
Abstract
Animal species with fission–fusion dynamics are characterized by variable group size and composition, as individuals move freely among a social network. Patterns consistent with fission–fusion dynamics appear to be widespread in bats and interest in these patterns has advanced our general understanding of proximate and ultimate explanations for fission–fusion dynamics. Here, we explore the following questions: (1) Why do bats switch roosts? (2) Why do not group members move together? (3) Do bats form social bonds despite frequent roost switching? (4) If group members do form social relationships, what shapes these relationships? (5) How are social relationships maintained in these dynamic systems? (6) What does our understanding about fission–fusion in bats tell us about the evolution of fission–fusion dynamics in general? In our review, we show that bats switch roosts for a combination of reasons, including to reduce predation, parasites, and disease, and to minimize commutes. The strongest evidence, however, suggests bats switch roosts to seek more suitable roosts when roost quality changes. However, the degree to which each of these factors influences roost switching varies between individuals, across species, and depends on preferred roost type and roost availability. Group members may not move together because they experience different perceived costs and benefits of remaining in a group. Bats apparently use a combination of spatial and sensory information to relocate group members and maintain social bonds. Bat populations with fission–fusion dynamics likely benefit from social thermoregulation, and share information about roosts and foraging sites. Although our understanding of these dynamics has improved in recent years, many questions remain. For instance, a better understanding of the proximate factors driving fission–fusion dynamics is required. Establishing the levels at which bats sharing the same roost interact, together with network analyses, would also advance our understanding of relationships within these dynamic groups. Together, answers to these and other questions will help us to better manage bat populations and their habitat.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Social systems, defined by group size, stability, composition, and interactions among group members (Hinde 1976), vary along a continuum. At one end of this continuum are the eusocial insects and mammals. With the exception of rare dispersal events, group size and composition vary only with births and deaths of group members in eusocial species, as the costs of dispersal are high. Consequently, individuals interact regularly, which favors cooperation among familiar group members. At the same time, group members are typically very closely related, favoring nepotism, where individuals preferentially cooperate with relatives. Ultimately, these conditions have favored altruism, and most individuals forego their own reproductive efforts and instead care for the offspring of a few breeding group members (Choe and Crespi 1997; Burda et al. 2000; Bourke 2014).
At the other end of the continuum are fission–fusion societies, where larger groups periodically split into multiple smaller groups that later regroup, or, in other instances, individuals move freely among a network of multiple social groups (reviewed in Aureli et al. 2008). Despite the dynamic nature of these groups, individuals interact nonrandomly, with some group members spending more time together than with others. In some cases, group members who interact regularly are also related, but relatedness is often highly variable within groups. Group members have the opportunity to engage in social behaviors, such as cooperation and nepotism, and therefore these groups represent more than passive aggregations around common resources. Although fission–fusion dynamics are assumed to be relatively widespread, they have been difficult to quantify making them less well understood compared to more stable social systems (reviewed in Aureli et al. 2008).
Nevertheless, fission–fusion dynamics have been quantified for a variety of primates, cetaceans, and ungulates (Aureli et al. 2008), with most of our understanding coming from studies of a few well-documented species, including African elephants (Loxodonta africana; Wittemyer et al. 2005; Archie et al. 2006), bottlenose dolphins (Tursiops truncatus; Connor et al. 2000; Lusseau et al. 2006), and chimpanzees (Pan troglodytes; Lehmann and Boesch 2004). Evidence from these species supports theoretical predictions that fission–fusion dynamics, like other social systems, are influenced by life history, reproductive strategy, phylogeny, and resource stability (Clutton-Brock and Janson 2012). For instance, fission–fusion has evolved primarily in long-lived animals with high offspring investment, high female philopatry, and where individuals occupy environments that vary within their lifetimes (Aureli et al. 2008; Couzin and Laidre 2009). At the same time, because related and unrelated group members interact repeatedly, cooperative behaviors, such as social grooming, cooperative offspring care, cooperative feeding, and group defense, have evolved in these seemingly random systems (Aureli et al. 2008).
Bats are also long-lived animals with high offspring investment, and many species show high female natal philopatry and occupy moderately variable environments (Kunz and Fenton 2003). Studies over the past 15 years have demonstrated that several species also live in fission–fusion societies where group members cooperate (Kerth 2008a). Fission–fusion in bats generally refers to the group dynamics of individuals found roosting together during the day, where subgroups or individual group members switch among roosts, leading to changes in group size and composition. In the neotropics, for example, females typically live in relatively stable year-round harems with one or two resident males, but they also move periodically. For example, female short-nosed bats (Cynopterus sphinx) switch as infrequently as every 4 years (Storz et al. 2000), while Spix’s disk-winged bats (Thyroptera tricolor) may switch roosts (harems) almost daily (Vonhof et al. 2004). In temperate regions group dynamics are highly seasonal and much more fluid. There, following hibernation, females form summer groups where they give birth and raise their offspring while males are typically solitary or live in small all-male groups. Throughout the summer, females switch roosts, but not as a cohesive group, resulting in variable group size over time. As a result, a colony consists of multiple social groups interconnected by the movement of group members between groups (Patriquin et al. 2010; Kerth and Van Schaik 2011).
While roost switching (or roost fidelity) is widely documented in bats (see Lewis 1995 and Barclay and Kurta 2007 for reviews), group size, stability, composition, and social relationships within groups have been quantified for only a few species, and most of our understanding comes from temperate vespertilionids (Table 4.1). Nevertheless, work with these species has revealed several trends, which we will draw on to answer the following questions: (1) Why do bats switch roosts? (2) Why do not group members switch roosts together? (3) Do bats form social bonds despite the fluid dynamics of fission–fusion? (4) If so, what shapes these bonds and how long do they last? (5) Is there evidence of cooperation and/or nepotism? From there, we will identify gaps in the current knowledge about fission–fusion in bats and offer suggestions for future directions. By examining fission–fusion in bats, we will then determine whether the factors shaping these systems are similar to those in other taxa, in turn providing further insight to the evolution of fission–fusion dynamics.
Fission–fusion dynamics in various taxa, including bats, has been discussed to varying degrees elsewhere (Couzin 2006; Aureli et al. 2008; Kerth 2008a; Couzin and Laidre 2009); our goal here is to synthesize and expand on previous discussions to provide a single, cogent review of our current knowledge with an aim to identify existing gaps and future directions. Our review will focus on sociality among females as much less is known about sociality in males (but see Safi 2008; August et al. 2014). Also, we will focus on dynamics in day roosts as little is known about group dynamics of bats in night roosts, with the exceptions of the false vampire bat (Megaderma lyra; Schmidt 2013) and the white-striped free-tailed bat (Tadarida australis; Rhodes 2007).
1 Why Do Bats Switch Roosts?
Roost switching may be costly (Lewis 1995; Ellison et al. 2007); therefore it likely reflects an individual’s attempt to balance the costs and benefits of group living (Lewis 1995; Kashima et al. 2013). For instance, bats appear to select roosts that offer protection from potential predators and that provide optimal conditions for thermoregulation (reviewed in Kunz and Lumsden 2003 and Barclay and Kurta 2007). At the same time, many species may live in groups, at least in part, to minimize thermoregulatory costs through group warming (Willis and Brigham 2007), but, as a result, may incur the costs of increased conspicuousness to predators and increased risk of transmission of diseases and parasites (Krebs and Davies 1995). Of course, living in social groups may also confer less tangible benefits, such as cooperation and nepotism (Carter and Wilkinson 2013a, b). As such, several explanations have been proposed for roost switching, none of which are necessarily mutually exclusive, including antipredation, reducing ectoparasite loads, reduced commuting distances between food and roosts, and change in roost quality (Barclay and Kurta 2007). We consider several plausible explanations in turn below.
1.1 Antipredation
As discussed in a recent review of the influence of predators on bat behavior, reduced predation risk is an often cited—but rarely tested—explanation for roost switching by bats (Lima and O’Keefe 2013). Certainly aerial and terrestrial predators attack bats emerging from roosts, and some predators have developed specialized hunting strategies for bats (Ancillotto et al. 2013; Mikula et al. 2013; Michaelsen et al. 2014; reviewed in Lima and O’Keefe 2013). Correspondingly, bats may select roosts, in part, to provide them protection from predators (reviewed in Kunz and Lumsden 2003 and Barclay and Kurta 2007), suggesting to some researchers that bats are likely “sensitive to day-to-day variation in the risk of predation” (Lima and O’Keefe 2013, p. 640). Moreover, a range of predators visit occupied roosts (simulated by placing fresh guano in roosts) more often than unoccupied roosts (Threlfall et al. 2013), supporting the idea that roost switching may be an effective antipredator strategy. Indeed bats leave roosts when directly attacked but there is little evidence that they respond to visual, acoustic, or olfactory cues of predators (Michaelsen et al. 2014; reviewed in Lima and O’Keefe 2013). Interestingly, Lima and O’Keefe (2013) argue that temperate bats are least likely to be influenced by predators, yet roost switching in bats is most common in these regions. Predation risk, by itself, does not appear sufficient to explain frequent roost switching.
The relative influence of predation risk on roost switching may vary between individuals and across species, and depends on roost type and the availability of alternative suitable roosts (reviewed in Lima and O’Keefe 2013). For several species, including little brown bats (Myotis lucifugus), pallid bats (Antrozous pallidus), Indiana bats (M. sodalis), and northern long-eared bats (M. septentrionalis), individuals may remain in a roost while other group members leave, and would therefore presumably still be at risk of predation (e.g., Barclay 1982; Lewis 1996; Carter and Feldhamer 2005; Patriquin et al. 2010), indicating that the relative costs and benefits of switching vary across individuals (reviewed in Barclay and Kurta 2007). Bats often show higher fidelity to man-made roosts, such as buildings and bat boxes (Lewis 1995; Kunz and Lumsden 2003; Evelyn et al. 2004; Lausen and Barclay 2006; Evans and Lumsden 2011), that are typically warmer and more stable and therefore result in higher offspring survival compared to natural roosts (Lausen and Barclay 2006). This despite some evidence suggesting predation risk is in fact higher at these structures (Michaelsen et al. 2014). Lausen and Barclay (2006), on the other hand, suggest bats in buildings are less vulnerable to terrestrial predators compared to bats in rock crevices, but these authors did not consider aerial predators. It appears, then, bats may trade-off predation risk in favor of optimal roost conditions (reviewed in Lima and O’Keefe 2013).
1.2 Minimize Commuting Distances Between Foraging and Roosting Sites
As Kunz and Lumsden (2003) point out, bats may switch roosts to be closer to feeding sites to minimize the costs of travel, yet there is little support for this hypothesis. Certainly, bats generally roost in areas closer to foraging habitat compared to randomly available roosts (Kingston 2013; Trevelin et al. 2013) and Lewis (1995) found several frugivorous species switch roosts to be closer to patches of fruit. Similarly, at least two insectivorous species, common pipistrelles (Pipistrellus pipistrellus) and soprano pipistrelles (P. pygmaeus), appear to switch to roosts that minimize commuting costs as distances between roosts and foraging sites are less than distances between consecutive roosts (Feyerabend and Simon 2000; Bartonička et al. 2008). However, when they moved to buildings during lactation, roosts were farther from their main foraging site (Bartonička et al. 2008), suggesting roost switching is not always driven by commuting costs. Indeed reviews of roost fidelity within roost areas have demonstrated that distances between roosts are typically less than distances between roosts and foraging sites, suggesting bats are not moving to be closer to foraging sites (Lewis 1995; Kunz and Lumsden 2003). In fact, distances traveled by common vampire bats (Desmodus rotundus) between roosts and foraging sites are often greater than between consecutive roosts (Wilkinson 1985) while no difference in distance between roosting and foraging sites was found for Hemprich’s long-eared bats (Otonycteris hemprichii) roosting in rock crevices (Korine et al. 2013). Female Bechstein’s bats (Myotis bechsteinii) found roosting together travel different distances to forage, with some traveling much farther than others (Kerth et al. 2001a, b), suggesting that in this species roost choice is not based on proximity to foraging sites, or at least not for all group members. Thus evidence that bats switch roosts to be closer to foraging sites is equivocal.
The relative importance of switching roosts to minimize commuting costs may vary with factors such as reproductive condition, roost type (and in turn availability), and diet. Soprano pipistrelles, among others, select roosts closer to foraging sites during lactation compared to postweaning (Bartonička et al. 2008), presumably because the costs of commuting during lactation are higher as females often return to roosts throughout the night to nurse their young (Kunz and Lumsden 2003). Some soprano pipistrelles move during lactation from bat boxes near their main foraging site during gestation to buildings farther away, which presumably offer different microclimates needed during lactation (Bartonička et al. 2008). Independent of reproductive condition or roost type, species like Hemprich’s long-eared bats and Bechstein’s bats may not benefit from attempting to roost closer to their prey that typically have unpredictable distributions, such as vertebrates or insects that do not swarm. Species like the Jamaican fruit-eating bat (Artibeus jamaicensis) may move closer to patchy, more predictable food, such as fruit or insect swarms (Barclay and Kurta 2007). Tent-making bats, which are capable of constructing roosts rather than relying on existing structures like trees, rocks, and buildings, could theoretically construct/switch roosts to be closer to foraging sites; this, however, has yet to be tested and evidence that bats are actually more likely to switch among relatively more permanent tents (Sagot and Stevens 2012) suggests tent-making and switching is not related to minimizing commuting distances. When examining factors that influence roost switching, and fission–fusion dynamics, it is important to consider species and population level differences in sex, reproductive condition, roost type, and diet.
1.3 Ectoparasites
Ectoparasites can impose significant costs to their hosts (Lourenço and Palmeirim 2007); as such, high parasite loads associated with a given roost may prompt bats to switch. A theoretical model demonstrates that bats may receive higher fitness benefits if they trade-off optimal roosts for lower ectoparasite loads (Kashima et al. 2013), which they can achieve by leaving roosts thus disrupting some parasites, particularly bat flies (Families Streblidae and Nycteribiidae), that must complete part of their life cycles on the roost substrate rather than on the host (Dick and Dittmar 2014). Empirical evidence suggests that the timing of roost switching by Bechstein’s bats, common pipistrelles, and soprano pipistrelles, as examples, coincides with peak infestations and that switching reduces ectoparasite numbers by as much as half (Reckardt and Kerth 2006; Bartonička and Gaisler 2007; Bartonička and Růžičková 2013). Moreover, it has been reported that pallid bats and Gould’s wattled bats (Chalinolobus gouldii) with higher ectoparasite loads switch roosts more often than those with lower parasite loads, suggesting bats leave roosts to escape high infestations (Lewis 1996; Evans and Lumsden 2011). Others argue that higher ectoparasite loads on Bechstein’s bats that switch roosts less frequently is evident that roost switching is effective at reducing ectoparasites (e.g., Reckardt and Kerth 2007). Interestingly, Bechstein’s bats appear to use information about prior roost occupancy to minimize their risk of reinfection (Reckardt and Kerth 2007). When provided a paired choice between new roost boxes and previously occupied, and thus potentially infested, roosts, Bechstein’s bats preferred new roosts. Moreover, when given a choice between recently occupied roosts and roosts occupied in previous years, Bechstein’s bats preferred older roosts that were less likely to be infested. Roost switching therefore appears to be an effective means for reducing parasite loads.
The relationship between roost switching and ectoparasite load is likely complicated by the kinds of ectoparasites involved. For instance, although bat flies move between bats via the roost substrate where they must complete part of their life cycle, mites move through direct contact between bats where they remain for their entire life cycle (Christe et al. 2000; Dick and Ditmar 2014). Also, unlike some mites, bat flies do not appear to elicit a grooming response (Godinho et al. 2013). It is therefore argued that roost switching is the only effective means for reducing the prevalence of bat flies but is unlikely to affect mites (e.g., ter Hofstede and Fenton 2005; Reckardt and Kerth 2007). However, roost microclimate affects both bat fly and mite survival (Bartonička and Gaisler 2007; Pearce and O’Shea 2007; Bartonicka 2010; Postawa et al. 2014). Thus roost switching may also serve to interfere with mite populations by subjecting them to fluctuating environmental conditions.
Roost microclimate and thus roost switching are also affected by roost structure, together with changes in ambient conditions and reproductive condition of bats. For example, conditions inside caves are typically more stable compared to buildings, which are in turn more stable compared to tree and rock cavities or foliage that vary in accord with ambient conditions (Kunz and Lumsden 2003; Postawa and Gas 2009). Correspondingly, temperate bats roosting in buildings switch roosts less frequently and often have higher parasite loads than do bats roosting in natural cavities, which in turn switch less frequently and have higher parasite loads than bats roosting in foliage (Lewis 1995; ter Hofstede and Fenton 2005; Patterson et al. 2007). Interestingly, black mastiff bats (Molossus rufus) roosting in buildings have lower ectoparasite loads compared to those roosting in natural tree hollows, which the authors attribute to lower contact rates in buildings where more space is available (Esbérard et al. 2005). Independent of roost structure, susceptibility to ectoparasites may also vary across the season with physiology, age, and reproductive condition (Pearce and O’Shea 2007). Many ectoparasites are less likely to attach to torpid bats (Montes et al. 2002; Turbill and Geiser 2006) and do not thrive in high temperatures (Bartonička and Gaisler 2007). Bat immunity also varies with reproductive condition (Christe et al. 2000); in particular, females may be less likely to groom themselves while nursing (Christe et al. 2007), and hairless young are more vulnerable than individuals with hair (Lewis 1995). Studies should therefore consider the time of year and reproductive condition of bats when drawing conclusions about roost-switching behaviors.
The relationship between roost-switching and ectoparasite loads is likely to be further complicated by phylogeny, group size, and species-specific behavior. Closely related bat species may be more likely to host similar ectoparasites and use similar roosts, thereby potentially exaggerating the relationship between roost-switching and ectoparasite loads (ter Hofstede and Fenton 2005). However, evidence that conspecifics using different types of roosts (e.g., big brown bats, Eptesicus fuscus, use tree cavities, rock cavities, and buildings) have different ectoparasite loads (Patterson et al. 2007; reviewed in Lewis 1995) and different rates of roost switching (reviewed in Lewis 1995) suggests the connection between ectoparasites and roost switching is not simply the result of phylogenetic inertia. Instead, differences in parasite loads across species and conspecific groups in different roosts may reflect group size. Transmission rate may be higher in larger groups typical of caves and buildings compared to smaller groups in foliage roosts (ter Hofstede and Fenton 2005). Larger groups also promote warmer microclimates that favor some ectoparasites (Postawa and Szubert-Kruszyńska 2014). Within groups, individuals may not be equally vulnerable to parasites: more exploratory group members may be more susceptible (Webber et al. 2015). In conclusion, roost switching may serve to reduce ectoparasite loads. However, the likelihood of switching roosts in response to ectoparasites within and across bat species should be expected to vary according to the types of ectoparasites and roosts being considered, geographic location and time of year, and group size and reproductive condition of the bat species under consideration.
1.4 Roost Suitability
It is often suggested that bats may switch roosts to locate alternative roosts for future inhabitation (e.g., Kerth and Reckardt 2003; Carter and Feldhamer 2005; Russo et al. 2005). Such scouting could be a very costly endeavor. For instance, because roosts are susceptible to decay or destruction, bats may have to move among several potential roosts to locate a subset of relatively safe, alternative roosts. Indeed, Bechstein’s bats living in more dynamic groups discover and occupy newly available roosts more quickly compared to brown long-eared bats living in more cohesive groups (Fleischmann and Kerth 2014), suggesting roost switching, and thus fission–fusion dynamics, may help bats locate new roosts. However, it has been suggested that a bat must spend at least 1 day in a particular roost to assess its suitability (Kerth et al. 2001a, b). As Kerth et al. (2001a, b) suggest, individuals face the risk of spending time in suboptimal conditions that could lead to increased energetic expenditure and reduced offspring survival. It therefore seems unlikely bats switch roosts simply to gather information about alternative roosts for potential future use.
Instead, bats may leave a roost as it becomes unsuitable, either because the structure is no longer available or conditions in the roost have changed for the worse. Certainly roost permanence and microclimate influence roost use by bats. Caves and buildings, on the one hand, are relatively permanent roosts that offer relatively stable microclimates, together with several alternative microclimates in the same roost thus reducing the need to switch roosts (Lausen and Barclay 2006). Crevices in rocks and trees, on the other hand, are subject to decay (Kunz and Lumsden 2003; Barclay and Kurta 2007) and, although microclimate inside these structures is more stable compared to external conditions, microclimate varies to some degree with changes in ambient conditions as well as roost structure (Sedgeley 2001; Clement and Castleberry 2013). Meanwhile foliage is typically ephemeral and the microclimate is strongly influenced by ambient conditions (Kunz and Lumsden 2003). As discussed above, bats switch less frequently among more permanent roosts, such as caves and buildings, compared to bats roosting in trees and foliage (Kunz and Lumsden 2003; Barclay and Kurta 2007). At the same time, however, it is generally thought more permanent roosts are typically more limited on the landscape compared to more ephemeral roosts, which in turn may limit opportunities for roost switching (Lewis 1995). Interestingly, the opposite is true of some tent-making bats; depending on the plant species and mode of construction, some tents can last 6 weeks while others last only days yet bats roosting in more stable tents generally live in more dynamic groups (Sagot and Stevens 2012). That said, within species using similar types of roosts, roost switching is in fact more frequent when roosts are more abundant compared to when roosts are limited in a managed landscape (e.g., Chaverri et al. 2007). Thus, at least for some species, roost permanence and availability influence roost switching.
Examples from vespertilionid bats, where roost switching is particularly well studied, suggest that they switch roosts in response to changes in roost suitability. Soprano pipistrelles move among alternative locations within attics and roofs when temperatures exceed 40 °C (Lourenço and Palmeirim 2004). Similarly, big brown bats switch among buildings primarily on hot days (Ellison et al. 2007). Northern long-eared bats switch among tree cavities almost daily in response to changes in temperature, wind, precipitation, and barometric pressure, but only if they are in trees that are presumably less buffered against fluctuations in ambient conditions (Patriquin 2012). By contrast, pallid bats do not appear to switch roosts in response to daily changes in temperature (Lewis 1996). Meanwhile, the number of bats in a roost affects roost microclimate, arguably to a greater degree than ambient conditions (e.g., Willis and Brigham 2007; Pretzlaff et al. 2010; but see Bartonička and Řehák 2007) yet individual benefits gained through group warming diminish as group size increases (Boyles et al. 2008). Group size will therefore fluctuate as individuals join or leave in an effort to seek an optimal group size (Krebs and Davies 1995). Roost suitability is therefore a dynamic quality influenced by synergistic effects of roost structure, ambient conditions, and the number of bats, which together may explain why bats switch roosts.
2 Why Do not Group Members Move Together?
Fission–fusion dynamics in bats are not only the result of roost switching, but more importantly the result of group members switching to different roosts at different times. It therefore seems unlikely that efforts to reduce predation or parasitism can explain why some individuals remain in a roost after part of the group has left, or why a group splits up to move to two or more separate roosts (e.g., Barclay 1982; Lewis 1996; Carter and Feldhamer 2005; Patriquin et al. 2010). Presumably all group members would benefit equally from reduced predation risk and parasitism and would gain these benefits if they moved together to the same roost (Barclay and Kurta 2007). However, group members may not move together if they have different information about roost suitability, including the risk of predation and parasitism. Furthermore, and as noted above, individual costs (e.g., predation and parasitism) and benefits (e.g., group warming and cooperation) of group living also vary dynamically with group size, along with individual differences in sex, age, and reproductive condition (Krause and Ruxton 2002; Terrien et al. 2011). Group members may experience different perceived costs and benefits of remaining in a group, or in moving to one particular roost, and therefore may not move together.
It may be necessary or beneficial for groups to temporarily split into smaller groups (Kerth 2010). For example, space inside roosts may be limited and may force a colony to split into multiple groups, as supported by observations of increasing group size with increasing tree diameter and cavity size (e.g., Lacki and Schwierjohann 2001; Willis et al. 2006; Olson and Barclay 2013). However, this pattern is not consistent across all species, as no relationship between group size and tree diameter was found for northern long-eared bats (Vonhof and Wilkinson 1999). That said, tree diameter may not be a reliable proxy for cavity size in some tree species and therefore may not predict group size (Willis et al. 2006). More than this, based on direct measures of cavity size, Willis and Brigham (2004) argue that the cavities they studied could have supported larger groups of female big brown bats than observed. Thus, it appears that limited space in roosts does not entirely explain why group size changes.
Instead, fission–fusion dynamics likely reflect individuals’ attempts to balance the costs and benefits of group living by adjusting group size (Olson and Barclay 2013). For example, the benefits of group warming diminish in larger groups while costs, such as competition, parasitism, and overheating, increase with group size (Olson and Barclay 2013 and references within). Group size is therefore expected to change as the relative costs and benefits of joining a group change, as supported by a direct relationship between group size, roost temperature, and ambient conditions for female temperate bats like Bechstein’s bats and big brown bats (Kerth and König 1999; Willis and Brigham 2007; Pretzlaff et al. 2010). This, however, fails to explain why group composition varies almost daily as the same subset of group members could presumably remain together with each fission event, suggesting additional factors influence group composition.
Theoretical evidence suggests group fission and corresponding changes in group composition occur when group members do not reach a consensus on when and where to move (Aureli et al. 2008; Sueur et al. 2011; Kao et al. 2014). As Sueur et al. (2011) point out, individual differences in preferences for when to move could theoretically be resolved by effectively taking the average preference across group members. It remains unclear when the decision is made to switch roosts: dawn swarms (Naďo and Kaňuch 2013) suggest the decision may be made in the morning, while other evidence suggests bats visit roosts throughout the night (Fleischmann and Kerth 2014). Regardless, taking an average decision on whether to move today or several days later could be very costly for bats (see above). Furthermore, it may be difficult to resolve conflict over where to move because spatial location attributes, such as patches of food, water, or shelter, cannot simply be averaged (Sueur et al. 2011). However, while roosts are discrete in space and therefore cannot be averaged, groups could theoretically choose a roost with conditions that best meet the average needs of all group members. Selecting an average roost may, however, result in costly, suboptimal conditions for some group members. Failure to reach group consensus may therefore occur when individual needs conflict with the needs of other group members.
Failure to reach a consensus on when and where to move may also occur when group members have different information about roost suitability. For example, female Bechstein’s bats collectively choose to abandon a roost when the majority of group members (more than 50 %) experience a simulated disturbance at the roost (e.g., blocked entrance or a puff of air; Kerth et al. 2006; Fleischmann et al. 2013; Fleischmann and Kerth 2014). However, initial consensus aside, some of these females then chose to return to disturbed roosts (Kerth et al. 2006; Fleischmann et al. 2013; Fleischmann and Kerth 2014). Moreover, Bechstein’s bats do not reach group consensus about abandoning a roost when only a minority (less than 50 %) of the group experiences a simulated disturbance at the roost (Kerth et al. 2006; Fleischmann et al. 2013; Fleischmann and Kerth 2014). Interestingly, female brown long-eared bats (Plecotus auritus) generally switch roosts less often and always reached a consensus to avoid a roost even when only a minority of group members were presented with an aversive stimulus (an air puff), and in turn live in more cohesive groups (Fleischmann and Kerth 2014). It therefore appears that fission–fusion dynamics occur (i) when individual information conflicts with social information about roost suitability (Kerth 2010; Fleischmann et al. 2013; Fleischmann and Kerth 2014) and (ii) when different individuals have different requirements, due to, for example, differences in age, sex, parasite load, and reproductive condition.
2.1 Social
Moving freely among groups and group members should allow bats to maintain social connections across a larger network of group members, thereby increasing their opportunities to cooperate, or share information, with more individuals (O’Donnell and Sedgeley 1999; Willis and Brigham 2004; Kerth and Van Schaik 2011). Theoretical predictions suggest living within diverse groups facilitates learning (Sueur et al. 2011). Network analyses for giant noctule bats (Nyctalus lasiopterus), northern long-eared bats, and Indiana bats have revealed that even group members that roost together only intermittently maintain social connections that may facilitate information flow (Fortuna et al. 2009; Patriquin et al. 2010; Silvis et al. 2014a). Both white-striped free-tailed bats and false vampires appear to increase their networks by roosting with different conspecifics at day roosts than they do at night roosts, as groups of conspecifics found in separate day roosts can be found together at night roosts (Rhodes 2007; Schmidt 2013).
3 Do Bats Form Social Bonds Despite Frequent Roost Switching?
To establish whether animals, including bats, with labile group dynamics live in passive aggregations or fission–fusion societies, we must establish whether group members form social bonds. The important distinction is that benefits derived in passive aggregations, such as risk dilution, increased vigilance, reduced energetic costs of movement, or enhanced thermoregulation, depend primarily on the number of group members and interaction among group members is more or less random or restricted to nearest neighbors (reviewed in Kao et al. 2014). By contrast, individuals living in social groups gain additional fitness benefits through cooperation and nepotism by interacting repeatedly with known individuals who may also be related (Kerth 2010). Thus, establishing whether bats form social bonds despite frequent roost switching allows us to differentiate between passive and social groups.
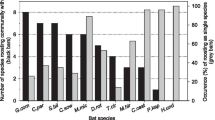
Here we define social bonds (i.e., affiliative social relationships) as nonrandom associations. Through labor-intensive radio tracking and the adoption of microchip technology, the amount of time individuals spend together in day roosts can be measured and compared to a random distribution of associations to determine if associations are truly social or a byproduct of mutual roost preferences (reviewed in Johnson et al. 2013). From this work, it is now evident that group members form preferred associations that cannot be explained by mutual roost preferences, with some group members forming stronger, more lasting relationships than others (Table 4.1). In fact, increasing evidence suggests that, much like other taxa with fission–fusion dynamics, several bat species appear to live in closed societies. Members of sympatric colonies of female Bechstein’s bats, big brown bats, and northern long-eared bats, for example, are rarely found roosting together (Kerth et al. 2000; Willis and Brigham 2004; Patriquin et al. 2010). Indeed, female Bechstein’s bats may even actively reject foreign conspecifics from roosts (Kerth et al. 2002a, b). Bats therefore appear to form social bonds despite frequent movements among roosts and roost groups.
Not only do some bats form nonrandom associations, these relationships can last for years. For example, Spix’s disk-winged bats, a tropical species that roosts in unfurling leaves, maintain preferred associations for at least 100 days, and up to 4 years (Vonhof et al. 2004). Several vespertilionid species in temperate regions all maintain preferred associations across multiple years even though they move to hibernacula for the winter (Patriquin et al. 2010; Kerth et al. 2011; August et al. 2014). Thus, bats form social bonds that can last years, suggesting the potential for cooperation among group members.
The persistence of these bonds has likely played a role in shaping the complex structure of colonies with fission–fusion dynamics. Evidence from Bechstein’s bats, Spix’s disk-winged bats, big brown bats, and northern long-eared bats, for example, suggest sympatric groups form socially distinct colonies, where conspecifics in one colony are rarely found roosting in adjacent colonies (Kerth et al. 2000; Vonhof et al. 2004; Willis and Brigham 2004; Patriquin et al. 2010). Colonies are therefore made up of a network of groups interconnected by the movement of individuals that in turn form social bonds. Cluster analyses and network analyses suggest subgroups of individuals collectively spend more time together over the summer compared to other group members, resulting in communities within a colony (Patriquin et al. 2010; Kerth et al. 2011). Thus bats with fission–fusion dynamics appear to live in colonies made up of multiple, interconnected subgroups or communities, within which group members form social bonds with each other.
4 What Shapes Social Bonds?
Understanding who is more likely to roost together may offer clues as to why we see labile groups, as well as who is more likely to cooperate within these groups. Based on kin selection theory, we might expect close relatives to form stronger social bonds, which may then facilitate nepotism (Hamilton 1964). Indeed various species with fission–fusion dynamics appear to live in matrilineal colonies (Burland et al. 1999; Kerth et al. 2000; Castella et al. 2001; Metheny et al. 2008a; Flanders et al. 2009; Kerth and Van Schaik 2011; Patriquin et al. 2013). In addition, it appears that new colonies of Bechtein’s bats and big brown bats are formed when matrilineal kin leave an existing colony to establish a new colony (Kerth 2008b; Metheny et al. 2008b), suggesting a preference to roost with relatives. Within colonies, at least some female Bechstein’s bats and northern long-eared bats with the strongest social bonds are also more closely related (Kerth et al. 2011; Patriquin et al. 2013). However, relatedness does not appear to explain social bonds in all colonies of Bechstein’s bats, nor in colonies of big brown bats (Kerth and König 1999; Metheny et al. 2008a; Kerth et al. 2011). Instead, the relative importance of relatedness in shaping social bonds may depend on group size or the relative stability of groups; bats in smaller or more stable groups may be able to rely on familiarity, but in larger or more variable groups may have to use additional cues, such as relatedness (Patriquin et al. 2013). Even in systems where relatedness does predict social bonds, females regularly roost with unrelated group members (Kerth et al. 2011; Patriquin et al. 2013). Relatedness therefore does not entirely explain social bonds in these dynamic systems.
We might also expect group members with similar demographics (e.g., reproductive condition and age), and therefore similar needs, to form social bonds. On average, female big brown bats and northern long-eared bats form stronger social bonds at different points in the summer corresponding to reproductive periods (e.g., gestation or lactation), suggesting bonds may be based in part on similar needs during these periods (Willis and Brigham 2004; Garroway and Broders 2007; Patriquin et al. 2010). However, females may not reproduce every summer and the synchrony of parturition varies across species and region, therefore some groups may be comprised of females in different reproductive states (Altringham 1996). Within these mixed groups, Bechstein’s bats in the same reproductive condition form stronger relationships (Kerth and König 1999), also suggesting bonds may be based on similar reproductive needs. However, reproductive and nonreproductive female Bechstein’s bats are regularly found roosting together (Kerth and König 1999; Kerth et al. 2011) and pairs of nonreproductive and pregnant big brown bats spent more time together than pairs of pregnant bats, suggesting social bonds are not based entirely on similar needs driven by reproduction (Willis and Brigham 2004). It also appears that social bonds are not based on preferences to associate with cohort members. For instance, Myotis spp. groups consist of a range of ages (Patriquin et al. 2010; Kerth et al. 2011) and no significant correlation was found between age and association for female Bechstein’s bats (Kerth et al. 2011) while older female northern long-eared bats spend more time with younger adult group members (Patriquin et al. 2010). Shared demography thus does not entirely explain social bonds among bats in fission–fusion systems.
It therefore appears that groups may form initially due to similar needs, but within these groups bats may form social bonds based on something more than similarity (Sueur et al. 2011). Like primates, it is possible that social relationships among bats living in fission–fusion systems are shaped by a combination of partner fidelity, or time spent together, and partner choice based on long-term “bookkeeping” of social interactions that may predict the likelihood of receiving cooperative behaviors (Schino and Aureli 2009; Campenni et al. 2015). Longer term studies, such as the work on Bechstein’s bats, are therefore necessary to better examine what shapes long-term relationships among bats.
5 How Are Social Bonds Maintained?
As in other taxa, long-term social relationships are no doubt facilitated through spatial cues together with mechanisms for individual and kin recognition (Krebs and Davies 1995). For instance, Bechstein’s bats, northern long-eared bats, and Indiana bats show interannual fidelity to summer roosting areas and specific roost trees which could allow summer colonies to reestablish social relationships when they come out from hibernation (Kerth and König 1999; Patriquin et al. 2010; Silvis et al. 2014a). Interannual fidelity to roost sites and particular roosts have been documented in various other species and therefore the long-term stability of social relationships in these species warrants further investigation (Barclay and Brigham 2001; Entwistle et al. 2000; O’Donnell 2000; Veilleux and Veilleux 2004; Willis and Brigham 2004; Arnold 2007). While site and roost fidelity may provide a means to locate and recognize group members, it may not allow individuals to reliably differentiate among group members, which would presumably be beneficial in affiliative relationships.
Given most bats rely heavily on echolocation to gain information about their environment; it is not surprising that vocalizations also play an important social role (reviewed in Kanwal et al. 2013 and Schmidt 2013). At least some bats can readily differentiate between conspecific and heterospecific calls at roosts (e.g., Schöner et al. 2010; Furmankiewicz et al. 2011). More importantly, however, several species produce group-specific echolocation and social calls that may allow individuals to differentiate between familiar and unfamiliar conspecifics, as well as individually distinct calls that may allow individuals to differentiate between familiar and unfamiliar group members (Pfalzer and Kusch 2003; Carter et al. 2008; Kazial et al. 2008; Melendez and Feng 2010; Schöner et al. 2010; Voigt-Heucke et al. 2010; reviewed in Carter and Wilkinson 2013a, b and Ross and Holdereid 2013). Evidence from common pipistrelles suggests different matrilines produce different calls that are not likely coded for by mtDNA and are therefore most likely learned from group members (Fornůsková et al. 2014). Vocalizations therefore appear to play an important role in group and individual recognition.
Behavioral evidence also supports predictions that bats use calls to differentiate between familiar and unfamiliar conspecifics. For instance, Spix’s disk-winged bats use social calls to maintain contact with group members while foraging (pers. obs. as cited in Chaverri et al. 2012), which may be akin to “grooming at a distance” in primates where group members that regularly groom one another maintain vocal contact when they are apart while foraging (reviewed in Dunbar and Shultz 2010). Maintaining contact while foraging may help bats ensure they later roost with preferred group members (Chaverri et al. 2012). In experimental studies, white-winged vampire bats (Diaemus youngi) engage in antiphonal calling when separated from group members and are capable of discriminating between familiar and unfamiliar conspecifics (Carter et al. 2008, 2009). Similarly, pallid bats are more likely to respond to playback calls of familiar group members at roosts (Arnold and Wilkinson 2011). Spix’s disk-winged bats in roosts produce “contact” calls in response to “inquiry” calls produced by group members flying in the area who then preferentially enter roosts of familiar group members (Chaverri and Gillam 2010; Chaverri et al. 2010, 2012). Thomas’ fruit-eating bats (Dermanura watsoni) and Honduran white bats (Ectophylla alba) also produce social calls at roosts (Gillam et al. 2013), and giant noctule bats and Daubenton’s bats (Myotis daubentonii) are attracted to playbacks of conspecific calls at roosts (Ruczyński et al. 2007, 2009), but whether they are used in group recognition remains to be tested.
Visual and olfactory cues may also allow group members to locate one another. For instance, swarming outside summer roosts documented for several species may serve to visually signal the location of group members (Russo et al. 2005; Kerth et al. 2006; Rhodes 2007; Lučan and Radil 2010; Naďo and Kaňuch 2013). Within roosts, olfaction may play an important role in group and individual recognition (Wilkinson 1986; Brooke 1997; Safi and Kerth 2003; Carter and Wilkinson 2013a, b). Gland secretions differ between Bechstein’s bat colonies, as well as among individuals within colonies (Safi and Kerth 2003). This, together with evidence that bats rub noses, particularly when they return to a roost, suggests odor cues may facilitate group and individual recognition in this species (Kerth et al. 2003; Safi and Kerth 2003). Additionally, experimental results suggest a range of species living in fission–fusion systems, including common pipistrelles, big brown, and Brazilian free-tailed bats (Tadarida brasiliensis), are capable of using olfactory cues to differentiate between familiar and unfamiliar conspecifics (De Fanis 1995; Bloss et al. 2002; Englert and Greene 2009), as well as to differentiate between different familiar group members (De Fanis 1995). Ancillotto and Russo (2014) suggest European free-tailed bats (Tadarida teniotis) use olfactory cues to differentiate between familiar and unfamiliar conspecifics, as evidenced by aggressive behavior toward unfamiliar conspecifics. Thus it appears bats may use a combination of spatial, vocal, visual, and olfactory cues to locate group members and maintain social bonds.
6 Are There Examples of Cooperation Among Bats Living in Fission–Fusion Systems?
As demonstrated in a recent review (Carter and Wilkinson 2013a, b), there is mounting evidence that bats benefit from cooperative behavior, such as social warming, social grooming, and social transmission of information about roosts and food. It is widely assumed that group warming is the primary benefit of group living in bats (Kerth 2008a), which is presumably dependent more on group size than group composition. However, nonreproductive Bechstein’s bats and big brown bats can be found roosting with reproductive conspecifics even though they are under different thermoregulatory constraints and should therefore select different roosts, suggesting they are in fact cooperating to contribute to group warming (Kerth and König 1999; Willis and Brigham 2004). Though studied more extensively in species that live in relatively stable social groups, such as common vampire bats and Kuhl’s pipistrelles (Pipistrellus kuhlii), social grooming (allogrooming) has also been documented in Bechstein’s bats that live in fission–fusion systems (Kerth et al. 2003; Ancillotto et al. 2012; Carter 2015; reviewed in Carter and Wilkinson 2013a, b). Social grooming in these dynamic systems may help build and maintain social bonds as it does in a range of other taxa (reviewed in Dunbar and Schultz 2010 and Russell and Phelps 2013).
Bearing in mind that bats can live up to 30 years or more, and females show strong natal philopatry to summer breeding areas and to roosts, it is possible they could build long-term knowledge about suitable roosts and share this information with younger, less experienced group members. Experimental evidence suggests bats are indeed capable of remembering spatial cues (reviewed in Ross and Holderied 2013) and theoretical models suggest remembering the location of roosts reduces search effort (Ruczyński and Bartoń 2012). Field observations of bats swarming outside roosts before entering for the day (Russo et al. 2005; Kerth et al. 2006; Rhodes 2007; Lučan and Radil 2010; Naďo and Kaňuch 2013) together with evidence of antiphonal calling by Spix’s disk-winged bats at roosts (Chaverri et al. 2010, 2012) and experimental studies demonstrating that experienced female Bechstein’s bats return to roosts with naïve group members prior to choosing a roost for the day (Kerth et al. 2006; Fleischmann et al. 2013; Fleischmann and Kerth 2014), suggest bats have the opportunity to learn about suitable roosts from group members.
Bats living in fission–fusion systems may also learn about foraging opportunities from group members. Several studies have demonstrated that bats from a range of social systems, including little brown bats, big brown bats, and Brazilian free-tailed bats that live in fission–fusion systems, can learn about food availability, and how to capture prey, through auditory, visual, and olfactory cues (Gaudet and Fenton 1984; Ratcliffe and ter Hofstede 2005; Page and Ryan 2006; Gillam 2007; Dechmann et al. 2009, 2010; Wright et al. 2011; Jones et al. 2014; O’Mara et al. 2014). However, it is possible that only species feeding on patchily distributed food, such as fruit or insect swarms, are likely to benefit from information sharing compared to gleaning species that feed on single prey, rather than patches, that are typically distributed less predictably.
Alloparental care, such as nursing and guarding nondescendant pups, has also been documented in bats. However, examples of these behaviors are restricted to species that live in year-round stable groups, including evening bats (Nycteris humeralis) (Wilkinson 1992; reviewed in Carter and Wilkinson 2013a, b) and greater spear-nosed bats (Phyllostomus hastatus) (Bohn et al. 2009; reviewed in Carter and Wilkinson 2013a, b). It therefore appears strong, predictable bonds may be needed to offset the costs of investing energy in nondescendant young.
7 What Does All of This Tell Us About Evolution of Fission–Fusion Dynamics?
Consistent with evidence from primates, cetaceans, and elephants, work with bats supports theoretical predictions that, like other social systems, fission–fusion dynamics are influenced by life history, reproductive strategy, phylogeny, and resource stability (Clutton-Brock and Janson 2012). For instance, fission–fusion has evolved primarily in long-lived animals with high offspring investment and high female philopatry occupying environments that vary within an individual animal’s lifetime (Aureli et al. 2008; Couzin and Laidre 2009). Together, these characteristics favor group living, cooperative behavior, and social learning to better adapt to changes in the environment (Whitehead 2007; Aureli et al. 2008). Moderate spatial and temporal variability then leads to changes in the relative costs and benefits of group living, resulting in fission–fusion dynamics with some level of consensus among group members (Sueur et al. 2011). This is consistent with the observation that fission–fusion dynamics predominate in bats living in temperate regions where there are both seasonal and daily fluctuations in the environment. By contrast, year-round stable groups predominate in the tropics where conditions are less variable. Of course, as highlighted above, there are exceptions: several tropical species, including Spix’s disk-winged bats and some tent-roosting bats, do live in fission–fusion systems. These species, however, tend to rely on ephemeral roosts which may necessitate roost switching; at the same time, despite frequent roost switching, tent-making species studied to date form groups that are relatively more stable compared to temperate bats. Predictions about resource stability and its influence on group structure therefore require further testing.
Consistent with theoretical predictions and empirical evidence from elephants and primates (Aureli et al. 2008), it appears that fission–fusion systems in bats favor individual recognition over group signatures. For example, species living in fission–fusion systems, such as Spix’s disk-winged bats, produce individually specific calls rather than group-specific calls (Gillam and Chaverri 2012). By contrast, species living in stable social groups, such as greater spear-nosed bats and lesser bulldog bats (Noctilio albiventris), rely on group-specific calls, with some also using individually distinct calls (Boughman 1997; Boughman and Wilkinson 1998; Voigt-Heucke et al. 2010). Moreover, scent profiles differ between colonies of Bechstein’s bats, but also across individuals within colonies (Safi and Kerth 2003). Thus, it appears individual recognition may be more important in fission–fusion systems where group members interact intermittently compared to stable groups where group members interact predictably.
8 Conclusions and Future Directions
Fission–fusion societies in bats appear to consist of colonies made up of multiple interconnected subgroups or communities consisting of multiple matrilines, within which group members form social bonds with each other based in part on relatedness and similarities in reproductive condition. The fundamental driver of these fission–fusion dynamics is frequent roost switching, likely due to a combination of predation risk, commuting costs, ectoparasitism, and changes in roost availability. Group composition changes when group members switch to different roosts at different times, likely due to benefits of temporarily splitting into smaller groups together with lack in group consensus on when and where to move. Despite the fluid nature of these groups, bats maintain long-term social bonds that may then favor the evolution of cooperation among group members. However, more work is needed to better understand both the proximate and ultimate causes of fission–fusion dynamics in bats, which we highlight below with suggestions for future work.
The causes of roost switching require closer examination. Most studies have approached the question of roost switching as a univariate problem, when in reality factors such as predation risk, commuting costs, and ectoparasites are likely not mutually exclusive and their relative influence will depend on the synergistic effects of roost type, environment, and demography. Ellison et al. (2007) suggested the strong effect of temperature on roost switching by big brown bats may have masked the effect of ectoparasites. Moreover, the influence of each of these factors will no doubt differ for bats roosting in foliage compared to those roosting in caves or buildings, due to differential exposure to predators, parasites, and the elements. For example, pallid bats using roosts with different characteristics show different roost-switching behavior and correspondingly different ectoparasite loads (reviewed in Lewis 1995). Similarly, the degree of roost switching varies with reproductive condition, due in part to physical constraints on movement when pregnant and transporting nonvolant young, but also due simply to differences in thermoregulatory demand. Thus, the costs of moving may simply outweigh the benefits of switching roosts during certain periods. Future studies should therefore expand on the work of Ellison et al. (2007) by quantifying all possible factors, including predator densities, ectoparasite loads, distances between roosts and feeding sites, changes in ambient conditions, and corresponding changes in roost quality. Furthermore, to better examine proximate factors shaping fission–fusion dynamics, researchers should use multivariate analyses to examine the relative effect of each factor while controlling for reproductive condition.
The influence of predation risk and ectoparasites on roost switching requires more careful examination to better assess their role in the evolution of fission–fusion dynamics. In particular, controlled experiments that manipulate predation risk and field studies that provide estimates of predator densities could help clarify the role of predation risk on roost switching (see Lima and O’Keefe 2013 for a detailed discussion of the effect of predators on bat behavior, and how to test these effects). Concurrently, efforts should be made to quantify all ectoparasites because strategies to reduce ectoparasites differ for mites and bat flies (ter Hofstede and Fenton 2005; Postawa and Furman 2014; Postawa et al. 2014). Further, experimental studies similar to that of Reckardt and Kerth (2007), which manipulated ectoparasite loads in roosts, would also help quantify the effect of parasites on roost-switching behavior. As with any rigorous study, a sufficient number of group members must be examined to account for individual variation, which may be particularly important in light of recent evidence that susceptibility to parasites varies with age, sex, and exploratory behavior (Webber et al. 2015).
The influence of ambient conditions on roost-switching behavior also requires more attention. For example, more studies from across a range of species and conspecifics occupying roosts that differ in vulnerability to changes in ambient conditions (e.g., buildings, trees, and foliage) would help determine whether environmental variability does in fact shape fission–fusion dynamics. Also, experimental manipulation of temperatures in bat boxes could provide valuable insight to the influence of microclimate on roost switching. To date, manipulations of bat boxes have demonstrated that bats prefer warmer roosts during lactation (Kerth et al. 2001a, b). This work could be expanded using a thermostat to regulate a constant optimal temperature inside bat boxes where bats would presumably remain for the entire season if changes in roost microclimate are the only predictors of roost switching. Also, creating a set of roosts where each have different temperatures could determine whether group consensus could be achieved if an “average” roost were available or whether it would still be too costly for some group members to move to a suboptimal roost.
Change in group size and composition after switching roosts is, of course, also fundamental to the evolution of fission–fusion dynamics. It appears that group size and composition change when it is beneficial to temporarily split into multiple smaller groups to offset the costs of group living and when group members cannot come to a group consensus on where to move (Kerth 2010). These conclusions, however, have been drawn based largely on work with a single species, Bechstein’s bats, living in bat boxes. Recent evidence of strong interspecific differences in group cohesion (Fleischmann and Kerth 2014) highlights the need for more extensive studies examining group consensus across a range of species and conspecific groups in different environments and that use different types of roosts. Fleischmann and Kerth (2014) suggest future work should also consider social and genetic relationships and how they affect the likelihood of reaching a group decisions as individuals with stronger social or genetic ties may be more likely to reach consensus. Behavioral syndromes may also influence decision-making (Fleischmann and Kerth 2014). Kilgour and Brigham (2013) experimentally demonstrated that some group members are more likely than others to explore and discover new food resources. Perhaps more exploratory individuals are more likely to leave group members to locate new roosts while others are more likely to stay behind, as suggested for Bechstein’s bats (Kerth and Reckardt 2003) and giant noctule bats (Fortuna et al. 2009). As future work clarifies the causes of fission–fusion dynamics, we can then better test hypotheses that fission–fusion dynamics allow group members to balance individual needs with the costs and benefits of group living (Kerth 2010).
Despite the fluid nature of these groups, individuals form both short- and long-term bonds. These bonds appear to be based in part on relatedness and shared needs shaped by similarities in reproductive condition. However, evidence that unrelated individuals and females in different reproductive condition form bonds suggests bonds are shaped by more than shared needs. Future work could expand on a recent experiment that provided big brown bats a binary choice between pairs of known conspecifics (Kilgour et al. 2013). Interestingly, a recent study suggests that early association shapes social bonds as juvenile Kuhl’s pipistrelles that roost close together form long-term relationships in adulthood (Ancillotto et al. 2012), which may then also explain why some group members remain together despite fission–fusion events. As discussed by Aureli et al. (2008), communication is also a key to group dynamics. Thus, more explicit tests are needed to examine how group members find each other. Relocation experiments similar to an experiment performed by Willis and Brigham (2004), for example, could prove valuable.
In their review, Dunbar and Schultz (2010) point out that within a social system, there are likely different levels of organization that may be akin to hierarchies. Indeed, within colonies of Bechstein’s bats and northern long-eared bats, distinct communities or subgroups exist where females are more likely to interact with one another compared to other group members (Patriquin et al. 2010; Kerth et al. 2011). Dunbar and Schultz (2010) therefore suggest that the level of social complexity can be compared across species by quantifying the size of the “base unit” (i.e., a summer colony of bats) and “the number of layers of acquaintanceship that emerge from it” (i.e., subgroups or communities in bats) (p. 795). Living in closed societies can be both beneficial and costly, as discussed in more detail by Kerth and Van Schaik (2011). For instance, living in closed societies consisting primarily of maternal kin may favor cooperation and nepotism and it appears to limit exposure to novel pathogens (Kerth and Van Schaik 2011). At the same time, however, these authors (Kerth and Van Schaik 2011) suggest bats in closed societies may not be able to disperse to new roosting areas following a disturbance, particularly given foreign individuals may be actively excluded from roosts (Kerth et al. 2002a, b). However, the case where socially distinct groups of Bechstein’s bats fused to form a single group following a population crash further refutes this hypothesis (Baigger et al. 2013).
Fission–fusion dynamics vary along a continuum, within and across species and over time (Aureli et al. 2008). Brown long-eared bats switch roosts less frequently and therefore live in more cohesive groups compared to Bechstein’s bats (Fleischmann and Kerth 2014). When big brown bat and Bechstein’s bat colonies become too large, they split into multiple new colonies, where matrilineal kin leave a colony to establish a new colony elsewhere (Kerth 2008b; Metheny et al. 2008b). By contrast, when Bechstein’s bat colony size decreased due to a sudden population crash, otherwise socially distinct communities merged (Baigger et al. 2013). Initially group members from each community spent more time with each other but, over time, these former preferred relationships broke down and new bonds formed (Baigger et al. 2013). We hypothesize that group mergers, together with a breakdown of preexisting relationships over time to form new social bonds, may then explain why colonies consist of multiple matrilines and why social bonds within these colonies are not necessarily predicted by relatedness (Kerth and König 1999; Metheny et al. 2008b; Kerth et al. 2011; Patriquin et al. 2013). We also propose that the degree of structuring (i.e., number of communities or subgroups) within a colony, as well as the factors shaping social relationships within colonies, such as relatedness, may differ depending on whether a colony has undergone a large-scale fission into multiple colonies or whether it has formed as a result of the fusion of multiple communities. The structure and relationships within colonies will also depend on the amount of time that has passed since the fission or fusion event. We might therefore expect findings to vary across species and conspecific groups in future studies.
Though evidence is mounting that bats form social bonds in dynamic fission–fusion systems, it is generally assumed that time spent in the same roost translates to social interactions such as cooperation, which may not be the case (Dunbar and Schultz 2010). For instance, at least one study suggests bats may line cavity walls rather than form clusters in roosts (unpublished data as cited in Willis and Brigham 2007), suggesting individuals within a roost do not necessarily interact equally. However, with greater affordability and advances in technology, we encourage future work to document interactions among bats (see Hristov et al. 2013 for a review and suggestions). In addition to suggestions offered by Hristov et al. (2013), we propose future work could mark individuals with a fluorescent powder and quantify the rate of transfer among group members to first establish whether bats roosting together do in fact interact and therefore potentially cooperate. At the same time, care must be taken when interpreting such data as strength of social relationships may not predict cooperation and reciprocity may not be tit for tat: for example, allogrooming may be exchanged for a different service, such as information about food (Carter and Wilkinson 2013a, b).
Network analyses are a valuable tool that can help reveal the complexities of fission–fusion dynamics, as well as the potential for cooperation and information flow among group members (Croft et al. 2008; Whitehead 2008). However, some caution must be exercised when using, and interpreting, these analyses (see Johnson et al. 2013 for suggestions). For instance, theoretical models have demonstrated that fission–fusion can emerge from random patterns as individuals aggregate around a common resource (Cross et al. 2005; Ramos-Fernández et al. 2006). Future work should therefore ensure observed association patterns are compared against random distributions to confirm they represent social preferences rather than shared interest in resources (Whitehead 2008). With proper care, future work could then test predictions about roles of individuals in networks by comparing the rate of spread of novel information among group members with different degrees of direct and indirect connections, or for individuals that may act as “brokers” between subgroups. It would also be interesting to look at reproductive success of females that are more central to networks compared to those at edges: evidence from baboons suggests females that spend more time with other group members, and more time allogrooming, have higher reproductive success (Silk 2007).
Future work should also examine sociality in male bats, which has received little attention. Comparative studies looking at roost switching and ectoparasites in male and female conspecifics, for example, would help tease apart the role of environment and ectoparasites in shaping roost switching and fission–fusion dynamics. Adult male Bechstein’s bats typically switch roosts less often than females and have more bat flies, suggesting more frequent roost switching may in fact serve to reduce parasite load (Reckardt and Kerth 2006).
Relationships at night roosts and hibernacula have also largely been ignored. It remains to be seen whether other species follow patterns similar to false vampires and white-striped free-tailed bats where groups of conspecifics found in separate day roosts can be found together at night roosts (Rhodes 2007; Schmidt 2013). Moreover, as Kerth and Van Schaik (2011) point out, it is not yet known whether groups found roosting together in the summer are also found hibernating together. Hibernating with summer groups could presumably allow bats to maintain the long-term social bonds that have been documented in some species. Bats are regularly aroused through winter in response to evaporative water loss during hibernation (Ben-Hamo et al. 2013 and references within), offering them the opportunity to interact with each other. If, summer groups do also hibernate together this could have devastating consequences in light of the emergence of white-nose syndrome. The rapid spread of this lethal fungus (Pseudogymnoascus destructans) throughout hibernacula in eastern North America has resulted in the loss of nearly six million bats since its first detection in 2006 (BCI 2014). If summer social groups also hibernate together and are subsequently lost to white-nose syndrome, we could see the loss of entire matrilines, resulting in reduced genetic variance. Moreover, we could see the loss of entire colonies in a particular area as populations of big brown bats, eastern small-footed myotis (Myotis leibii), little brown bats, northern long-eared bats, Indiana bats, and tricolored bats (Perimyotis tricolor) are all declining as a result of white-nose syndrome (Langwig et al. 2012). This then raises the question whether summer colonies of each of these species will also merge as observed in Bechstein’s bats following a population crash.
Though our understanding of fission–fusion dynamics in bats has grown over the past 15 years, much remains to be learned. Most of what we know is based on only a few well-studied species, and Bechstein’s bats in particular, and therefore conclusions are somewhat speculative at this time. As suggested for future directions for work with primates, we should be testing predictions about selection on group structure by performing comparative studies across species that consider ecology and phylogeny, and compare this to work in other taxa (Clutton-Brock and Janson 2012). This highlights the need for more extensive studies across species, as well as across conspecifics occupying different environments and sympatric groups of conspecifics using roosts that vary in stability. Comparative studies of heterospecifics living in different social systems but in similar environments would also help to further understand the evolution of fission–fusion dynamics in bats. Also, as can be seen in Table 4.1, measures of association and definitions of social groups vary widely across studies. We therefore encourage future work to use consistent terminology and methodology to better allow comparisons across studies.
9 Conservation Implications
Bats living in fission–fusion systems need a multitude of roosts to support social relationships and thermoregulatory needs (O’Donnell 2000; Willis and Brigham 2004; Russo et al. 2005; Popa-Lisseanu et al. 2008; Fortuna et al. 2009; Patriquin et al. 2010). Previous conservation efforts have endeavored to quantify key roost characteristics to conserve particular types of trees. However, evidence of high reuse of particular trees in some areas together with observations that bats use a large set of roosts within and across years suggests they require a set of roosts with different characteristics (Barclay and Kurta 2007). Some of these roosts appear to act as “central” or “primary” roosts as they are used repeatedly throughout a summer and by more group members than other roosts (Barclay and Kurta 2007; Popa-Lisseanu et al. 2008; Silvis et al. 2014a; Patriquin pers. obs). Loss of these central roosts results in dramatically fragmented social networks for Indiana bats (Silvis et al. 2014a) and the loss of roosts in general could result in smaller home ranges together with smaller colony sizes, in turn increasing the risk of local extinction (Borkin and Parsons 2011, 2014).
Susceptibility to disturbance may also depend on the shape of the network. Simulations suggest Indiana bat networks are more sensitive to roost loss, as networks were fragmented into multiple, smaller groups following the removal of 10 % of roosts, whereas the removal of more than 20 % of northern long-eared bat roosts was needed before colonies became fragmented (Silvis et al. 2014a, b). Colonies were less likely to fragment when bats shared a higher number of roosts (Silvis et al. 2014a). A further test of this could compare the effect of roost loss on Bechstein’s bat and brown long-eared bat networks, which exhibit different levels of fission–fusion dynamics and correspondingly different abilities to discover and occupy new roosts (Fleischmann and Kerth 2014). In conclusion, how bats determine whether they should stay or should go is a multifaceted question, one that requires careful attention by researchers to a wide variety of biotic and abiotic factors, and one that likely differs across species, populations, and individuals and varies over time.
References
Altringham JD, Senior P (2006) Social systems and ecology of bats. In: Ruckstuhl KE, Neuhaus P (eds) Sexual segregation in vertebrates: ecology of the two sexes. Cambridge University Press, Cambridge, xii + 488 pp, pp 280–302
Altringham JD (1996) Bats: biology and behaviour. Oxford University Press, Inc., New York, ix + 262 pp
Ancillotto L, Russo D (2014) Selective aggressiveness in European free-tailed bats (Tadarida teniotis): influence of familiarity, age and sex. Naturwissenschaften 101:221–228
Ancillotto L, Serangeli MT, Russo D (2012) Spatial proximity between newborns influences the development of social relationships in bats. Ethology 118:331–340
Ancillotto L, Serangeli MT, Russo D (2013) Curiosity killed the bat: domestic cats as bat predators. Mamm Biol 78:369–373
Archie EA, Moss CJ, Alberts SC (2006) The ties that bind: genetic relatedness predicts the fission and fusion of social groups in wild African elephants. Proc R Soc B 273:513–522
Arnold BD (2007) Population structure and sex-biased dispersal in the forest dwelling vespertilionid bat, Myotis septentrionalis. Am Midl Nat 157:374–384
Arnold BD, Wilkinson GS (2011) Individual specific contact calls of pallid bats (Antrozous pallidus) attract conspecifics at roosting sites. Behav Ecol Sociobiol 65:1581–1593
August TA, Nunn MA, Fensome AG, Linton DM, Mathews F (2014) Sympatric woodland myotis bats form tight-knit social groups with exclusive roost home ranges. PLoS ONE 9:e112225
Aureli F, Schaffner CM, Boesch C, Bearder SK, Call J, Chapman CA, Connor R, Di Fiore A, Dunbar RIM, Henzi SP, Holekamp K, Korstjens AH, Layton R, Lee P, Lehmann J, Manson JH, Ramos-Fernandez G, Strier KB, Van Schaik CP (2008) Fission-fusion dynamics: new research frameworks. Curr Anthropol 49:627–654
Baigger A, Perony N, Reuter M, Leinert V, Melber M, Grünberger S, Fleischmann D, Kerth G (2013) Bechstein’s bats maintain individual social links despite a complete reorganisation of their colony structure. Naturwissenschaften 100:895–898
Barclay RMR (1982) Night roosting behavior of the little brown bat, Myotis lucifugus. J Mammal 63:464–474
Barclay RMR, Brigham RM (2001) Year-to-year reuse of tree-roosts by California bats (Myotis californicus) in southern British Columbia. Am Midl Nat 146:80–85
Barclay RMR, Kurta A (2007) Ecology and behavior of bats roosting in tree cavities and under bark. In: Lacki MJ, Hayes JP, Kurta A (eds) Bats in forests: conservation and management. Johns Hopkins University Press, Baltimore, Maryland, pp 17–59
Bartonička T (2010) Survival rate of bat bugs (Cimex pipistrelli, Heteroptera) under different microclimatic conditions. Parasitol Res 107:827–833
Bartonička T, Gaisler J (2007) Seasonal dynamics in the numbers of parasitic bugs (Heteroptera, Cimicidae): a possible cause of roost switching in bats (Chiroptera, Vespertilionidae). Parasitol Res 100:1323–1330
Bartonička T, Řehák Z (2007) Influence of the microclimate of bat boxes on their occupation by the soprano pipistrelle Pipistrellus pygmaeus: possible cause of roost switching. Acta Chiropterologica 9:517–526
Bartonička T, Růžičková L (2013) Recolonization of bat roost by bat bugs (Cimex pipistrelli): could parasite load be a cause of bat roost switching? Parasitol Res 112:1615–1622
Bartonička T, Bielik A, Řehák Z (2008) Roost switching and activity patterns in the soprano pipistrelle, Pipistrellus pygmaeus, during lactation. Ann Zool Fenn 45:503–512
BCI—Bat Conservation International (2014) White-nose syndrome resources. http://www.batcon.org/resources/for-specific-issues/white-nose-syndrome
Ben-Hamo M, Munoz-Garcia A, Williams JB, Korine C, Pinshow B (2013) Waking to drink: rates of evaporative water loss determine arousal frequency in hibernating bats. J Exp Biol 216:573–577
Bloss J, Acree TE, Bloss JM, Hood WR, Kunz TH (2002) Potential use of chemical cues for colony-mate recognition in the big brown bat, Eptesicus fuscus. J Chem Ecol 28:819–834
Bohn KM, Moss CF, Wilkinson GS (2009) Pup guarding by greater spear-nosed bats. Behav Ecol Sociobiol 63:1693–1703
Borkin KM, Parsons S (2011) Sex-specific roost selection by bats in clear-fell harvested plantation forest: improved knowledge advises management. Acta Chiropterologica 13:373–383
Borkin KM, Parsons S (2014) Effects of clear-fell harvest on bat home range. PLoS ONE 9:e86163
Boughman JW (1997) Greater spear-nosed bats give group-distinctive calls. Behav Ecol Sociobiol 40:61–70
Boughman JW, Wilkinson GS (1998) Greater spear-nosed bats discriminate group mates by vocalizations. Anim Behav 55:1717–1732
Bourke AFG (2014) Hamilton’s rule and the causes of social evolution. Philos Trans R Soc B 369:20130362
Boyles JG, Storm JJ, Brack V (2008) Thermal benefits of clustering during hibernation: a field test of competing hypotheses on Myotis sodalis. Funct Ecol 22:632–636
Brooke AP (1997) Social organization and foraging behaviour of the fishing bat, Noctilio leporinus (Chiroptera: Noctilionidae). Ethology 103:421–436
Burda H, Honeycutt RL, Begall S, Locker-Grutjen O, Scharff A (2000) Are naked and common mole-rats eusocial and if so, why? Behav Ecol Sociobiol 47:293–303
Burland TM, Barratt EM, Beaumont MA, Racey PA (1999) Population genetic structure and gene flow in a gleaning bat, Plecotus auritus. Proc R Soc B 266:975–980
Campenni M, Manciocco A, Vitale A, Schino G (2015) Exchanging grooming, but not tolerance and aggression in common marmosets (Callithrix jacchus). Am J Primatol 77:222–228
Carter GG (2015) Cooperation and social bonds in common vampire bats. PhD thesis
Carter TC, Feldhamer GA (2005) Roost tree use by maternity colonies of Indiana bats and northern long-eared bats in southern Illinois. For Ecol Manage 219:259–268
Carter GG, Wilkinson GS (2013a) Food sharing in vampire bats: reciprocal help predicts donations more than relatedness or harassment. Proc R Soc 280:20122573
Carter GG, Wilkinson GS (2013b) Cooperation and conflict in the social lives of bats. In: Adams RA, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, xvi + 547 pp, pp 225–242
Carter GG, Skowronski MD, Faure PA, Fenton B (2008) Antiphonal calling allows individual discrimination in white-winged vampire bats. Anim Behav 76:1343–1355
Carter GG, Fenton MB, Faure PA (2009) White-winged vampire bats (Diaemus youngi) exchange contact calls. Can J Zool 87:604–608
Castella V, Ruedi M, Excoffier L (2001) Contrasted patterns of mitochondrial and nuclear structure among nursery colonies of the bat Myotis myotis. J Evol Biol 14:708–720
Chaverri G (2010) Comparative social network analysis in a leaf-roosting bat. Behav Ecol Sociobiol 64:1619–1630
Chaverri G, Gillam EH (2010) Cooperative signaling behavior of roost location in a leaf roosting bat. Communicative Integr Biol 3:599–601
Chaverri G, Gillam EH, Vonhof MJ (2010) Social calls used by a leaf-roosting bat to signal location. Biol Lett 6:441–444
Chaverri G, Gillam EH, Kunz TH (2012) A call-and-response system facilitates group cohesion among disc-winged bats. Behav Ecol 24:481–487
Choe JC, Crespi BJ (eds) (1997) The evolution of social behavior in insects and arachnids. Cambridge University Press, Cambridge, vii + 541 pp
Christe P, Arlettaz R, Vogel P (2000) Variation in intensity of a parasitic mite (Spinturnix myoti) in relation to the reproductive cycle and immunocompetence of its bat host (Myotis myotis). Ecol Lett 3:207–212
Christe P, Glaizot O, Evanno G, Bruyndonckx N, Devevey G, Yannic G, Patthey P, Maeder A, Vogel P, Arlettaz R (2007) Host sex and ectoparasites choice: preference for, and higher survival on female hosts. J Anim Ecol 76:703–710
Clement MJ, Castleberry SB (2013) Tree structure and cavity microclimate: implications for bats and birds. Int J Biometeorol 57:437–450
Clutton-Brock T, Janson C (2012) Primate socioecology at the crossroads: past, present, and future. Evol Anthropol 21:136–150
Connor RC, Wells RS, Mann J, Read AJ (2000) The bottlenose dolphin. Social relationships in a fission-fusion society. In: Mann J, Connor RC, Tyack PL, Whitehead H (eds) Cetacean societies: field studies of dolphins and whales. University of Chicago Press, Chicago, xiv + 433 pp, pp 91–126
Couzin ID (2006) Behavioral ecology: social organization in fission-fusion societies. Curr Biol 16:R169–R171
Couzin ID, Laidre ME (2009) Fission–fusion populations. Curr Biol 19:633–635
Croft DP, James R, Krause J (2008) Exploring animal social networks. Princeton University Press, Princeton, vii + 192 pp
Cross PC, Lloyd-Smith JO, Getz WM (2005) Disentangling association patterns in fission-fusion societies using African buffalo as an example. Anim Behav 69:499–506
De Fanis E (1995) The role of odour in the discrimination of conspecifics by pipistrelle bats. Anim Behav 49:835–837
Dechmann DKN, Heucke SL, Giuggioli L, Safi K, Voigt CC, Wikelski M (2009) Experimental evidence for group hunting via eavesdropping in echolocating bats. Proc R Soc B 276:2721–2728
Dechmann DKN, Kranstauber B, Gibbs D, Wikelski M (2010) Group hunting-a reason for sociality in molossid bats? PLoS ONE 5:e9012. doi:10.1371/journal.pone.0009012
Dick CW, Dittmar K (2014) Parasitic bat flies (Diptera : Streblidae and Nycteribiidae): host specificity and potential as vectors. In: Klimpel S, Mehlhorn H (eds) Bats (Chiroptera) as vectors of diseases and parasites, Parasitology research monographs 5. Springer, New York. xii + 187 pp, pp 131–156
Dunbar RIM, Shultz S (2010) Bondedness and sociality. Behaviour 147:775–803
Ellison LA, O’Shea TJ, Neubaum DJ, Bowen RA (2007) Factors influencing movement probabilities of big brown bats (Eptesicus fuscus) in buildings. Ecol Appl 17:620–627
Englert AC, Greene MJ (2009) Chemically-mediated roostmate recognition and roost selection by Brazilian free-tailed bats (Tadarida brasiliensis). PLoS ONE 4:e7781
Entwistle AC, Racey PA, Speakman JR (2000) Social and population structure of a gleaning bat, Plecotus auritus. J Zool 252:11–17
Esbérard CEL, Jesus AC, Motta AG, Bergallo HG, Gettinger D (2005) Hesperoctenes fumarius (Hemiptera: Polyctenidae) infesting Molossus rufus (Chiroptera: Molossidae) in Southeastern Brazil. J Parasitol 91:465–467
Evans LN, Lumsden LF (2011) A comparison of the roosting behaviour of Gould’s wattled bats Chalinolobus gouldii using bat boxes and tree hollows in suburban Melbourne. In: Law B, Eby P, Lunney D, Lumsden L (eds) The biology and conservation of Australian bats. Royal Zoological Society of New South Whales, New South Whales. xi + 498 pp, pp 289–296
Evelyn MJ, Stiles DA, Young RA (2004) Conservation of bats in suburban landscapes: roost selection by Myotis yumanensis in a residential area in California. Biol Conserv 115:463–473
Feyerabend F, Simon M (2000) Use of roosts and roost switching in a summer colony of 45 kHz phonic type pipistrelle bats (Pipistrellus pipistrellus Schreber, 1774). Myotis 38:51–59
Flanders J, Jones G, Benda P, Dietz C, Zhang S, Li G, Sharifi M, Rossiter SJ (2009) Phylogeography of the greater horseshoe bat, Rhinolophus ferrumequinum: contrasting results from mitochondrial and microsatellite data. Mol Ecol 18:306–318
Fleischmann D, Kerth G (2014) Roosting behavior and group decision making in 2 syntopic bat species with fission-fusion societies. Behav Ecol 25:1–8
Fleischmann D, Baumgartner IO, Erasmy M, Gries N, Melber M, Leinert V, Parchem M, Reuter M, Schaer P, Stauffer S, Wagner I, Kerth G (2013) Female bechstein’s bats adjust their group decisions about communal roosts to the level of conflict of interests. Curr Biol 23:1658–1662
Fornůsková A, Petit EJ, Bartonička T, Kaňuch P, Butet A, Řehák Z, Bryja J (2014) Strong matrilineal structure in common pipistrelle bats (Pipistrellus pipistrellus) is associated with variability in echolocation calls. Biol J Linn Soc 113:1115–1125
Fortuna MA, Popa-Lisseanu AG, Ibáñez C, Bascompte J (2009) The roosting spatial network of a bird-predator bat. Ecology 90:934–944
Furmankiewicz J, Ruczyński I, Urban R, Jones G (2011) Social calls provide tree-dwelling bats with information about the location of conspecifics at roosts. Ethology 117:480–489
Garroway CJ, Broders HG (2007) Nonrandom association patterns at northern long-eared bat maternity roosts. Can J Zool 85:956–964
Gaudet CL, Fenton MB (1984) Observational learning in three species of insectivorous bats (Chiroptera). Anim Behav 32:385–388
Gillam EH (2007) Eavesdropping by bats on the feeding buzzes of conspecifics. Can J Zool 85:795–801
Gillam EH, Chaverri G (2012) Strong individual signatures and weaker group signatures in contact calls of Spix’s disc-winged bat, Thyroptera tricolor. Anim Behav 83:269–276
Gillam EH, Chaverri G, Montero K, Sagot M (2013) Social calls produced within and near the roost in two species of tent-making bats, Dermanura watsoni and Ectophylla alba. PLoS ONE 8:e61731
Godinho LN, Cripps JK, Coulson G, Lumsden LF (2013) The effect of ectoparasites on the grooming behaviour of Gould’s wattled bat (Chalinolobus gouldii): an experimental study. Acta Chiropterologica 15:463–472
Hamilton WD (1964) The genetical evolution of social behaviour. I. J Theor Biol 7:1–16
Hillen J, Kiefer A, Veith M (2010) Interannual fidelity to roosting habitat and flight paths by female western barbastelle bats. Acta Chiropterologica 12:187–195
Hinde RA (1976) Interactions, relationships and social structure. Man 11:1–17
Hristov NI, Allen LC, Chadwell BA (2013) New advances in the study of group behavior in bats. In: Adams R, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, xvi + 547 pp, pp 271–292
Johnson JS, Kropczynski JN, Lacki MJ (2013) Social network analysis and the study of sociality in bats. Acta Chiropterologica 15:1–17
Johnson JS, Kropczynski JN, Lacki MJ, Langlois GD (2012) Social networks of Rafinesque’s big-eared bats (Corynorhinus rafinesquii). J Mammal 93:1545–1558
Jones PL, Ryan MJ, Page RA (2014) Population and seasonal variation in response to prey calls by an eavesdropping bat. Behav Ecol Sociobiol 68:605–615
Kanwal JS, Zhang Z, Feng J (2013) Decision-making and socioemotional vocal behavior in bats. In: Adams R, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, xvi + 547 pp, pp 243–270
Kao AB, Miller N, Torney C, Hartnett A, Couzin ID (2014) Collective learning and optimal consensus decisions in social animal groups. PLoS Comput Biol 10:e1003762
Kashima K, Ohtsuki H, Satake A (2013) Fission-fusion bat behavior as a strategy for balancing the conflicting needs of maximizing information accuracy and minimizing infection risk. J Theor Biol 318:101–109
Kazial KA, Kenny TL, Burnett SC (2008) Little brown bats (Myotis lucifugus) recognize individual identity of conspecifics using sonar calls. Ethology 114:469–478
Kerth G (2008a) Causes and consequences of sociality in bats. Bioscience 58:737–746
Kerth G (2008b) Animal sociality: bat colonies are founded by relatives. Curr Biol 18:R740–R742
Kerth G (2010) Group decision-making in fission-fusion societies. Behav Process 84:662–663
Kerth G, König B (1999) Fission, fusion and nonrandom associations in female Bechstein’s bats (Myotis bechsteinii). Behaviour 136:1187–1202
Kerth G, Petit E (2005) Colonization and dispersal in a social species, the Bechstein’s bat (Myotis bechsteinii). Mol Ecol 14:3943–3950
Kerth G, Reckardt K (2003) Information transfer about roosts in female Bechstein’s bats: an experimental field study. Proc R Soc B 270:511–515
Kerth G, Van Schaik J (2011) Causes and consequences of living in closed societies: lessons from a long-term socio-genetic study on Bechstein’s bats. Mol Ecol 21:636–646
Kerth G, Mayer F, König B (2000) Mitochondrial DNA (mtDNA) reveals that female Bechstein’s bats live in closed societies. Mol Ecol 9:793–800
Kerth G, Wagner M, König B (2001a) Roosting together, foraging apart: information transfer about food is unlikely to explain sociality in female Bechstein’s bats (Myotis bechsteinii). Behav Ecol Sociobiol 50:283–291
Kerth G, Weissmann K, König B (2001b) Day roost selection in female Bechstein’s bats (Myotis bechsteinii): a field experiment to determine the influence of roost temperature. Oecologia 126:1–9
Kerth G, Mayer F, Petit E (2002a) Extreme sex-biased dispersal in the communally breeding, nonmigratory Bechstein’s bat (Myotis bechsteinii). Mol Ecol 11:1491–1498
Kerth G, Safi K, König B (2002b) Mean colony relatedness is a poor predictor of colony structure and female philopatry in the communally breeding Bechstein’s bat (Myotis bechsteinii). Behav Ecol Sociobiol 52:203–210
Kerth G, Almasi B, Ribi N, Thiel D, Lupold S (2003) Social interactions among wild female Bechstein’s bats (Myotis bechsteinii) living in a maternity colony. Acta Ethologica 5:107–114
Kerth G, Ebert C, Schmidtke C (2006) Group decision making in fission-fusion societies: evidence from two-field experiments in Bechstein’s bats. Proc R Soc B 273:2785–2790
Kerth G, Perony N, Schweitzer F (2011) Bats are able to maintain long-term social relationships despite the high fission-fusion dynamics of their groups. Proc R Soc B 278:2761–2767
Kilgour RJ, Brigham RM (2013) The relationships between behavioural categories and social influences in the gregarious big brown bat (Eptesicus fuscus). Ethology 119:189–198
Kilgour RJ, Faure PA, Brigham RM (2013) Evidence of social preferences in big brown bats (Eptesicus fuscus). Can J Zool 760:756–760
Kingston T (2013) Response of bat diversity to forest disturbance in Southeast Asia: insights from long-term research in Malaysia. In: Adams R, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, xvi + 547 pp, pp 169–186
Korine C, Daniel S, Pinshow B (2013) Roost selection by female Hemprich’s long-eared bats. Behav Process 199:131–138
Krause J, Ruxton GD (2002) Living in groups. Oxford University Press, Oxford, xiv + 228 pp
Krebs JR, Davies NB (1995) An introduction to behavioural ecology, 3rd edition. Blackwell, Oxford, xii + 420 pp
Kunz TH, Fenton MB (eds) (2003) Bat ecology. University of Chicago Press, Chicago, xix, 779 pp
Kunz TH, Lumsden LF (2003) Ecology of cavity and foliage roosting bats. In: Kunz TH, Fenton MB (eds) Bat ecology. University of Chicago Press, Chicago, xix, 779 pp, pp 3–89
Lacki MJ, Schwierjohann JH (2001) Day-roost characteristics of northern bats in mixed mesophytic forest. J Wildl Manage 65:482–488
Langwig KE, Frick WF, Bried JT, Hicks AC, Kunz TH, Kilpatrick MA (2012) Sociality, density-dependence and microclimates determine the persistence of populations suffering from a novel fungal disease, white-nose syndrome. Ecol Lett 15:1050–1057
Lausen CL, Barclay RMR (2006) Benefits of living in a building: big brown bats (Eptesicus fuscus) in rocks versus buildings. J Mammal 87:362–370
Lehmann J, Boesch C (2004) To fission or to fusion: effects of community size on wild chimpanzee (Pan troglodytes verus) social organisation. Behav Ecol Sociobiol 56:207–216
Lewis SE (1995) Roost fidelity of bats: a review. J Mammal 76:481–496
Lewis SE (1996) Low roost-site fidelity in pallid bats: associated factors and effect on group stability. Behav Ecol Sociobiol 39:335–344
Lima SL, O’Keefe JM (2013) Do predators influence the behaviour of bats? Biol Rev 88:626–644
Lourenço SI, Palmeirim JM (2004) Influence of temperature in roost selection by Pipistrellus pygmaeus (Chiroptera): relevance for the design of bat boxes. Biol Conserv 119:237–243
Lourenço SI, Palmeirim JM (2007) Can mite parasitism affect the condition of bat hosts? Implications for the social structure of colonial bats. J Zool 273:161–168
Lusseau D, Wilson B, Hammond PS (2006) Quantifying the influence of sociality on population structure in bottlenose dolphins. J Anim Ecol 75:14–24
Lučan RK, Radil J (2010) Variability of foraging and roosting activities in adult females of Daubenton’s bat (Myotis daubentonii) in different seasons. Biologia 65:1072–1080
Melber M, Fleischmann D, Kerth G (2013) Female Bechstein’s bats share foraging sites with maternal kin but do not forage together with them—results from a long-term study. Ethology 119:793–801
Melendez KV, Feng AS (2010) Communication calls of little brown bats display individual-specific characteristics. J Acoust Soc Am 128:919–923
Metheny JD, Kalcounis-Rueppell MC, Willis CKR, Kolar KA, Brigham RM (2008a) Genetic relationships between roost-mates in a fission–fusion society of tree-roosting big brown bats (Eptesicus fuscus). Behav Ecol Sociobiol 62:1043–1051
Metheny JD, Kalcounis-Rueppell MC, Bondo KJ, Brigham RM (2008b) A genetic analysis of group movement in an isolated population of tree-roosting bats. Proc R Soc B 275:2265–2272
Michaelsen TC, Jensen KH, Högstedt G (2014) Roost site selection in pregnant and lactating soprano pipistrelles (Pipistrellus pygmaeus Leach, 1825) at the species northern extreme: the importance of warm and safe roosts. Acta Chiropterologica 16:349–357
Mikula P, Hromada M, Tryjanowski P (2013) Bats and swifts as food of the European kestrel (Falco tinnunculus) in a small town in Slovakia. Ornis Fennica 90:178–185
Montes C, Cuadrillero C, Vilella D (2002) Maintenance of a laboratory colony of Cimex lectularius (Hemiptera: Cimicidae) using an artificial feeding technique. J Med Entomol 39:675–679
Naďo L, Kaňuch P (2013) Dawn swarming in tree-dwelling bats—an unexplored behaviour. Acta Chiropterologica 15:387–392
O’Donnell CFJ (2000) Cryptic local populations in a temperate rainforest bat Chalinolobus tuberculatus in New Zealand. Anim Conserv 3:287–297
O’Donnell CFJ, Sedgeley JA (1999) Use of roosts by the long-tailed bat, Chalinolobus tuberculatus, in temperate rainforest in New Zealand. J Mammal 80:913–923
O’Mara MT, Dechmann DKN, Page RA (2014) Frugivorous bats evaluate the quality of social information when choosing novel foods. Behav Ecol 25:1233–1239
Olson CR, Barclay RMR (2013) Concurrent changes in group size and roost use by reproductive female little brown bats (Myotis lucifugus). Can J Zool 91:149–155
Page RA, Ryan MJ (2006) Social transmission of novel foraging behavior in bats: frog calls and their referents. Curr Biol 16:1201–1205
Papadatou E, Butlin RK, Altringham JD (2008) Seasonal roosting habits and population structure of the long-fingered bat Myotis capaccinii in Greece. J Mammal 89:503–512
Patriquin KJ (2012) The causes and consequences of fission-fusion dynamics in female northern long-eared bats (Myotis septentrionalis). PhD thesis
Patriquin KJ, Leonard ML, Broders HG, Garroway CJ (2010) Do social networks of female northern long-eared bats vary with reproductive period and age? Behav Ecol Sociobiol 64:899–913
Patriquin KJ, Palstra F, Leonard ML, Broders HG (2013) Female northern myotis (Myotis septentrionalis) that roost together are related. Behav Ecol 24:949–954
Patterson BD, Dick CW, Dittmar K (2007) Roosting habits of bats affect their parasitism by bat flies (Diptera: Streblidae). J Trop Ecol 23:177–189
Pearce RD, O’Shea TJ (2007) Ectoparasites in an urban population of big brown bats (Eptesicus fuscus) in Colorado. J Parasitol 93:518–530
Pfalzer G, Kusch J (2003) Structure and variability of bat social calls: implications for specificity and individual recognition. J Zool Lond 261:21–33
Popa-Lisseanu AG, Bontadina F, Mora O, Ibáñez C (2008) Highly structured fission–fusion societies in an aerial-hawking, carnivorous bat. Anim Behav 75:471–482
Postawa T, Furman A (2014) Abundance patterns of ectoparasites infesting different populations of Miniopterus species in their contact zone in Asia Minor. Acta Chiropterologica 16:387–395
Postawa T, Gas A (2009) Do the thermal conditions in maternity colony roost determine the size of young bats? Comparison of attic and cave colonies of Myotis myotis in Southern Poland. Folia Zool 58:396–408
Postawa T, Szubert-Kruszyńska A (2014) Is parasite load dependent on host aggregation size? The case of the greater mouse-eared bat Myotis myotis (Mammalia: Chiroptera) and its parasitic mite Spinturnix myoti (Acari: Gamasida). Parasitol Res 113:1803–1811
Postawa T, Szubert-Kruszynska A, Ferenc H (2014) Differences between populations of Spinturnix myoti (Acari: Mesostigmata) in breeding and non-breeding colonies of Myotis myotis (Chiroptera) in central Europe: the effect of roost type. Folia Parasitol 61:581–588
Pretzlaff I, Kerth G, Dausmann KH (2010) Communally breeding bats use physiological and behavioural adjustments to optimise daily energy expenditure. Die Naturwissenschaften 97:353–363
Ramos-Fernández G, Boyer D, Gómez VP (2006) A complex social structure with fission–fusion properties can emerge from a simple foraging model. Behav Ecol Sociobiol 60:536–549
Ratcliffe JM, ter Hofstede HM (2005) Roosts as information centres: social learning of food preferences in bats. Biol Lett 1:72–74
Reckardt K, Kerth G (2006) The reproductive success of the parasitic bat fly Basilia nana (Diptera: Nycteribiidae) is affected by the low roost fidelity of its host, the Bechstein’s bat (Myotis bechsteinii). Parasitol Res 98:237–243
Reckardt K, Kerth G (2007) Roost selection and roost switching of female Bechstein’s bats (Myotis bechsteinii) as a strategy of parasite avoidance. Oecologia 154:581–588
Rhodes M (2007) Roost fidelity and fission-fusion dynamics of white-striped free-tailed bats (Tadarida australis). J Mammal 88:1252–1260
Ross G, Holderied MW (2013) Learning and memory in bats: a case study on object discrimination in flower-visiting bats. In: Adams R, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, xvi + 547 pp, pp 207–224
Ruczyński I, Bartoń KA (2012) Modelling sensory limitation: the role of tree selection, memory and information transfer in bats’ roost searching strategies. PLoS ONE 7:e44897
Ruczyński I, Kalko EKV, Siemers BM (2007) The sensory basis of roost finding in a forest bat, Nyctalus noctula. J Exp Biol 210:3607–3615
Ruczyński I, Kalko EKV, Siemers BM (2009) Calls in the forest: a comparative approach to how bats find tree cavities. Ethology 115:167–177
Russell YI, Phelps S (2013) How do you measure pleasure? A discussion about intrinsic costs and benefits in primate allogrooming. Biol Philos 28:1005–1020
Russo D, Cistrone L, Jones G (2005) Spatial and temporal patterns of roost use by tree-dwelling barbastelle bats Barbastella barbastellus. Ecography 28:769–776
Safi K (2008) Social bats: the males’ perspective. J Mammal 89:1342–1350
Safi K, Kerth G (2003) Secretions of the interaural gland contain information about individuality and colony membership in the Bechstein’s bat. Anim Behav 65:363–369
Sagot M, Stevens RD (2012) The evolution of group stability and roost lifespan: perspectives from tent-roosting bats. Biotropica 44:90–97
Schino G, Aureli F (2009) Reciprocal altruism in primates: partner choice, cognition, and emotions. Adv Study Behav 39:45–69
Schmidt S (2013) Beyond echolocation: emotional acoustic communication in bats. In: Altenmüller E, Schmidt S, Zimmermann E (eds) Evolution of emotional communication: from sounds in nonhuman mammals to speech and music in man. Oxford University Press, Oxford, xiv + 376 pp, pp 301–315
Schöner CR, Schöner MG, Kerth G (2010) Similar is not the same: social calls of conspecifics are more effective in attracting wild bats to day roosts than those of other bat species. Behav Ecol Sociobiol 64:2053–2063
Sedgeley JA (2001) Quality of cavity microclimate as a factor influencing selection of maternity roosts by a tree-dwelling bat, Chalinolobus tuberculatus, in New Zealand. J Appl Ecol 38:425–438
Silk J (2007) The adaptive value of sociality in mammalian groups. Philos Trans R Soc B 362:539–559
Silvis A, Ford WM, Britzke ER, Johnson JB (2014a) Association, roost use and simulated disruption of Myotis septentrionalis maternity colonies. Behav Process 103:283–290
Silvis A, Kniowski AB, Gehrt SD, Ford WM (2014b) Roosting and foraging social structure of the endangered Indiana bat (Myotis sodalis). PLoS ONE 9:e96937
Storz JF, Bhat HR, Kunz TH (2000) Social structure of a polygynous tent-making bat, Cynopterus sphinx (Megachiroptera). J Zool 251(2):151–165
Sueur C, King AJ, Conradt L, Kerth G, Lusseau D, Mettke-Hofmann C, Schaffner CM, Williams L, Zinner D, Aureli F (2011) Collective decision-making and fission-fusion dynamics: a conceptual framework. Oikos 120:1608–1617
ter Hofstede HM, Fenton MB (2005) Relationships between roost preferences, ectoparasite density, and grooming behaviour of neotropical bats. J Zool 266:333–340
Terrien J, Perret M, Aujard F (2011) Behavioral thermoregulation in mammals: a review. Front Biosci 16:1428–1444
Threlfall C, Law B, Banks PB (2013) Odour cues influence predation risk at artificial bat roosts in urban bushland. Biol Lett 9:20121144
Trevelin LC, Silveira M, Port-Carvalho M, Homem DH, Cruz-Neto AP (2013) Use of space by frugivorous bats (Chiroptera: Phyllostomidae) in a restored Atlantic forest fragment in Brazil. For Ecol Manage 291:136–143
Trousdale AW, Beckett DC, Hammond SL (2008) Short-term fidelity of Rafinesque’s big-eared bat (Corynorhinus rafinesquii). J Mammal 89:477–484
Turbill C, Geiser F (2006) Thermal physiology of pregnant and lactating female and male long-eared bats, Nyctophilus geoffroyi and N. gouldi. J Comp Physiol B 176:165–172
Veilleux JP, Veilleux SL (2004) Intra-annual and interannual fidelity to summer roost areas by female eastern pipistrelles, Pipistrellus subflavus. Am Midl Nat 152:196–200
Voigt-Heucke SL, Taborsky M, Dechmann DKN (2010) A dual function of echolocation: bats use echolocation calls to identify familiar and unfamiliar individuals. Anim Behav 80:59–67. doi:10.1016/j.anbehav.2010.03.025
Vonhof MJ, Wilkinson L (1999) Roosting habitat requirements of northern long-eared bats (Myotis septentrionalis) in the boreal forests of northeastern British Columbia: year 2. Report prepared for the B.C. Ministry of Environment, Lands and Parks, Fort St. John, 88 pp
Vonhof MJ, Whitehead H, Fenton MB (2004) Analysis of Spix’s disc-winged bat association patterns and roosting home ranges reveal a novel social structure among bats. Anim Behav 68:507–521
Webber QMR, McGuire LP, Smith SB, Willis CKR (2015) Host behaviour, age and sex correlate with ectoparasite prevalence and intensity in a colonial mammal, the little brown bat. Behaviour 152:83–105
Whitehead H (2007) Learning, climate and the evolution of cultural capacity. J Theor Biol 245:341–350
Whitehead H (2008) Analyzing animal societies: quantitative methods for vertebrate social analysis. University of Chicago Press, Chicago 336 pp
Wilkinson GS (1985) The social organization of the common vampire bat. I. Pattern and cause of association. Behav Ecol Sociobiol 17:111–121
Wilkinson GS (1986) Social grooming in the common vampire bat, Desmodus rotundus. Anim Behav 34:1880–1889
Wilkinson GS (1992) Communal nursing in the evening bat, Nycticeius humeralis. Behav Ecol Sociobiol 31:225–235
Willis CKR, Brigham RM (2004) Roost switching, roost sharing and social cohesion: forest-dwelling big brown bats, Eptesicus fuscus, conform to the fission-fusion model. Anim Behav 68:495–505
Willis CKR, Brigham RM (2007) Social thermoregulation exerts more influence than microclimate on forest roost preferences by a cavity-dwelling bat. Behav Ecol Sociobiol 62:97–108
Willis CKR, Voss CM, Brigham RM (2006) Roost selection by forest-living female big brown bats (Eptesicus fuscus). J Mammal 87:345–350
Wittemyer G, Douglas-Hamilton I, Getz WM (2005) The socioecology of elephants: analysis of the processes creating multitiered social structures. Anim Behav 69:1357–1371
Wright GS, Wilkinson GS, Moss CF (2011) Social learning of a novel foraging task by big brown bats (Eptesicus fuscus). Anim Behav 82:1075–1083
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Patriquin, K.J., Ratcliffe, J.M. (2016). Should I Stay or Should I Go? Fission–Fusion Dynamics in Bats. In: Ortega, J. (eds) Sociality in Bats. Springer, Cham. https://doi.org/10.1007/978-3-319-38953-0_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-38953-0_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-38951-6
Online ISBN: 978-3-319-38953-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)