
Abstract
Web building has been such a highly successful foraging innovation among spiders that the vast majority of extant spiders are web builders. The structure of spider webs varies substantially between species, and web building has even been lost completely in some clades. Examples of different web forms include the classic orb webs, which may be orientated vertical to the ground or horizontal, sheet webs, and cobwebs, which consist of three-dimensional meshwork and ascending sticky threads for support and capture of prey. The architecture of webs may also vary within clades and even within species. This may be a consequence of: (i) individuals adapting their web structures to the environment; e.g., larger webs are built in areas where more space is available, (ii) spiders varying their webs to tune its performance, e.g., when spiders are exposed to different prey, or (iii) silk expression constraints, e.g., when on diets lacking certain nutrients. We review the literature, focusing on contributions from the Neotropical region, showing that spider webs vary in structure and function at multiple levels and so must be considered a dynamic, variable, extended phenotype of its builder. Webs accordingly depict the foraging, mating, and defensive strategies, and physiological status, of the spider.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Spiders of the infraorder Araneomorphae, which are often misleadingly (as not all members build webs) called web-building or true spiders (Turnbull 1973), comprise 93.9% of all extant spider species, and thus represent by far the most diverse spider group. The webs of Araneomorph spiders are highly distinguishable and found in almost every ecosystem on Earth, but they are particularly abundant in the Neotropics.
The primarily function of spider webs is to capture prey. However, they can serve other functions, including acting as a sensory system, a courtship and/or mating platform, thermoregulatory platform, and antipredator barrier. Despite their ubiquitousness across environments, the ecological and structural importance of spider webs within ecosystems is poorly known. This is partly because the ecological, evolutionary, and biophysical aspects of webs for individual spiders, populations, and species are largely unexplored. A significant portion of what is known about spider web diversity, evolution, ecology, and building behaviours is a consequence of over a century of detailed observations on Neotropical spiders.
Probably the most readily recognizable form of spider web is the orb web. It was once thought to be the pinnacle of spider web evolution. New molecular evidence has nonetheless suggested two alternative scenarios (Bond et al. 2014; Fernández et al. 2014): (1) that the orb web evolved earlier than originally postulated and may represent the ancestral form of all spider webs, or (2) the orb web has had multiple independent origins. The latter hypothesis would conform with hypotheses formed prior to the advancement of more methodical cladistic analyses (e.g., Lehtinen 1967) but seems to be the less likely. We do not weigh into the debate regarding the origins of orb webs herein. Rather, we explore how studies using Neotropical spiders have assisted the development of the various hypotheses over the years.
In addition to our understanding of the evolutionary trajectories of spider webs, our understanding of the function of spider webs has been significantly enhanced of late (see Blackledge et al. 2011; Harmer et al. 2011). Still lacking, however, is a clear understanding of the genetic underpinnings of spider webs and web building. Our knowledge of the genetic underpinnings of various spider silks has expanded of late (Prosdocimi et al. 2011; Clarke et al. 2015; Collin et al. 2016), and this knowledge might provide insights into the genetic influences over web building and web functionality. Here, we overview spider web and silk diversity and variability, with specific reference to the plenitude of work on Neotropical spiders.
Neotropical Araneomorph Diversity
We know that up to half of all genera of orb-web spiders are found in the Neotropics. Brazil alone has arguably the world’s greatest web-building spider diversity, containing up to 72 of the 112 known families of Araneomorphs (World Spider Catalog 2016). Such an impressive diversity qualifies the Neotropics as a spider biodiversity hotspot and highlights the importance of on-going research in the region (see Chap. 1).
A wide variety of web types and foraging modes are found among Neotropical Araneomorphs. These include the use of silken aerial and ground webs, snares and trip lines, sit-and-wait and cursorial foraging. Among the webs, a diversity of web architectures are found (depicted in Fig. 6.1), from two-dimensional planar horizontal or vertical orb webs, to three-dimensional sheet webs and cobwebs, to elongated two dimensional ‘ladder webs’, highly modified webs, and webs comprising but a few capture threads.

Examples of the diversity of Neotropical spider web architectures, showing (clockwise from the upper left-hand side) two-dimensional Araneid planar horizontal orb webs, an example of a derived orb web, Theridiid cobwebs with gumfooted threads, three-dimensional sheet webs, and three-dimensional orb webs of Cyrtophora and Mecynogea. An example of a ‘ladder web’ is shown on the trunk of the tree
Some Examples of Well-Known Araneomorph Groups
Synotaxidae
The Synotaxidae are a family of spiders found in South America, Australia, and New Zealand (Griswold et al. 1998; Agnarsson 2003a). Spiders of this group were once thought to belong to the Theridiidae and to be related to the Argyrodinids (Forster et al. 1990; Griswold et al. 1998; Agnarsson 2003a). Nevertheless, upon detailed assessments of leg and abdomen micro-characteristics, spiders in the group were designated to an independent family (Forster et al. 1990; Agnarsson 2003a). A diagnostic characteristic of the Synotaxidae is the unique ‘chicken-wire’ web comprising irregularly meshed silk with vertically and horizontally aligned sticky silk threads (Eberhard 1977, 1995; Griswold et al. 1998; Dimitrov et al. 2017) (Fig. 6.2). The web building, eggsac production and prey-wrapping behaviours in Synotaxus ecuadorensis are well described and seem to be typical of the group (Eberhard 1995; Barrantes and Eberhard 2007).

The unique ‘chicken-wire’ web of the Synotaxidae, comprising irregularly meshed silk with vertically and horizontally aligned sticky silk threads (From http://tolweb.org/Synotaxidae/93137)
RTA Clade
The retrolateral tibial apophysis (RTA) is a megadiverse clade containing almost half of all Araneomorph spiders. The majority of species in the RTA clade do not build webs. The latest genomic analyses have nevertheless placed the clade sister to the web-building Deinopoidae (Bond et al. 2014; Fernández et al. 2014). This suggests that the RTA clade represent a group that lost the ability to build webs. The few RTA spiders that build webs today are thus interesting subjects for testing hypotheses about the costs and benefits of web building.
The Neotropics are rich in RTA clade spiders (Santos and Brescovit 2001; Santos 2007; Silva et al. 2008). Significantly, the Neotropics has good representations of web-building RTA species, including species of web-building pisaurids (Pisauridae) and wolf spiders (Lycosidae). Accordingly, the region seems to be a hotbed for studies testing hypotheses about the evolutionary significance of web building (Stefani et al. 2001; Macrini et al. 2015).
The genus Aglaoctenus (Lycosidae), for example, is widely distributed across South America (Uruguay to Colombia) and found in a variety of different environments (Santos and Brescovit 2001; Piacentini 2011; González et al. 2015a, b). This is particularly interesting because the genus includes species that build tube-shaped webs and display sub-social behaviour (Macrini et al. 2015). Spiders of the genus belong to an ancient subfamily of wolf spiders (Sosippinae). Accordingly, it may be hypothesized that the modern wolf spider had at some time jettisoned web building (Murphy et al. 2006). The webs of Aglaoctenus spp. are comprised of a series of attachments, a meshed sheet, and a funnel in which the spider can retreat (Stefani and Del-Claro 2015) (Fig. 6.3). The tube-shaped webs are built low to the ground, and may be a modification of a web resembling that of some ecribellate spiders (Murphy et al. 2006).

Web of Aglaoctenus lagotis (Lycosidae) from Serra do Japi, SP, Brazil. (Photo: M.O. Gonzaga)
A preliminary silk gene expression analysis for Aglaoctenus lagotis and A. oblongus from Uruguay shows the presence of the MaSp2 protein (spidroin) gene (Blamires, unpublished data). This finding, if confirmed, is interesting because the MaSp2 protein was thought to appear first in the ecribellate spiders (Hinman and Lewis 1992; Blackledge et al. 2012; Blamires et al. 2017). Clearly more comparative genetic and behavioural analyses are needed to reveal the evolutionary origins of the various silk proteins, the RTA spider webs, cribellate spider webs, and modern ecribellate spider webs.
Uloboridae
Neotropical spiders of the family Uloboridae usually spin horizontal orb webs containing a fibrous dry ecribellate adhesive silk (see section on ‘Spider silks’) (Lehtinen 1967; Eberhard 1972; Opell 1987). Modifications of the usual horizontally aligned orb webs, such as the highly reduced webs of Polenecia spp. (Wiehle 1927), Hyptiotes spp. (Peters 1938; Opell 1982), and Miagrammopes (Pickard-Cambridge 1904; Lubin et al. 1978; Opell 1987), are well described (see also Nascimento and Gonzaga 2015, Santos and Gonzaga 2017 for variations).
There are numerous publications on the web-building behaviours of Neotropical Uloborids thanks to almost 50 years of meticulous observations by Eberhard (Eberhard 1972, 1973, 1990a; Lubin et al. 1978; Opell and Eberhard 1984; Eberhard and Barrantes 2015). Significantly meticulous behavioural observations of Neotropical Uloborids were among the first to document variability in web building and web architectures across environments and contexts (Eberhard 1990a). These studies have been integral to our understanding of the evolution of different Araneomorph web forms (Eberhard 1990b; Eberhard 2014; Hormiga and Griswold 2014; Eberhard and Barrantes 2015).
Theridiidae
Sociality has evolved four times within the theridiid clade, suggesting the clade contains characters and behaviours that promote sociality (Agnarsson 2002; Agnarsson et al. 2006, 2007). Members of the genus Anelosimus have evolved a unique sociality, one with no hierarchy but the sharing of brood care, prey capture, and web construction. As a consequence their behaviours and communal webs are exceptionally well studied (Nentwig and Christenson 1986; Agnarsson 2002; Whitehouse and Lubin 2005; Yip et al. 2008) (see Chap. 13).
Theridiids of the genus Argyrodes have attracted similar interest because of their kleptoparasitic lifestyle (Agnarsson 2002; Su and Smith 2014). Interestingly, phylogenetic analyses of both sociality and kleptoparasitism show similar origins, and both may have diverged from some form of maternal care (Agnarsson 2002). Unlike sociality, kleptoparasitism arose only once in the Theridiidae (Agnarsson 2002). Neotropical species of the genus Argyrodes inhabit the webs of larger spiders, most commonly Nephila clavipes, and steal food from the host’s web. While Argyrodes kleptoparasites have a distribution beyond the Neotropics, the Neotropical species have been most extensively studied (Vollrath 1979; Agnarsson 2002, 2004, 2011). From these studies we know much about the behaviours of Aryrodes spp. and the type of interactions they have with their host spider and its web (Vollrath 1979; Higgins and Buskirk 1998; Agnarsson 2002, 2003b, 2011). These studies by and large suggest that Argyrodes negatively influences the fitness of the host (Vollrath 1979; Higgins and Buskirk 1998). However, it seems that under certain ecological circumstances some Argyrodes can benefit the host spider (Elgar 1994; Peng et al. 2013).
The cobwebs and combfoot webs made by theridiid spiders are well-known. These include the webs built by Neotropical Latrodectus spp., Steatoda spp. and Theridion spp. These spiders all construct three-dimensional ‘irregular sheet’ webs (Benjamin and Zschokke 2002). The cobweb typically has four additional components: (i) a tangled retreat, a small ‘pocket’ consisting of supporting tangle threads and a non-sticky sheet where the spider hides, (ii) supporting threads, which form a large inaccessible tangle, (iii) gumfooted threads which vertically descend from the tangled retreat and interact with prey crawling below the web, and (iv) gumfooted gluey silk droplets at the base of the ascending threads, which adhere to the crawling prey (Fig. 6.4). Distinctive web patterns in Neotropical theridiids, however, have been previously described. This is the case of the webs of Helvibis longicauda and Chrysso intervales, which are entirely composed of viscid silk lines (Gonzaga et al. 2006).

(a) Diagram of a cobweb showing the architectural components: the retreat consisting of supporting tangle threads and a non-sticky sheet (a), b vertically descending gumfooted threads, and c gumfoot sticky droplets deposited at the base of the gumfooted threads. (b) Web of Achaearanea tingo (Theridiidae) from Parque Estadual Intervales, Ribeirão Grande, SP, Brazil (Photo: M.O. Gonzaga)
Araneidae
Spiders in this family are abundant in Neotropical ecosystems (Bonaldo et al. 2007; Baldissera et al. 2008). The depth of spider research in the region means that there is a dearth of information on the diversity, behaviours, ecology, and phenology of Neotropical Araneid spiders. There is information, however, on several aspects of their biology in the Neotropics, such as foraging strategies (e.g., Moura et al. 2016; Rito et al. 2016), habitat selection (e.g., Messas et al. 2014; Souza et al. 2015), web structures and architecture (e.g., Eberhard 1988b, 2008, 2014; Xavier et al. 2017), sexual selection and parental care (e.g., Moura et al. 2017; Moura and Gonzaga 2007), defences against natural enemies (e.g., Eberhard 2003; Gonzaga and Vasconcellos-Neto 2005; Magalhaes and Santos 2012), among others.
Nephila clavipes is one of the most well-known of all Neotropical orb-web Araneids due to its size, abundance, and ubiquitousness throughout the Neotropics. The species builds a large (often several metres in width and height), tightly meshed, two-dimensional orb web (often with an accompanying three-dimensional ‘barrier web’ that may contain stacked prey carcasses, Higgins 1992) across high canopy gaps in the forest, where they potentially capture almost all insects prey that flies through the corridor (Vasconcellos-Neto and Lewinsohn 1984). They have also been known to capture and consume small bats and birds (Brooks 2012; Nyffeler and Knornschild 2013) and are a common host of Argyrodes kleptoparasites (Vollrath 1979). Like other species in the genus (Blamires et al. 2010), N. clavipes often aggregates its web. The behaviours and ecology of N. clavipes are so well-studied that it serves as a model for a plenitude of web and silk studies, and behavioural phenomena, such as centrally placed foraging, sexual cannibalism, and sexual-size dimorphism (Herberstein and Hebets 2013).
Tetragnathidae
Spiders of the genus Leucauge are very common in Neotropical forests and are identified by their distinctive silver, black, green, and/or yellow body colouration (Tso et al. 2006) and their sub-canopy, horizontally aligned two-dimensional orb webs (Eberhard 1990a, b; Hénaut et al. 2006; Briceno and Eberhard 2011). They may be solitary, aggregated, or subsocial (Eberhard 1990b; Salomon et al. 2010).
The web-building behaviours of L. mariana, for example, have been the subject of close observation as they make precision movements to produce webs with highly consistent spacing between spiral threads despite evidently constructing the web ‘blindly’ (Eberhard 1987, 1988a, b; Briceno and Eberhard 2011). The extra reach of their extensively long legs I and II are thought to facilitate rapid grasping of the radial threads and precise placement of the spiral threads during construction (Briceno and Eberhard 2011).
Spider Silks
Araneomorphs have evolved a ‘toolkit’ of silks, with each silk having particular properties for particular uses (Blackledge and Hayashi 2006). We will overview the work on silks used predominantly in webs, namely the major ampullate (MA), aciniform, and the cribellate and ecribellate gluey silks. While minor ampullate, pyriform, and some other silks (e.g., pseudoflagelliform and ribbon silks of filistatids) may function within webs, there is scant documentation of their functions within Neotropical spider webs.
Major Ampullate Silk
The longstanding model organism for studies of MA silk proteins, protein structures, and physical properties is Nephila clavipes. This is partly because its large body size makes it amenable for keeping in the laboratory and repeatedly extracting large quantities of silk, which is essential for most chemical assays, and partly because the pioneering work was done using this species (Lewis 1992).
The interest in MA silk properties emanates from its unique combination of high strength, extensibility, and toughness (Lewis 1992; Craig 2003; Harmer et al. 2011; Blamires et al. 2017). Accordingly, there are a multitude of potential commercial implications associated with understanding the mechanisms by which MA silk performs. Ecologically, the properties of MA silk enable it to function efficiently as a safety line, a propagator of tactile information, as a supporting frame for the web, and as impact-absorbing radial threads in orb webs (Osaki 1996; Sensenig et al. 2012; Blamires et al. 2017).
Phylogenetic analyses show that a stiff MA silk precursor was secreted by the major ampullate glands prior to the evolution of aerial web building (Garb et al. 2007, 2010; Prosdocimi et al. 2011). MA silk appeared in the first webs as moorings and supports (Denny 1976; Craig 2003). The proline-rich, highly elastic protein MaSp2 was first incorporated into MA silk by orb-web-building spiders (Hinman and Lewis 1992; Blackledge et al. 2012). Its high extensibility meant that incorporating radial threads into orb webs remarkably improved the web’s ability to absorb the kinetic energy imparted by impacting prey (Denny 1976; Blackledge et al. 2012; Sensenig et al. 2012). Furthermore, the inclusion of the MaSp2 protein induced MA silks to shrink upon exposure to water, a phenomena called supercontraction (Boutry and Blackledge 2010). Supercontraction made it possible for MA silk to become stretched in humid environments, and for the web to tighten when the humidity fell (Boutry and Blackledge 2010, 2013). A finding of the MaSp2 gene in the Uruguayan wolf spider (RTA clade) Aglaoctenus lagotis, and the prsence of a MaSp2-like protein in cribellate silks (Piorkowski and Blackledge 2017), suggests that MaSp2 may have evolved prior to the building of two-dimensional, vertically aligned ecribellate orb webs.
Aciniform Silk
Aciniform silk is used by most Araneomorph spiders for wrapping and immobilizing prey, building sperm webs, as an egg-case liner, and for web decorations in Argiope argentata (Blackledge and Hayashi 2006). Aciniform silk, like major ampullate silk, is comprised of a core of spidroins (Hayashi et al. 2004). The aciniform spidroin, AcSp1, of Latrodectus spp. contains poly-alanine motifs similar to major ampullate silk, albeit containing fewer glycine motifs (Ayoub et al. 2013). Aciniform silk of the widely distributed Argiope trifasciata is thought to be as tough as major ampullate silk (Hayashi et al. 2004). This high toughness seems to be functionally important for protecting eggs (Blackledge and Hayashi 2006; Hsia et al. 2011). Nevertheless, when aciniform silk is used as web decorations by Argiope spp. it plays no role in the web’s mechanical performance (Herberstein et al. 2000a; Blackledge and Hayashi 2006). It is probably utilized because it reflects ultraviolet light more strongly than the web silks, and so is useful as a silk-based signal to predators, prey, or perhaps both (Cheng et al. 2010).
Cribellate Silk
Spiders from the Deinopidae and Uloboridae families produce dry cribellar silks made of thousands of silk fibrils that surround supporting axial fibres (Opell and Bond 2000). These spiders produce their cribellar silks by drawing fibrils from spigots on the cribellum using the calamistrum, a comb of setae on the metatarsus of the fourth legs (Hawthorn and Opell 2003). The fine fibrils combine with thicker supporting strands to produce the characteristic wooliness of the silk (Eberhard and Pereira 1993; Hawthorn and Opell 2003).
Cribellate silk are used as capture threads in the horizontal orb webs of Uloborids. These wool-like threads are relatively stiff and inelastic compared to those of ecribellate orb webs (Lubin 1986; Kohler and Vollrath 1995; Blackledge and Hayashi 2006). They adhere to insects that get caught in the web via van der Waals and hygroscopic attractive forces between the thread and insect cuticle (Hawthorn and Opell 2003; Sahni et al. 2014a). Opell (1994a, b), examined the properties of the spiral threads of Neotropical Miagrammopes spp., Hyptiotes spp., and Uloborus spp., and found differences in stickiness across species. Web architecture appeared to be a major factor correlating with stickiness, with the more reduced webs of Miagrammopes and Hyptiotes being the stickiest.
Ecribellate Spiral Threads
The spiral threads of the derived Araneoid spiders, including orb weavers and cobweb builders, are made from either pseudoflagelliform or flagelliform silk coated with viscid aggregate silks (Townley and Tillinghast 2013; Sahni et al. 2014a, b). The flagelliform and aggregate silk genes and their orthologues were first identified and described for the Neotropical orb web spider Nephila clavipes (Hayashi and Lewis 1998; Choresh et al. 2009; Collin et al. 2016).
The viscid coated threads are produced by a triad of two aggregate-secreting and one flagelliform-secreting spigots located on the posterior lateral spinneret (Sahni et al. 2014a). The aggregate silk coating the flagelliform thread is highly hygroscopic and absorbs atmospheric moisture immediately upon spinning, thus swelling before transfiguring into regularly distributed ellipsoid droplets along the thread that resemble beads along a string (Edmonds and Vollrath 1992; Sahni et al. 2014a; Townley et al. 2006).
The stickiness of the viscid spiral threads is due to the presence of glycoproteins (Opell and Hendricks 2010; Townley and Tillinghast 2013; Sahni et al. 2014a, b). The aggregate silk additionally contains inorganic salts, as well as organics salts, amino acids (e.g., glycine), neurotransmitters and saturated fatty acids (Vollrath et al. 1990). The viscid threads are produced more quickly and are less visible to insects than cribellate sticky silks (Craig 1986; Opell 1996, 1998). This advantage may have driven the immense diversity of ecribellate spiders that exists today.
Other Silks
A defining feature of Araneomorphs is their ability to spin multiple task-specific silks (Vollrath and Knight 2001; Blackledge and Hayashi 2006; Blamires et al. 2017). Silks utilized in addition to major ampullate, aciniform, and different kinds of sticky silks include tubuliform silk, which has high stiffness and low toughness (Blackledge and Hayashi 2006) and is used to form the inner coating of eggsacs (Hu et al. 2005; Tian and Lewis 2005; Gnesa et al. 2012). A phylogenetic examination across the major Araneomorph families (including Deinopidae, Uloboridae, Theridiidae, Araneidae) shows the TuSp1 gene to be, despite its property differences, monophyletic and closely related to all of the other spidroin genes (Tian and Lewis 2005; Garb et al. 2010).
Minor ampullate (MiA) silk is used as a temporary capture spiral in orb webs, for prey-wrapping by theridiids, and as a component of dragline threads by most spiders (Vollrath and Knight 2001; La Mattina et al. 2008; Blamires et al. 2017). While most of the mechanical properties of minor ampullate silk are comparable with major ampullate silk (Blackledge and Hayashi 2006), it does not, unlike major ampullate silk, supercontract in water (Blamires et al. 2017). The spidroin-coding genes of minor ampullate (MiSp1 and MiSp2) spidroins have been identified in Nephila clavipes and seem to be MaSp1 orthologues (Colgin and Lewis 1998).
Pyriform silks cement the dragline and web frame to substrates, and glue silk threads together during web construction (Perry et al. 2010; Wolff et al. 2015). The spidroin PySp1 is manufactured exclusively in the pyriform gland of black widow spiders (Blasingame et al. 2009). A comparative PCR analysis of the Argiope trifasciata, Nephila clavipes, and Nephilingis cruentata PySp1 protein found two amino acid motifs unique to the spidroin: (1) a motif where every other amino acid is proline, and (2) a glutamine-rich motif of 6–8 amino acids (Perry et al. 2010). Nevertheless, relatively little is known of the properties of this spidroin.
A ribbon-like prey-wrapping silk is found in among spiders from the family Filistatidae (Eberhard and Pereira 1993). There is conflicting evidence whether or not these silks are cribellar threads that have been smoothed as a consequence of a division in the cribellum or whether it is composed of individually rippled tubuliform fibres (Eberhard and Pereira 1993; Lopardo and Ramirez 2007).
Web Function
The primary function of spider webs is to catch flying insect prey. Prey capture by a spider web involves three actions — prey interception, stopping, and retention (Eberhard 2014; Zaera et al. 2014), with the different web silks modified in a way within webs to perform one or more of these steps (Blackledge and Hayashi 2006; Blamires et al. 2017).
The architectural components of all two-dimensional orb webs are: (i) the capture surface area, (ii) the width of the spaces between the sticky spirals or mesh size, (iii) the number of radii that traverse the spiral thread, and (iv) the length or pattern of any decorations (stabilimenta) added to the web (Fig. 6.5).

Diagram of a vertical two-dimensional orb web, showing the architectural components: (A) capture surface area, (B) space between the sticky spirals (mesh size), (C) radii traversing the spiral thread and (D) decoration (stabilimenta)
Uloborid Orb Webs
Uloborid orb webs tend to be aligned horizontal to the ground. The reduced webs of Miagrammopes spp. and Hyptiotes spp., however, are aligned vertical to the ground, which might be facilitated by their greater cribellate thread stickiness (Opell 1994a, b). Recently Santos and Gonzaga (2017) described a new Uloborid genus (Uaitemuri) from Southeastern Brazil which also builds vertical orb webs. The horizontal orientation seems to reduce the prey interception rates of webs (Bishop and Connolly 1992) but, considering prey retention is entirely attributable to benign Van der Waals forces (Hawthorn and Opell 2003), it might be utilized out of necessity to retain prey in the web.
Uloborids, such as Philoponella undulata, often aggregate their webs or build webs in close proximity to the webs of other spiders (Finke 1981). When multiple webs are found in close proximity, insects that bounce off or fly through any one web are likely to have their velocity reduced, thereby rendering them more easily caught by a nearby web (Uetz 1989; Yip et al. 2008; Blamires et al. 2010). The more webs in the vicinity, the more likely it is that an insect will eventually be caught. This phenomenon is described as the ‘ricochet effect’ and has been proposed as having an evolutionary benefit associated with spider web aggregations (Uetz 1989).
Deinopid Net-Casting Webs
The Neotropical net-casting spiders Deinopis spinosa and D. longipes (Deinopidae) produces cribellate capture threads, and they have a unique prey-capturing method. They position themselves on vegetation and spin a rectangular-shaped capture net of cribellate silk (Fig. 6.6), releasing the net as an insect moves beneath. The net surrounds the insect, which becomes entangled in the woolly cribellate silk (Opell 1994a; Getty and Coyler 1996). Large flying insects, such as moths, are caught by flicking the cribellate silk backwards (Getty and Coyler 1996). The silk reflects ultraviolet light (Craig et al. 1994) which might be utilized to lure prey toward the net.

The cribellate capture net of Deinopis sp., showing the spider in position so to release the net as an insect moves beneath.
Theridiid Cobwebs
The gumfoot threads of a cobweb extend downward from the tangled retreat (see Fig. 6.4). The gumfoot glue droplets at the base of the thread adhere to prey crawling along the ground, and when the prey struggle, the thread is released from the substrate. Upon release from its pyriform attachment, a gumfoot thread transmits vibrational stimuli toward the cobweb to inform the spider that prey has been captured (Peters 1987). Viscid globules have been found within the tangled retreat of Achaeranea tesselata webs (Barrantes and Weng 2006). Nevertheless, the function of these globules remains unclear. Their small size suggests that they are of little value in prey retention, but this function should not be ruled out (Benjamin et al. 2002; Barrantes and Weng 2006).
Cobweb Function Compared to Orb Webs
In both spiral and gumfoot threads, the glue coalesces under surface forces into droplets that disperse along the axial thread (Opell and Hendricks 2010; Sahni et al. 2011; Blamires et al. 2014a) (Fig. 6.7). The glues of both types of thread are comprised of an aqueous solution of glycoproteins and low molecular weight organic and inorganic salts (Sahni et al. 2014a, b). Gumfoot glue, however, contains additional water-soluble peptides (Hu et al. 2007; Sahni et al. 2011, 2014b). While orb web axial threads are comprised of extensible flagelliform silk (Peters 1987), gumfoot axial threads consist of stiffer major ampullate gland (MA) threads (Sahni et al. 2014b). The mechanical properties of spiral and gumfoot threads differ, most likely as a consequence of the different properties of the respective axial silks (Peters 1987) although differences in the biochemistry of the glues may also play a role (Blamires et al. 2014a).

The gluey silks and underlying axial threads of (a) orb webs and (b) cobwebs, showing that the glues of both are comprised of an aqueous gluey silk, comprising a solution of and low molecular weight organic and inorganic salts and glycoproteins surrounding flagelliform (orb-web spiral threads) or major ampullate (cobweb gumfoot threads)
Araneidae
Vertically Aligned Orb Webs
Orb webs aligned vertical to the ground, such as those spun by Neoscona, Argiope, Nephila, Araneus, and Larinia, appear to be adapted for the capture of high-kinetic-energy prey (Kohler and Vollrath 1995; Harmer et al. 2011; Sensenig et al. 2012). The radial threads are the threads that play the greatest role in stopping prey, as the energy absorption capacity of their silks is an order of magnitude greater than that of the viscid silks (Vollrath 1994). The initial softness and ultimate strength of radial threads provides the inelastic absorption for high-kinetic-energy interception (Denny 1976; Craig 1987; Harmer et al. 2011; Sensenig et al. 2012). The energy absorbed depends where on the web’s surface area, the force applied, and the angle of interception (Craig 1987). Covering the flagelliform threads with aqueous aggregate glue causes the flagelliform silk to plasticise and become highly extensible. This enables the kinetic energy of impacting prey to be imparted onto the web, reducing the probability of the prey bouncing off or flying through the web (Boutry and Blackledge 2013).
The primary function of the sticky spiral threads nonetheless is to retain prey (Sahni et al. 2014b). Some insects, e.g., flies, are better retained by webs with cribellate silk while others, e.g., bugs and beetles, are better retained by webs with aggregate silk (Vollrath 1994). Other insects, e.g., moths, have low adhesion to both (Vollrath 1994; Opell and Schwend 2007). The spiral silk’s ability to maintain tension when stretched and relaxed in rapid succession is a critical feature of its performance. (Zhou and Zhang 2005).
Modified Orb Webs
Neotropical spiders of the genus Scoloderus build an elongated (up to 1 m long but 0.2 m wide) web with the hub located toward the bottom (Eberhard 1975) (Fig. 6.8). This is in contrast with other Araneids that build elongated orb webs, such as the Australian ladder-web spiders, Telaprocera spp. which build elongated webs with the hub positioned at the top of it (Harmer et al. 2011). The reason the hub of Scoloderus webs are at the bottom is because they specialize in capturing moths. When a moth strikes the web it sheds scales in its struggle and rather than breaking free tumbles down the web (Stowe 1978). The extreme elongation of the web means the moths will continue to tumble until they finally adhere to the spirals near the hub (Eisner et al. 1964), thus enabling the spider to rapidly attack its prey (Stowe 1978).

A diagram of the elongated (up to 1 m long) webs of spiders from the Neotropical spider genus Scoloderus (Reproduced from Eberhard (1975))
Cyrtophora citricola is native to Africa and the Middle-East (Lubin 1974) but has been recently documented living in colonies in Columbia, the Dominican Republic, Cuba, and Florida (Levi 1997; Pulido 2002; Alayón 2003). They build a web containing a horizontally aligned prey-catching orb that lacks sticky silks (Levi 1997; Eberhard 1990a; Blamires et al. 2013), onto which it adds a three-dimensional silk barrier structure that extends vertically upward to ~1 m (Lubin 1974, 1980; Berry 1987; Blamires et al. 2013). The spiders position themselves on the underside of the orb and catch the prey that fall into it from above upon striking the barrier structure’s threads.
Much of what we know about the function of their distinctive ‘tent web’ comes from examination of the New World congener Cyrtophora moluccensis (Lubin 1980; Berry 1987; Blamires et al. 2013, 2014b). The architecture of these ‘tent webs’ fundamentally differs to those of closely related orb-web spiders, such as Argiope spp. (Eberhard 1990a; Blackledge et al. 2011). It has been supposed that the barrier structure is used to support the horizontal orb (Berry 1987) but there is no empirical evidence for this function. Most likely it provides protection, and enables the spiders to more effectively capture large-bodied prey, such as moths, which might fly through regular orb webs or fail to adhere to the sticky spirals (Blamires et al. 2013).
Horizontally Aligned Webs
Webs built in horizontal alignment to the ground, such as those of Leucauge spp. are generally not in the flight path of flying insects so intercept less prey (Bishop and Connolly 1992). Accordingly, they appear to be less effective than webs built aligned vertical to the ground (Craig 1987; Blackledge et al. 2011). Nonetheless, horizontally aligned webs appear to have some advantages over vertically aligned web, including protection from inadvertent damage from wind or flying animals, the exploitation of low-lying prey habitats or prey flight patterns not exploited by vertical webs, ability to better cope with the spider’s weight, and freedom from the need to absorb enormous amounts of kinetic energy, since insects are rarely caught in full flight, and tensioning of the web by supercontraction (Craig 1987; Blackledge et al. 2011; Bishop and Connolly 1992; Boutry and Blackledge 2013). There is nevertheless currently no definitive evidence of an advantage for building horizontally aligned webs over vertically aligned webs. In fact, a study that manipulated Leucauge regnyi webs into various alignments found vertically aligned webs to catch fewer prey less efficiently than horizontal webs (Bishop and Connolly 1992).
Web Plasticity
Orb Webs
The architectural components of orb webs are the most readily identifiable and quantifiable among spider webs. Measurements used to quantify orb-web architectural variability (i.e., plasticity) include counting the number of radial threads and any decorations (stabilimenta), measuring the widths and heights of the arc encapsulated by spirals, and calculating parameters such as mesh height and capture area using various formulae (Herberstein and Tso 2000; Blackledge and Gillespie 2002; Tso et al. 2007). Environmental factors that can be ascribed as influential over the relative number and size of orb-web architectural components include: (i) the presence of predators, (ii) ambient temperature, wind, and light levels, and (iii) prey availability, prey types, and prey nutrient value (Heiling and Herberstein 2000; Boutry and Blamires 2013). Change in web architecture has also been documented across ontogeny in orb-web spiders (Hesselberg 2010; Kuntner et al. 2010; Escalante 2013).
Observations of Neotropical Araneids, Uloborids and Tetragnathids were the first to document variability in web building and web architectures across environments and ontogeny (Eberhard 1990a, b). Subsequent studies have expanded to ecribellate orb web spiders, such as Nephila clavipes and Eustala illicita (Hesselberg 2010; Hesselberg and Triana 2010). An orb-web-building spider generally follows the following steps sequentially in building an orb web: (i) exploration, (ii) frames construction, (iii) building of a proto hub, (iv) radii construction, (v) the construction of a spiral scaffold, and (vi) the construction of the spiral threads. Nephila clavipes differs from other orb web spiders in that they do not remove the spiral scaffolding upon placement of the spiral thread (Eberhard 1990b). Variations in behaviours at any one of these steps will result in measurable variation to the web architectural components (Eberhard 1990a; Zschokke and Vollrath 1995). Decorations are added to completed orb webs by Neotropical Cyclosa spp. using detritus, with the size and shape of the detritus decoration dependent upon the availability of prey remains (Gonzaga and Vasconcellos-Neto 2012). Other plastic behaviours associated with web building include variability in predatory behaviours. For instance, the order, frequency, and timing of prey-attacking behaviours can vary across spider ontogeny (Castanho and Oliveira 1997; Japyassu and Caires 2008).
Web plasticity may influence web structural variation across the spider phylogeny (Blackledge et al. 2009, 2011; Kuntner et al. 2010). Accordingly, some clades might exhibit relatively minor variability in web forms across species, environments, and ontogeny. While others, e.g., Theridiidae, might exhibit exceptional variability in web forms across species, environments, and ontogeny (Forster and Forster 1985; Eberhard et al. 2008).
Diet-Induced Web Plasticity
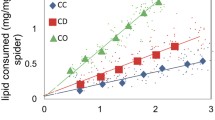
Satiated spiders build webs with smaller capture areas than starved spiders (Sandoval 1994; Herberstein et al. 2000b). The capture area of a web is often associated with a reduction in mesh height (Sandoval 1994; Tso et al. 2007). If the spider expects to feed on a homogeneous prey type it will not change mesh height, but it will invest in longer spiral threads to accommodate the web size change (Sandoval 1994). Spiders might also alter mesh height to tune the web for catching a specific type of prey (Sandoval 1994; Blackledge and Zevenbergen 2006; Blamires et al. 2011). Whether spiders adjust their web architectures under different diets because of constraints placed on the production of certain silks or to adjust the performance of the web is difficult to ascertain, because web and silk properties strongly co-vary across diets (Tso et al. 2007; Blamires et al. 2015, 2016).
Cobwebs
Cobwebs are the specialized three-dimensional webs of spiders of Theridiidae (Blackledge et al. 2005; Eberhard et al. 2008; Boutry and Blamires 2013). A number of studies have examined cobweb architectural plasticity (Blackledge and Zevenbergen 2007; Salomon 2007; Zevenbergen et al. 2008). As with orb webs, spatial constraints, hunger, and the type of prey consumed influence cobweb architectures (Jorger and Eberhard 2006; Blackledge and Zevenbergen 2007; Zevenbergen et al. 2008). For instance, satiated Lactodectus hesperus build webs with fewer gumfooted threads, with each thread containing fewer sticky droplets, than do those of starved L. hesperus (Blackledge and Zevenbergen 2007). The sizes of the individual gumfoot droplets nevertheless do not differ between satiated and starved spiders (Blamires et al. 2014a). Temperature and light influence cobweb architecture, most likely by affecting the spider’s ability to spin silk or to build webs (Lubin et al. 1991; Blackledge and Zevenbergen 2007; Zevenbergen et al. 2008).
Plasticity in Other Webs
The Pholcid Physocyclus globosus builds a finely meshed, irregular, domed sheet web below a veiled tangle web. While the web form does not differ substantially, the web-building behaviours vary significantly across ontogeny in this species. Juveniles lay 91% of their threads within the sheet chamber, while adult females lay 55% and adult males 41%; adult males more often utilize exploration threads (Escalante 2013). Variations in behaviour by different individuals have been shown to result in variable web architectures (Madrigal-Brenes and Barrantes 2009). Female web-building wolf spiders of the genus Aglaocetenus may abandon web building at certain times of year or in certain locations, or vary their web building, i.e., shorten the attachment threads or deposit more fine threads without any apparent function, in the presence of male conspecifics or other spiders, e.g., Uloborids (González et al. 2014).
Conclusion
The diversity of spider webs and web-building spiders from the Neotropics has been well described owing to over a century of detailed observations. We have given an overview of some examples of the multitude of different web forms found in the Neotropics, highlighting some striking web forms and their structural and functional variability. Since the Neotropics contains up to half of all genera of orb-web spiders, the spider web forms and the various modifications thereof described herein will be a close representation of overall spider web diversity.
A spider’s web is an extended phenotype depicting its foraging, mating, and defensive strategies, and physiological status. Spiders exhibiting web plasticity can continue to build functional webs across highly variable environments. The seasonality and geography of the Neotropical region, and the array of spider predators and prey that can be found there, have undoubtedly shaped the unique diversity of the region’s spiders.
References
Agnarsson I (2002) Sharing a web: on the relation of sociality and kleptoparasitism in theridiid spiders (Theridiidae, Araneae). J Arachnol 30:181–188
Agnarsson I (2003a) The phylogenetic placement and circumscription of the genus Synotaxus (Araneae: Synotaxidae), a new species from Guyana, and notes on theridioid phylogeny. Invertebr Syst 17:719–734
Agnarsson I (2003b) Spider webs as habitat patches — the distribution of kleptoparasites (Argyrodes, Theridiidae) among host webs (Nephila, Tetragnathidae). J Arachnol 31:344–349
Agnarsson I (2004) Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea, Theridiidae). Zool J Linnean Soc 141:447–626
Agnarsson I (2011) Habitat patch size and isolation as predictors of occupancy and number of argyrodine spider kleptoparasites in Nephila webs. Naturwissenschaften 98:163–167
Agnarsson I, Avilés L, Coddington JA, Maddison WP (2006) Sociality in Theridiid spiders: repeated origins of an evolutionary dead end. Evolution 60:2342–2351
Agnarsson I, Maddison WP, Avilés L (2007) The phylogeny of the social Anelosimus spiders (Araneae: Theridiidae) inferred from six molecular loci and morphology. Mol Phylogenet Evol 43:833–851
Alayón GG (2003) Cyrtophora citricola (Araneae: Araneidae), registro neuvo de araña para Cuba. Cocuyo 13:14
Ayoub NA, Garb JE, Kuelbs A, Hayashi CY (2013) Ancient properties of spider silks revealed by the complete gene sequence of the prey-wrapping silk protein (AcSp1). Mol Biol Evol 30:589–601
Baldissera R, Ganade G, Brescovit AD, Hartz SM (2008) Landscape mosaic of Araucaria forest and forest monoculture influencing understory spider assemblages in southern Brazil. Austral Ecol 33:45–54
Barrantes G, Eberhard WG (2007) The evolution of prey-wrapping behaviour in spiders. J Nat Hist 41:1631–1658
Barrantes G, Weng JL (2006) Viscid globules in webs of the spider Achaearanea tesselata (Araneae: Theridiidae). J Arachnol 34:480–482
Benjamin SP, Duggelin M, Zschokke S (2002) Fine structure of sheet-webs of Linyphia triangularis (Clerck) and Microlinyphia pusilla (Sundevall), with remarks on the presence of viscid silk. Acta Zool 83:49–59
Benjamin SP, Zschokke S (2002) Untangling the tangle web: web construction behaviours of the gumfooted spider Steatoda triangulos and comments on phylogenetic implications (Araneae: Theridiidae). J Insect Behav 15:791–809
Berry JW (1987) Notes on the life history and behavior of the communal spider Cyrtophora moluccensis (Doleschall) (Araneae, Araneidae) in Yap, Carolina Islands. J Arachnol 15:309–319
Bishop L, Connolly SR (1992) Web orientation, thermoregulation, and prey capture efficiency in a tropical forest spider. J Arachnol 20:173–178
Blackledge TA, Gillespie RG (2002) Estimation of capture areas of spider webs in relation to web asymmetry. J Arachnol 30:70–77
Blackledge TA, Hayashi CY (2006) Silken toolkits: biomechanics of silk fibers spun by the orb web spider Argiope argentata (Fabricius 1775). J Exp Biol 209:2452–2461
Blackledge TA, Zevenbergen JM (2006) Mesh width influences prey retention in spider orb webs. Ethology 112:1194–1201
Blackledge TA, Zevenbergen JM (2007) Condition-dependent spider web architecture in the western black widow, Latrodectus hesperus. Anim Behav 73:855–864
Blackledge TA, Summers AP, Hayashi CY (2005) Gumfooted lines in black widow cobwebs and the mechanical properties of spider capture silk. Zoology 108:41–46
Blackledge TA, Scharff N, Coddington JA, Szuts T, Wenzel JW, Hayashi CY, Agnarsson I (2009) Reconstructing web evolution and spider diversification in the molecular era. Proc Natl Acad Sci USA 106:5229–5234
Blackledge TA, Kuntner M, Agnarsson I (2011) The form and function of spider orb webs: evolution from silk to ecosystems. Adv Insect Physiol 41:175–262
Blackledge TA, Perez-Riguiero J, Plaza GR, Perea B, Navarro A, Guinea GV, Elices M (2012) Sequential origin in the high performance properties of orb spider dragline silk. Sci Rep 2:782
Blamires SJ, Lee YH, Chang CM, Lin IT, Cheg JA, Lin TY, Tso IM (2010) Multiple structures interactively influence prey capture efficiency in spider orb webs. Anim Behav 80:947–953
Blamires SJ, Chao IC, Liao CP, Tso IM (2011) Multiple prey cues induce foraging flexibility in a trap-building predator. Anim Behav 81:955–961
Blamires SJ, Hou C, Chen LF, Liao CP, Tso IM (2013) Three-dimensional barricading of a predatory trap reduces predation and enhances prey capture. Behav Ecol Sociobiol 67:709–714
Blamires SJ, Sahni V, Dhinojwala A, Blackledge TA, Tso IM (2014a) Nutrient deprivation induces property variations in spider gluey silk. PLoS One 9:e88487
Blamires SJ, Hou C, Chen LF, Liao CP, Tso IM (2014b) A predator’s body coloration enhances its foraging profitability by day and night. Behav Ecol Sociobiol 68:1253–1260
Blamires SJ, Piorkowski D, Chuang A, Tseng YH, Toft S, Tso IM (2015) Can differential nutrient extraction explain property variations in a predatory trap? R Soc Open Sci 2:140479
Blamires SJ, Tseng YH, Wu CL, Toft S, Raubenheimer D, Tso IM (2016) Spider silk and web performance landscapes across nutrient space. Sci Rep 6:26383
Blamires SJ, Blackledge TA, Tso IM (2017) Physico-chemical property variation in spider silks: ecology, evolution and synthetic production. Annu Rev Entomol 62:443–460
Blasingame E, Tuton-Blasingame T, Falick AM, Zhao L, Fong J, Vaidyanathan V, Vispers A, Guerts P, Hu XW, LaMattina C, Viera C (2009) Pyriform spidroin 1, a novel member of the silk gene family that anchors dragline silk fibers in attachment discs of the black widow spider, Latrodectus hesperus. J Biol Chem 284:29097–29108
Bonaldo AB, Marques MAL, Pinto-da-Rocha R, Gardner T (2007) Species richness and community structure of arboreal spider assemblages in fragments of three vegetational types at Banhado Grande wet plain, Gravataí River, Rio Grande do Sul, Brazil. Iheringia 97:143–151
Bond JE, Garrison NL, Hamilton CA, Godwin RL, Hedin M, Agnarsson I (2014) Phylogenomics resolves a spider backbone phylogeny and rejects a prevailing paradigm for orb web evolution. Curr Biol 24:1765–1771
Boutry C, Blackledge TA (2010) Evolution of supercontraction in spider silk: structure–function relationship from tarantulas to orb-weavers. J Exp Biol 213:3505–3514
Boutry C, Blackledge TA (2013) Wet webs work better: humidity, supercontraction and the performance of spider orb webs. J Exp Biol 216:3606–3610
Boutry C, Blamires SJ (2013) Plasticity in spider webs and silk: an overview of current evidence. In: Santerre M (ed) Spiders: morphology behavior and geographic distribution. Nova, New York, pp 1–46
Briceno RD, Eberhard WG (2011) The hub as a launching platform: rapid movements of the spider Leucauge mariana (Araneae: Tetragnathidae) as it turns to attack prey. J Arachnol 39:102–112
Brooks DM (2012) Birds caught in spider webs: a synthesis of patterns. Wilson J Ornithol 124:345–353
Castanho LM, Oliveira PS (1997) Biology and behaviour of the neotropical ant-mimicking spider Aphantochilus rogersi (Araneae: Aphantochilidae): nesting, maternal care and ontogeny of ant-hunting techniques. J Zool 242:643–650
Cheng RC, Yang EC, Lin EP, Herberstein ME, Tso IM (2010) Insect form vision as one potential shaping force of spider web decoration design. J Exp Biol 213:759–768
Choresh O, Bayarmagnai B, Lewis RV (2009) Spider web glue: two proteins expressed from opposite strands of the same DNA sequence. Biomacromolecules 10:2852–2856
Clarke TH, Garb JE, Hayashi VY, Arensberger P, Ayoub NA (2015) Spider transcriptomes identify ancient large-scale gene duplication event potentially important in silk gland evolution. Genome Biol Evol 7:1856–1870
Colgin MA, Lewis RV (1998) Spider minor ampullate silk proteins contain new repetitive sequences and highly conserved non-silk like “spacer regions”. Protein Sci 7:667–672
Collin MA, Clarke TH, Ayoub NA, Hayashi CY (2016) Evidence from multiple species that spider silk glue component ASG2 is a spidroin. Sci Rep 6:21589
Craig CL (1986) Orb-web visibility: the influence of insect flight behaviour and visual physiology on the evolution of web designs within the Araneoidea. Anim Behav 34:54–68
Craig CL (1987) The ecological and evolutionary interdependence between web architecture and web silk spun by orb web weaving spiders. Biol J Linn Soc 30:135–162
Craig CL (2003) Spiderwebs and silk: tracing evolution from molecules to genes to phenotypes. Oxford University Press, Oxford
Craig CL, Bernard GD, Coddington JA (1994) Evolutionary shifts in the spectral properties of spider silks. Evolution 48:287–296
Denny MW (1976) The physical properties of spider’s silk and their role in the design of orb webs. J Exp Biol 65:483–506
Dimitrov D, Benavides LR, Arnedo MA, Giribet G, Griswold CE, Scharff N, Hormiga G (2017) Rounding up the usual suspects: a standard target-gene approach for resolving the interfamilial phylogenetic relationships of ecribellate orb-weaving spiders with a new family–rank classification (Araneae, Araneoidea). Cladistics 33:221–250. https://doi.org/10.1111/cla.12165
Eberhard WG (1972) The web of Uloborus diversus (Araneae : Uloboridae). J Zool 166:417–465
Eberhard WG (1973) Stabilimenta on the webs of Uloborus diversus (Araneae: Uloboridae) and other spiders. J Zool 171:367–384
Eberhard WG (1975) The ‘inverted ladder’ orb web of Scoloderus sp. and the intermediate orb of Eustala (?) sp. Araneae : Araneidae. J Nat Hist 9:93–106
Eberhard WG (1977) Rectangular orb-webs of Synotaxus (Araneae: Theridiidae). J Nat Hist 11:501–507
Eberhard WG (1987) Effects of gravity on temporary spiral construction by Leucauge mariana (Araneae: Araneidae). J Ethol 5:29–36
Eberhard WG (1988a) Memory of distances and directions moved as cues during temporary spiral construction in the spider Leucauge mariana (A., Araneidae). J Insect Behav 1:51–66
Eberhard WG (1988b) Behavioral flexibility in orb web construction: effects of supplies of different silk glands and spider size and weight. J Arachnol 16:295–302
Eberhard WG (1990a) Function and phylogeny of spider webs. Annu Rev Ecol Syst 21:341–372
Eberhard WG (1990b) Early stages of orb construction by Philoponella vicina, Leucauge mariana, and Nephila clavipes (Araneae, Uloboridae and Tetragnathidae), and their phylogenetic implications. J Arachnol 18:205–234
Eberhard WG (1995) The web and building behavior of Synotaxus ecuadorensis (Araneae, Synotaxidae). J Arachnol 23:25–30
Eberhard WG (2003) Substitution of silk stabilimenta for egg sacs by Allocyclosa bifurca (Araneae: Araneidae) suggests that silk stabilimenta function as camouflage devices. Behaviour 140:847–868
Eberhard WG (2008) Araneus expletus (Araneae, Araneidae): another stabilimentum that does not function to attract prey. J Arachnol 36:191–194
Eberhard WG (2014) A new view of orb webs: multiple trap designs in a single structure. Biol J Linn Soc 111:437–449
Eberhard WG, Barrantes G (2015) Cues guiding uloborid construction behavior support orb web monophyly. J Arachnol 43:371–387
Eberhard WG, Pereira F (1993) Ultrastructire of cribellate silk of nine species in eight families and possible taxonomic implications (Araneae: Amaurobiidae, Deinopidae, Desidae, Dictynidae, Filistatidae, Hypochilidae, Stiphidiidae, Tengellidae). J Arachnol 21:161–174
Eberhard WG, Agnarsson I, Levi HW (2008) Web forms and the phylogeny of theridiid spiders (Araneae: Theridiidae): chaos from order. Syst Biodivers 6:1–61
Edmonds DT, Vollrath F (1992) The contribution of atmospheric water vapour to the formation and efficiency of a spider’s capture web. Proc R Soci B 248:145–148
Eisner T, Alsop R, Ettershank G (1964) Adhesiveness of spider silk. Science 146:1058–1061
Elgar MA (1994) Experimental evidence of a mutualistic association between two web-building spiders. J Anim Ecol 63:880–886
Escalante I (2013) Ontogenetic and sexual differences in exploration and web construction in the spider Physocyclus globosus (Araneae: Pholcidae). Arachnology 16:61–68
Fernández R, Hormiga G, Giribet G (2014) Phylogenomic analysis of spiders reveals nonmonophyly of orb weavers. Curr Biol 24:1772–1777
Finke OM (1981) An association between two Neotropical spiders (Araneae: Uloboridae and Tengellidae). Biotropica 13:301–307
Forster LM, Forster RR (1985) A derivative of the orb web and its evolutionary significance. N Z J Zool 12:455–465
Forster RR, Platnick NI, Coddington JA (1990) A proposal and review of the spider family Synotaxidae (Araneae, Araneoidea), with notes on theridiid interrelationships. Bull Am Mus Nat Hist 193:1–116
Garb JE, DiMauro T, Lewis RV, Hayashi CY (2007) Expansion and intragenic homogenization of spider silk genes since the Triassic: evidence from Mygalomorphae (tarantulas and their kin) spidroins. Mol Biol Evol 24:2454–2464
Garb JE, Ayoub NA, Hayashi CY (2010) Untangling spider silk evolution with spidroin terminal domains. BMC Evol Biol 10:243
Getty RM, Coyler FA (1996) Observations on prey capture and anti-predator behaviors of ogre-faced spiders (Dienopis) in southern Costa Rica (Araneae, Dienopidae). J Arachnol 24:93–100
Gnesa E, Hsia Y, Yarger JL, Weber W, Lin-Cereghino J, Lin-Cereghino G, Tang S, Agari K, Viera C (2012) Conserved C-terminal domain of spider tubuliform spidroin 1 contributes to extensibility in synthetic fibers. Biomacromolecules 13:304–312
Gonzaga MO, Vasconcellos-Neto J (2005) Testing the functions of detritus stabilimenta in webs of Cyclosa fililineata and Cyclosa morretes (Araneae : Araneidae): do they attract prey or reduce the risk of predation? Ethology 111:479–491
Gonzaga MO, Vasconcellos-Neto J (2012) Variation in the stabilimenta of Cyclosa fililineata Hingston, 1932, and Cyclosa morretes Levi, 1999 (Araneae: Araneidae), in southeastern Brazil. Psyche 2012:396594
Gonzaga MO, Leiner NO, Santos AJ (2006) On the stick cobwebs of two theridiid spiders (Araneae: Theridiidae). J Nat Hist 40:293–306
González M, Costa FG, Peretti AV (2014) Strong phenological differences between two populations of a Neotropical funnel-web wolf spider. J Nat Hist 48:2183–2197
González M, Peretti AV, Costa FG (2015a) Reproductive isolation between two populations of Aglaoctenus lagotis, a funnel-web wolf spider. Biol J Linn Soc 114:646–658
González M, Costa FG, Peretti AV (2015b) Funnel-web construction and estimated immune costs in Aglaoctenus lagotis (Araneae: Lycosidae). J Arachnol 43:158–167
Griswold CE, Coddington JA, Hormiga G, Scharff N (1998) Phylogeny of the orb-web building spiders (Araneae, Orbiculariae: Deinopoidea, Araneoidea). Zool J Linnean Soc 123:1–99
Harmer ATM, Blackledge TA, Madin JS, Herberstein ME (2011) High-performance spider webs: integrating biomechanics, ecology and behaviour. J R Soc Interface 8:457–471
Hawthorn AC, Opell BD (2003) Van der Waals and hygroscopic forces of adhesion generated by spider capture threads. J Exp Biol 206:3905–3911
Hayashi CY, Lewis RV (1998) Evidence from flagelliform silk cDNA for the structural basis of elasticity and modular nature of spider silks. J Mol Biol 275:773–784
Hayashi CY, Blackledge TA, Lewis RV (2004) Molecular and mechanical characterization of aciniform silk: uniformity of iterated sequence modules in a novel member of the spider silk fibroin gene family. Mol Biol Evol 21:1950–1959
Heiling AM, Herberstein ME (2000) Interpretations of orb-web variability: a review of past and current ideas. Ekologia 19:97–106
Hénaut Y, Garcia-Ballinas JA, Alauzet C (2006) Variations in web construction in Leucauge venusta (Araneae, Tetragnathidae). J Arachnol 34:234–240
Herberstein ME, Hebets EA (2013) Behaviour: why are spiders good models for research? In: Penny D (ed) Spider research in the 21st century: trends and perspectives. SIRI Scientific Publishing, Manchester, pp 230–250
Herberstein ME, Tso IM (2000) Evaluation of formulae to estimate the capture area and mesh height of orb webs (Araneoidea, Araneae). J Arachnol 28:180–184
Herberstein ME, Craig CL, Coddington JA, Elgar MA (2000a) The functional significance of silk decorations of orb-web spiders: a critical review of the empirical evidence. Biol Rev 75:649–669
Herberstein ME, Craig CL, Elgar MA (2000b) Foraging strategies and feeding regimes: web and decoration investment in Argiope keyserlingi Karsch (Araneae: Araneidae). Evol Ecol Res 2:69–80
Hesselberg T (2010) Ontogenetic changes in web design in two orb-web spiders. Ethology 116:535–545
Hesselberg T, Triana E (2010) The web of the acacia orb-spider Eustala illicita (Araneae: Araneidae) with notes on its natural history. J Arachnol 38:21–26
Higgins LE (1992) Developmental changes in barrier web structure under different levels of predation risk in Nephila clavipes (Araneae: Tetragnathidae). J Insect Behav 5:635–655
Higgins LE, Buskirk RE (1998) Spider-web kleptoparasites as a model for studying producer-consumer interactions. Behav Ecol 9:384–387
Hinman MB, Lewis RV (1992) Isolation of a clone encoding a second dragline silk fibroin: Nephila clavipes dragline silk is a two-protein fiber. J Biol Chem 267:19320–19324
Hormiga GC, Griswold E (2014) Systematics, phylogeny, and evolution of orb-weaving spiders. Annu Rev Entomol 59:487–512
Hsia Y, Gnesa E, Jeffrey F, Tang S, Viera C (2011) Spider silk composites and applications. In: Cuppoletti J (ed) Metal ceramic and polymeric composites for various uses. Intech, Rijeka, pp 303–324
Hu XW, Lawrence BD, Kohler K, Falick AM, Moore AMF, McMullen E, Jones PR, Viera C (2005) Araneoid egg case silk: a fibroin with novel ensemble repeat units from the black widow spider, Latrodectus hesperus. Biochemistry 44:10020–10027
Hu XW, Yuan J, Wang X, Vasanthavada K, Falick AM, Jones PR, La Mattina C, Viera C (2007) Analysis of aqueous glue coating proteins on the silk fibers of the cob weaver, Latrodectus hesperus. Biochemistry 46:3294–3303
Japyassu HF, Caires RA (2008) Hunting tactics in a cobweb spider (Araneae-Theridiidae) and the evolution of behavioral plasticity. J Insect Behav 21:258–284
Jorger KM, Eberhard WG (2006) Web construction and modification by Achaearanea tesselata (Araeae, Theridiidae). J Arachnol 34:511–523
Kohler T, Vollrath F (1995) Thread biomechanics in the two orb-weaving spiders Araneus diadematus (Araneae, Araneidae) and Uloborus walckenaerius (Araneae, Uloboridae). J Exp Zool 271:1–17
Kuntner M, Kralj-Fiser S, Gregorič M (2010) Ladder webs in orb-web spiders: ontogenetic and evolutionary patterns in Nephilidae. Biol J Linn Soc 99:849–866
La Mattina C, Reza R, Hu XW, Falick AM, Vasanthavada K, McNary S, Yee R, Viera C (2008) Spider minor ampullate silk proteins are constituents of prey wrapping silk in the cob weaver Latrodectus hesperus. Biochemistry 47:4692–4700
Lehtinen PT (1967) Classification of the cribellate spiders and some allied families, with notes on the evolution of the suborder Araneomorpha. Ann Zool Fenn 4:199–468
Levi HW (1997) The American orb weavers of the genera Mecynogea, Manogea, Kapogea, and Cyrtophora (Araneae: Araneidae). Bull Mus Comp Zool 155:215–255
Lewis RV (1992) Spider silk: the unraveling of a mystery. Acc Chem Res 25:392–398
Lopardo L, Ramirez MJ (2007) The combing of cribellar silk by the prithine Misionella mendensis, with notes on other Filistatid spiders (Araneae: Filistatidae). Am Mus Novit 3563:1–14
Lubin YD (1974) Adaptative advantages and the evolution of colony formation in Cyrtophora (Araneae, Araneidae). Zool J Linnean Soc 54:321–339
Lubin YD (1980) The predatory behavior of Crytophora (Araneae: Araneidae). J Arachnol 8:159–185
Lubin YD (1986) Web building and prey capture in the Ulobidae. In: Shear WA (ed) Spiders: Webs Behavior and Evolution. Stanford University Press, Stanford, pp 132–171
Lubin YD, Eberhard WG, Montgomery GG (1978) Webs of Miagrammopes (Araneae , Uloboridae) in the Neotropics. Psyche 85:1–23
Lubin Y, Kotzman M, Ellner S (1991) Ontogenetic and seasonal changes in webs and websites of a desert widow spider. J Arachnol 19:40–48
Macrini CMT, Peres EA, Solferini VN (2015) Cryptic diversity of Agloactenus lagotis (Araneae, Lycosidae) in the Brazilian Atlantic rainforest: evidence from microsatellite and mitochondrial DNA sequence data. J Appl Biol Biotechol 3:009–014
Madrigal-Brenes R, Barrantes G (2009) Construction and function of the web of Tidarren sisyphoides (Araneae: Theridiidae). J Arachnol 37:306–311
Magalhaes I, Santos AJ (2012) Phylogenetic analysis of Micrathena and Chaetacis spiders (Araneae: Araneidae) reveals multiple origins of extreme sexual size dimorphism and long abdominal spines. Zool J Linn Soc-Lond 166:14–53
Messas YF, Souza HS, Gonzaga MO, Vasconcellos-Neto J (2014) Spatial distribution and substrate selection by the orb-weaver spider Eustala perfida Mello-Leitão, 1947 (Araneae: Araneidae). J Nat Hist 48:2645–2660
Moura RR, Gonzaga MO (2007) Temporal variation in size-assortative mating and male mate choice in a spider with amphisexual care. Sci Nat 104:28. https://doi.org/10.1007/s00114-017-1448-6
Moura RR, Leal LC, Kloss TG (2016) Does nutritional status constrain adoption of more costly and less risky behaviour in an Amazonian shelter-building spider? J Nat Hist 50:2829–2837
Moura RR, Vasconcellos-Neto J, Gonzaga MO (2017) Extended male care in Manogea porracea (Araneae: Araneidae): the exceptional case of a spider with amphisexual care. Anim Behav 123:1–9
Murphy NP, Framenau VW, Donnellan SC, Harvey MS, Park YC, Austin AD (2006) Phylogenetic reconstruction of the wolf spiders (Araneae: Lycosidae) using sequences from the 12S rRNA, 28S rRNA, and NADH1 genes: implications for classification, biogeography, and the evolution of web building behavior. Mol Phylogenet Evol 38:583–602
Nascimento AL, Gonzaga MO (2015) Maternal defensive behaviors of Uloborus sp. (Aranea, Uloboridae): behavioral repertoire and influence of clutch size and female size on female aggressiveness. Acta Ethol 19:33–41
Nentwig W, Christenson TE (1986) Natural history of the non-solitary sheetweaving spider Anelosimus jucundus (Araneae: Theridiidae). Zool J Linnean Soc 87:27–35
Nyffeler M, Knornschild M (2013) Bat predation by spiders. PLoS One 8:e58120
Opell BD (1982) Post-hatching development and web production of Hyptiotes cavatus (Hentz) (Araneae, Uloboridae). J Arachnol 10:185–191
Opell BD (1987) The new species Philoponella herediae and its modified orb-web (Araneae , Uloboridae). J Arachnol 15:59–63
Opell BD (1994a) Factors governing the stickiness of cribellar prey capture threads in the spider family Uloboridae. J Morphol 222:111–119
Opell BD (1994b) Increased stickiness of prey capture threads accompanying web reduction in the spider family Uloboridae. Funct Ecol 8:85–90
Opell BD (1996) Functional similarities of spider webs with diverse architectures. Am Nat 148:630–648
Opell BD (1998) Economics of spider orb-webs: the benefits of producing adhesive capture threads and of recycling silk. Funct Ecol 12:613–624
Opell BD, Bond JE (2000) Capture thread extensibility of orb-weaving spiders: testing punctuated and associative explanations of character evolution. Biol J Linn Soc 70:107–120
Opell BD, Eberhard WG (1984) Resting postures of orb-weaving uloborid spiders (Araneae,Uloboridae). J Arachnol 11:369–376
Opell BD, Hendricks ML (2010) The role of granules within viscous capture threads of orb-weaving spiders. J Exp Biol 213:339–346
Opell BD, Schwend HS (2007) The effect of insect surface features on the adhesion of viscous capture threads spun by orb-weaving spiders. J Exp Biol 210:2352–2360
Osaki S (1996) Spider silk as a mechanical lifeline. Nature 384:419
Peng P, Blamires SJ, Agnarsson I, Lin HC, Tso IM (2013) A colour-mediated mutualism between two arthropod predators. Curr Biol 23:172–176
Perry DJ, Bittencourt D, Liberels-Stilberg J, Rech EL, Lewis RV (2010) Pyriform spider silk sequences reveal unique repetitive elements. Biomacromolecules 11:3000–3006
Peters HM (1938) Ober das Netz der Dreieckspinne, Hyptiotes paradoxus. Zool Anz 121:49–59
Peters HW (1987) Fine structure and function of capture threads. In: Nentwig W (ed) Ecophysiology of spiders. Springer-Verlag, Berlin, pp 187–202
Piacentini L (2011) Three new species and new records in the wolf spider subfamily Sosippinae from Argentina (Araneae: Lycosidae). Zootaxa 3018:27–49
Pickard-Cambridge FO (1904) Arachnida — Araneida and Opiliones. In: Biologia Centralia-Americana. Zoology, London, pp 465–560
Piorkowski D, Blackledge TA (2017) Punctuated evolution of viscid silk in orb web spiders supported by mechanical behavior of wet cribellate silk. Sci Nat 6: doi:10.1007/s00114-017-1489-x
Prosdocimi F, Bittencourt D, Rodrigues da Silva F, Kirst M, Motta PC, Rech EL (2011) Spinning gland transcriptomics from two main clades of spiders (order: Araneae) — insights on their molecular, anatomical and behavioral evolution. PLoS One 6:e21634
Pulido FJI (2002) Manejo de la araña del Mediterraneo o araña parda enredadora. Instituto Colombiano Agropecuario report. 5p
Rito KF, Hanashiro FTT, Peixoto EC, Gonzaga MO (2016) Optimal foraging or predator avoidance?: why does the Amazon spider Hingstepeira foliscens (Araneae: Araneidae) adopt alternative foraging behaviors? Zoologia 33, e20150147
Sahni V, Blackledge TA, Dhinojwala A (2011) Changes in the adhesive properties of spider aggregate glue during the evolution of cobwebs. Sci Rep 1:41
Sahni V, Dhinojwala A, Opell BD, Blackledge TA (2014a) Prey capture adhesives produced by orb-weaving spiders. In: Asakura T, Miller T (eds) Biotechnology of silk. Springer, Dordrecht, pp 203–217
Sahni V, Miyoshi T, Chen K, Jain D, Blamires SJ, Blackledge TA, Dhinojwala A (2014b) Direct solvation of glycoproteins by salts in spider silk glues increases adhesion and helps to explain the evolution of modern spider orb webs. Biomacromolecules 15:1225–1232
Salomon M (2007) Western black widow spiders express state-dependent web-building strategies tailored to the presence of neighbours. Anim Behav 73:865–875
Salomon M, Sponarski C, Larocque A, Aviles L (2010) Social organization of the colonial spider Leucauge sp in the Neotropics: vertical stratification within colonies. J Arachnol 38:446–451
Sandoval CP (1994) Plasticity in web design in the spider Parawixia bistriata: a response to variable prey type. Funct Ecol 8:701–707
Santos AJ (2007) A revision of the Neotropical nursery-web spider genus Architis (Araneae: Pisauridae). Zootaxa 1578:1–40
Santos AJ, Brescovit AD (2001) A revision of the South American spider genus Aglaoctenus Tullgren, 1905 (Araneae, Lycosidae, Sosippinae). Andrias 15:75–90
Santos AJ, Gonzaga MO (2017) Systematics and natural history of Uaitemuri, a new genus of the orb-weaving spider family Uloboridae (Araneae: Deinopoidea) from south-eastern Brazil. Zool J Linnean Soc 180:155–174
Sensenig A, Lorentz KA, Kelly SP, Blackledge TA (2012) Spider orb webs rely on radial threads to absorb prey kinetic energy. J R Soc Interface 9:1880–1891
Silva ELC, Lise A, Carico JE (2008) Revision of the Neotropical spider genus Enna (Araneae, Lycosoidea, Trechaleidae). J Arachnol 36:76–110
Souza HS, Messas YF, Gonzaga MO, Vasconcellos-Neto J (2015) Substrate selection and spatial segregation by two congeneric species of Eustala (Araneae: Araneidae) in southeastern Brazil. J Arachnol 43:59–66
Stefani V, Del-Claro K (2015) The effects of forest fragmentation on the population ecology and natural history of a funnel-web spider. J Nat Hist 49:211–223
Stefani V, Del-Claro K, Silva LA, Guimarães B, Tizo-Pedroso E (2001) Mating behaviour and maternal care in the tropical savanna funnel-web spider Aglaoctenus lagotis Holmberg (Araneae: Lycosidae). J Nat Hist 45:1119–1129
Stowe MK (1978) Observations of two nocturnal orbweavers that build specialized webs: Scoloderus cordatus and Wixia ectypa (Araneae: Araneidae). J Arachnol 6:141–146
Su YC, Smith D (2014) Evolution of host use, group-living and foraging behaviours in kleptoparasitic spiders: molecular phylogeny of the Argyrodinae (Araneae : Theridiidae). Invertebr Syst 28:415–431
Tian M, Lewis RV (2005) Molecular characterization and evolutionary study of spider tubuliform (eggcase) silk protein. Biochemistry 44:8006–8012
Townley MA, Tillinghast EK (2013) Aggregate silk gland secretions of Araneoid spiders. In: Nentwig W (ed) Spider ecophysiology. Springer, Berlin, pp 283–302
Townley MA, Tillinghast EK, Neefus CD (2006) Changes in composition of spider orb web sticky droplets with starvation and web removal and synthesis of sticky droplet compounds. J Exp Biol 209:1463–1486
Tso IM, Liao CP, Huang RP, Yang EC (2006) Function of being colorful in web spiders: attracting prey or camouflaging oneself? Behav Ecol 17:606–613
Tso IM, Chiang SY, Blackledge TA (2007) Does the giant wood spider Nephila pilipes respond to prey variation by altering web or silk properties? Ethology 113:324–333
Turnbull AL (1973) Ecology of the true spiders (Araneomorphae). Annu Rev Entomol 18:305–348
Uetz GW (1989) The ‘ricochet effect’ and prey capture in colonial spiders. Oecologia 81:154–159
Vasconcellos-Neto J, Lewinsohn TM (1984) Discrimination and release of unpalatable butterflies by Nephila clavipes, a Neotropical orb-weaving spider. Ecol Entomol 9:337–344
Vollrath F (1979) Behaviour of the kleptoparasitic spider Argyrodes elevatus (Araneae, theridiidae). Anim Behav 27:515–521
Vollrath F (1994) General properties of some spider silks. In: Kapaln DL, Adams WW, Farmer B, Viney C (eds) Silk polymers: materials science and biotechnology. American Chemical Society, Washington, D.C, pp 17–28
Vollrath F, Knight DP (2001) Liquid crystalline spinning of spider silk. Nature 410:541–548
Vollrath F, Fairbrother WJ, Williams RJP, Tillinghast EK, Bernstein DT, Gallagher KS, Townley MA (1990) Compounds in the droplets of the orb spider’s viscid spiral. Nature 345:526–528
Whitehouse MEA, Lubin YD (2005) The functions of societies and the evolution of group living: spider societies as a test case. Biol Rev 80:347–361
Wiehle H (1927) Beitrage zur Kenntnis Uloboriden. Z Morph Okol Tiere 8:468–537
Wolff JO, Grawe I, Writh M, Karstedt A, Gorb SN (2015) Spider’s super-glue: thread anchors are composite adhesives with synergistic hierarchical organization. Soft Matter 11:2394–2403
World Spider Catalog (2016) Natural History Museum Bern, version 16.5. Available from http://research.amnh.org/entomology/spiders/catalog/index. Accessed 28 Mar 2017
Xavier GM, Moura RR, Gonzaga MO (2017) Orb web architecture of Wixia abdominalis O. Pickard-Cambridge, 1882 (Araneae: Araneidae): intra-orb variation of web components. J Arachnol (in press) 45: 160–165
Yip EC, Powers KS, Aviles L (2008) Cooperative capture of large prey solved scaling challenge faced by spider societies. Proc Natl Acad Sci USA 105:11818–11822
Zaera R, Solar A, Teus J (2014) Uncovering changes in spider orb-web topology owing to aerodynamic effects. J R Soc Interface 11:20140484
Zevenbergen JM, Schneider NK, Blackledge TA (2008) Fine dining or fortress? Functional shifts in spider web architecture by the western black widow Latrodectus hesperus. Anim Behav 76:823–829
Zhou H, Zhang Y (2005) Hierarchical chain model of spider capture silk elasticity. Phys Rev Lett 94:028104
Zschokke S, Vollrath F (1995) Web construction patterns in a range of orb-weaving spiders (Araneae). Eur J Entomol 92:523–541
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter

Cite this chapter
Blamires, S.J., Zhang, S., Tso, IM. (2017). Webs: Diversity, Structure and Function. In: Viera, C., Gonzaga, M. (eds) Behaviour and Ecology of Spiders. Springer, Cham. https://doi.org/10.1007/978-3-319-65717-2_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-65717-2_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-65716-5
Online ISBN: 978-3-319-65717-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)