
Abstract
The homologies between the malleus, incus, and ectotympanic bones of the mammalian middle ear and the articular, quadrate, and angular bones of the tetrapod palatoquadrate, respectively, are well-established by embryonic evidence. The evolutionary question of how the implied transition occurred is less clear and, historically, there have been two general views. The first view is that the ancestral state was a reptile-like tympanic membrane behind the quadrate that activated the stapes, and that the jaw hinge bones were subsequently incorporated between tympanic membrane and stapes. The second view is that in the ancestor, low-frequency ground-borne sound was received by the lower jaw and transmitted via the hinge bones and stapes to the fenestra ovalis. The anatomy of the middle ear region of the known sequence of fossil stem-group mammals—pelycosaurs, basal therapsids, and several cynodonts—is reviewed in this chapter. As with almost all recent authors, the interpretation offers support for the second view. Within the cynodont grades, decreasing mass of the postdentary bones relative to the dentary is part of a complex of changes in the feeding mechanism but also implies increasing sensitivity to airborne sound. As long as the jaw hinge continued to perform a mechanical role in mandibular function, the bones were too massive to be receptive to higher frequency sound, and therefore an air-filled tympanic cavity and tympanic membrane were unlikely to have evolved. This latter stage awaited the origin of the new mammalian jaw hinge between the dentary and squamosal bones in mammaliaforms.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Cynodonts
- Dicynodonts
- Evolution of hearing
- Origin of mammals
- Pelycosaurs
- Synapsids
- Synapsid ear
- Therapsids
- Therocephalians
5.1 Introduction
The presence of a chain of three middle ear ossicles, the malleus, incus, and stapes in the mammals compared to the single columella auris in living amphibians, reptiles, and birds, offers a classic example of the power of comparative embryology to reveal obscure homologies, as iconic in its context as, say, Archaeopteryx is in the context of the power of paleontology. The pre-Darwinian anatomist Karl Reichert (1837) is credited with the discovery that the malleus and incus of the mammals develop from the same parts of the first visceral arch that become the jaw hinge bones, the articular and quadrate respectively, in the reptiles (Fig. 5.1A). The mammalian stapes and the reptilian columella auris are formed within the second visceral arch, so they too were inferred to be homologous. As reviewed by a number of authors (e.g., Anthwal et al. 2013; Maier 1990; Takechi and Kuratani 2010), further comparative anatomical and embryological evidence subsequently accrued that continued to support this conclusion, such as the pattern of innervation and the musculature. Most notably, Gaupp (1913) demonstrated the additional homologies of the angular bone of the reptilian jaw with the mammalian ectotympanic bone, and the reptilian prearticular bone with the mammalian goniale that is attached to the malleus (Fig. 5.1B).

(A) Skull of the embryo of an armadillo (Tatusia hybrida) showing Meckel’s cartilage (mk) with manubrium (ml), incus (in), stapes (st), and ectotympanic (a.ty). For other abbreviations, see original publication. (B) Comparison of the posterior end of a cynodont jaw (left) with the posterior end of the jaw and ear ossicles of a late embryo of the marsupial Perameles (right). (A, B from Goodrich 1930)
The homologies of the parts of the middle ear originally were established at a time when the cause and meaning of homology was not clear. The evolutionary question only subsequently arose as to how the transformation from the basal amniote condition of jaw hinge bones to the mammalian accessory ear ossicles occurred. This is a functional as well as an anatomical question and embraces the extent to which, if at all, the tympanic ear of the mammal is homologous as a sound-receiving structure with those of other tetrapods. Two alternative broad scenarios emerged during the 20th century. One (e.g., Goodrich 1930; Parrington 1946; Hopson 1966) presumed that the mammalian tympanic membrane is a postquadrate tympanum homologous with that of modern reptiles and birds. In the common ancestor, the tympanic membrane would have been connected directly by the stapes to the fenestra ovalis (FO), and only later in the mammalian lineage did a reduced articular and quadrate became incorporated, which enhanced the impedance matching between the airborne sound waves received by the tympanic membrane and the waves transmitted into the fluid-filled cochlea.
The second evolutionary scenario (e.g., Gaupp 1913; Tumarkin 1955; Allin 1975) is that the amniote-grade common ancestor of reptiles and mammals lacked a tympanic membrane, and that in the mammalian lineage sound waves were originally received by the lower jaw and transmitted via the angular, articular, and quadrate bones to the stapes and FO. In this view, the evolution of the definitive mammalian ear had to await the origin of a new jaw articulation between the dentary and the squamosal, from which point there followed a reduction in the mass of the postdentary bones and quadrate and loss of their connection to the jaws, but no change in their essential topological relationships to one another. The mammalian tympanic membrane is seen, therefore, as a neomorph and not homologous with that of any other living tetrapod taxon.
More recent developmental and genetic evidence supports the second view—the mammalian tympanic membrane is not homologous with the tympanic membrane of reptiles and birds. Takechi and Kuratani (2010) showed that certain genes associated with development of the malleus and incus also affect the tympanic membrane, suggesting a developmental relationship between these elements that is presumably absent in non-mammalian amniotes. Going further, Kitazawa et al. (2015), by inactivating certain genes involved in jaw development, found that formation of the tympanic membrane of the mouse was associated with correct lower jaw development but was associated with upper jaw development in the chick. This is what would be expected if the mammalian tympanic membrane was historically attached to the angular bone, while that of the non-mammalian amniotes to the quadrate of the upper jaw.
Thompson and Tucker (2013) described the development of the middle ear cavity in mammals. It involves breakdown of part of the endodermal epithelial lining, invasion by a mass of neural crest mesenchyme cells, and subsequent cavitation of the latter. None of these processes occur in the non-mammalian amniotes, which implies that the cavity of the two respective taxa is not homologous and, therefore, the tympanic membrane bounding the cavity must have had a separate evolutionary origin.
Fossil members of the mammalian lineage, colloquially referred to as “mammal-like reptiles” and more formally as the paraphyletic taxon non-mammalian Synapsida, were first collected from Upper Permian rocks in South Africa in the middle of the 19th century and described by Richard Owen (1845), who initially regarded them only as a new kind of reptile. By the time of the publication of Owen’s catalogue of fossil reptiles in the British Museum (Natural History), a considerably greater diversity of what are now referred to as therapsids was known and, furthermore, their mammalian affinity had become recognized (Owen 1876). Meanwhile in North America, members of even more basal synapsids, the peleycosaurs, were being discovered in Upper Carboniferous and Lower Permian rocks and described by Cope (1877) and later by Case (1907). However, it was not until 1910, when the South African paleontologist Robert Broom visited the USA that the opportunity arose to compare the North American and South African fossils. He demonstrated beyond any subsequent serious doubt that the two are related and together illustrate stages in the evolution of the mammalian lineage: the stem mammals (Broom 1910).
Over the intervening century, the non-mammalian synapsid record has expanded into numerous taxa representing some dozen grades (Fig. 5.2) between the basalmost pelycosaurs, which possessed only two or three definitive mammalian characters, and the earliest mammaliaforms. This has had profound consequences for understanding how the structure of the middle ear evolved in the mammalian lineage, to culminate in the ancestral mammaliamorph grade represented by, for example, the Upper Triassic Morganucodon.

Phylogenetic relationships of the synapsids
However, anatomy aside, there have been very few serious attempts at functional analyses of hearing in the fossil synapsids. The majority of opinions expressed have been based on little more than subjective assumptions about whether there is evidence for the attachment of, or indeed enough space to accommodate, a tympanic membrane large enough to have activated the stapes in question. There have also been a number of vague assertions about the possible extent of non-tympanic and seismic sound reception. Exceptions include the studies by Hotton (1959), using a physical model of a pelycosaur middle ear; by Kemp (2007), who attempted to quantify some of the acoustic properties of a eucynodont; and by Laass (2014) who considered a dicynodont skull from the point of view of design for various possible modes of sound reception. The issue of non-mammalian synapsid hearing is colored, inevitably perhaps, by considerations of the transition to the mammalian system. It is rarely addressed in its own right.
5.2 The Pelycosaur Grade
The stapes of pelycosaurs (Fig. 5.3) is invariably described as massive in comparison with the relatively slender structure of those found in subsequent synapsids and is presumed to be the ancestral condition for the lineage. In the case of Dimetrodon (Romer and Price 1940), a roughly circular footplate fits over the FO from which the shaft extends ventrolaterally toward, but does not reach, the lower part of the quadrate (Fig. 5.3D). This end is tapered and consists of unfinished bone, suggesting continuation by cartilage in life. There is a stout dorsal process near to the footplate that articulates with the overlying paraoccipital process and a stapedial foramen pierces the stapes. In the relatively few cases known, other pelycosaurian stapes have a similar form, although in smaller species the stapes is more horizontally disposed (Fig. 5.3E) (Hotton 1959).

Pelycosaurian grade. Dimetrodon. (A) Reconstruction of the skeleton. (B) Skull in left-lateral view. (C) Lateral (above) and medial (below) views of the lower jaw. (D) Posterior view of the skull with Hotton’s interpretation of the position of a tympanic membrane added on the right. (E) Posterior view of the skull and jaws of Varanops. (A from Clack 2012, used with permission; B, C original from Kemp; D, E from Romer and Price 1940 and Hotton 1959)
As reviewed comprehensively by Hotton (1959), there has been much speculation in the past about the function of the stapes in pelycosaurs, mainly concerning whether a tympanic membrane existed at all and, if so, where it was located. In their definitive description, Romer and Price (1940) felt that the mass of the stapes excluded it from a sound transmission function altogether, although they are silent on what precisely the function of the bone might have been. Watson (1952) also believed that there was inadequate space for a sufficiently large tympanic membrane and that there is no positive osteological indication of where one might have been attached. On the other hand, Parrington (1955, 1979) proposed that the distal end of the stapes was continued as a cartilaginous extrastapes reaching to a tympanic membrane low down behind the quadrate, a view influenced by his belief that both the non-synapsid predecessors and the therapsid successors of pelycosaurs possessed a postquadrate tympanic membrane.
Hotton (1959) constructed a mechanical model to test whether a stapes of the mass found in pelycosaurs could be activated to a significant extent by a membrane of a size that could be accommodated medial or posterior to the quadrate. His results showed that the model membrane did in fact enhance sound conduction via the stapes, though not to any great degree. However, there are serious difficulties in interpreting the results, for example the extent to which the parts of the model realistically match the physical properties of the living system, and the inability to distinguish between sound transmission due to the vibration of the stapes as a whole and intramolecular sound transmission within it. Actually fitting a similarly sized tympanic membrane perpendicular to the stapes shaft in the actual skull is also difficult, and Hotton’s reconstruction has the membrane of Dimetrodon rather improbably in a more or less transverse plane, attached along the dorsolateral margin of the stapes shaft rather than terminally (Fig. 5.3D).
The reflected lamina of the angular of the lower jaw first appears in the sphenacodont pelycosaurs such as Dimetrodon, as a ventral keel that is set slightly laterally to the major plane of the jaw (Fig. 5.3B). Generally, its function is regarded as an area for the insertion of internal mandibular musculature, particularly the anterior pterygoideus muscle descending posteroventrally from the palate (Barghusen 1968) and, perhaps, intermandibular musculature. The external surface of the angular and its lamina, in common with the external surface of the whole of the postdentary region of the jaw, is finely rugose, indicative of tightly applied skin rather than either muscle or any form of tissue that could be regarded as a mandibular tympanic membrane. The quadrate is firmly sutured to the squamosal and, like the stapes, is of a size to render it impossible that it could have vibrated as a whole bone to transmit sound.
Further work on the possible functioning of the stapes and associated structures in sound conduction in pelycosaurs is long overdue. Currently the most reasonable view is that no tympanic membrane was present, and low-frequency, high-amplitude ground- and airborne sound were received and transmitted via intra-bone conduction, from mandible to quadrate and quadrate to stapes. However, certain anatomical features are not readily explained by this hypothesis, notably the indication of a cartilaginous rather than a bony connection between the quadrate and stapes.
5.3 Basal Therapsids
Therapsida is a more derived clade whose closest relatives are the sphenacodont pelycosaurs (Fig. 5.2) with which they share several synapomorphic characters. Most prominently, these include the reflected lamina of the angular, which had become greatly enlarged in therapsids, and the enlargement of teeth in the canine region of both upper and lower jaws. Indeed, therapsid ancestry probably lies within the Sphenacodontia, rendering the latter technically paraphyletic (Benson 2012; Huttenlocker and Rega 2012).
During the Middle Permian, therapsids radiated into several lineages more or less simultaneously. The interrelationships among these lineages are not completely resolved and the phylogenetic pattern approaches the condition of a soft polytomy (Fig. 5.2), the phenomenon in which there are inadequate shared characters to distinguish with confidence between a finely spaced sequence of dichotomous branchings and a truly simultaneous multiple split (Kemp 2009a, 2012). The implication of this pattern of therapsid radiation is that the taxon rapidly differentiated into a series of different specialist ecotypes, some carnivore and some herbivore. This is indicated by the considerable variation found in the anatomy of the dentition, jaws, reconstructed jaw musculature, and jaw articulation. The structure of these parts of the animal clearly reflects the function in their primary role as the feeding mechanism: collecting different kinds of food, generating bite and triturating forces, manipulating food in the oral cavity, etc.
Given this variation in the postdentary bones and quadrate, if these also had a role in sound reception, then it must have involved a mode of hearing in which the precise design was not particularly critical. Indeed, the stapes too varies to a surprising extent in size and structure both among and within different therapsid taxa. Detection of high-frequency airborne sound does require an adequately designed system, and so it follows that hearing in basal therapsids was probably limited to detection of low-frequency vibrations via intra-bone conduction. This requires a greatly less critical design than does the tympanic-activated, whole bone vibration type of middle ear of modern amniotes including mammals. The lack of a close correspondence between the form of an intra-bone low-frequency sound transmission route and its potential effectiveness, however, does make it difficult to assess the hearing performance of therapsids.
5.3.1 Biarmosuchia
Ignoring a few very poorly preserved specimens, the most basal therapsids are the biarmosuchians (Fig. 5.4). From them is inferred the ancestral therapsid structure of the jaw, jaw articulation, and middle ear (Ivakhnenko 1999; Sidor and Rubidge 2006). The stapes is very much reduced compared to the pelycosaur condition, and it extends laterally from the FO to contact the medial side of the quadrate as a fairly delicate rod. The presence of a stapedial foramen appears to be variable (Sidor and Rubidge 2006). The quadrate has not been fully described in any specimen, but appears to be relatively large and firmly fixed to the squamosal behind. The most prominent new character in this region of the skull is the huge expansion of the reflected lamina of the angular. It consists of a broad, but very thin, vertical sheet that extends below the level of the ventral margin of the body of the angular to which it is attached anteriorly. The posterodorsal and posterior margins are free edges. A very narrow cleft exists between the lamina externally and the body of the jaw internally, which is termed the angular recess. The lamina itself is marked by two corrugations radiating from around the center and presumably designed to increase the strength and/or stiffness of the structure. The function of the reflected lamina of therapsids, including its possible role in sound reception, remains a matter of unresolved debate. In part, this is because there is no comparable structure on the jaw of any living amniote and, in part, because of its status as the homologue of the otherwise entirely dissimilar, tiny ectotympanic bone of mammals that supports the tympanic membrane.

No functional consideration of hearing specifically in biarmosuchians has yet been presented, although since all the more derived therapsid taxa are assumed to have evolved from a generalized biarmosuchian morphotype, comments about function in the former may well apply also to the latter.
5.3.2 Gorgonopsia
The gorgonopsians (Fig. 5.5) were hypercarnivores of the basal therapsid radiation (Kemp 1969). The dentition was modified by a huge enlargement of the upper and lower canine teeth for prey disablement, along with facultative interdigitation between prominent lower and upper incisors for food processing. A highly specialized jaw articulation met the simultaneous requirements for a gape of at least 90°, which was necessary for the canines to clear one another during food capture and for propalinal (back and forth) movement of the lower jaw so the lower incisors could become engaged with the upper incisors. To achieve the wide gape there is a screw-shaped jaw articulation (Fig. 5.5B) between the quadrate and articular (Parrington 1955). The propaliny resulted from the quadrate being moveably attached to the squamosal (Fig. 5.5C) in a fashion that allowed the jaw hinge to rotate forward and backward (Kemp, 1969). The reflected lamina of the angular possessed a strong vertical ridge on its lateral face to which attached a large new part of the adductor musculature, originating from the squamosal anterolaterally to the jaw articulation. Although comparable in position and function to the masseter of the mammals, this muscle evolved convergently in gorgonopsids.

Gorgonopsian grade. (A) Reconstruction of the skull in left-lateral and dorsal views showing main jaw adductor musculature. (B) Anterior view of the left squamosal showing the recess in which the quadrate complex lay, with anterior (left) and posterior (right) views of the corresponding quadrate below. (C) Diagram of the lateral view of the quadrate rotation in the squamosal recess to achieve a forward shift of the quadrate condyles. (D) Medial and anterior views of the left stapes of a gorgonopsian. ART, articular; Q, quadrate; SQ, squamosal; m, muscle vectors causing the rotation. (A original drawing; B–D from Kemp 1969)
The stapes (Fig. 5.5D), in the few cases where it has been adequately described (Parrington 1955; Kemp, 1969), is a flat bone lying in a transverse plane and extending laterally to a recess on the medial side of the quadrate. There is a substantial stapedial foramen occupying the medial half of the bone and a prominent dorsal process toward the distal end of the shaft in contact with the overlying paroccipital process.
Parringon (1955) was convinced that there was a small, reptilian tympanic membrane in gorgonopsians located at the terminal end of the stapes, low down and immediately behind the quadrate. This position coincides with the inner, ventral termination of a broad, open sulcus, or channel, where the squamosal sutures with the lateral edge of the tabular bone, which he interpreted as the route of a tubular external auditory meatus. The arguments against the existence of the postquadrate tympanic membrane are, as in other basal synapsids, that the tympanic membrane would have been too small in area to have been functional and that there is no osteological indication of its support. The moveable nature of the quadrate in its socket in the squamosal could perhaps be regarded as a precondition for eventually evolving a whole-bone vibrating element for sound transmission, but at this stage the quadrate was far too massive to have functioned in this manner. On the other hand, the form of the squamosal sulcus remains unexplained if it did not house an external auditory meatus: possibly it was related to the architecture of the occipital and neck musculature.
5.3.3 Dicynodontia
The specialized herbivorous taxon Dicynodontia (Fig. 5.6) was by far the most abundant and diverse lineage of therapsids of the Upper Permian. The stapes lacks a stapedial foramen in almost all cases except for the basal genus Eodicynodon (Rubidge 1990) and the specialist digging genera Cistecephalus and Kawingosaurus (Cox 1972; Laass 2014), in which a presumably secondarily derived perforate condition may be associated with subterranean sound detection. The orientation of the stapes is approximately perpendicular to the sagittal plane, and both the footplate and distal end are expanded. The latter abuts firmly up against the medial side of the quadrate. The quadrate, along with the intimately attached quadratojugal on its lateral side, is large, flattish, and set in a shallow fossa on the more or less transverse anterior face of the squamosal. There is no firm suture between the quadrate complex and squamosal, suggesting that a small degree of movement between the two was possible. The articulation between the quadrate and the articular is relatively elongated and open, allowing some anteroposterior motion of the lower jaw (Crompton and Hotton 1967) in association with the action of the horny tooth-plates that are assumed to have occupied the jaws in lieu of most or all of the dentition. The dicynodont reflected lamina of the angular is large and smooth surfaced, enclosing a substantial angular recess. The lower part tends to curve medially below the ventral margin of the jaw.

Dicynodont grade. (A) Ventral view of the posterior half of the skull of Lystrosaurus, showing the preserved ceratohyal. (B) left-lateral and ventral views of Dicynodontoides (Kingoria). (C) Possible routes of seismic sound reception by bone conduction to the ear (shaded) according to Laass (2014). Top: arrows indicate compressional bone conduction from the burrow wall to the skull roof detected due to the inertia of the mandible causing relative movement between the articular and the quadrate (double arrows). Center: arrows indicate conduction from the ground via the lower jaw, articular, quadrate, and stapes, relying on inertia of the rest of the skull. Bottom: arrows indicate sound conduction from the ground or the air via the mandible, jaw articulation, and stapes. F a , direction of air borne sound pressure; F g , direction of ground-borne sound pressure; q, quadrate; qj, quadratojugal; sta, stapes. (A from Barry 1968; B from Cluver and King 1983; C from Laass 2014; all used with permission)
In an early contribution to the functioning of the middle ear in dicynodonts, Barry (1968) described what he took to be an ossified ceratohyal attached toward the inner end of the stapes and extending forward into the throat region in the Lower Triassic Lystrosaurus (Fig. 5.6A). In comparison with the condition in the modern tuatara Sphenodon, Barry (1968) concluded that it was involved in seismic sound reception and, furthermore, that no tympanic membrane was present. However, as no such ossification has been reported in other dicynodonts, and as Lystrosurus is a specialized, burrowing form, its middle ear must be assumed to be specialized for that particular mode of life. In contrast, Cox (1959) noted a small, posteriorly projecting process on the occiput of Dicynodontoides (Kingoria). It lies close to the lateral end of the paroccipital process (Fig. 5.6B), and he interpreted it as partial support for a tympanic membrane. However, in the light of its absence from other dicynodont taxa, and the evidence otherwise against the presence of a postquadrate tympanic membrane in basal therapsids generally, it is more probable that the process represents the point of attachment of a ligament associated with head support.
The most recent and the most detailed functional study of a dicynodont middle ear is that of the digging dicynodont Kawingasaurus by Laass (2014), based on his neutron tomography investigation of the structure. He concluded that there were at least two routes by which seismic sound could be detected (Fig. 5.6C). One was by inertial bone conduction from the flat, massive skull roof in contact with the wall of a burrow. In this case, the non-vibrating, fixed reference structure was the mandible, which was acoustically isolated from the cranium by the relatively highly compliant articular-quadrate contact. The second route was a direct one, and it consisted of seismic sound waves being received by the ventral margin of the mandible and transmitted via the quadrate and stapes to the FO. Support for the existence of this route is the ventrolateral orientation of the stapes toward the quadrate attachment, the relatively large size of the stapedial footplate, and the massiveness of the quadrate and stapes.
For these latter reasons, Laass (2014) also concluded that detection of airborne sound was poor and limited at best to low frequencies. As well as this high inertia of the bones, he calculated that there could have been a pressure transformer ratio of no more than 2–3, based on the estimated area of the angular and its reflected lamina as the presumed site of airborne sound reception (25–47 mm2) and the area of the stapes footplate (13.2 mm2). This figure is well below that of any mammal and also of eucynodonts, in which airborne sound probably was received and transmitted by the postdentary bones, as discussed in Sect. 5.4.3.
Although no similar study has been made for non-burrowing dicynodonts, their anatomy is sufficiently comparable to Kawingasaurus to suggest that detection of seismic sound and only low-frequency airborne sound was the norm. The difference is that in non-burrowing forms the stapes is more or less horizontal and so perpendicular to the plane of the postdentary bones, which might imply that airborne sound was of somewhat greater significance.
5.3.4 Therocephalia
The therocephalians (Fig. 5.7) constitute the next grade in the mammaliaform lineage known from the fossil record and represent the presumed condition of the middle ear region ancestral to that of the cynodonts (Kemp 1972b; Maier and van den Heever 2002). Notably the reflected lamina is particularly large and thin compared to other therapsids, and differs from those of gorgonopsians and dicynodonts in being free of the body of the angular bone along all of its upper margin. It is strengthened by a substantial pattern of corrugations, which led Kemp (1972a) to propose that its function was primarily to provide insertion for part of the mandibular muscle complex. The glenoid fossa of the articular bone is broad, widely open, and faces posteriorly and medially. The quadrate–quadratojugal complex (Fig. 5.7B) appears to have been moveably attached, but in a quite different manner from that of gorgonospsids. The quadratojugal component was held in a slit in the squamosal in such a way that rotation of the complex about a longitudinal axis was possible (Fig. 5.7C). Kemp (1972a) proposed that both the orientation of the jaw articulation surfaces (Fig. 5.7D) and the form of the mobility of the quadrate complex were designed to resist a large bite force that was generated by adductor musculature running dorsomedially from its insertion on the mandible. As it happens, the ability of the quadrate to rotate about a longitudinal axis, whatever its initial mechanical function may have been, is a necessary precursor to later stages in the evolution of cynodont and eventually mammalian hearing as described in Sect. 5.4 (Luo and Crompton 1994). The therocephalian stapes (Fig. 5.7E) is an imperforate, dumbbell-shaped bone that has a broad contact with the medial side of the quadrate (Kemp, 1972a; van den Heever 1994; Maier and van den Heever 2002). There appears to have been no possible room for a postquadrate tympanic membrane near the distal end of the stapes.

Therocephalian grade: (A) Left-lateral and dorsal views of the skull with principal jaw adductor musculature shown. (B) Posterior view of the right quadrate complex and its contacts. (C) Diagram of the mobility of the quadrate complex about a longitudinal axis at x, as viewed from behind of Theriognathus (Whaitsia). (D) Diagram of the dorsal (above) and posterior (below) views of the skull and jaws to show the orientation of the axis of rotation of the jaw articulation. Because of the oblique nature of the axis, mobility of the quadrate on the squamosal is necessary to permit opening and closing of the jaws. (E) Ventral view of the posterior part of the skull of Tetracynodon. m, muscle force; r, hinge reaction force. (A original drawing; B, C and D from Kemp 1972a; E from Sigurdsen et al. 2012)
Whatever role the angular and its reflected lamina may have had in jaw mechanics does not exclude a secondary role in sound reception (Allin 1975; Maier and van den Heever 2002). The open form of the jaw articulation and the considerable mass of the hinge bones militate against any role in sound transmission by whole bone vibrations from the angular via the articular, quadrate, and stapes to the FO, but intra-bone conduction of low-frequency sound collected by the reflected lamina would have been possible. Maier and van den Heever (2002) calculated the ratio of the area of the reflected lamina to that of the stapes footplate as about 4.0, giving a very poor potential sound pressure level ratio, one that could only have been associated with sensitivity to low-frequency, high amplitude sound transmission from postdentary bones to stapes.
5.4 Non-Mammalian Cynodonts
Much attention has been paid to the middle ear of the non-mammalian cynodont therapsids because it is within them that the transition from the basal therapsid anatomy to one recognizably approaching mammalian anatomy can be followed. In the earliest members of the Cynodontia, the quadrate and postdentary bones are not greatly reduced compared to therocephalians, although the reflected lamina of the angular is much smaller and the stapes has a large stapedial foramen. However, within the taxon there was a progressive reduction in these elements as illustrated by several grades of fossils up to the definitive mammaliamorph stage. By this time, the new, mammalian jaw hinge between dentary and squamosal had evolved and the function of the articular and quadrate, now referred to as the malleus and incus, was exclusively in sound reception.
5.4.1 Procynosuchus
Procynosuchus (Figs. 5.8, 5.9) is by far the best known of the basalmost cynodonts (Kemp 1979). The stapes possesses a very large stapedial foramen between quite delicate anterior and posterior arms (Fig. 5.9C). The footplate is a little larger than the FO (Fig. 5.9B) and so made contact with the bony margin of the latter, no doubt by means of a cartilaginous cap. The distal end also lacks a periosteal layer and partially fits into a shallow recess in the lower medial part of the quadrate but with about half of its face extending behind the quadrate. A ridge on the dorsal surface of the stapes lying distal to the foramen represents a dorsal process contacting the overlying paroccipital process. The quadrate-quadratojugal complex lies against the anterior face of a shallow squamosal recess (Fig. 5.9B) without forming a suture and is similar to, but relatively larger than, that of the better known Thrinaxodon, described in Sect. 5.4.2. The articulation of the quadrate with the articular bone of the lower jaw is somewhat similar to that of therocephalians. There are two quadrate condyles that face anteroventrally rather than ventrally, and the corresponding posterodorsally facing articular condyle lacks a hind rim. The angular bone is dominated by a broad shallow recess over its lateral face, indicating that adductor musculature had expanded its insertion over the external surface of the jaw. There is a very reduced and simplified reflected lamina of the angular restricted to the anteroventral region of the main angular bone and, therefore, leaving freely exposed most of what had been the angular recess.

(A) Preserved skeleton of Procynosuchus delaharpae from the Upper Permian of Zambia (TSK collection field number TSK34, Natural History Museum, London). (B) Model of Procynosuchus in life based on the skeleton, by Richard Hammond

Procynosuchus. (A) Reconstruction of the lateral and dorsal views of the skull with the principal jaw adductor muscles. (B) Ventro-lateral view of the skull to show fenestra ovalis. (C) Stapes in dorsal (d), ventral (v), posterior (p), anterior (a), proximal (prox), and distal views. (B, C from Kemp 1979)
Kemp (1979) argued that the stapes had a dual role. One concerned the mechanics of the jaws and was to give some support to the mobile quadrate. The second, as inferred from its reduced mass and direct contact with the quadrate, was the conduction of sound waves to the FO. Following the theory of Allin (1975), Kemp proposed that this included ground-borne and low-frequency airborne sound, detected by the postdentary bones and transmitted by the small, loosely attached quadrate. More improbably, Kemp also thought it possible that a tympanic membrane existed behind the lower end of the quadrate, even though there is no indication of where exactly it might have attached, on the grounds that this would explain why half the distal end of the stapes lay behind the quadrate and shows signs of completion in cartilage, presumably as a cartilaginous extrastapes.
5.4.2 Thrinaxodon
The posterior jaw and middle ear region of the epicynodont Thrinaxodon (Fig. 5.10) (Parrington 1946; Luo and Crompton 1994) are similar to those of Procynosuchus except that the postdentary bones and jaw hinge are relatively smaller compared to the dentary (Fig. 5.11A). The dorsoventrally flattened stapes (Fig. 5.10B), with a large stapedial foramen, abuts against the medial face of the quadrate in the same way. The quadrate lies against the anterior face of the squamosal, but there is a significant notch in the ventral border of the latter bone exposing part of the back of the quadrate (Fig. 5.10B). The manner in which the quadratojugal attached is also further modified. It is a tall, narrow blade wrapped around the lateral edge of the quadrate, and it lies in a near vertical extension of the quadrate notch. As in Procynosuchus, the articulating region of the quadrate consists of lateral and medial condyles aligned to face anteroventrally and corresponding to the posterodorsally facing articular condyle. The reflected lamina of the angular is small, thin, and ventrally placed, exposing almost all of the lateral face of the main body of the angular. The postdentary complex is not strongly sutured to the dentary but must have been attached only by soft, relatively compliant tissue, giving it the potential for vibrational movements independently of the rest of the jaw.

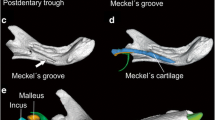
Thrinaxodon. (A) Lateral view of the skull with Hopson’s reconstruction of an external auditory meatus leading to a post-quadrate tympanic membrane. (B) Ventral view of the skull with an enlargement of the postero-lateral corner. (C) Luo and Crompton’s interpretation of the ear region as a model showing the vibrational motion of the quadrate as activated by a tympanic membrane supported by the angular and its reflected lamina; see source for abbreviations. (D) Photograph of a skull in left-lateral (top), right-lateral (middle), and ventral views (bottom). (C from Luo and Crompton 1994, used with permission; D specimen T815, Cambridge University Museum of Zoology, photographed by J.A. Clack)

(A) Parrington’s interpretation of the ear region of a cynodont. (B–E) Allin’s interpretation of the ear region of cynodonts: (B) Thrinaxodon grade, (C) eucynodont grade, (D) basal mammaliamorph grade, and (E) modern mammal grade. ang.pr, angular process of the dentary; ART, articular; art.pr, articular process of the dentary; D, dentary; dep.mand, depressor mandibuli muscle; ECTO, ectotypmpanic; I, incus; M, malleus; Q, quadrate; rart.pr, retroarticular process; refl.lam, reflected lamina of the angular; SA, surangular; SQ, squamosal; STA, stapes; tym, tympanic membrane. (A from Parrington 1979, used with permission; B–D from Kemp 1982, modified from Allin 1975)
Parrington (1946), in his classic description of the skull of Thrinaxodon, was convinced that, as in all cynodonts, the stapes possessed a cartilaginous extrastapes attached to a tympanic membrane positioned low down behind the quadrate (Fig. 5.11A). Furthermore, he accepted the interpretation of the sulcus on the back of the squamosal as the route of an external auditory meatus leading to the supposed tympanic membrane. In his view, the angular and its reflected lamina were not involved in sound reception. Hopson’s (1966) reconstruction of the middle ear of Thrinaxodon was essentially the same with a tympanic membrane attached to the squamosal, completely behind the quadrate (Fig. 5.10A).
In contrast to this view, Allin (1975) developed the idea of a postdentary tympanic membrane attached to the upper edge of the angular bone, further supported by the reflected lamina below and the retroarticular process of the articular behind (Fig. 5.11B). The squamosal sulcus was interpreted as the site of origin of a depressor mandibuli muscle, admittedly surprisingly large, from the occiput to the retroarticular process of the articular. He assumed there was an air-filled recessus mandibularis, equivalent to a tympanic cavity, bounded medially by the lateral face of the articular. Allin’s interpretation, like the various forerunners of the general idea, is predicated upon the particular homologies of the individual bones with those of mammals, but his version also includes an explicit claim of functional continuity. The angular and its reflected lamina had the same function of supporting the tympanic membrane as the incompletely circular ectotympanic bone of the mammal, while the retroarticular process of the articular had the function of the mammalian manubrium of attaching to the center of the tympanic membrane as a lever arm of the incus. The nature of the quadrate-stapes contact is inferred to have been functionally the same as in mammals. Allin never completely abandoned the possibility of some sort of postquadrate tympanic membrane also being present, a view earlier proposed by Watson (1952). Indeed, in later publications (Allin 1986; Allin and Hopson 1992), he reconstructed a rather awkward-looking arrangement in which both kinds of tympanic membrane were present and suggested, with little evidence, that they may have been sensitive to different respective frequency ranges.
The most recent contribution to the function of the quadrate of Thrinaxodon is that of Luo and Crompton (1994). They demonstrated that the quadrate would have been capable of rotating about a longitudinal axis whose fulcrum was the contact with the posteriorly extending parts of the prootic and pterygoid bones (Fig. 5.10C). Rotational vibration of the bone about this axis would be caused by sound waves vibrating the postdentary bones and the tympanic membrane it supported. The quadrate vibrations would then be transmitted directly to the stapes where it abutted onto the quadrate’s medial side and thence via the footplate to the FO. As so interpreted, the functional relations of postdentary bones, quadrate, and stapes were fundamentally the same as the ectotympanic, malleus, incus, and stapes of mammals.
5.4.3 Eucynodontia
The clade Eucynodontia (Figs. 5.11C, 5.12) is the next grade in the trend in mandibular and jaw articulation structure seen through therocephalians, Procynosuchus and Thrinaxodon. The postdentary bones are reduced to a small, narrow, transversely flattened rod preserved loosely in a horizontal trough along the lower medial side of the dentary (Figs. 5.11C, 5.12B). It was held in place in life by soft tissue rather than firm bone-to-bone suturing. The reflected lamina of the angular, on the rare occasions when it is completely preserved, appears as a thin, delicate process extending backwards from the anteroventral corner of the body of the angular adjacent to the dentary. What had once been the angular recess between the main body and the sheet-like reflected lamina of the angular is by this stage completely open laterally. The quadrate bone (Fig. 5.12D) is similarly reduced in relative size, although attached in essentially the same way as in Thrinaxodon (Luo and Crompton 1994). The principal qualitative development in this region of the skull (Figs. 5.12C, D) is a new ligamentous contact between a small boss on the surangular of the postdentary rod and a facet on a flange of the squamosal immediately lateral to the quadrate-articular hinge (Crompton 1972; Kemp 2007).

Eucynodont grade. (A) Lateral view of a cast of the skull of the Middle Triassic South African Trirachodon (original specimen South African Museum No. K12168). (B) Kemp’s reconstruction of the postdentary bones and quadrate of Chiniquodon as a unit vibrating about an axis. (C) Ventral view of the posterolateral part of the skull of Chiniquodon showing lower jaw in situ. (D) Ventral view of the skull with enlargement of the posterolateral region of Probainognathus. ang.pr.d., angular process of the dentary; bear, bearing point of postdentary complex on dentary; BSP, basisphenoid; COR, coronoid; D, dentary; ext.au.me, possible site of an external auditory meatus; fen.ov., fenestra ovalis; jug.f., jugular foramen; p.pr., paroccipital process; Q, quadrate; QJ, quadratojugal; q.ra.pt., quadrate ramus of the pterygoid; rart.pr., retroarticular process; refl.lam., reflected lamina of the angular; sa.boss, boss on surangular for ligament attachment; sa.lig., surangular ligament; SPL, splenial; SQ, squamosal; STA, stapes; ‘tymp.m'.’ functional tympanic membrane. (B, C from Kemp 2007; D from Luo and Crompton 1994, used with permission)
Historically, the range of functional interpretations of the middle ear of eucynodonts is much the same and associated with the same authors as for Thrinxodon. One of the contentious anatomical points is whether the sulcus at the back of the squamosal housed an internal auditory meatus leading to a tympanic membrane (Watson 1952; Hopson 1966; Parrington 1979) or, alternatively, was the site of origin of a depressor mandibuli muscle that inserted onto the back of the articular bone (Allin 1975). Neither of these explanations is entirely satisfactory. Within the Eucynodontia, the sulcus in some taxa, such as Diademodon, was extraordinarily large, which provokes the argument that an associated depressor mandibuli muscle occupying the sulcus would have been far too large, either for an insertion on the very small retroarticular process of the angular or, indeed, for any feasible functional requirements of the lower jaw. However, the size and the curiously twisted course of the sulcus appears equally inappropriate for an air-filled external auditory meatus. It may be the case, therefore, that the sulcus had a quite different explanation. Its anatomy is highly variable in different eucynodonts, from the extreme prominence in diademodontids that have grinding herbivorous postcanine teeth, to a more moderate form in the carnivorous chiniquodontids, and the virtual absence from the high occiput of the apparently hypercarnivorous cynognathids. The variation in its architecture suggests that the form of this region of the skull reflects differing geometry of the jaw musculature and force patterns within the temporal fenestra and occiput in different kinds of feeding design rather than having anything to do with hearing.
For Allin (1975), an important key to understanding the middle ear of eucynodonts (Fig. 5.11C) is his argument that the reduction of the relative size of what were to become the mammalian middle ear ossicles—ectotympanic, malleus, and incus (Fig. 5.11B–E)—was the result of positive selection for sensitivity to higher frequency airborne sound, a shift permitted by the reduced mechanical requirements of the jaw hinge consequent upon the evolution of greater differentiation of the adductor mandibuli musculature (Crompton 1963; Kemp 1980). The temporalis muscle attached high up on the very prominent coronoid process, and had a posteriorly directed force vector, while the masseter muscle attached to the lateral face of the angular region of the dentary, giving it a dorsally directed force vector. The effect of this reorientation of the adductor musculature was to concentrate all the generated muscle force between occluding upper and lower postcanine teeth, with no reaction at all at the jaw hinge. With little reaction stress to be resisted, the articular and quadrate were free to reduce in size. Luo and Crompton (1994) demonstrated in more detail that the quadrate of a eucynodont such as Probainognathus (Fig. 5.12D) was not only reduced in size, but also its contact with the squamosal was modified to make it more mobile and, therefore, more able to be set into vibration about the longitudinal axis.
In a detailed consideration of the possible auditory function of the postdentary bones in the eucynodont Chiniquodon, Kemp (2007) showed that the postdentary rod was only loosely attached anteriorly by connective tissue within the dentary trough, while its posterior end was supported by the newly evolved ligament connecting the surangular with the squamosal, adjacent to the articular-quadrate articulation. Therefore, the rod was largely acoustically isolated from the dentary by the soft tissue connection and could have undergone rotational vibration about a longitudinal axis along its dorsal margin (Fig. 5.12B), just as the malleus essentially does in mammals. The quadrate, as noted, is also moveably attached by soft tissue to the squamosal. In principle, therefore, the postdentary rod and quadrate together formed a compliant malleus-incus unit as in mammals. Airborne sound falling on the angular part acting like a tympanic membrane would set the unit as a whole vibrating about its long axis, and this movement would be passed via the quadrate part to the head of the stapes. The stapes in turn would vibrate like a piston in the FO.
Based on this proposed mechanism, the impedance matching function of the middle ear of the specimen can be calculated as a pressure transformer ratio. The ratio between the area of the lateral surface of the angular bone acting as a tympanic membrane and the area of the FO is about 20. There is also a lever effect arising from the force arm from the estimated position of the axis of rotation of the postdentary bones to the center of the tympanic membrane being 1.5 times as long as the lever arm from the axis to the point at which the quadrate contacts the end of the stapes. The effective transformer ratio, as the product of these two values, is therefore around 1:30, which is within the range of modern mammals. On the basis of this calculation, the postdentary rod, quadrate, and stapes are well designed to transform airborne sound waves into fluid waves in the vestibule.
However, the postdentary rod and quadrate still had some residual mechanical functions associated with the jaw mechanics. Actively opening the jaw necessarily generates stress at the hinge, as does biting using the more anterior teeth and, in the case of carnivores, grasping struggling live prey between the jaws. For this reason, the quadrate and articular had to retain far greater mass than would be optimal in their use for sound reception alone. The actual mass of the postdentary rod plus the quadrate, as estimated from the volume of the elements detached from the specimen, is about 0.5 gm, which is around ten times the value found in typical mammals. The implication is that the inertia of the system would have been too high for effective transmission of high-frequency vibrations, although it would have functioned adequately at lower frequencies, perhaps up to about 2 kHz.
A further conclusion from Kemp’s (2007) study is that no air-filled tympanic cavity would have been present at this stage. Such a cavity lateral to the angular, as envisaged in Allin’s (1975) reconstruction, is problematic because the role of a tympanic cavity is to increase the compliance of the system for increased sensitivity to higher frequency sound. If the high moment of inertia of the rest of the system forbade higher frequency sound reception anyway, then a tympanic cavity would have been functionally redundant. Low-frequency sound waves acting on the surface of the tissue covering the angular bone and the small ventral continuation supported by the reflected lamina would have caused enough vibration of the bones to be detectable.
Tritheledontids (Fig. 5.13) are small, highly derived eucynodonts with a number of new mammalian characters, such as loss of the postorbital bar and the presence of prismatic enamel on the teeth. They are generally regarded as the sister group of mammaliaforms (Bonaparte et al. 2003). Most significantly, the dentary extended a little further posteriorly and made a simple contact with the squamosal, which is an incipient version of the new mammalian jaw hinge. The quadrate is reduced further in size compared to earlier eucynodonts, and it was attached even more loosely to the squamosal (Luo and Crompton 1994). Presumably, the effect was to increase its compliance, which, along with its reduced moment of inertia, would have enhanced sensitivity to high-frequency sound waves. From this point, the new squamosal-dentary contact was free to evolve into a fully mammalian glenoid articulation and so relieve the quadrate and articular from any further masticatory function. It may be speculated that acquisition of this newly acquired sensitivity to higher frequencies was the point at which an air-filled tympanic cavity would be beneficial, increasing the compliance, and therefore sensitivity, of what had become a dedicated chain of auditory ossicles.

Tritheledontid grade. Ventral and left-lateral views of the skull of Pachygenelus. (From Bonaparte et al. 2003, used with permission)
5.5 Summary
There are formidable difficulties in the way of a detailed understanding of hearing in non-mammalian synapsids. First, although the formal homologies of the relevant entities are well established, their actual anatomy is profoundly different from any living tetrapod, amphibian or amniote. For example, no living form has anything resembling the reflected lamina of the angular bone or a stapes that firmly abuts against the quadrate. There are simply no modern functional analogues with which to compare the mammal-like reptiles.
Second, the functional design for intra-bone sound conduction of low-frequency sound waves within skulls is not very critical: to some degree virtually any bony structure can transmit vibrations to anywhere with which it is sufficiently stiffly connected. Therefore, it is difficult to infer transmission routes of such sound from the anatomy. Yet most probably the ancestral synapsid mode of auditory function largely, if not entirely, consisted of just such intra-bone transmission of low-frequency seismic or airborne sound waves.
Third, the structures in question, stapes, articular, and quadrate hinge bones, and postdentary bones, also had mechanical functions in the context of mandibular action in feeding, so any design for hearing must have been extensively compromised by the more stringent optimal requirements for this second biological role. Finally, interpretations have been influenced by prejudgments concerning the transition from the basal tetrapod system to the mammalian condition—different assumptions point to different conclusions.
Most of the earlier hypotheses about how basal synapsids heard were based on little more than speculation about the position and the size of a possible tympanic membrane within the skull and how massive a stapes it would have been capable of actuating. This follows from an early presumption that a tympanic membrane and terrestrial hearing were primitive for tetrapods, dating from the origin of the group in the mid-Paleozoic (see Schoch and Anderson, Chap. 11; Sect. 5.2). In fact, about the only point of universal agreement is that a stapes whose footplate is applied to an open FO leading into the vestibular cavity must have been involved in some way in the mechanoreception of sound waves. Yet whether the stapes acted as the vibrating element relative to the braincase as the fixed element, or acted as the fixed element relative to vibrating cranial elements, may be debated.
Notwithstanding earlier beliefs of several authors, there seems little doubt now that the stapes of pelycosaurs, representing the ancestral synapsid condition, primarily served a mechanical role in the structure of the skull, although exactly what role remains unclear. The high mass of the stapes and consequent high moment of inertia, plus its firm attachment to the surrounding cranial bones and consequent low compliance, indicate that it would take enormous energy to set it into whole-bone vibration. Furthermore, there is no indication of either room for, or bony support of, a tympanic membrane likely to have been large enough to be capable of activating it. All of this points to hearing that was limited to the detection of low-frequency seismic and, perhaps, loud airborne sound waves. Although it is difficult to identify any specific route, the stapes presumably acted as the final element in transmitting such sound waves by intra-bone conduction from the lower jaw and other parts of the skull to the inner ear.
Evolution of the therapsid grade saw the start of major modifications to the jaw mechanics with larger and more complex adductor musculature generating increased and more controlled bite forces. The rapid differentiation of the therapsids into several specialized lineages is manifested by the variation amongst them of the anatomy and inferred functioning of the jaw musculature. The differences in the enlarged reflected lamina probably related to different patterns of intermandibular and internal adductor muscles, and the form of the jaw hinge reflected such aspects of feeding as the directions of the forces of the major adductor muscles and whether there was a requirement for horizontal as well as adductor movement of the mandible. Although the changes in these structures were primarily adaptations for feeding strategies, in the light of their subsequent evolution, it appears there was also a serendipitous effect on their role in sound detection. Indeed, the subsequent evolution of the middle ear and sound detection ability can only be fully appreciated in the context of correlated evolution of dentition, jaw musculature, jaw hinge, and stapes. Two new developments were of particular significance. The enlargement of the reflected lamina of the angular, for whatever primary reason, generated a greater area for receiving airborne sound waves able to set in motion the part of the mandible behind the dentary that could later become partially acoustically isolated from the dentary. At the same time, the tendency to evolve a moveable quadrate as a means of elaborating biting and food manipulation created the initial conditions for eventual acoustic isolation of this bone, along with the attached postdentary bones and stapes, from the rest of the skull. Thanks to these various modifications, significant detection of low-frequency airborne and seismic sound had probably become a significant part of the sensory repertoire of the more basal therapsids, such as gorgonopsians, dicynodonts, and therocephalians.
The trend initiated within therapsids continued within the cynodonts. From the relatively therocephalian-like procynosuchian grade there was continued enlargement of the adductor muscles and their geometrical reorganization into a posteriorly oriented temporalis muscle on the coronoid process and dorsally oriented masseter muscle on the angular region of the dentary. The effect of these changes was to allow a stronger but also more precisely applied bite action in association with larger, more complex occluding teeth, enhancing the function of food assimilation. It also reduced the stresses acting at the jaw articulation during biting and mastication.
Another trend in the cynodonts was increasingly to shift the insertion of the remaining parts of the adductor musculature that inserted on the postdentary bones onto the dentary. This relative enlargement of the dentary and reduction of the postdentary complex is hard to account for solely in terms of jaw mechanics because, as far as the size and geometry of the muscles is concerned, it is their attachment to appropriate mandibular lever arms that matters. It should be irrelevant whether those lever arms are part of the same bone or parts of separate but strongly sutured bones. However, the trend can be explained by the potential of these changes to enhance the sensitivity of airborne sound reception. By reducing the feeding stresses at the jaw hinge, the postdentary bones and the quadrate could reduce in mass and become loosely attached as a unit to the dentary, increasing their compliance relative to the latter. To this end, a secondary articulation between the surangular and the squamosal was first established as a means of further isolating the postdentary rod from the dentary. At the same time, the quadrate reduced in mass and increased in compliance relative to the squamosal and other cranial contacts by loosening its attachment. By the full expression of the eucynodont, the system was able to detect and transmit somewhat higher frequency, lower amplitude sound waves to the FO by means of whole bone vibration of these elements and the stapes.
The next stage in the transition to the mammalian middle ear is represented by tritheledontids. The dentary expanded posteriorly until it came to span what had already become a relatively very small gap to the squamosal. A simple contact between the two bones formed, from which moment this new squamosal-dentary contact was free to evolve into a fully mammalian glenoid articulation and so relieve the quadrate and articular from any remaining masticatory function. Rapid reduction in both the relative and the absolute mass of the hinge bones was now possible, along with the appearance of the definitive mammalian air-filled tympanic cavity. Still higher frequency sound could henceforth be detected, a conclusion supported by the beginning of the enlargement of the cochlear recess in mammaliaforms, and possibly associated, along with the enlarged cerebral hemispheres, with more complex activities such as nocturnal foraging and social behavior (Kemp 2009b).
References
Allin, E. F. (1975). Evolution of the mammalian ear. Journal of Morphology, 147, 403–438.
Allin, E. F. (1986). The auditory apparatus of advanced mammal-like reptiles and early mammals. In N. I. Hotton, P. D. MacLean, J. J. Roth, & E. C. Roth (Eds.), The ecology and biology of mammal-like reptiles (pp. 283–294). Washington: Smithsonian Institution Press.
Allin, E. F., & Hopson, J. A. (1992). Evolution of the auditory system in Synapsida (“mammal-like reptiles” and primitive mammals) as seen in the fossil record. In D. B. Webster, R. R. Fay, & A. N. Popper (Eds.), The evolutionary biology of hearing (pp. 587–614). New York: Springer-Verlag.
Anthwal, N., Joshi, L., & Tucker, A. S. (2013). Evolution of the mammalian middle ear and jaw: Adaptations and novel structures. Journal of Anatomy, 222, 147–160.
Barghusen, H. R. (1968). The lower jaw of cynodonts (Reptilia, Therapsida) and the evolutionary origin of mammal-like adductor jaw musculature. Postilla, 116, 1–49.
Barry, T. H. (1968). Sound conduction in the fossil anomodont Lystrosaurus. Annals of the South African Museum, 50, 275–281.
Benson, R. B. J. (2012). Interrelationships of basal synapsids: Cranial and postcranial partitions suggest different toplogies. Journal of Systematic Palaeontology, 10(4), 601–624.
Bonaparte, J. F., Martinelli, A. G., Schultz, C. L., & Rubert, R. (2003). The sister group of mammals: Small cynodonts from the Late Triassic of southern Brazil. Revista Brasiliera de Paleontologia, 5, 5–27.
Broom, R. (1910). A comparison of the Permian reptiles of North America with those of South Africa. Bulletin of the American Museum of Natural History, 28, 197–234.
Case, E. C. (1907). Revision of the Pelycosauria of North America. Washington, D.C.: Carnegie Institute.
Cluver, M. A., & King, G. M. (1983). A reassessment of the relationships of Permian Dicynodontia (Reptilia: Theraapsida) and a new classification of dicynodonts. Annals of the South African Museum, 91, 195–272.
Cope, E. D. (1877). Descriptions of extinct Vertebrata from the Permian and Triassic formations of the United States. Proceedings of the American Philosophical Society, 17, 182–193.
Cox, C. B. (1959). On the anatomy of a new dicynodont genus with evidence of the position of the tympanic membrane. Proceedings of the Zoological Society of London, 132, 321–367.
Cox, C. B. (1972). The pectoral region and habits of a new digging dicynodont. In K. A. Joysey & T. S. Kemp (Eds.), Studies in vertebrate evolution (pp. 173–189). Edinburgh: Oliver and Boyd.
Crompton, A. W. (1963). On the lower jaw of Diarthrognathus and the origin of the mammalian lower jaw. Proceedings of the Zoological Society of London, 140, 697–753.
Crompton, A. W. (1972). The evolution of the jaw articulation of cynodonts. In K. A. Joysey & T. S. Kemp (Eds.), Studies in vertebrate evolution (pp. 231–251). Edinburgh: Oliver and Boyd.
Crompton, A. W., & Hotton, N. I. (1967). Functional morphology of the masticatory apparatus of two dicynodonts. Postilla, 109, 1–51.
Gaupp, E. (1913). Die Reichertsche Theorie. Archive fur Anatomie und Entwicklungsgeschichte, 1912(Supplement), 1–416.
Goodrich, E. S. (1930). Studies on the structure and development of vertebrates. London: Macmillan.
Hopson, J. A. (1966). The origin of the mammalian middle ear. Americal Zoologist, 6, 437–450.
Hotton, N. I. (1959). The pelycosaur tympanic membrane and early evolution of the middle ear. Evolution, 13(1), 99–121.
Huttenlocker, A. K., & Rega, E. (2012). The paleobiology and bone microstructure of pelycosaurian-grade synapsids. In A. Chinsamy-Turan (Ed.), Forerunners of mammals: Radiation, histology, biology (pp. 91–119). Bloomington: Indiana University Press.
Ivakhnenko, M. F. (1999). Biarmosuches from the Ocher Faunal assemblage of Eastern Europe. Paleontological Journal, 33, 289–296.
Kemp, T. S. (1969). On the functional morphology of the gorgonopsid skull. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 256, 1–83.
Kemp, T. S. (1972a). The jaw articulation and musculature of the whaitsiid Therocephalia. In K. A. Joysey & T. S. Kemp (Eds.), Studies in vertebrate evolution (pp. 213–251). Edinburgh: Oliver and Boyd.
Kemp, T. S. (1972b). Whaitsiid Therocephalia and the origin of cynodonts. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 264, 1–54.
Kemp, T. S. (1979). The primitive cynodont Procynosuchus: Functional anatomy of the skull and relationships. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 285, 73–122.
Kemp, T. S. (1980). Aspects of the structure and functional anatomy of the Middle Triassic cynodont Luangwa. Journal of Zoology, 191, 193–239.
Kemp, T. S. (1982). Mammal-like reptiles and the origin of mammals. London: Academic Press.
Kemp, T. S. (2007). Acoustic transformer function of the postdentary bones and quadrate of a non-mammalian cynodont. Journal of Vertebrate Paleontology, 27(2), 431–441.
Kemp, T. S. (2009a). Phylogenetic interrelationships and pattern of evolution of the therapsids: Testing for polytomy. Palaeontologica Africana, 44, 1–12.
Kemp, T. S. (2009b). The endocranial cavity of a nonmammalian eucynodont, Chiniquodon theotenicus, and its implications for the origin of the mammalian brain. Journal of Vertebrate Paleontology, 29(4), 1188–1198.
Kemp, T. S. (2012). The origin and radiation of therapsids. In A. Chinsamy-Turan (Ed.), Forerunners of mammals: Radiation, histology, biology (pp. 3–28). Bloomington: Indiana University Press.
Kitazawa, T., Takechi, M., Hirasawa, T., Adachi, N., et al. (2015). Developmental genetic bases behind the independent origin of the tympanic membrane in mammals and diapsids. Nature Communications, 6(6853), 1–7.
Laass, M. (2014). Bone-conduction hearing and seismic sensitivity of the Late Permian anomodont Kawingasaurus fossilis. Journal of Morphology, 276, 121–143.
Luo, Z., & Crompton, A. W. (1994). Transformation of the quadrate (incus) through the transition from non-mammalian cynodonts to mammals. Journal of Vertebrate Paleontology, 14, 341–374.
Maier, W. (1990). Phylogeny and ontogeny of mammalian middle ear structures. Netherlands Journal of Zoology, 40, 55–74.
Maier, W., & van den Heever, J. (2002). Middle ear structures in the Permian Glanosuchus sp. (Therocephalia, Therapsida), based on thin sections. Mitteilungen aus dem Museum für Naturkunde in Berlin Geowissenschaftliche Reihe, 5, 309–318.
Owen, R. (1845). Description of certain fossil crania, discovered by A. G. Bain, Esq., in sandstone rocks at the south-eastern extremity of Africa, referable to different species of an extinct genus of Reptilia (Dicynodon), and indicative of a new tribe or sub-tribe of Sauria. Transactions of the Geological Society of London, 7, 59–84.
Owen, R. (1876). Descriptive and illustrated catalogue of the fossil Reptilia of South Africa in the collection of the British Museum. London: The British Museum, Natural History.
Parrington, F. R. (1946). On the cranial anatomy of cynodonts. Proceedings of the Zoological Society of London, 116, 181–197.
Parrington, F. R. (1955). On the cranial anatomy of some gorgonopsids and the synapsid middle ear. Proceedings of the Zoological Society of London, 125, 1–40.
Parrington, F. R. (1979). The evolution of the mammalian middle and outer ears: A personal view. Biological Reviews, 54, 369–387.
Reichert, K. B. (1837). Über die Visceralbogen der Wirbelthiere im allgemeinen und deren Metamorphosen bei den Vögeln und Suäugethieren, 1837. Archive für Anatomie, Physiologie, und wissenschaftliche Medizin, 1837, 120–220.
Romer, A. S., & Price, L. W. (1940). Review of the Pelycosauria. Special Papers of the Geological Society of America, 28, 1–538.
Rubidge, B. S. (1990). Redescription of the cranial morphology of Eodicynodon oosthuizeni (Therapsida: Dicynodontia). Navorsinge van die Nasionale Museum Bloemfontein, 7, 1–25.
Sidor, C. A., & Rubidge, B. S. (2006). Herpetoskylax hopsoni, a new biarmosuchian (Therapsida: Biarmosuchia) from the Beaufort Group of South Africa. In M. T. Carrano, T. J. Gaudin, R. W. Blob, & J. R. Wible (Eds.), Amniote paleobiology: Perspectives on the evolution of mammals, birds, and reptiles (pp. 76–113). Chicago: Chicago University Press.
Sigurdsen, T., Huttenlocker, A. K., Modesto, S. P., Rowe, T. B., & Damiani, R. (2012). Reassessment of the morphology and paleobiology of the therocephalian Tetracynodon darti (Therapsida), and the phylogenetic rerlationships of Baurioidea. Journal of Vertebrate Paleontology, 32(5), 1113–1134.
Takechi, M., & Kuratani, S. (2010). History of studies on mammalian ear evolution: A comparative morphological and developmental perspective. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 341B: 417–433.
Thompson, H., & Tucker, A. S. (2013). Dual origin of the epithelium of the mammalian middle ear. Science, 339, 1453–1456.
Tumarkin, A. (1955). On the evolution of the auditory conducting apparatus: A new theory based on functional considerations. Evolution, 9(3), 221–243.
van den Heever, J. (1994). The cranial anatomy of the early Therocephalia (Amniota: Therapsida). Annals of the University of Stellenbosch, 1994(11), 1–59.
Watson, D. M. S. (1952). The evolution of the mammalian ear. Evolution, 7, 159–177.
Compliance with Ethics Requirements
T. S. Kemp has declared there were no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing AG
About this chapter
Cite this chapter
Kemp, T.S. (2016). Non-Mammalian Synapsids: The Beginning of the Mammal Line. In: Clack, J., Fay, R., Popper, A. (eds) Evolution of the Vertebrate Ear . Springer Handbook of Auditory Research, vol 59. Springer, Cham. https://doi.org/10.1007/978-3-319-46661-3_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-46661-3_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-46659-0
Online ISBN: 978-3-319-46661-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)