
Abstract
The intestinal epithelial barrier is critical for maintaining a balance between potentially noxious luminal contents of the gut and the mucosal immune system. Defects in barrier function are associated with both Crohn’s disease and ulcerative colitis, and are also present in some healthy first-degree relatives. Impaired barrier function is associated with increased risk of Crohn’s disease relapse of patients in clinical remission. The tight junction, which seals the space between adjacent epithelial cells, is the primary determinant of permeability in the absence of epithelial injury, e.g. ulceration. The tight junction is formed by a complex of occludin, claudins, ZO-1, and the actomyosin cytoskeleton; the interactions between components are dynamically regulated to modify paracellular flux. The functional properties of the tight junction can be modified in response to either physiological stimuli of cytokines, e.g. TNF, IFNγ, and IL-13. Under pathological conditions, increased paracellular permeability may allow luminal material to access the lamina propria, thereby enhancing immune activation and triggering a recurrent cycle of barrier dysfunction and inflammation. Tight junction dysregulation may thereby play a central role in inflammatory bowel disease by linking mucosal immune responses to barrier function. As such, the potential of the tight junction as a target for therapeutic intervention should be considered.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others
Keywords
Mucosal surfaces are lined by epithelial cells, which form a barrier between the internal and external environment . The integrity of this barrier is critical, particularly within the intestine, where the luminal environment includes potentially pathogenic antigens and microorganisms. Although the term “barrier” may imply that the mucosa simply acts as a static impediment, preventing all transepithelial flux, this is not the case, as the epithelium must absorb and secrete the water, ions, and macromolecules that are necessary to maintain intestinal homeostasis and overall nutrition. Thus, the mucosa must integrate active, vectorial, and passive transport with a selectively permeable barrier that prevents entry of noxious luminal materials.
“Intestinal barrier function ” is often used indiscriminately but is commonly thought of as either (1) the epithelial cells and materials they secrete, which provide a physical impediment to ion, water, and macromolecular flux, or (2) the defense provided by the mucosal immune system, which protects the host from potentially pathogenic luminal contents that have crossed the physical barrier. While both types of barriers are essential to epithelial homeostasis, the primary focus of this chapter is the former, i.e. the barrier formed by epithelial cells and the materials they secrete. The latter include mucins, secreted by intestinal goblet cells, which form a hydrated gel over the epithelial surface. As discussed elsewhere defects in mucin synthesis and the extracellular mucin layer have been associated with inflammatory bowel disease (IBD) in humans and spontaneous colitis in mice.
While mucin contributes to the development of the unstirred layer by limiting passage of large materials and trapping some bacteria, it is the epithelial cells that form the principal barrier to water and solute flux. The lipid bilayer of epithelial cells serves as a barrier to most hydrophilic solutes, and therefore, the majority of passive flux across epithelia occurs via the paracellular route. Changes in epithelial state can affect three distinct flux routes : the paracellular pore and leak pathways and the unrestricted pathway. The pore pathway is a high-capacity, size- and charge-selective route that mediates flux of small ions and water. In contrast, the leak pathway is a low-capacity, relatively size- and charge-non-selective pathway that facilitates passive transport of larger macromolecules. In the presence of erosions or ulcerations the epithelium is lost and, therefore, does not contribute to barrier function. In this case, flux of water, ions, macromolecules, and bacteria is unrestricted in terms of both capacity and selectivity . Molecules that permeate denuded epithelia are therefore said to cross the “unrestricted pathway.” It is important to note that these pathways are not absolute, and as discussed below, there is cross-talk between elements of each pathway. When the unrestricted pathway is sealed, i.e. in the presence of an intact epithelial layer, the intercellular tight junctions define the selective permeability of the mucosal barrier .
The Epithelial Barrier Is Regulated in Response to Physiologic Stimuli
The intestinal epithelium is charged with the complex task of serving as a barrier that separates internal body cavities from potentially noxious luminal contents while simultaneously facilitating nutrient, ion, and water absorption and secretion. Physiologic regulation of epithelial barrier function has been studied extensively in the context of Na+-glucose cotransport [1–4]. Upon activation of Na+-glucose cotransport, a transepithelial osmotic gradient is developed and epithelial myosin light chain kinase (MLCK) is activated. Together, these changes result in increased pore pathway flux, passive water absorption, and solvent-drag-mediated absorption of nutrient-sized molecules, such as glucose [1].
The Intestinal Barrier Is Compromised in IBD
Epithelial barrier function is compromised in many intestinal disorders. The association between decreased barrier function and intestinal disease was first reported in the early 1980s [5–7]. These studies, which made use of both ex vivo and in vivo approaches, demonstrated increased permeability in Crohn’s disease (CD ) and ulcerative colitis (UC ) in the absence, as well as the presence, of ulcerations. Barrier defects are also present in conditions not typically associated with erosion and ulceration, e.g. celiac disease [8, 9], and in some healthy first-degree relatives of CD patients [10].
In patients, measurement of intestinal permeability typically involves measuring fractional urinary excretion of orally administered small molecules, such as 51Cr-EDTA , polyethylene glycols , mannitol, lactulose, sucralose, or creatinine [11–14]. Intestinal absorption of these molecules occurs freely across erosions. When the epithelium is intact absorption is more restricted and defined by the tight junctions . These tracers are not metabolized in the circulation and are freely filtered at the glomerulus. Thus, fractional urinary recovery can be used as a noninvasive measure of intestinal permeability, assuming normal vascular perfusion, renal function, and intestinal motility. Lactulose and mannitol are used most commonly. With a small radius, mannitol is capable of crossing via the paracellular pore pathway, while lactulose is larger and can cross at sites of epithelial damage or through the paracellular leak pathway. Fractional excretion of lactulose can therefore be thought of as a measure of leak pathway permeability and intestinal damage while mannitol excretion can be used as a measure of intestinal surface area. Assessing the lactulose to mannitol ratio (LAMA) therefore corrects for confounding factors such as epithelial surface area, intestinal transit time, and renal function [12]. However, lactulose and mannitol are partially degraded by luminal bacteria within the colon, making these probes unsuitable analysis of colonic permeability [15, 16]. As a result, permeability has been studied to a far greater extent in patients with small intestinal Crohn's disease than in patients with Crohn's colitis or ulcerative colitis.
The contribution of the intestinal barrier to IBD was first suggested following the observation that increased permeability is present in a subset of healthy, asymptomatic first-degree relatives of CD patients [10, 17, 18]. The exact nature of these barrier defects is not known, and increased intestinal permeability may stem from increased susceptibility to epithelial ulceration, tight junction barrier defects, or subclinical immune activation. However, analysis of some of these healthy subjects failed to identify erosions or ulcers. Nevertheless, one clinical trial found hypersensitivity to NSAID-mediated increases in intestinal permeability in subjects with genetic susceptibility to CD, i.e. first-degree relatives [19]. Additional studies have linked barrier defects in healthy first-degree relatives to specific CD-associated mutations in NOD2, an immunoregulatory gene [20]. Finally, barrier loss in IBD has been correlated with altered tight junction organization, protein composition, cytoskeletal regulation, and epithelial damage [21–28]. It is important to note that these findings are not mutually exclusive as heightened immune activation may lead to increased tight junction permeability and ultimately epithelial damage.
Although the contribution of barrier defects in disease pathogenesis is unclear, it is interesting to note a case report describing the development of CD in a previously healthy relative with increased intestinal permeability [29]. While this single patient could be interpreted as support for a link between abnormal intestinal permeability and subsequent disease development, it is important to keep in mind that risk of disease was increased in this individual on a genetic basis. Indeed, first-degree relatives of patients affected by CD have an elevated risk of developing IBD [30]. Unfortunately, no studies have compared the long-term risk of developing CD in healthy relatives with or without increased permeability. However, increased small intestinal permeability is a known marker of impending relapse in CD patients with inactive disease [31].
The Epithelial Monolayer
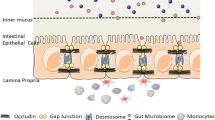
In a reductionist view, the intestinal barrier is formed by a single layer of epithelial cells linked together by intercellular junctions. In the absence of specific transporters, the epithelial plasma membrane serves as a barrier to most hydrophilic molecules; thus, intercellular junctions are the limiting factor in transepithelial permeability. The most critical intercellular junctions are the tight junctions, adherens junctions, and desmosomes, which form the apical junctional complex [32]. The tight junction and adherens junctions are associated with a dense perijunctional ring of actin and myosin II that encircles the apical aspect of each epithelial cell. These form a network that connects neighboring cells and forms the selectively permeable paracellular barrier (Fig. 3.1a).

Structure of the epithelial barrier . (a) Transmission electron micrograph depicting the apical junctional complex between two mouse enterocytes. From apical to basal, the apical junctional complex consists of the zonula occludens, also referred to as the tight junction (TJ); the zonula adherens, also referred to as the adherens junction (AJ); and macula adherens, also referred to as the desmosome. Microvilli (Mv) line the luminal surface of intestinal epithelia and are labeled for orientation. (b) The tight junction is spanned by occludin and claudin family members, which dynamically regulate permeability between cells. The plaque protein zonula ocludens-1 (ZO-1) interacts with transmembrane tight junction proteins and with the actomyosin cytoskeleton to regulate tight junction function. Myosin light chain kinase (MLCK) controls junction-associated myosin phosphorylation and myosin ATPase activity to regulate barrier function. (c) The cytosolic tight junction scaffolding protein ZO-1 is essential for cortical actin organization. Fluorescence microscopy (top two panels) and scanning electron microscopy (bottom two panels) of confluent epithelial monolayers show that ZO-1 deletion results in marked abnormalities of apical structure. Top panels are maximum intensity projections of confocal z-series through the entire epithelial monolayer. Scale bars = 5 μm
Molecular Anatomy of the Tight Junction
The adherens junction is formed by homotypic interactions between the epithelial cadherin isoform E-cadherin (Fig. 3.1B). The cytoplasmic domains of the cadherins interact with the catenins, which in turn bind to the actin cytoskeleton to regulate cell structure and function [33–35]. The tight junction consists of many transmembrane proteins, including occludin and claudins [36–39] (Fig. 3.1b). Transmembrane tight junction proteins are linked to one another and to the actomyosin cytoskeleton by the cytosolic scaffolding protein zonula occludens (ZO)-1, thereby forming an elaborate protein network at the apical junctional complex [40–43]. The extracellular loops of claudins mediate homotypic interactions to form high-capacity size- and charge-selective paracellular pores [44, 45]. In contrast, occludin , ZO-1, and the actomyosin cytoskeleton are known to regulate leak pathway permeability [46–49]. These molecular components interact and can regulate both pathways, as occludin and ZO-1, which primarily regulate the leak pathway, can also affect claudin pore function [50]. ZO-1 may affect paracellular permeability through regulation of cortical actomyosin organization and contractile activity [43, 51]. Moreover, by integrating cortical actomyosin function, ZO-1 directs a diverse array of cellular processes; ZO-1 depletion has marked effects on epithelial morphology (Fig. 3.1c).
In vitro and in vivo studies have revealed that tight junction assembly is closely linked to adherens junction integrity, as confluent epithelial monolayers lacking E-cadherin or alpha-catenin are unable to efficiently recruit tight junction proteins or form effective barriers to ions and macromolecular flux [52–55].
Intercellular Junctions and IBD
The critical role of the adherens junction in intestinal physiology was demonstrated using chimeric mice in which some, but not all, villous enterocytes expressed a dominant-negative cytoplasmic tail of N-cadherin [52]. Expression of the dominant-negative N-cadherin tail disrupted E-cadherin function and resulted in the loss of intercellular junctions, aberrant epithelial differentiation, Crohn’s-like disease, and epithelial dysplasia [52]. E-cadherin may also be relevant to human disease as data from recent genome-wide association studies (GWAS) suggest that polymorphisms within CDH1, the gene encoding E-cadherin, may be associated with ulcerative colitis [56].
Despite this critical role of the adherens junction , the tight junction is the primary determinant of paracellular permeability . It is, therefore, not surprising that alterations in tight junction structure and function are associated with increased permeability in IBD. For example, the normally complex anastomosing network of tight junction strands seen by freeze-fracture electron microscopy is simplified in IBD [26]. Moreover, the expression and distribution of individual tight junction proteins is altered in IBD [21, 22, 25]. Finally, increased expression and enzymatic activity of myosin light chain kinase, a key regulator of the perijunctional actomyosin ring and tight junction permeability, have been associated with active IBD [24]. As discussed below, this appears to be the primary intracellular signaling pathway by which TNF and related cytokines disrupt tight junction barrier function [48, 49, 57, 58].
Discovery of the claudins , a large family of proteins now recognized to define tight junction ion selectivity and to be essential for the development of the paracellular barrier, was a major breakthrough in tight junction biology [38]. The availability of reagents suitable for analyzing individual claudin proteins in human specimens revealed that expression of claudin-2 was upregulated in IBD [21, 23]. In parallel, in vitro studies demonstrated that claudin-2 expression increased the paracellular flux of cations, e.g. Na+, and small uncharged molecules [59–61]. Furthermore, in vitro and in vivo studies have shown that claudin-2 upregulation alone is responsible for the increased pore pathway flux induced by IL-13 [61]. Thus, increased flux through claudin-2 is a potential mechanism for the elevated pore pathway paracellular permeability observed in IBD. IL-13 may not be the only mediator of claudin-2 induction in IBD [62, 63], but it is interesting to note that claudin-2 expression is greater in UC than CD, and that there is a greater elevation of IL-13 production by lamina propria mononuclear cells from UC, relative to CD [22, 23]. However, as discussed below, increased claudin-2 expression cannot fully explain the barrier defects present in IBD, and it remains uncertain whether this represents a beneficial adaptive, or detrimental maladaptive, process.
While elevated levels of IL-13 are highly relevant to IBD, the impact of IL-13 neutralization on intestinal permeability and disease activity in human IBD is untested. In contrast, TNF neutralization reduces disease severity and also restores intestinal barrier function in IBD patients [64]. While this does, in part, reflect healing of mucosal ulcers and global immune downregulation , TNF also plays a critical role in tight junction regulation. For example, studies in mice have shown that acute T cell activation causes a TNF-dependent increase in intestinal permeability. This is associated with endocytosis of the tight junction protein occludin and redistribution of the cytoplasmic plaque protein ZO-1 [47, 49]. Occludin internalization is directly related to TNF-induced barrier loss, as occludin overexpression in transgenic mice limits increases in permeability and prevents diarrhea following acute TNF exposure [47]. More recent in vitro studies have shown that occludin knockdown also protects from such barrier loss as occludin deficient monolayers are insensitive to TNF exposure. This barrier regulation relies on direct interactions between occludin and ZO-1 [46, 65]. This failure of occludin-deficient epithelia to regulate tight junction permeability following TNF exposure is likely explained by the fact that these monolayers have increased tight junction leak pathway permeability at baseline [46]. Notably, occludin expression is reduced in chronic experimental colitis as well as human IBD [22, 66].
TNF-induced barrier loss has also been shown, in vitro and in vivo, to require myosin light chain kinase- (MLCK-) dependent myosin II regulatory light chain (MLC) phosphorylation [48, 49]. TNF-induced MLCK activation precedes occludin internalization , as genetic or pharmacological MLCK inhibition prevents occludin redistribution, barrier loss, and diarrhea [49]. In contrast, inhibition of occludin endocytosis prevents barrier loss but does not block MLC phosphorylation. Beyond enzymatic activation, TNF increases MLCK transcription and protein synthesis in vitro and in vivo [67, 68]. Moreover, MLCK expression and enzymatic activity are both increased in human IBD, particularly in association with active disease [24]. MLCK activation is also required for barrier dysregulation by other cytokines relevant to IBD, including lymphotoxin-like inducible protein that competes with glycoprotein D for herpes virus entry on T cells (LIGHT), and interleukin-1β (IL-1β) [58, 69, 70]. As discussed below, these and other data [28] indicate that MLCK inhibition may have therapeutic efficacy in IBD, particularly as a maintenance therapy.
The ability of pro-inflammatory cytokines to regulate tight junction barrier function may explain the presence of increased permeability in human IBD. For example, the permeability increases that precede CD reactivation may be secondary to limited immune activation. This model is also consistent with the observation that barrier defects in first-degree relatives of CD patients are associated with mutations in NOD2, a known immunoregulatory gene [20]. However, the fact that these relatives, as well as other individuals with reduced barrier function, do not have disease as well as the absence of spontaneous disease in animal models of impaired intestinal tight junction barrier function demonstrate that increased intestinal permeability alone is insufficient to cause IBD [10, 71, 72]. How is it possible to integrate these data and understand the relationship between intestinal barrier function and disease? One animal model that clarifies the relationship between immune signaling and barrier function in disease is the interleukin-10 knockout (IL-10 KO) mouse . This model is relevant to human disease, as polymorphisms of both the IL-10 promoter and the IL-10 receptor have been linked to ulcerative colitis and very eraly onset IBD [73, 74].
IL-10 knockout mice develop spontaneous colitis when housed under ‘normal’ specific pathogen-free conditions. However, like human IBD, development of disease in IL-10 KO mice is clearly multifactorial. Disease penetrance and presentation vary with genetic background and among different animal facilities, and disease does not develop at all in germ-free mice. Moreover, when disease does occur in IL-10 KO mice, clinical symptoms are not present at birth, but develop only after weeks or months [75]. Intestinal barrier defects are detectable in a large proportion of IL-10 KO mice within 4 weeks of birth, but are absent in antibiotic-treated or germ-free mice [76]. Thus, given that the primary defect in these mice is loss of the immunoregulatory cytokine IL-10 , one can conclude that intestinal permeability defects arise secondary to an interaction between the immune system and luminal microbiota. This suggests that the barrier defects in IL-10 KO mice are a sensitive indicator of mucosal immune activation. Another study suggests that enhancing mucosal barrier function by undefined mechanisms may limit disease in IL-10 KO mice [77]. Thus, early permeability defects may contribute to disease progression in IL-10 KO mice.
Another observation that suggests a contribution of barrier defects to disease evolution in IL-10 KO mice is the synchronization of disease onset by treatment with the nonsteroidal anti-inflammatory drug (NSAID) piroxicam [78]. While the mechanisms by which piroxicam exerts these effects are unclear, NSAIDs are known to cause epithelial injury, including erosions that can enhance mucosal bacterial invasion in IL-10 KO mice. Thus, one might hypothesize that piroxicam triggers disease in IL-10 KO mice by enhancing intestinal permeability, although not by increasing tight junction permeability . This suggests that a transient elevation of intestinal permeability in a genetically susceptible host, e.g. IL-10 KO mice , is sufficient to initiate chronic disease. This model could also explain the observation, made in humans, that an acute episode of bacterial gastroenteritis is associated with increased risk of developing IBD [79]. Thus, the IL-10 KO mouse model emphasizes the interaction between genetic susceptibility, immune activation, intestinal microbiota, and epithelial barrier function in development of human IBD.
Barrier Defects and Mucosal Immune Regulation
The data discussed above suggest that barrier function can be a sensitive indicator of mucosal immune activation and that increased permeability may be able to amplify the effects of such immune activation. However, while the role of primary epithelial barrier dysfunction in IBD pathogenesis remains controversial, the associations are striking. For example, barrier defects precede clinical manifestations of disease in IL-10 KO mice , in which the primary defect is immunological [76]. Increased intestinal permeability has also been reported prior to clinical disease onset in the outbred SAMP1/YitFc mouse model of IBD [80]. Bone marrow chimera studies suggest that the primary defect in these mice is present within a radioresistant , non-bone marrow-derived cell population [80]. However, the precise abnormality that causes disease in SAMP1/YitFc mice has not been identified.
Studies of mice with targeted barrier defects provide further evidence that tight junction permeability may contribute to disease progression. One informative example comes from mice lacking junctional adhesion molecule A (JAM-A) , an immunoglobulin superfamily member that facilitates tight junction assembly and leukocyte transmigration across endothelia and epithelia [72, 81]. Loss of JAM-A expression within intestinal epithelia led to increased colonic epithelial apoptosis in colitis, providing one potential mechanism by which JAM-A deficiency augments disease. Consistent with this, colonic neutrophil accumulation was increased in JAM-A KO mice , and these mice were also hypersensitive to dextran sulfate sodium- (DSS-) induced epithelial injury and colitis [72]. Because JAM-A is normally expressed ubiquitously, defects in these mice could not be linked directly to loss of intestinal epithelial JAM-A . However, endothelial specific JAM-A KO mice did not demonstrate increased DSS susceptibility, demonstrating that endothelial JAM-A loss was not responsible for the observed phenotype. In addition, the observation that intestinal epithelial, but not endothelial, JAM-A expression is reduced in human and experimental IBD supports the hypothesis that loss of intestinal epithelial JAM-A is responsible for the phenotype of JAM-A KO mice [72]. Although the phenotype of the JAM-A KO mice is informative and implicates tight junction proteins in epithelial repair processes, the cause of JAM-A loss in human and experimental IBD remains unclear. Moreover, despite the experimental simplicity of the model, the DSS model of colitis primarily reflects responses to acute epithelial damage and does not reflect the pathogenesis of human IBD. Some workers have attempted to correct this using a chronic DSS model, but that model suffers from the same problem in that it is caused by ongoing direct epithelial injury, which is unlikely to be the cause of human IBD.
Increased transcription and enzymatic activity of intestinal epithelial MLCK are well-documented in human and experimental IBD, and are regulated by proinflammatory cytokines such as TNF. Moreover, MLCK is required for in vitro and in vivo TNF-induced barrier loss [48, 49]. To model this form of barrier loss, transgenic mice that express constitutively active MLCK (CA-MLCK ) from an intestinal epithelial-specific promoter were created [71]. These mice have increased small intestinal and colonic paracellular permeability and exhibit mucosal immune activation, as evidenced by elevated mucosal IFNγ (gamma), TNF, IL-10, and IL-13 transcription and increased numbers of colonic lamina propria T cells. However, despite mucosal immune activation, the mice thrive and do not develop spontaneous disease. While this absence of disease may reflect the activity of immunoregulatory cells, such as those responsible for the increase in IL-10 transcription, expression of CA-MLCK in RAG1-deficient (RAG1−/−) mice, which lack regulatory T cells, also fails to induce disease [71]. However, when effector T lymphocytes are introduced into CA-MLCK/RAG1−/− mice, using the CD4+CD45Rbhi T lymphocyte adoptive transfer model of colitis, the onset of disease is accelerated and severity is increased relative to RAG1−/− littermates that lack CA-MLCK [71]. Therefore, these CA-MLCK transgenic mice recapitulate the observation in humans that primary barrier dysfunction is insufficient to cause disease, e.g. in healthy relatives of CD patients, and also provide evidence that barrier dysfunction can accelerate the onset and enhance the severity of immune-mediated colitis.
This interplay between TNF exposure, immune activation, MLCK, and intestinal barrier function has been further studied using immune-mediated adoptive transfer colitis in RAG1 knockout mice lacking either TNF receptor 1 (TNFR1) or TNFR2 [28]. CD4+CD45Rbhi T lymphocyte adoptive transfer into RAG1 deficient mice results in increased MLCK protein expression along with increased transcription of TNFR2 but not TNFR1 [28]. Adoptive transfer into RAG1 knockout mice lacking TNFR2 failed to induce MLCK expression, and mice lacking either TNFR2 or MLCK were at least partially protected from immune-mediated colitis [28]. These data, along with in vitro observations showing that TNF-induced, MLCK-dependent barrier loss requires TNFR2 function indicate that it is epithelial TNFR2, and not TNFR1, that drives MLCK transcription [28]. Remarkably, colitis-associated claudin-2 upregulation was also limited in MLCK-deficient mice (Fig. 3.2), demonstrating the in vivo interplay between paracellular leak and pore pathways [28]. Nevertheless, mice lacking either epithelial MLCK or TNFR2 did eventually develop colitis and unrestricted pathway barrier loss [28]. This occurred as a consequence of immune-mediated epithelial damage. Consistent with this, epithelial MLCK knockout mice were not protected from acute, TNF-induced epithelial apoptosis or DSS colitis [28]. These data reinforce the notion that MLCK inhibition may be beneficial in preventing relapse or progression to advanced IBD but may not be useful in treatment of active disease with ulcerations.

Both leak and pore paracellular pathways are altered in immune-mediated colitis. All mice are RAG1−/− to allow for adoptive transfer colitis. Myosin light chain is phosphorylated in immune-mediated colitis in an MLCK-dependent manner (top panels). Transgenic, intestinal epithelial-restricted expression of constitutively active-MLCK restores disease-associated MLC phosphorylation to long MLCK−/− mice. Claudin-2 expression parallels MLC phosphorylation in CD4+CD45RBhi colitis (bottom panels). Increases in MLC phosphorylation and claudin-2 expression correspond with worsened clinical outcomes in early stages of colitis. Scale bar = 10 μm. From Su et al., 2013
Isolated Barrier Defects Are Insufficient to Cause Disease
While data above link increased intestinal permeability to disease severity and relapse, mild barrier defects may actually trigger regulatory processes that prevent inappropriate immune responses in immunocompetent hosts. The clearest example of this may be data indicating that, despite the presence of increased permeability in patients with infectious gastroenteritis, most of these individuals recover fully and do not develop IBD [79]. Recent studies in mice have provided some insight into the immunoregulatory mechanisms that prevent development of chronic disease in this situation. In wild-type mice, when mucosal damage was induced by intrarectal ethanol administration, the resulting transient increase in permeability induced a population of regulatory T cells characterized by the surface expression of the TGFβ pro-peptide latency-associated peptide (LAP) and also increased IL-10 production by lamina propria mononuclear cells [82]. Moreover, this mild mucosal damage protected mice from colitis induced by subsequent intrarectal TNBS administration. While the detailed mechanisms by which transient increases in permeability invoke immunoregulatory responses are incompletely characterized, it is notable that mucosal IL-10 production was also increased in CA-MLCK transgenic mice [71]. Thus, chronic barrier defects may also induce immunoregulatory responses that prevent disease. On this basis, it can be inferred that, in an immunocompetent host , limited barrier defects induce a robust immunoregulatory response that prevents disease. If correct, this hypothesis suggests that the difference between CD patients and their healthy relatives with increased permeability may be the quality of this regulatory response and the ability to manage inflammation induced by barrier loss. Overall, these studies emphasize the need for further clarification of the complex interactions between the intestinal epithelial barrier and mucosal immune system.
A Multifactorial Model of IBD
The data discussed above suggest a model of IBD in which paracellular defects, mucosal immune activation, and the luminal microbiota are interrelated and act cooperatively in the prevention, induction, and progression of disease (Fig. 3.3). The epithelial barrier balances the interaction between immunostimulatory luminal materials and the mucosal immune system. In immunocompetent subjects, mild barrier defects activate regulatory immune responses that compensate for and prevent excessive immune activation. In contrast, barrier defects may trigger inappropriate immune activation and cytokine release in subjects who are unable to elicit effective immunoregulatory responses. These cytokines may then signal to epithelial cells to cause further increases in paracellular permeability, which, in turn, allows further transmucosal passage of immunostimulatory materials and an even greater degree of immune activation. Thus defects in either barrier function or immune activation may initiate the cycle of barrier-immune dysregulation, but defects in both are necessary for perpetuation of this cycle and disease pathogenesis. This model also suggests that, in addition to the immunosuppressive and immunomodulatory agents that have been central to IBD therapy, it may be possible to break the cycle by restoring barrier function prior to onset of clinical disease.

The epithelial barrier regulates the balance between the luminal environment and mucosal immunity . Breaches in the epithelial barrier allow luminal material to cross the epithelium, where it may then activate mucosal immune responses. Under conditions of appropriate immune regulation, regulatory T cells (Tregs) produce anti-inflammatory signals to counter the pro-inflammatory immune response. A subset of these Tregs, those expressing latency-associated peptide (LAP), secrete IL-10 and TGF-β to attenuate disease. In contrast, under disease conditions, luminal material may activate Th1 or Th2 T cells. Th1 cells secrete TNF and IFNγ, which increase paracellular leak pathway permeability via MLCK-dependent mechanisms. Alternatively, Th2 cells secrete IL-13, which increases pore pathway permeability by upregulating claudin-2 expression. These increases in paracellular permeability may allow the passage of additional luminal material across the barrier. Amplification of this cycle of inflammation eventually leads to apoptosis of epithelial cells. Under these conditions, intestinal permeability is largely tight-junction independent and unrestricted. Due to the tight junction-independent nature of this unrestricted pathway in more advanced disease, therapeutics designed to restore tight junction barrier function will likely be most useful in early disease or in maintaining remission. Adapted from Su et al. 2013
Barrier Restoration as a Future Therapeutic Intervention
Current immunosuppressive and immunomodulatory approaches are often effective in IBD management, but they treat the inflammation responsible for tissue damage rather than the underlying cause of inflammation. It has been recognized for some time that the epithelial barrier can be dysregulated by proinflammatory stimuli. The model described, which explains available data from in vitro and in vivo models as well as human studies, suggests that the barrier also regulates the immune response. Although increased paracellular permeability is a normal physiological response that is often beneficial and, in an immunocompetent host, triggers an immunoregulatory response, reduced barrier function may also be critical to disease pathogenesis. This is particularly true in individuals with immunoregulatory defects. For example, barrier loss precedes colitis in IL-10-deficient mice. Conversely, barrier restoration may blunt disease development. Similarly, genetic deletion of intestinal epithelial long MLCK, which is necessary for TNF-induced barrier loss, partially protects immunodeficient mice from colitis induced by CD4+CD45Rbhi adoptive transfer. These data suggest that barrier restoration may be therapeutically useful, particularly in patients with IL-10 receptor mutations or single nucleotide polymorphisms (SNPs) within the IL10 promoter.
Development of barrier restorative agents will require targeting the pore, leak, and unrestricted paracellular pathways, all of which are disrupted at different stages of colitis. While claudin-2 must be responsible for some of the increases in pore pathway permeability in colitis, no modulators of claudin-2 pore function are currently available.
Epithelial MLCK is clearly an attractive druggable target to restore leak pathway permeability early during disease development and promote maintenance of remission. However, the different MLCK isoforms expressed in smooth muscle and nonmuscle cells are encoded by a single gene and, therefore, have common catalytic and calmodulin-dependent regulatory domains [83, 84]. Knockout mouse studies clearly show that inhibition of smooth muscle MLCK has disastrous consequences [84]. Thus, alternatives to currently available MLCK inhibitors , all of which target the catalytic domain, will be necessary if MLCK is to be a viable therapeutic target.
Tight junctions play a lesser role in intestinal barrier loss in more advanced disease when increased permeability is primarily driven by epithelial damage and increased flux across the unrestricted pathway. Some of the epithelial wounds present are repaired by an actomyosin purse string-dependent mechanism that also requires epithelial MLCK activity [85]. Therefore, in order restore tight junction barrier function without disrupting other tissues or epithelial wound repair, it will be critical to develop a more targeted approach to inhibiting tight junction regulatory activities of epithelial MLCK in response to pathophysiologic stimuli, such as TNF, without impairing wound healing. If this can be achieved, barrier restoration may have a promising future in IBD therapy.
References
Turner JR, Rill BK, Carlson SL, Carnes D, Kerner R, Mrsny RJ, et al. Physiological regulation of epithelial tight junctions is associated with myosin light-chain phosphorylation. Am J Physiol. 1997;273:C1378–85.
Shiue H, Musch MW, Wang Y, Chang EB, Turner JR. Akt2 phosphorylates ezrin to trigger NHE3 translocation and activation. J Biol Chem. 2005;280:1688–95.
Turner JR, Black ED, Ward J, Tse CM, Uchwat FA, Alli HA, et al. Transepithelial resistance can be regulated by the intestinal brush border Na+-H+ exchanger NHE3. Am J Physiol Cell Physiol. 2000;279:C1918–24.
Turner JR, Cohen DE, Mrsny RJ, Madara JL. Noninvasive in vivo analysis of human small intestinal paracellular absorption: regulation by Na + -glucose cotransport. Dig Dis Sci. 2000;45:2122–6.
Ukabam SO, Clamp JR, Cooper BT. Abnormal small intestinal permeability to sugars in patients with Crohn’s disease of the terminal ileum and colon. Digestion. 1983;27:70–4.
Hollander D. Crohn’s disease—a permeability disorder of the tight junction? Gut. 1988;29:1621–4.
Pearson AD, Eastham EJ, Laker MF, Craft AW, Nelson R. Intestinal permeability in children with Crohn’s disease and coeliac disease. Br Med J (Clin Res Ed). 1982;285:20–1.
Schulzke JD, Bentzel CJ, Schulzke I, Riecken EO, Fromm M. Epithelial tight junction structure in the jejunum of children with acute and treated celiac sprue. Pediatr Res. 1998;43:435–41.
Madara JL, Trier JS. Structural abnormalities of jejunal epithelial cell membranes in celiac sprue. Lab Invest. 1980;43:254–61.
Hollander D, Vadheim CM, Brettholz E, Petersen GM, Delahunty T, Rotter JI. Increased intestinal permeability in patients with Crohn’s disease and their relatives. A possible etiologic factor. Ann Intern Med. 1986;105:883–5.
Jenkins RT, Ramage JK, Jones DB, Collins SM, Goodacre RL, Hunt RH. Small bowel and colonic permeability to 51Cr-EDTA in patients with active inflammatory bowel disease. Clin Invest Med. 1988;11:151–5.
Bijlsma PB, Peeters RA, Groot JA, Dekker PR, Taminiau JA, Van Der Meer R. Differential in vivo and in vitro intestinal permeability to lactulose and mannitol in animals and humans: a hypothesis. Gastroenterology. 1995;108:687–96.
Arslan G, Atasever T, Cindoruk M, Yildirim IS. (51)CrEDTA colonic permeability and therapy response in patients with ulcerative colitis. Nucl Med Commun. 2001;22:997–1001.
Bjarnason I. Intestinal permeability. Gut. 1994;35:S18–22.
Keighley MR, Taylor EW, Hares MM, Arabi Y, Youngs D, Bentley S, et al. Influence of oral mannitol bowel preparation on colonic microflora and the risk of explosion during endoscopic diathermy. Br J Surg. 1981;68:554–6.
Vince A, Killingley M, Wrong OM. Effect of lactulose on ammonia production in a fecal incubation system. Gastroenterology. 1978;74:544–9.
Katz KD, Hollander D, Vadheim CM, McElree C, Delahunty T, Dadufalza VD, et al. Intestinal permeability in patients with Crohn’s disease and their healthy relatives. Gastroenterology. 1989;97:927–31.
May GR, Sutherland LR, Meddings JB. Is small intestinal permeability really increased in relatives of patients with Crohn’s disease? Gastroenterology. 1993;104:1627–32.
Hilsden RJ, Meddings JB, Sutherland LR. Intestinal permeability changes in response to acetylsalicylic acid in relatives of patients with Crohn’s disease. Gastroenterology. 1996;110:1395–403.
Buhner S, Buning C, Genschel J, Kling K, Herrmann D, Dignass A, et al. Genetic basis for increased intestinal permeability in families with Crohn’s disease: role of CARD15 3020insC mutation? Gut. 2006;55:342–7.
Prasad S, Mingrino R, Kaukinen K, Hayes KL, Powell RM, MacDonald TT, et al. Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells. Lab Invest. 2005;85:1139–62.
Heller F, Florian P, Bojarski C, Richter J, Christ M, Hillenbrand B, et al. Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology. 2005;129:550–64.
Zeissig S, Burgel N, Gunzel D, Richter J, Mankertz J, Wahnschaffe U, et al. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut. 2007;56:61–72.
Blair SA, Kane SV, Clayburgh DR, Turner JR. Epithelial myosin light chain kinase expression and activity are upregulated in inflammatory bowel disease. Lab Invest. 2006;86:191–201.
Weber CR, Nalle SC, Tretiakova M, Rubin DT, Turner JR. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab Invest. 2008;88:1110–20.
Schmitz H, Barmeyer C, Fromm M, Runkel N, Foss HD, Bentzel CJ, et al. Altered tight junction structure contributes to the impaired epithelial barrier function in ulcerative colitis. Gastroenterology. 1999;116:301–9.
Zeissig S, Bojarski C, Buergel N, Mankertz J, Zeitz M, Fromm M, et al. Downregulation of epithelial apoptosis and barrier repair in active Crohn’s disease by tumour necrosis factor alpha antibody treatment. Gut. 2004;53:1295–302.
Su L, Nalle SC, Shen L, Turner ES, Singh G, Breskin LA, et al. TNFR2 activates MLCK-dependent tight junction dysregulation to cause apoptosis-mediated barrier loss and experimental colitis. Gastroenterology. 2013;145:407–15.
Irvine EJ, Marshall JK. Increased intestinal permeability precedes the onset of Crohn’s disease in a subject with familial risk. Gastroenterology. 2000;119:1740–4.
Sands BE. Inflammatory bowel disease: past, present, and future. J Gastroenterol. 2007;42:16–25.
Wyatt J, Vogelsang H, Hubl W, Waldhoer T, Lochs H. Intestinal permeability and the prediction of relapse in Crohn’s disease. Lancet. 1993;341:1437–9.
Farquhar M, Palade G. Junctional complexes in various epithelia. J Cell Biol. 1963;17:375–412.
Ojakian GK, Nelson WJ, Beck KA. Mechanisms for de novo biogenesis of an apical membrane compartment in groups of simple epithelial cells surrounded by extracellular matrix. J Cell Sci. 1997;110:2781–94.
Nejsum LN, Nelson WJ. A molecular mechanism directly linking E-cadherin adhesion to initiation of epithelial cell surface polarity. J Cell Biol. 2007;178:323–35.
Buckley CD, Tan J, Anderson KL, Hanein D, Volkmann N, Weis WI, et al. Cell adhesion. The minimal cadherin-catenin complex binds to actin filaments under force. Science. 2014;346:1254211.
Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita S, et al. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123:1777–88.
McCarthy KM, Skare IB, Stankewich MC, Furuse M, Tsukita S, Rogers RA, et al. Occludin is a functional component of the tight junction. J Cell Sci. 1996;109(Pt 9):2287–98.
Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–50.
Furuse M, Sasaki H, Fujimoto K, Tsukita S. A single gene product, claudin-1 or -2, reconstitutes tight junction strands and recruits occludin in fibroblasts. J Cell Biol. 1998;143:391–401.
Fanning AS, Anderson JM. Zonula occludens-1 and -2 are cytosolic scaffolds that regulate the assembly of cellular junctions. Ann N Y Acad Sci. 2009;1165:113–20.
Furuse M, Hata M, Furuse K, Yoshida Y, Haratake A, Sugitani Y, et al. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: a lesson from claudin-1-deficient mice. J Cell Biol. 2002;156:1099–111.
Umeda K, Ikenouchi J, Katahira-Tayama S, Furuse K, Sasaki H, Nakayama M, et al. ZO-1 and ZO-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell. 2006;126:741–54.
Ikenouchi J, Umeda K, Tsukita S, Furuse M. Requirement of ZO-1 for the formation of belt-like adherens junctions during epithelial cell polarization. J Cell Biol. 2007;176:779–86.
Van Itallie CM, Anderson JM. Claudins and epithelial paracellular transport. Annu Rev Physiol. 2006;68:403–29.
Suzuki H, Nishizawa T, Tani K, Yamazaki Y, Tamura A, Ishitani R, et al. Crystal structure of a claudin provides insight into the architecture of tight junctions. Science. 2014;344:304–7.
Buschmann MM, Shen L, Rajapakse H, Raleigh DR, Wang Y, Wang Y, et al. Occludin OCEL-domain interactions are required for maintenance and regulation of the tight junction barrier to macromolecular flux. Mol Biol Cell. 2013;24:3056–68.
Marchiando AM, Shen L, Graham WV, Weber CR, Schwarz BT, Austin 2nd JR, et al. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J Cell Biol. 2010;189:111–26.
Zolotarevsky Y, Hecht G, Koutsouris A, Gonzalez DE, Quan C, Tom J, et al. A membrane-permeant peptide that inhibits MLC kinase restores barrier function in in vitro models of intestinal disease. Gastroenterology. 2002;123:163–72.
Clayburgh DR, Barrett TA, Tang Y, Meddings JB, Van Eldik LJ, Watterson DM, et al. Epithelial myosin light chain kinase-dependent barrier dysfunction mediates T cell activation-induced diarrhea in vivo. J Clin Invest. 2005;115:2702–15.
Raleigh DR, Boe DM, Yu D, Weber CR, Marchiando AM, Bradford EM, et al. Occludin S408 phosphorylation regulates tight junction protein interactions and barrier function. J Cell Biol. 2011;193:565–82.
Van Itallie CM, Fanning AS, Bridges A, Anderson JM. ZO-1 stabilizes the tight junction solute barrier through coupling to the perijunctional cytoskeleton. Mol Biol Cell. 2009;20:3930–40.
Hermiston ML, Gordon JI. Inflammatory bowel disease and adenomas in mice expressing a dominant negative N-cadherin. Science. 1995;270:1203–7.
Hermiston ML, Gordon JI. In vivo analysis of cadherin function in the mouse intestinal epithelium: essential roles in adhesion, maintenance of differentiation, and regulation of programmed cell death. J Cell Biol. 1995;129:489–506.
Capaldo CT, Macara IG. Depletion of E-cadherin disrupts establishment but not maintenance of cell junctions in Madin-Darby canine kidney epithelial cells. Mol Biol Cell. 2007;18:189–200.
Maiers JL, Peng X, Fanning AS, DeMali KA. ZO-1 recruitment to alpha-catenin—a novel mechanism for coupling the assembly of tight junctions to adherens junctions. J Cell Sci. 2013;126:3904–15.
Barrett JC, Hansoul S, Nicolae DL, Cho JH, Duerr RH, Rioux JD, et al. Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat Genet. 2008;40:955–62.
Al-Sadi R, Guo S, Ye D, Dokladny K, Alhmoud T, Ereifej L, et al. Mechanism of IL-1beta modulation of intestinal epithelial barrier involves p38 kinase and activating transcription factor-2 activation. J Immunol. 2013;190:6596–606.
Schwarz BT, Wang F, Shen L, Clayburgh DR, Su L, Wang Y, et al. LIGHT signals directly to intestinal epithelia to cause barrier dysfunction via cytoskeletal and endocytic mechanisms. Gastroenterology. 2007;132:2383–94.
Furuse M, Furuse K, Sasaki H, Tsukita S. Conversion of zonulae occludentes from tight to leaky strand type by introducing claudin-2 into Madin-Darby canine kidney I cells. J Cell Biol. 2001;153:263–72.
Amasheh S, Meiri N, Gitter AH, Schoneberg T, Mankertz J, Schulzke JD, et al. Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells. J Cell Sci. 2002;115:4969–76.
Weber CR, Raleigh DR, Su L, Shen L, Sullivan EA, Wang Y, et al. Epithelial myosin light chain kinase activation induces mucosal interleukin-13 expression to alter tight junction ion selectivity. J Biol Chem. 2010;285:12037–46.
Suzuki T, Yoshinaga N, Tanabe S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J Biol Chem. 2011;286:31263–71.
Mankertz J, Amasheh M, Krug SM, Fromm A, Amasheh S, Hillenbrand B, et al. TNFalpha up-regulates claudin-2 expression in epithelial HT-29/B6 cells via phosphatidylinositol-3-kinase signaling. Cell Tissue Res. 2009;336:67–77.
Suenaert P, Bulteel V, Lemmens L, Noman M, Geypens B, Van Assche G, et al. Anti-tumor necrosis factor treatment restores the gut barrier in Crohn’s disease. Am J Gastroenterol. 2002;97: 2000–4.
Van Itallie CM, Fanning AS, Holmes J, Anderson JM. Occludin is required for cytokine-induced regulation of tight junction barriers. J Cell Sci. 2010;123:2844–52.
Wang F, Schwarz BT, Graham WV, Wang Y, Su L, Clayburgh DR, et al. IFN-gamma-induced TNFR2 expression is required for TNF-dependent intestinal epithelial barrier dysfunction. Gastroenterology. 2006;131:1153–63.
Wang F, Graham WV, Wang Y, Witkowski ED, Schwarz BT, Turner JR. Interferon-gamma and tumor necrosis factor-alpha synergize to induce intestinal epithelial barrier dysfunction by up-regulating myosin light chain kinase expression. Am J Pathol. 2005;166:409–19.
Graham WV, Wang F, Clayburgh DR, Cheng JX, Yoon B, Wang Y, et al. Tumor necrosis factor-induced long myosin light chain kinase transcription is regulated by differentiation-dependent signaling events. Characterization of the human long myosin light chain kinase promoter. J Biol Chem. 2006;281:26205–15.
Al-Sadi R, Ye D, Dokladny K, Ma TY. Mechanism of IL-1beta-induced increase in intestinal epithelial tight junction permeability. J Immunol. 2008;180:5653–61.
Al-Sadi R, Ye D, Said HM, Ma TY. IL-1beta-induced increase in intestinal epithelial tight junction permeability is mediated by MEKK-1 activation of canonical NF-kappaB pathway. Am J Pathol. 2010;177:2310–22.
Su L, Shen L, Clayburgh DR, Nalle SC, Sullivan EA, Meddings JB, et al. Targeted epithelial tight junction dysfunction causes immune activation and contributes to development of experimental colitis. Gastroenterology. 2009;136:551–63.
Vetrano S, Rescigno M, Rosaria Cera M, Correale C, Rumio C, Doni A, et al. Unique role of junctional adhesion molecule-a in maintaining mucosal homeostasis in inflammatory bowel disease. Gastroenterology. 2008;135:173–84.
Tedde A, Laura Putignano A, Bagnoli S, Congregati C, Milla M, Sorbi S, et al. Interleukin-10 promoter polymorphisms influence susceptibility to ulcerative colitis in a gender-specific manner. Scand J Gastroenterol. 2008;43:712–8.
Glocker EO, Kotlarz D, Boztug K, Gertz EM, Schaffer AA, Noyan F, et al. Inflammatory bowel disease and mutations affecting the interleukin-10 receptor. N Engl J Med. 2009;361:2033–45.
Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. 1993;75: 263–74.
Madsen KL, Malfair D, Gray D, Doyle JS, Jewell LD, Fedorak RN. Interleukin-10 gene-deficient mice develop a primary intestinal permeability defect in response to enteric microflora. Inflamm Bowel Dis. 1999;5:262–70.
Arrieta MC, Madsen K, Doyle J, Meddings J. Reducing small intestinal permeability attenuates colitis in the IL10 gene-deficient mouse. Gut. 2009;58:41–8.
Narushima S, Spitz DR, Oberley LW, Toyokuni S, Miyata T, Gunnett CA, et al. Evidence for oxidative stress in NSAID-induced colitis in IL10−/− mice. Free Radic Biol Med. 2003;34:1153–66.
Gradel KO, Nielsen HL, Schonheyder HC, Ejlertsen T, Kristensen B, Nielsen H. Increased short- and long-term risk of inflammatory bowel disease after salmonella or campylobacter gastroenteritis. Gastroenterology. 2009;137:495–501.
Olson TS, Reuter BK, Scott KG, Morris MA, Wang XM, Hancock LN, et al. The primary defect in experimental ileitis originates from a nonhematopoietic source. J Exp Med. 2006;203:541–52.
Woodfin A, Reichel CA, Khandoga A, Corada M, Voisin MB, Scheiermann C, et al. JAM-A mediates neutrophil transmigration in a stimulus-specific manner in vivo: evidence for sequential roles for JAM-A and PECAM-1 in neutrophil transmigration. Blood. 2007;110:1848–56.
Boirivant M, Amendola A, Butera A, Sanchez M, Xu L, Marinaro M, et al. A transient breach in the epithelial barrier leads to regulatory T-cell generation and resistance to experimental colitis. Gastroenterology. 2008;135:1612–23.
Clayburgh DR, Rosen S, Witkowski ED, Wang F, Blair S, Dudek S, et al. A differentiation-dependent splice variant of myosin light chain kinase, MLCK1, regulates epithelial tight junction permeability. J Biol Chem. 2004;279:55506–13.
He WQ, Peng YJ, Zhang WC, Lv N, Tang J, Chen C, et al. Myosin light chain kinase is central to smooth muscle contraction and required for gastrointestinal motility in mice. Gastroenterology. 2008;135:610–20.
Russo JM, Florian P, Shen L, Graham WV, Tretiakova MS, Gitter AH, et al. Distinct temporal-spatial roles for rho kinase and myosin light chain kinase in epithelial purse-string wound closure. Gastroenterology. 2005;128:987–1001.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Odenwald, M.A., Turner, J.R. (2017). Understanding the Epithelial Barrier in IBD. In: Baumgart, D. (eds) Crohn's Disease and Ulcerative Colitis. Springer, Cham. https://doi.org/10.1007/978-3-319-33703-6_3
Download citation
DOI: https://doi.org/10.1007/978-3-319-33703-6_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-33701-2
Online ISBN: 978-3-319-33703-6
eBook Packages: MedicineMedicine (R0)