
Abstract
In haptic perception sensory signals depend on how we actively move our hands. For textures with periodically repeating grooves, movement direction can determine temporal cues to spatial frequency. Moving in line with texture orientation does not generate temporal cues. In contrast, moving orthogonally to texture orientation maximizes the temporal frequency of stimulation, and thus optimizes temporal cues. Participants performed a spatial frequency discrimination task between stimuli of two types. The first type showed the described relationship between movement direction and temporal cues, the second stimulus type did not. We expected that when temporal cues can be optimized by moving in a certain direction, movements will be adjusted to this direction. However, movement adjustments were assumed to be based on sensory information, which accumulates over the exploration process. We analyzed 3 individual segments of the exploration process. As expected, participants only adjusted movement directions in the final exploration segment and only for the stimulus type, in which movement direction influenced temporal cues. We conclude that sensory signals on the texture orientation are used online during exploration in order to adjust subsequent movements. Once sufficient sensory evidence on the texture orientation was accumulated, movements were directed to optimize temporal cues.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others


Keywords
1 Introduction
Moving and sensing mutually influence each other in haptic perception [1, 2]. On the one hand, it is movements that generate sensations during active exploration [3]. On the other hand, humans systematically vary exploratory movements depending on the object property of interest, and, thus, depending on the required sensory input. Texture judgments are typically associated with lateral movements over surfaces [4]. Previously, texture properties were reported to be associated with changes of individual movement parameters, as the exploratory force [5–7] or movement velocity [8]. This study aims to investigate whether movement direction is a parameter, which is adjusted to the texture. We examine how texture orientation influences movement directions used over the course of exploration in a frequency discrimination task.
As for vision, we know that the direction of eye movements depends on the orientation of depicted textures [9–11]. Although, haptic perception relies even more on presence of movements than vision, haptic research did not focus on the relationship between texture orientation and movement directions so far. Finger movements can be essential for the haptic perception of textures. Striking over the textures produces temporal patterns of stimulation, i.e. temporal cues. Especially for fine-texture discrimination performance is seriously impaired without temporal cues [12]. For less fine surfaces spatial and temporal cues have been reported to be combined [13]. Gamzu and Ahissar [8] demonstrated for a frequency discrimination task that poor haptic performers can improve by using strategies which accentuate temporal cues. In a study of Lamb [14], participants explored textures, which incorporated stripes of raised dots. The spacing between stripes was either modified along the direction of the movement track or perpendicular to the movement track. Discrimination performance was better for manipulations along the track of finger movement than perpendicular to it. This performance increment can be attributed to the additional existence of temporal cues in the case of variations along the movement track. Thus, when exploration generates temporal cues the precision of texture perception can be increased.
It was previously reported that when exploration movements can be chosen freely participants aim to enhance the precision of perception [15]. Consequently, we expect that participants will choose those movements, which generate temporal cues in unconstrained texture exploration. For a spatial frequency discrimination task, haptic textures can be composed of periodic parallel grooves. Here, texture orientation is defined by the groove orientation. In this case, a movement in the direction of the texture orientation does not contain temporal cues. In contrast, every other movement direction produces temporal cues. In the special case of the movement directed orthogonally to the texture orientation, the stimulation is maximized which also leads to maximal differences in the stimulations induced by two different textures. Thus, orthogonal movement potentially optimizes the temporal cues that are useful for texture discrimination. In the present experiment we investigate whether in the discrimination of textures exploration movements are adapted to optimize temporal cues and therefore are directed against the texture orientation.
In order to adjust exploratory movements to texture orientation, sensory information about texture orientation needs to be gathered first. It is, hereby, important to consider that integration of sensory information not only occurs within one movement but also over several movements [16]. Haptic perception has been reported to be more precise with more elongated exploration [17]. Sensory signals are accumulated over the exploration. Consequently, movement adjustments based on the sensory signals are assumed to be more profound at the end of an exploration, as more sensory evidence is captured in that moment. Some studies already demonstrated that exploration movements are adjusted based on previously accumulated sensory signals. Saig and colleagues [1] reported online adjustments of movement parameters induced by sensory processing during exploratory behavior in a haptic localization task. We demonstrated motor adjustment based on sensory signals in softness exploration [2]. Here, participants applied systematically lower forces for softer objects after having gathered sensory signals on the objects softness.
In the present study we will focus on the adjustment of movement direction to texture orientation during a spatial frequency discrimination task. We expect that participants direct their exploration orthogonal to texture orientation, but not before they have accumulated sufficient sensory information from the textures. Thus, we expect that initial movements during the course of exploration are not dependent on texture orientation. In contrast, final exploration movements should be directed orthogonal to the texture orientation and, thus, optimize temporal cues to spatial frequency.
2 Experiment
We created haptic texture stimuli by 3D modeling and printing (Stratasys Objet 30 Pro). Participants explored a standard and a comparison stimulus grating and judged which of the two had a higher spatial-frequency. Standard gratings consisted of a groove pattern following the sine-wave function along one dimension (periods 1.27 and 1.44 mm; see Fig. 1). The texture pattern of comparison gratings consisted of the intersections of two orthogonal sine-wave function patterns (periods: 1.02 to 1.69 mm). A finger movement across a standard in direction of the texture orientation would generate no temporal signals on the texture’s spatial frequency. A movement orthogonal to texture orientation would be associated with optimal temporal cues. In this specific case, stimulation and differences between stimulations produced by two different patterns are maximized. Those relationships between movement direction and temporal cues do not hold for comparison stimuli. As a consequence of their construction, movements in orthogonal directions over comparison stimuli would provide similar temporal cues to spatial frequency. We manipulated the orientation of the standard and the comparison in each trial, and measured movement direction for individual strokes. We predict that movements across the standard gratings will be preferentially executed in direction orthogonal to the texture’s orientation. We expect to find this effect especially in the final movements of each single exploration, but not in the first movements.

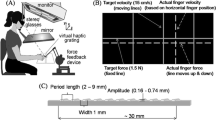
Sketch of setup and stimuli. Stimulus location and shape were visually represented on a monitor and were seen through a mirror and stereo glasses. The real grating stimuli were placed on a force sensor next to each other. The participant’s right index finger was connected to the PHANToM via an adapter. The PHANToM measured the finger position.
2.1 Participants
The sample consisted of thirteen right-handed participants (average age: 25.4 years, range: 19–32 years; 7 females). All were naïve to the purpose of the experiment and were paid for participating. None of them reported sensory or motor impairments or recent injuries of the right index finger. We tested two-point-discrimination thresholds at the finger pad of the right index finger. All participants had a two-point discrimination threshold of 3 mm or lower. Participants gave written informed consent and the study was conducted in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
2.2 Apparatus and Stimuli
Participants sat in front of a custom-made visuo-haptic workbench (see Fig. 1), which comprised a PHANToM 1.5A haptic force feedback device, force sensor (682 Hz, resolution: 0.05 N) and a 22″-computer screen (120 Hz, 1024 × 1280 pixel). The right index finger was connected to the PHANToM via an adapter, which allows for free finger movements having all six degrees of freedom in a 38 × 27 × 20 cm3 workspace. The PHANToM device was only used to measure finger position. The adapter was connected with double-faced adhesive tape to the nail, which left the finger pad bare. Simultaneously, the participants looked through stereoglasses and via a mirror onto the screen (40 cm viewing distance). The mirror prevented participants from seeing their hand or the real stimuli and enabled spatial alignment of the 3D-visual representation with the haptic display. The participants’ heads were stabilized by a chinrest. A custom-made software controlled the experiment, collected responses, and recorded the data from the PHANToM and the sensor with recording intervals of 3 ms. Headphones and ear plugs masked sounds from haptic exploration. The stimuli were presented next to each other in front of the participant. They were placed on the force sensor, which measured the executed finger force. The stimuli were displayed in virtual 3D-scene as three dimensional cylindrical discs with a border. The visual representation did not display the texture pattern or orientation. Position and size of the ‘visual’ stimuli corresponded to those of the real ones. Outside of the stimulus area present finger position was visible as a small sphere (8 mm diameter).
Haptic grating stimuli were created using the OpenSCAD software and 3D printing. The 3D printer (Objet 30 Pro, Stratasys Ltd., United States) creates arbitrary 3D objects from 3D digital data. In this method 3D objects are build drop wise with support and model photopolymer material (VeroClear) having a build resolution of 600 × 600 × 1600 dpi (x-, y-, z-axis). The stimuli were 4 mm high (z-axis) grating discs with a texture diameter of 90.7 mm and a total diameter including the border of 100.7 mm. A 10 × 5 mm grip indicated the texture orientation for the experimenter (see Fig. 1). We created two types of stimuli, standard and comparison gratings. Standard gratings consisted of a groove pattern following the sine-wave function. Texture height was defined as a sine-wave function with the peak amplitude (A) of 0.3 mm, see Eq. 1, depending only on one of the other two dimensions. We defined two standard stimuli with the periods (P) of 1.27 mm and 1.44 mm. For comparison stimuli, we computed texture height from two overlaid sine-wave functions that were oriented perpendicular to each other. The intersection of both textures defined the comparison. Thus, the texture height was at each point the minimum of the two functions, see Eq. 2. A cut through two orthogonal axes of comparison stimuli would result in identical images (see Fig. 1). We created 5 comparison gratings with periods of 1.02, 1.19, 1.35, 1.52 and 1.69 mm. For each of the two standards we used three comparisons. Two comparisons were defined by ±20 % of the standard’s period, because 20 % corresponds to the Weber fraction in active touch (as assessed e.g. from [18]). The third comparison was the same stimulus for both standards (1.35 mm); it has 6 % lower period than the standard of 1.44 mm and 6 % higher period than the standard of 1.27 mm. Consequently, the frequency comparisons should be too difficult for static touch only. Based on the stimulus construction we defined texture orientation in standard gratings as the orientation of the parallel grooves. By definition, comparison gratings had two equal groove orientations. In the following, we will refer to one of them as the texture orientation (see Fig. 1).
2.3 Design and Procedure
In each trial a stimulus pair, consisting of one standard and one comparison stimulus, was explored. The participant judged which of the two had a higher spatial-frequency regardless of other differences between the textures. We explained spatial frequency as the number of experienced grooves over a certain distance. Thus, stimuli with a higher period have lower spatial frequencies. We manipulated the standard stimulus (period of 1.27 or 1.44 mm) and the orientation of the stimulus pair on the force sensor (15°, 45°, 75°, 105°, 135°, and 165°; in Fig. 1 depicted in the orientation of 75°). The dependent variable was the movement direction used over standard and comparison grating. We focused on the first, middle and last stroke, as they represent movement adjustments at different moments during exploration regardless of the exact stroke number in a trial.
Each standard was paired with one of three comparisons (standard 12.7 mm with comparisons with periods of 1.02, 1.35, 1.52 mm and standard 1.44 mm with 1.19, 1.35, 1.69 mm). The standard grating was either presented at the right or the left side. Standard and comparison grating were always placed in the same orientation. Note that due to its definition the comparison grating felt the same when being explored along or against its orientation and also stimulation from explorations along oblique axes was only moderately different in temporal frequency (factor below 1.4). Additionally, to keep motivation high over the entire experiment, we introduced the experiment as a game. Participants could earn 10 or 100 points with a correct answer. Overall, there were 2 [standards] × 3 [comparisons] × 6 [orientations] × 2 [standard left or right] × 2 [10 or 100 points] = 144 trials. The order of trials was randomized and trials were presented in 3 successive blocks of 48 trials. Participants were instructed to take a break of at least two minutes between each two blocks. The experiment lasted 2–3 h.
Before starting the exploration of a trial, the screen indicated how many points a correct response would correspond to (10 or 100). The exploration randomly started either with the right or the left stimulus on a random start position at the border of this stimulus (20°–350°, in steps of 30°). Then, participants were free to perform as many strokes and to switch as often between stimuli as they wanted. During the exploration no visual information about the stimulus was given. The basic payment was 16€. An additional euro was gained, whenever the participant accumulated 500 points. A visual and auditory feedback was given 1–3 trials after the points had been accumulated. Pure guessing would have led to a total payment of approximately 23€ and perfect task performance to a total of about 31€.
2.4 Data Analysis
We segmented the exploration behavior on each stimulus into single strokes. A stroke was defined as a continuous movement over the texture in one direction. We analyzed 3 strokes (first, middle, last) of the exploration of each stimulus. In case of even numbers of strokes, the middle stroke was defined by the later one of the two possible. For the definition of a stroke we considered those parts of the exploration, in which the finger was touching the stimulus area with at least 0.1 N of force. We detected strokes as continuous movements either from one texture border to another or between two movement turns, which we extracted by zero crossings in y- or x-velocity. For each stroke we derived its movement direction and duration (restricted to minimum duration: 200 ms). Only those trials entered in the analysis, in which the participant at least performed two strokes on each grating. In order to collapse data over trials, we aligned all stimulus orientations with an orientation of 0°. Therefore, stroke directions were rotated by their corresponding texture orientation in the opposite direction. As strokes differed highly in their duration, we weighted individual strokes with their duration. Technically, strokes were duplicated in accordance to their duration. Based on this data we calculated histograms of movement directions (bin size: 15°) for each participant, grating and exploratory segment. The resulting histograms represented which proportion of exploration time one participant followed a specific direction for this stroke (first, middle or last) in this grating (standard or comparison). In an overall participant analyses we computed average histograms to keep statistical power equal with the individual participant analyses. We conducted circular statistics on these binned data separately for each exploratory segment (first, middle, last) and grating type (standard, comparison). For this purpose, we used the V-test, a variant of the Rayleigh test, which tests the alternative hypothesis that the population is not distributed uniformly around the circle but has a specified mean direction [19], 90° in our case. For standard gratings, we predicted that over the course of exploration movement directions should get more and more non-uniformly distributed. Thus, we expect significant values especially for the last stroke in the exploration of the standard.
3 Results
3.1 Exploration and Task Performance
On average, participants performed 3.9 strokes (SD = 1.8) on the standard grating and 3.7 strokes (SD = 1.7) on the comparison grating, and they switched twice between the stimuli (M = 2.05, SD = 0.82): once from first to the second stimulus and then once back to the first stimulus. An average stroke took 1.6 s (SD = 0.6) for the standard grating and 1.7 s (SD = 0.6) for the comparison grating. Participants used more strokes for the exploration of standard gratings, t(12) = 2.26, p = 0.04, while stroke duration was higher for comparison gratings, t(12) = −3.04, p = 0.01. On average participants gave 59.2 % correct answers (SD = 8 %), which is significantly higher than guessing (50 %), t(12) = 3.88, p < 0.01.
3.2 Movement Directions
The angular distributions of movement directions for the first, middle and last stroke exploring the standard or the comparison grating are plotted for all participants in Fig. 2. We calculated the V-test based on the binned data presented in Fig. 2. This test reports V-values, which increase with the deviation of the empirical distribution from a uniform distribution and with the consistence of the empirical mean direction with a predicted one. A non-significant test could either be due to a uniform distribution or a distribution with a mean that deviates from the predicted direction of 90°. For the first stroke V-values were not significant for both gratings (standard: V = −6.15, p = 0.82; comparison: V = 0.59, p = 0.47). Also in the middle stroke values were not significant, although Fig. 2 appears to indicate a tendency in the standard stimulus (standard: V = 6.93, p = 0.16; comparison: V = −1.23, p = 0.57). As predicted in the last stroke participants showed a significant non-uniformity in their movement directions and moved orthogonally (90°) to the standard grating, V = 17.85, p < 0.01. For the comparison stimulus this non-uniformity did not reach significance in the last stroke, V = 7.19, p = 0.15. This result is well reflected in the individual participant analyses. We found significant movement adjustments to the texture orientation for 9 of 13 participants. None of the participants significantly adjusted the first stoke to the texture orientation. Additionally, no one showed more significant adjustments in the comparison than in the standard. The data of two participants revealed the same pattern as the average data. Four participants significantly adjusted their middle and last stroke to the standard and only one of the strokes to the comparison. Three participants showed significant adjustments to standard and comparison (2 in middle and last stroke; 1 in last stroke only). Taken together, adjustments of movement direction were not found in the beginning of the exploration. However, for standard gratings, the last stroke was adjusted to move orthogonal to the texture orientation. So that in this case 25 % of the movements had a direction between 75°–105°.

Movement direction histograms including all participant data for each exploration segment and texture type separately. All textures were aligned to a 0° orientation. Possible movement directions varied between 0–180° and were mirrored on the lower part of the figure.
4 Discussion
We investigated whether movement directions are changed over the course of natural exploration in order to move orthogonally to a texture. In our experiment participants compared the spatial frequency of two types of gratings. For one of the two gratings moving along the texture orientation produced only spatial cues and no temporal cues on spatial frequency. In contrast, moving orthogonally to the texture orientation produced optimal temporal cues. This simple relationship between movement and texture orientation did not hold for the second grating type, where two orthogonal moving directions resulted in the same cues. As predicted, participants changed their movement direction across the oriented textures over the course of exploration. In the initial segments of exploration movement directions were not dependent on texture type or orientation, but rather uniformly distributed. However, participants moved orthogonally to the textures of the first grating type in final exploration segments. Thus, sensory signals about texture orientation seem to be taken into account for the following control of exploration movements. Movements were chosen to produce improved temporal cues, when the availability of cues depended on the movement direction.
According to our data profound changes in movement direction only appeared in the final segment of exploration. In the middle stroke, and therefore after at least one stroke with an average duration of more than 1.5 s, movement directions were not significantly aimed towards orthogonal exploration. This finding is especially interesting in light of a recent study [20], which described that edge orientation is already coded by the intensity and temporal structure of the responses of first-order tactile neurons. However, our result is very unlikely to be attributed to the duration of motor responses, as those were demonstrated to take only 100–200 ms [21]. Thus, although sensory signals are available early on, we think that within finger movements signal reliability is low. Presumably, temporal signal integration processes are necessary to produce an as reliable estimate of texture orientation as required for movement adjustments.
The results revealed the expected pattern, in which movements were chosen orthogonally to the texture orientation only in the case of a unique orientation (i.e. for standard gratings) and only in the final exploration segments. From the first sight the remaining 5 experimental cases seemed not to look highly uniform in their distribution. In order to control for possible directional preferences other than the predicted one we computed simple Rayleigh tests for these cases. None of the 5 distributions showed a significant non-uniformity, z ≤ 1.80, p ≥ 0.16. Correspondingly, our results primarily reflect the expected bottom-up effects on movement direction. However, there are also some hints that chosen movement directions might not only be based on bottom-up stimulus information. In our comparison gratings a movement in 0° direction and a movement orthogonal to that (90°) generated precisely the same cues. However, as it can be seen from Fig. 2 movements in these two directions (90° ± 30° vs. 0° ± 30°) are not represented equally often (37 % vs. 28 %) during the final stroke. This difference shows a statistical trend, t(12) = 1.96; p = 0.06 in a paired t-test on the rationalized arcsine transformation of the individual proportional data. Perhaps, the 90° direction was chosen more often, in order to move in a similar way as in the standard grating of the same trial. This observation indicates that the task to compare two stimuli has a top-down influence on movement directions and fits well to recently reported results on movement control [22].
As we reported in the results about 25 % of the last movements were at directions ±15° from orthogonality. Even about 48 % of the last movements were at directions ±30° from orthogonality. We assumed that orthogonal movements maximize temporal cues and therefore optimize sensory input. Based on our data we can perform post-hoc analyses to compare whether the movements approximately orthogonal to the texture orientation (±30°) led to better task performance, measured by the proportion of correct responses, than other movements. A paired-sample t-test on the rationalized arcsine transformation of the individual proportions of correct responses reveals significantly better performance for trials with approximately orthogonal movement direction, t(12) = 2.43, p = 0.03 in comparison to other movements. Both groups of trials did not differ in their average number of strokes used, t(12) = −0.05, p = 0.96. However, it needs to be considered that this experiment was not designed for this analysis. We plan future experiments where exploration length and movement direction are manipulated independently to better address this question. We will also address the question of how much the previous movement direction influenced the following movement.
Our initial expectation of directing movements orthogonal to the texture orientation was based on the idea, that orthogonal movement optimizes temporal cues. However, an alternative explanation for choosing orthogonal movement direction could be that participants tried to avoid exploration movements in line with the texture that generate no temporal cues on spatial frequency. If our system takes into account motor and perceptual noise, than aiming for orthogonal movements would maximize the chance to avoid exploration in line with the texture. In order to distinguish between both possibilities, future research could use more complex virtual textures, in which the movement direction generating maximal temporal stimulation is not orthogonal to the direction generating no temporal cues.
5 Conclusion
This study asked the fundamental question of whether texture orientation influences movement directions used in a frequency discrimination task. We focused on natural and unconstrained exploration of two types of textures, one with and one without a unique orientation. Participants adjusted movement direction over the course of exploration depending on the texture they encountered. In the first exploration segment movements were uniformly distributed along all directions regardless of the texture type. However, in cases in which the texture had a unique orientation, movement directions changed until the final exploration segment. In the last stroke participants moved orthogonally to the texture orientation. This result indicates that sensory signals on texture orientation were accumulated over the course of exploration and influenced motor control. We suggest that the reason for this adjustment is the optimization of sensory cues needed to perform the task. Taken together, this study strongly supports the idea of a sensorimotor control loop. Sensory signals are used to perform those movements, which optimize the sensory feedback itself.
References
Saig, A., Gordon, G., Assa, E., Arieli, A., Ahissar, E.: Motor-sensory confluence in tactile perception. J. Neurosci. 32(40), 14022–14032 (2012)
Lezkan, A., Drewing, K.: Predictive and sensory signals systematically lower peak forces in the exploration of softer objects. In: 2015 IEEE World Haptics Conference (WHC), pp. 69–74. IEEE, June 2015
Gibson, J.J.: The Senses Considered as Perceptual Systems. Houghton-Mifflin, Boston (1966)
Lederman, S.T., Klatzky, R.L.: Hand movements: a window into haptic object recognition. Cogn. Psychol. 19, 342–368 (1987)
Lederman, S.J.: Tactile roughness of grooved surfaces: the touching process and effects of macro- and microsurface structure. Percept. Psychophys. 16, 385–395 (1974)
Gibson, G.O., Craig, J.C.: The effect of force and conformance on tactile intensive and spatial sensitivity. Exp. Brain Res. 170, 172–181 (2006)
Nefs, H.T., Kappers, A.M.L., Koenderink, J.J.: Frequency discrimination between and within line gratings by dynamic touch. Percept. Psychophys. 64, 969–980 (2002)
Gamzu, E., Ahissar, E.: Importance of temporal cues for tactile spatial-frequency discrimination. J. Neurosci. 21, 7416–7427 (2001)
Wismeijer, D.A., Gegenfurtner, K.R.: Orientation of noisy texture affects saccade direction during free viewing. Vis. Res. 58, 19–26 (2012)
Wismeijer, D.A., Erkelens, C.J., Van Ee, R., Wexler, M.: Depth cue combination in spontaneous eye movements. J. Vis. 10(6), 25 (2010)
Wexler, M., Ouarti, N.: Depth affects where we look. Curr. Biol. 18(23), 1872–1876 (2008)
Hollins, M., Risner, S.R.: Evidence for the duplex theory of tactile texture perception. Percept. Psychophys. 62(4), 695–705 (2000)
Cascio, C.J., Sathian, K.: Temporal cues contribute to tactile perception of roughness. J. Neurosci. 21(14), 5289–5296 (2001)
Lamb, G.D.: Tactile discrimination of textured surfaces: psychophysical performance measurements in humans. J. Physiol. 338(1), 551–565 (1983)
Kaim, L., Drewing, K.: Exploratory strategies in haptic softness discrimination are tuned to achieve high levels of task performance. IEEE Trans. Haptics 4(4), 242–252 (2011)
Henriques, D.Y., Soechting, J.F.: Approaches to the study of haptic sensing. J. Neurophysiol. 93, 3036–3043 (2005)
Drewing, K., Lezkan, A., Ludwig, S.: Texture discrimination in active touch: effects of the extension of the exploration and their exploitation. In: 2011 IEEE World Haptics Conference (WHC), pp. 215–220. IEEE, June 2011
Nefs, H.T., Kappers, A.M., Koenderink, J.J.: Amplitude and spatial-period discrimination in sinusoidal gratings by dynamic touch. Perception 30(10), 1263–1274 (2001)
Mardia, V., Jupp, P.: Directional Statistics, 2nd edn. Wiley Ltd., London (2000)
Pruszynski, J.A., Johansson, R.S.: Edge-orientation processing in first-order tactile neurons. Nat. Neurosci. 17, 1404–1409 (2014)
Johansson, R.S., Westling, G.: Signals in tactile afferents from the fingers eliciting adaptive motor responses during precision grip. Exp. Brain Res. 66(1), 141–154 (1987)
Callier, T., Saal, H.P., Davis-Berg, E.C., Bensmaia, S.J.: Kinematics of unconstrained tactile texture exploration. J. Neurophysiol. 113(7), 3013–3020 (2015)
Acknowledgements
We would like to thank Maria Ebsen and Claire Weyel for their help in collecting the data and Steffen Bruckbauer for help with the figures. This research was supported by the German Research Foundation (DFG; grant SFB/TRR135/1, A05).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this paper
Cite this paper
Lezkan, A., Drewing, K. (2016). Going Against the Grain – Texture Orientation Affects Direction of Exploratory Movement. In: Bello, F., Kajimoto, H., Visell, Y. (eds) Haptics: Perception, Devices, Control, and Applications. EuroHaptics 2016. Lecture Notes in Computer Science(), vol 9774. Springer, Cham. https://doi.org/10.1007/978-3-319-42321-0_40
Download citation
DOI: https://doi.org/10.1007/978-3-319-42321-0_40
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-42320-3
Online ISBN: 978-3-319-42321-0
eBook Packages: Computer ScienceComputer Science (R0)