability (Neutel et al. 2002) is disputed, as well as the necessity for dominance of weak cycles since the predator-prey loops have a relatively large weight in the system (Allesina and Pascual 2008). In a more in-depth study on the role of modularity in networks, a less clear result emerged, with stability of networks depending on its modularity only under certain conditions, namely the size of the subsystems and the mean interaction strength (Grilli et al. 2016).
In addition to studies on the dynamic stability of networks calculated from link weight, loop length, and modularity, resilience measures can also be calculated for networks from its weighted-link distribution between all pairs of nodes. Thus, it is not necessarily restricted to certain types of interactions (predator-prey), or certain loop configurations. Therefore, the approach here does not follow directly from the use of ecological networks to assess subsystem stability, but rather builds on whole-system energy-flow and information-theory based ecological network analysis.
Ecological Network Analysis (ENA) is one way to analyse both the direct and indirect interactions in such networks. The network configuration that comprises a certain number of nodes and links, and often a weighted link distribution, are at the centre of the ENA to calculate descriptors (including resilience) for any type of network. Such resilience measures are based on constraints on transfers of material flow along links between a source node’s output and a recipient node’s input (ascendency based, Ulanowicz 1986, 2009), and have been applied to ecological and socioeconomic networks (e.g. Chrystal and Scharler 2014; Goerner et al. 2009; Kharrazi et al. 2013).
Resilience measures that describe the constraint of flows along links or pathways in networks are dependent on the network’s connectivity, and the respective interaction between pairs of nodes (Ulanowicz 1986). Of prime interest regarding the interactions is the degree of uniformity of the flow distribution. When the total output of a source node and the total input into a recipient node are the same, the constraint on the flow is maximal. In contrast, when a source node donates only part of its output to a recipient node, and the latter receives more input from other nodes, the interaction is much less constrained. Such flow and interaction distributions can be quantified by defining the probability of input into a node and the output from the source node. These two probabilities are the same in the example of maximal constrained flows, but different in the example of the less constrained interaction between two nodes. Therefore, whenever flows are maximally constrained, each node only has one output to a receiving node, and only one input from a source node. Such systems are constrained to such an extent that there is no resiliency left in the system in case of perturbations—there is no possibility to channel energy along any other link in the system should a particular link be lost. The receiver node thus loses its entire input. In the less constrained network, nodes are more diversely interlinked, which leads to parallel pathways in that an output from a source node can reach a recipient node by different pathways, also via other nodes, or ‘detours’. Such pathways build redundancy into a network and its resilience increase as it is has a higher chance to buffer against a link loss. Such a state is therefore more desirable compared to a maximally constrained state. On the other extreme, however, a minimally constrained state prevents efficient transfers along links, a high dependency of a particular node on many other nodes, and consequently reduced functioning and a lower resilience (Goerner et al. 2009; Ulanowicz 2009). For instance, higher trophic levels in food webs, or receivers at the end of product chains, are not well served with minimally constrained networks.
Mathematically, this constraint-induced trade-off between redundancy and efficiency can be calculated using information theory. Specifically, according to Rutledge et al. (1976) the information can be determined as the reduction of uncertainty. Using the conditional probabilities of flows based on a particular network configuration, we arrive at the following equations (Ulanowicz 2004):
where H is the total system flow diversity, Φ is the system redundancy, AMI is the system average mutual information, T ij the flow from node i to j and T.. the sum of all internal and boundary flows (referred to as Total System Throughput—TST). It has been shown (Ulanowicz 1986) that H = Φ + AMI. These quantities were used to create a metric that can assess the trade-off between the system redundancy (high number of pathways with more uniform flow) versus systems with high mutual information (articulated pathways with more asymmetric flow). This new metric was first termed “fitness for change” (Ulanowicz 2009; Ulanowicz et al. 2009), subsequently sustainability (Goerner et al. 2009), and lastly system robustness (e.g. Mukherjee et al. 2015; Kharrazi et al. 2016). Quantitatively, it was derived by multiplying the ratio of AMI/H by the Boltzmann measure of disorder (−k log(a), Ulanowicz 2009) and is here termed system robustness, designated R:
where α = AMI/H.
The robustness curve shows increasing degree of order (higher value of AMI relative to H) on the x-axis as determined by AMI/H (Fig. 11.1). Work on empirical ecosystem networks revealed that their sustainability values clustered around a narrow region near the maximum of the curve, and this region has been referred to as “The Window of Vitality”. This window describes an optimum balance between redundancy and efficiency in a network that results in the highest values of sustainability or resilience. To the contrary, sustainability is lowest for both highly and minimally constrained networks compared to intermediate constrained networks (Ulanowicz 2009; Ulanowicz et al. 2009; Goerner et al. 2009; Fig. 11.1).

Robustness (Fitness in Ulanowicz 2009) as a function of efficiency and resilience. X-axis: AMI/H. Y-axis:\(-\upalpha{ \log }\left(\upalpha \right)\). Reprinted from Ecological Modelling, with permission from Elsevier
Ecological relations are direct and indirect processes that have effects on the resilience of ecosystems, as they exert some control on the interaction between components of the system. Indirect effects in ecosystems play an important part in their functioning, as they depict the wider impact in the system of changes to specific nodes or links. Examples of indirect effects include node-node interactions, trophic cascades, apparent competition, indirect mutualism and others (Wootton 1994; Higashi and Nakajima 1995; Szyrmer and Ulanowicz 1987; Shetsov and Rael 2015; Salas and Borrett 2011; Higashi and Patten 1989). Many of the studies attribute a dominance of indirect effects in ecosystems over direct effects.
In this chapter, we explore two different categories of indirect effects, firstly the influence of keystone species (Libralato et al. 2006) on ecosystems and their resilience, specifically changes in biomass and consequently the related flows. Secondly, we explore indirect effects including direct and indirect competition and mutualism, and other interrelations effects (combinations of +, −, 0 relations) (Ulanowicz and Puccia 1990; Fath 1998).
For networks, indirect effects can be calculated as follows. First, the direct negative impacts of the consumer on the resource, and the positive impacts of the resource on the consumer are calculated from the flow matric with elements T ij representing the flow from node i to node j.
\(g_{ij} = \frac{{T_{ij} }}{{\mathop \sum \nolimits_{k} T_{kj} }}\) represents the amount of all prey (i) consumed by a predator (j), whereas \(g_{ij} = \frac{{T_{ij} }}{{\mathop \sum \nolimits_{m} T_{im} }}\) represents the portion of the prey’s production that is consumed by all predators.
The net impact is achieved by subtraction: \(q_{ij} = g_{ij} - f_{ij}\), and the resulting Q matrix represents all direct impacts between two nodes. Second, the matrix of direct and indirect impacts, or total impact between two nodes, is calculated by \(M = \sum\nolimits_{h - 1}^{\infty } {Q^{h} }\), representing all powers of Q, and therefore all pathway lengths between any two nodes (Ulanowicz and Puccia 1990).
The elements of the M matrix can then be used to calculate the keystoneness of a node (KS i ) in the network (Libralato et al. 2006) as:
where\(\varepsilon_{i} = \sqrt {\sum\nolimits_{j \ne 1}^{n} {m_{ij}^{2} } }\), not including the effect of a node on itself (i.e. m ij ) Here, m ij represents the entries of the M matrix of total impacts, and p i the contribution of a node’s biomass to the total biomass of the network:
These approaches, employing the information-theory based trade-off of redundancy and efficiency and the energy-flow based indirect relations assessment (i.e., keystoneness), are used in a number of case studies involving ecological and socio-ecological systems.
4 Case Studies
Here we present several case studies that illustrate various applications of systems analysis and the exploration of the resilience concepts. These include two ecological examples, that of Manwan Dam, China and Mdloti Estuary, South Africa, which were both investigated in terms of their resilience over a time series of data and under perturbation. Furthermore, a third example included a socioeconomic system, the Heihe River Basin, China, as a Virtual Water Network. Indirect effects, on the other hand, were investigated for an ecological system (keystone species, Mdloti Estuary) and a socioeconomic system (mutualisms and competition on the systems-level) in Beijing, China.
4.1 Ecosystem Perturbation Examples: China and South Africa
We engaged in analysing ecological and economic networks for their robustness, manipulated networks to analyse their changing robustness, and described the related values of Average Mutual Information (AMI), Total System Throughput (TST), Flow Diversity (H), Ascendency (A = AMI × TST) and Development Capacity (DC = H × TST) (Ulanowicz 1986). Two types of ecological systems included the South African Mdloti estuary and Manwan Dam, China, and the socioeconomic system representing the Virtual Water Network of the Heihe River Basin, China.
4.1.1 Manwan Dam, China
In Manwan Dam, a reservoir on the Lancang River in southwest China, we investigated the effects of perturbation on robustness indicators by comparing the original network to perturbed networks in which (1) the flow structure of the food web was changed and (2) species were removed from the system. The network representing Manwan Dam (Chen et al. 2011) featured ten compartments, 41 within-system energy flows (kJ m−2 y−1), three boundary inputs, and ten boundary losses. Flows to and from the basis of the food web, i.e., detritus and phytoplankton, were removed one by one to examine the effect on overall system robustness. As a first scenario, we removed flows to and from the primary producer and detritus compartments (Fig. 11.2). Removal of the largest flow in the network—input into phytoplankton (Flow #5)—resulted in the largest drop in the system robustness. This flow into phytoplankton is the main energy source of the system and its loss therefore substantially decreased robustness of the system. Removal of the output flow from detritus (Flow #3), flow from phytoplankton to detritus (Flow #2), input flow into detritus (Flow #1), and output flow from phytoplankton (Flow #4), resulted in a sequential increase in the system robustness compared to the unperturbed ecosystem. This may be attributed to the fact that the four flows have medium flow values, and their removal led the flow distribution in the food web to become more uneven compared to the original unperturbed network.

Robustness values for the original unperturbed Manwan Dam network (green), after the removal of the input flow into detritus (1), of the flow phytoplankton → detritus (2), of the output from detritus (3), of the output from phytoplankton (4) and of the input into phytoplankton (5)
In the next scenario, the system was isolated from its environment by removing all boundary flows (Fig. 11.3). The most obvious impact of flow removal on robustness was apparent not from a compartment removal but the boundary flow removal (1), which represents isolating the system from its environment (Fig. 11.3). Thereafter, we removed nodes from the system, as an extreme scenario of perturbation. Removal of the phytoplankton node from the system led to the loss of an important energy source to the rest of the system, which also decreased its robustness. Compared with the scenario of losing total input and output, the robustness variability is relatively low, indicating that primary production is not the only source of energy to the food web. Also, the flow distribution became less uneven by removing the phytoplankton node compared to removing all boundary flows. When we removed detritus (3) in the system, as the most connected compartment in the system, the flow distribution became more efficient and the robustness of the system decreased slightly (Fig. 11.3).

Robustness values for the original unperturbed Manwan Dam network (green), after the removal of all boundary flows (1), removal of the phytoplankton node (2), detritus node (3), zooplankton node (4) and other microalgae node (5)
4.1.2 Mdloti Estuary, South Africa
Even naturally stressed ecosystems such as estuaries show various degrees of resilience. As they undergo a constant change in the physical environment, certain species have adapted to cope with changing salinity and conductivity, water levels, turbidity and other physical factors. It is often difficult to distinguish impacts in such systems as natural and anthropogenic impacts are difficult to discern (Elliott and Quintino 2007). Scenarios of real-world impacts on estuaries concerning the base of the food web (primary producers, detritus) and fishing impacts revealed interesting results on the robustness in the South African subtropical Mdloti estuary (Mukherjee et al. 2015). For this purpose, networks representing the carbon exchanges among compartments for two different seasons [dry, wet; described in Scharler (2012)] were transferred to the Ecopath software (Christensen and Walters 2004), in which the changes to the food web were implemented. The impact of perturbations on the food web network was investigated and assessed using the information theory based indices AMI, H, and Robustness. We considered the sensitivity of the network under three scenarios: (a) autotrophic biomass increase and decrease from 10 to 99 in 10% intervals; (b) an increase in fish yield from 10 up to 99% in 10% intervals; (c) an increase and decrease of the detritus import to the system from 10 to 99% in 10% intervals. The changes in network indices AMI and H (Ulanowicz 1986) and robustness (Ulanowicz et al. 2009) were calculated.
Robustness is a state of system’s health which indicates the balance between system efficiency and redundancy (see also above). All indices indicated that robustness of the system increased with a rise in autotroph biomass (Scenario a). In the fish harvest scenario (Scenario b), robustness of the system decreased as the top predators (fish) are reduced in biomass, and consequently their throughput. In the detritus import scenario (Scenario c), the relationship of robustness and A/C show a similar pattern in the two seasonal networks (Fig. 11.4). In both phases, robustness decreases with the perturbations, and the ratio of the network indices showed a downward trend in each scenario (Fig. 11.4). System robustness increased with an increase in autotrophic biomass, decreased with larger fish harvest, and also decreased with change in detritus import. Of the three different scenarios, the robustness values did not differ much from the original values in the autotrophic biomass scenario but changed comparatively more in the other two scenarios—fish harvest and detritus import. These results clearly indicate that the robustness measure is able to reflect the information about the system even when it faces both smaller and larger scale perturbations. The system maintained a balance in the face of stress, and only beyond a threshold are significant changes in robustness apparent.

Degrees of order (AMI/H, or A/C) and corresponding magnitudes of robustness for the Mdloti estuarine system for networks of the open (network 1) and closed (network 2) phases for a autotrophic biomass change scenario, b fish harvest scenario, and c detritus import scenario. Reprinted from Mukherjee et al. (2015), with permission from Elsevier
4.2 Socioeconomic Example: Water Network in China
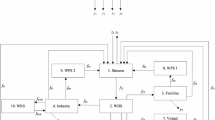
The same information theory-based network resilience measures were applied to a socioeconomic system, a Virtual Water Network (VWN) from the Heihe River basin in China (Fang et al. 2014). Unlike fresh water management strategies, which mainly focus on the efficiency of the target production processes of water utilisation, the VWN provides an integral view via the link of virtual water flows with different socioeconomic activities. The notions of virtual water flows provide important indicators to manifest the water consumption and allocation between different sectors via product transactions, as all production processes utilise water to varying degrees. During this study of Ganzhou District in the Heihe River Basin, we investigated configurations of a virtual water network (VWN) to identify the water network efficiency and stability in the socioeconomic system.
The system was divided into six sectors, representing the Ganzhou District economy. These include farming (1), livestock (2), other agricultural (3), industry (4), construction (5), and services (6), the three sectors related to agriculture emphasize the important role of this sector in the district (Fang et al. 2014), and also its comparatively high water consumption (Cheng et al. 2006). The system was then analysed over four phases representing the period 2002–2010. Results based on ENA show that the agricultural sector is the major local economic sector with high-intensity water consumption. Changes in the network structure occurred in the years 2003, 2005, and 2007. The number of links declined from 31 in 2002, to 30 in 2003/4 with the disappearance of the link from livestock (2) to other agriculture (3). The lowest number of links (26) occurred from 2005 to 2007 as the flows from livestock (2) to farming (1), other agricultural (3) to farming (1), other agricultural (3) to livestock (2), and the self-loop of construction sector (5) disappeared. In the last period (2008–2010), the flow from livestock (2) to other agricultural (3) is again present. Overall, the main changes in links occurred among the three agricultural sectors, i.e., farming (1), livestock (2), and other agricultural (3).
Although the total fresh water consumption declined over the time period of the studies, the efficiency of VWN was still low, and overall robustness declined over the study period (Fig. 11.5). Due to the low efficiency, the robustness of VWN remains on the left-side of the robustness curve (Fig. 11.6), implying that the VWN has a higher redundancy level due to stable circulation among various sectors but lower system efficiency, which is contrary to the intuition that human-designed networks should be more efficient. This occurs because, except for the freshwater flowing through specific supply chains, the water hidden in the products or services is circulating within diverse sectors with more exchanges along various pathways, which results in higher redundancy. The current policies for water management emphasize the control of the total amount of water consumption (with the aim of reduction), ignoring the balance between efficiency and redundancy for virtual water circulation within the socioeconomic system.

Trend in robustness values for the socioeconomic water system for the four periods from 2002 to 2010

Robustness of the networks in the four phases from 2002 to 2010. Note that each year is depicted by a circle
4.3 Relational Analysis
4.3.1 Keystone Species Analysis: South Africa
Here we present applications of the ecological relations analyses for ecological and socioeconomic systems, including the application to a perturbed estuarine systems in South Africa and to the Metropolitan area of Beijing, China. In the ecological study of the Mdloti estuary, South Africa, we investigated the impacts of keystone species on a time series of networks representing various seasons. Keystone species were identified using the method described above (Libralato et al. 2006; Ulanowicz and Puccia 1990).
The networks were perturbed to investigate possible temporal changes of their impacts on other network nodes and on the system as a whole. After the keystone species had been identified, its biomass was increased and decreased in 10% intervals to calculate their impacts on other nodes in the system. For five different time steps representing different seasons from March 2002 to March 2003, the identified keystone species were all fish: Argyrosomus japonicus (in two of the time steps), Monodactylus falciformis (in two of the time steps), and Caranx sexfasciatus (in one time step), ranging from trophic level 2.9 to 3.7 as calculated from the respective networks. Keystone species in the five time steps thus differed (the keystone species in one network is not necessarily the keystone in another), however each keystone species in a given network maintained its keystoneness rank over the perturbation scenarios.
The system level impact of the keystone species was at times prominent but not consistent through time (Fig. 11.7). In addition, there was no consistent impact on species belonging to similar trophic levels. Previous studies have shown that this system is very robust in nature (Mukherjee et al. 2015; Scharler 2012) and although variations in the presence of the keystone species affected some components, the resilience of the system as a whole counters these adverse effects and it does not collapse when the keystone species are perturbed or removed. This point is further supported by the change of the keystoneness of the species over time in this system.

System-level impacts on the total number of positive (black diamonds) and negative (grey squares) relations (y-axes) after perturbation of keystone species biomass in various seasons of the Mdloti Estuary. a March 2002, b June 2002, c September 2002, d December 2002, e March 2003
From this particular study it could be concluded that although there were significant effects of the keystone species in the ecosystem, that these effects were not consistent throughout the seasonal variation (Fig. 11.7). The ecosystem continues to function under the perturbations where keystone biomass is increased or depleted, but that was within a specific boundary beyond which the system could not persist (i.e. could not be mass balanced).
4.3.2 Relational Analysis: China
Another network indicator, related to system resilience, concentrates on the types of interaction between nodes (whether they are positive, negative, or neutral) (Fath 1998; Ulanowicz and Puccia 1990). A novel application of this approach was applied to assess the relation of carbon change due to land use change over time.
The acceleration of urbanisation has greatly changed the morphology of terrestrial surfaces, and about one-third of urban carbon emission results from land use/cover changes (LUCC) such as the replacement of vegetated surfaces with built-up land (Denman et al. 2007). Previous studies have shown a “carbon sink” effect caused by LUCC that promotes carbon sequestration by forests and grasslands after the conversion of farmland to forest or grassland at high latitudes, and a “carbon source” effect caused by the loss of these ecosystems (Kauppi et al. 1992). Beijing is a typical city whose distributions of LUCC vary greatly, which strongly affects the spatial and other characteristics of urban carbon emission and sequestration. Overall, urban resilience will be enhanced with improved carbon emission and sequestration management.
To achieve the research goal, first one needs to characterize the increase or decrease of carbon release and absorption under the existing LUCC conditions. And then, this needs to be analysed in context of the relationships between the social economic systems and the environment that causes these changes. Finally, the direction and size of carbon reduction can be determined based on the spatial distribution of these relationships. In urban systems, these relationships usually include mutualism, competition, exploitation, control, and neutralism (see Zhang et al. 2014).
In contrast to the robustness measure which explored the link distribution between pairs of nodes, utility analysis includes both direct and indirect pathways and how they affect the interaction signs between nodes (or sectors) of a system. For instance, direct feeding relations may reveal positive effects of prey to predator, and negative effects of predators on prey. These represent direct relations, but indirect relations may be classified by exploring the relationship between predators. If they are feeding on the same prey, then their relation is one of competition (negative effects both ways, i.e. −, −) although they are not directly linked through a feeding relation. As such, relations between nodes in a network according to positive and negative impacts describe the type of relation for each pair in a network either as positive (+), negative (−), or neutral (0), via all direct pathways. The relations may also be calculated over indirect pathways, in order to capture any indirect effects that travel over several pathway lengths between nodes (Ulanowicz and Puccia 1990; Fath 1998). The nature of the relations is important to understand the interaction between the nodes that may be mutualistic (+, +), competitive (−, −), absent (0, 0), or any other combination of positive, negative, and neutral (Table 11.1). As positive interactions, especially those configured into so-called autocatalytic loops, are thought to benefit an ecosystem (Ulanowicz and Puccia 1990), in economic sectors these relations may illustrate those sectors that have consistent positive or negative impacts on other sectors, from which may arise management scenarios to either maintain, or improve, certain relations.
The calculation of this particular relational analysis (Utility Analysis: Fath 1998) is very similar to that of the trophic impacts analysis (Ulanowicz and Puccia 1990) described above, and the similarities and differences are treated in Scharler and Fath (2009).
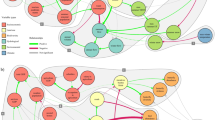
An application of utility analysis to the city of Beijing as a network of the different sectors illustrates the type of interactions between socioeconomic sectors. As urban areas contribute the majority of carbon emissions (IPCC 2007), this study illustrated how the various sectors interact, how each of them expanded or decreased their influence in the network, and which type of interactions were prominent among which sectors. Beijing’s urban area expanded to such an extent in recent decades that between 1992 and 2008 about 20% of cultivated land and 28% of the forested land was converted into constructed land (Miao et al. 2011). Within the time period studied here, major changes in land-use and cover occurred before 2000. After 2000, relationships between sectors changed mainly in the central and south-eastern parts of Beijing due to land-use and cover changes in these areas (Xia et al. 2016). Overall, there was a significant shift in the extent of area between sectors, and consequently a difference in the carbon emission and sequestration of each. The various sectors included in this analysis included urban land, rural land (residential area outside of urban land), transportation and industrial land outside urban and rural residential area, cultivated land, bare land, forest and water (reservoirs, rivers). Many of these had subcomponents to further identify the various sectors, resulting in 18 component networks replicated within and between four time periods representing the timespan from 1990 to 2010 (Xia et al. 2016) (Fig. 11.8).

The 18 component network representing the socioeconomic carbon metabolism network of Beijing. B1 sand, B2 barren earth, B3 bare exposed rock, C1 irrigated cultivated land, C2 dry cultivated land, F1 forest, F2 shrub land, F3 open woodland, F4 other woodland, G1 high-coverage grassland (vegetation cover >50%), G2 medium-coverage grassland (vegetation cover 20–50%), G3 low-coverage grassland (vegetation cover 5–20%), R rural, T transportation and infrastructure, U urban, W1 rivers, W2 lakes and reservoirs, W3 intermittently flooded plain, z ij input flow from j to i, y j , output flow from j to i. Reprinted from Xia et al. (2016), with permission from Elsevier
Major changes that occurred during the study period were apparent from the mutualistic relations (+, +) between the natural and socioeconomic sectors. The extent of mutualistic relations decreased substantially by more than 40%, which was mainly disturbed by urban expansion (Table 11.2). Mutualistic relations between the socioeconomic sectors were less easily disturbed. Mutualistic relations were also the most fluctuating in that they changed to a large extent between the time periods. Exploitative relations (+, −) occurred mainly between two components of the socioeconomic sector, transportation and industrial land. As these sectors are maintained and regulated by managers, they may therefore also provide an opportunity for regulation to reduce the extent of the exploitative relationship. Competitive relationships (−, −) arose mainly between natural components, or between cultivated land and natural components. Such relations were also the most stable over time (Table 11.2), which indicates that once established they are difficult to change. Before 2000, more positive than negative relations were found in the entire system, whereas negative relations mostly outweighed positive ones after 2000 (Xia et al. 2016).
Not only did the ecological relations change overall in the entire system, but the spatially explicit analysis revealed a concentration of change in certain areas of Beijing after 2000. Exploitation (+, −), control (−, +), and mutualism (+, +) relationships became concentrated in the southeast of the urban area, whereas competition relations (−, −) dominated its northwest expanse. The increase in competitive relations over time was a consequence of urban expansion competing for land, and increased use of water resources. The results provide an objective basis for planning adjustments to Beijing’s land-use patterns to improve its carbon metabolism and reduce carbon emission.
5 Discussion and Conclusions
Resilience has been an important topic in ecology for many decades, and has garnered attention and application in social and economic systems. In both domains, the same basic system-dynamic question applies in terms of the ability of a system to continue to function in the face of perturbations. Throughout its history, the resilience concept has included the premise of a steady state and return to that state, but recently has been expanded to encompass natural change and adaptation. A resilient system is not one that necessarily stays in the same place. Quite the contrary, the system must remain flexible and adaptive to changing conditions. This refers to either a return to the previous state or the continuation to a new state through a regime change. For example, as referenced above, Fath et al. (2015) frame resilience in terms of the adaptive cycle and a system’s ability to survive all phases of growth, conservation, collapse, and reorganization. In addition, similar features are seen as key to develop resilient systems such as incorporating functional diversity and also providing a balanced trade-off between efficiency and redundancy. Analysis of socioeconomic systems, using the information-based measure of balanced trade-off between efficiency and redundancy has shown that these systems have lower resilience (robustness) due to comparatively high redundancy (Kharrazi et al. 2013). In any event, the concern for resilience and the need for better metrics of resilience in socioeconomic systems provides a rich area for further research. Therefore, a main goal of this research was to illustrate the application of systems analysis concepts by providing additional case studies using a suite of resilience metrics derived from ecological network analysis.
The information-theory based analysis was applied to the reservoir ecosystem impacted by the Manwan Dam in Southwest China. This example showed a relatively high level of robustness in the unperturbed state (although not at the optimum which could be because the dam itself is a perturbation from the natural condition). Scenarios to remove functional groups showed that the robustness can be lowered most notably by altering flows and nodes regarding the phytoplankton, which is the key entry point for energy flow in the ecosystem. Application of the methodology to a human designed water supply network, also in China, exhibited a much lower overall robustness measure. The metric was tracked during four phases of water network development from 2002 to 2010 which showed a decrease in the overall robustness during this time. In this case, the goal of the regional water network was delivery for agricultural usage, and while water delivery was achieved it came at the expense of a more vulnerable network. Diversity and redundancy are two features which allow a system to respond effectively to perturbations. These key features must be balanced against efficiency in a trade-off that produces more resilient and robust systems.
These robustness/resilience values used herein have originally been conceived from the analysis of several ecological networks. The networks used to define the “Window of Vitality” as the region of highest robustness and sustainability, in a different approach (Zorach and Ulanowicz 2003) also match a confined region defined by minimum and maximum link density and effective number of trophic levels (Pimm 1982; Ulanowicz 2002; Wagensberg et al. 1990). As the robustness curve is not completely symmetrical, redundancy has a larger influence on resilience than efficiency [although the impact of a marginal change depends on which side of the optimum the system lies (Kharrazi et al. 2017)]. However, the authors (Goerner et al. 2009) concede that the optimum has not yet been defined due to a lack of data. The optimal balance between efficiency and redundancy, as well as the variability that may accompany such a balance is as yet to be explored. This research adds additional case studies employing this method showing that the ecological network of Manwan Dam (unperturbed) lies near this optimum region, and an agricultural water network lies off the optimum on the heavily redundant side, indicating it can improve its efficiency. The methodology allowed for tracking and improvement of resilience or efficiency in the respective networks.
Two additional case studies applied an energy-flow based network analysis that determines the direct and indirect relations between any two nodes in the network. In the South Africa case, this was used to reveal the keystone species in the Mdloti Estuary during five sampling periods from March 2002 until March 2003. Perturbations on the network had variable impacts on the presence of the keystone species, although a general pattern was not obvious. Lastly, the relational analysis was applied to carbon sequestration ability due to land use change around Beijing. Results showed that the number of mutualistic relations decreased over time and since the assessment was spatially explicit, we were able to show that competition relations increased in regions that experienced heavy urban expansion. This is a novel approach to use network analysis for assessing the carbon metabolism due to land use change.
In summary, networks are useful tools to describe the interactions between their constituents, and resilience measures applicable to both ecological and socioeconomic systems have been presented. They are derived from the efficiency of energy and material transfers throughout the networks, as well as the proportion of “detours”, or redundant pathways ensuring resilience. Positive and negative relations between sectors of economic systems, or ecosystems, highlight the influence of various economies and species on one another, resulting in a comprehensive picture of relations, impacts and therefore management options to achieve balance between sectors. Both relational analyses are related to resilience by describing the proportions of types of interactions in systems and their response to change and perturbation.
References
Admiraal, J. F., Wossink, A., De Groot, W. T., & De Snoo, G. R. (2013). More than total economic value: How to combine economic valuation of biodiversity with ecological resilience. Ecological Economics, 89, 115–122.
Allesina, S., & Pascual, M. (2008). Network structure, predator—prey modules, and stability in large food webs. Theoretical ecology, 1, 55–64.
Carpenter, S., Walker, B., Anderies, J. M., & Abel, N. (2001). From metaphor to measurement: Resilience of what to what? Ecosystems, 4, 765–781.
Chapin IIIi, F. S., Zavaleta, E. S., Eviner, V. T., Naylor, R. L., Vitousek, P. M., Reynolds, H. L., Hooper, D. U., Lavorel, S., Sala, O. E., Hobbie, S. E., Mack, M.C., & Díaz, S. (2000). Consequences of changing biodiversity. Nature 405, 235–242.
Chen, S., Fath, B. D., Chen, B. (2011). Information-based network environ analysis: A system perspective for ecological risk assessment. Ecological Indicators, 11(6), 1664–1672.
Chen, X., & Cohen, J. E. (2001). Transient dynamics and food-web complexity in the Lotka-Volterra cascade model. Proceedings of the Royal Society of London. Series B, Biological Sciences, 268, 869–877.
Cheng, G. D., Xiao, H. L., Xu, Z. M., Li, J. X., & Lu, M. F. (2006). Water issue and its counter-measure in the inland river basins of Northwest China—a case study in Heihe River Basin. Journal of Glaciology and Geocryology, 3, 406–413.
Christensen, V., & Walters, C. J. (2004). Ecopath with ecosim: Methods, capabilities and limitations. Ecological Modelling, 172, 109–139.
Chrystal, R. A., & Scharler, U. M. (2014). Network analysis indices reflect extreme hydrodynamic conditions in a shallow estuarine lake (Lake St Lucia), South Africa. Ecological indicators, 38, 130–140.
Denman, K. L., Brasseur, G., Chidthaisong, A., Ciais, P., Cox, P. M., Dickinson, R. E., Hauglustaine, D., Heinze, C., Holland, E., Jacob, D., Lohmann, U., Ramachandran, S., da Silva Dias, P. L., Wofsy, S. C., & Zhang, X. (2007). Couplings between changes in the climate system and biogeochemistry. In S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor, & H. L. Miller (Eds.), Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment. Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press.
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z.-I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A.-H., Soto, D., Stiassny, M.L.J., Sullivan, C.A. (2006). Freshwater biodiversity : Importance, threats, status and conservation challenges. Biological reviews of the Cambridge Philosophical Society, 81(2), 163–182.
Elliott, M., & Quintino, V. (2007). The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Marine Pollution Bulletin, 54, 640–646.
Elmqvist, T., Folke, C., Nyström, M., Peterson, G., Bengtsson, J., & Walker, B. (2003). Response diversity, ecosystem change, and resilience. Frontiers in Ecology and the Environment, 1, 488–494.
Elton, C. S. (1958). Ecology of invasions by animals and plants. London: Chapman & Hall.
Fang, D., Fath, B. D., Chen, B., & Scharler, U. M. (2014). Network environ analysis for socio-economic water system. Ecological indicators, 47, 80–88.
Fath, B. (1998). Network synergism: Emergence of positive relations in ecological systems. Ecological Modelling, 107, 127–143.
Fath, B. D., Scharler, U., Ulanowicz, R. E., & Hannon, B. (2007). Ecological network analysis: network construction. Ecological Modelling, 208, 49–55.
Fath, B. D., Dean, C. A., & Katzmair, H. (2015). Navigating the adaptive cycle: An approach to managing the resilience of social systems. Ecology and Society, 20(2), 24.
Folke, C. (2006). Resilience: The emergence of a perspective for social—ecological systems analyses. Global Environmental Change, 16, 253–267.
Folke, C., Carpenter, S., Walker, B., Scheffer, M., Elmqvist, T., Gunderson, L., et al. (2004). Regime shifts, resilience, and biodiversity in ecosystem management. Annual Review of Ecology Evolution and Systematics, 35, 557–581.
Gamfeldt, L., & Hillebrand, H. (2008). Biodiversity effects on aquatic ecosystem functioning—maturation of a new paradigm. International Review of Hydrobiology, 93, 550–564.
Goerner, S. J., Lietaer, B., & Ulanowicz, R. E. (2009). Quantifying economic sustainability: Implications for free-enterprise theory, policy and practice. Ecological Economics, 69, 76–81.
Grilli, J., Rogers, T., & Allesina, S. (2016). Modularity and stability in ecological communities. Nature Communications, 7, 12031.
Gunderson, L. H., & Holling, C. S. (Eds.). (2002). Panarchy: Understanding transformations in human and natural systems. Washington DC: Island Press.
Higashi, M., & Patten, B. C. (1989). Dominance of indirect causality in ecosystems. The American Naturalist, 133, 288–302.
Higashi, M., & Nakajima, H. (1995). Indirect effects in ecological interaction networks. I. The chain rule approach. Mathematical Biosciences, 130, 99–128.
Holling, C. S. (1973). Resilience and the stability of ecological systems. Annual Review of Ecology and Systematics, 4, 1–23.
Holling, C. S. (1986). The resilience of terrestrial ecosystems: local surprise and global change. In W. C. Clark & R. E. Munn (Eds.), Sustainable Development of the Biosphere. London: Cambridge University Press.
Hughes, T. P., Baird, A. H., Bellwood, D. R., Card, M., Connolly, S. R., Folke, C., et al. (2003). Climate change, human impacts, and the resilience of coral reefs. Science, 301, 929–934.
IPCC—Report of the intergovernmental panel on climate change (2007). Fourth Assessment Report. Climate change 2007: Synthesis report. Cambridge University Press. ISBN 92-9169-122-4.
Jørgensen, S. E., Fath, B. D., Nielsen, S. N., Pulselli, F., Fiscus, D., Bastianoni, S. (2015). Flourishing within limits to growth: Following nature’s way. Earthscan Publisher. 220 p.
Kharrazi, A., Rovenskaya, E., Fath, B. D., Yarime, M., & Kraines, S. (2013). Quantifying the sustainability of economic resource networks: An ecological information-based approach. Ecological Economics, 90, 177–186.
Kharrazi, A., Fath, B. D., & Katzmair, H. (2016). Advancing empirical approaches to the concept of resilience: A critical examination of panarchy, ecological information, and statistical evidence. Sustainability, 8, 935.
Kharrazi, A., Rovenskaya, E., & Fath, B. D. (2017). Network structure impacts global commodity trade growth and resilience. PLoS ONE, 12(2), e0171184. https://doi.org/10.1371/journal.pone.0171184.
Kauppi, P. E., Mielikainen, K., & Kuusela, K. (1992). Biomass and carbon budget of European forest, 1971 to 1990. Science, 256, 70–74.
Libralato, S., Christensen, V., & Pauly, D. (2006). A method for identifying keystone species in food web models. Ecol. Modell., 195, 153–171.
Loreau, M. (2000). Biodiversity and ecosystem functioning: Recent theoretical advances. Oikos, 91, 3–17.
MacArthur, R. H. (1955). Fluctuations of animal populations and a measure of community stability. Ecology, 36, 533–536.
May, R. M. (1972). Will a large complex system be stable? Nature, 238, 413–414.
McCann, K. S. (2000). The diversity-stability debate. Nature, 405, 228–233.
McNaughton, S. J. (1977). Diversity and stability of ecological communities: A comment on the role of empiricism in ecology. The American Naturalist, 111(979), 515–525.
MEA. (2005). Millenium ecosystem assessment, 2005. http://www.millenniumassessment.org.
Meadows, D. H., Meadows, D. L., Randers, J., & Behrens, W. W., III. (1972). The limits to growth. New York: Universe Books.
Miao, L. J., Cui, L. F., Luan, Y. B., & He, B. (2011). Similarities and differences of Beijing and Shanghai’s land use changes induced by urbanization. Chinese Journal of Metal Science and Technology, 31(4), 398–404.
Moore, J. C., & de Ruiter, P. C. (2012). Energetic Food Webs. An analysis of real and model ecosystems: Oxford University Press.
Moore, J. C., de Ruiter, P. C., & Hunt, H. W. (1993). Influence of productivity on the stability of real and model ecosystems. Science, 261, 906–908.
Mori, A. S., Furukawa, T., & Sasaki, T. (2013). Response diversity determines the resilience of ecosystems to environmental change. Biological Reviews of the Cambridge Philosophical Society, 88, 349–364.
Müller, F., Bergmann, M., Dannowski, R., Dippner, J. W., Gnauck, A., Haase, P., et al. (2016). Assessing resilience in long-term ecological data sets. Ecological indicators, 65, 10–43.
Mukherjee, J., Scharler, U. M., Fath, B. D., & Ray, S. (2015). Measuring sensitivity of robustness and network indices for an estuarine food web model under perturbations. Ecological Modelling, 306, 160–173.
Naeem, S., Chapin III, F. S., Costanza, R., Ehrlich, P. R., Golley, F. B., Hooper, D. U., Lawton, J. H., O’Neill, R. V., Mooney, H. A., Sala, O. E., Symstad, A. J., & Tilman, D. (1999). Biodiversity and ecosystem functioning: Maintaining natural life support processes. Issues in Ecology, 4, 1–11. Published by the Ecological Society of America.
Neutel, A.-M., Heesterbeek, J. A. P., & De Ruiter, P. C. (2002). Stability in real food webs: Weak links in long loops. Science, 296, 1120–1123.
Pimm, S. L. (1982). Foodwebs. London: Chapman and Hall.
Power, M. E., Tilman, D., Estes, J. A., Menge, B. A., Bond, W. J., Mills, L. S., et al. (1996). Challenges in the quest for keystones. BioScience, 46, 609–620.
Rist, L., Felton, A., Nyström, M., Troell, M., Sponseller, R. A., Bengtsson, J., et al. (2014). Applying resilience thinking to production ecosystems. Ecosphere, 5, 1–11.
Rutledge, R. W., Basore, B. L., & Mulholland, R. J. (1976). Ecological stability: An information theory viewpoint. Journal of Theoretical Biology, 57, 355–371.
Salas, A. K., & Borrett, S. B. (2011). Evidence for dominance of indirect effects in 50 trophic ecosystem networks. Ecological Modelling, 222, 1192–1204.
Scharler, U. M. (2012). Ecosystem development during open and closed phases of temporarily open/closed estuaries on the subtropical east coast of South Africa. Estuarine, Coastal Shelf Science, 108, 119–131.
Scharler, U. M., & Fath, B. D. (2009). Comparing network analysis methodologies for consumer—resource relations at species and ecosystems scales. Ecological Modelling, 220, 3210–3218.
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., & Walker, B. (2001). Catastrophic shifts in ecosystems. Nature, 413, 591–596.
Shevtsov, J., & Rael, R. (2015). Indirect energy flows in niche model food webs: Effects of size and connectance. PLoS ONE, 10(10), e0137829.
Szyrmer, I., & Ulanowicz, R. E. (1987). Total flows in ecosystems. Ecological Modelling, 35, 123–136.
Tilman, D. (1999). The ecological consequences of changes in biodiversity: A search for general principles. Ecology, 80, 1455–1474.
Ulanowicz, R. E. (1986). Growth and development. New York: Springer.
Ulanowicz, R. E. (2002). Information theory in ecology. Journal of Computational Chemistry, 25, 393–399.
Ulanowicz, R. E. (2004). Quantitative methods for ecological network analysis. Computers and Chemistry, 28, 321–339.
Ulanowicz, R. E. (2009). The dual nature of ecosystem dynamics. Ecological Modelling, 220(16), 1886–1892.
Ulanowicz, R. E., & Puccia, C. J. (1990). Mixed trophic impacts in ecosystems. COENOSES, 5, 7–16.
Ulanowicz, R., Goerner, S., Lietaer, B., & Gomez, R. (2009). Quantifying sustainability: Resilience, efficiency and the return of information theory. Ecological Complexity, 6, 27–36.
Wackernagel, M., Schulz, N. B., Deumling, D., Linares, A. C., Jenkins, M., Kapos, V., et al. (2002). Tracking the ecological overshoot of the human economy. PNAS, 99, 9266–9271.
Wagensberg, J., Garcia, A., & Sole, R. V. (1990). Connectivity and information transfer in flow networks: Two magic numbers in ecology? Bulletin of Mathematical Biology, 52, 733–740.
Wootton, J. T. (1994). The nature and consequences of indirect effects in ecological communities. Annual Review of Ecology and Systematics, 25(1), 443–466.
Xia, L., Fath, B. D., Scharler, U. M., & Zhang, Y. (2016). Science of the total environment spatial variation in the ecological relationships among the components of Beijing’s carbon metabolic system. Science of the Total Environment, 544, 103–113.
Zhang, Y., Xia, L. L., & Xiang, W. N. (2014). Analyzing spatial patterns of urban carbon metabolism: A case study in Beijing, China. Landscape and Urban Planning, 130, 184–200.
Zorach, A. C., Ulanowicz, R. E. (2003). Quantifying the complexity of flow networks: How many roles are there?. Complexity, 8(3), 68–76.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Scharler, U.M. et al. (2018). Resilience Measures in Ecosystems and Socioeconomic Networks. In: Mensah, P., Katerere, D., Hachigonta, S., Roodt, A. (eds) Systems Analysis Approach for Complex Global Challenges. Springer, Cham. https://doi.org/10.1007/978-3-319-71486-8_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-71486-8_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-71485-1
Online ISBN: 978-3-319-71486-8
eBook Packages: EngineeringEngineering (R0)