
Abstract
The immune system plays a key role in the development, establishment, and progression of head and neck cancer. A greater understanding of the dysregulation and evasion of the immune system in the evolution and progression of head and neck cancers provides the basis for improved therapies and outcomes for patients. Head and neck cancer evades the host’s immune system on different levels: (1) manipulation of its own immunogenicity, (2) production of immunosuppressive molecules, and (3) promotion of immunomodulatory cell types. Through the tumor’s influence on the microenvironment, the immune system can be exploited to promote metastasis, angiogenesis, and growth. In this chapter, we review basic immunology as it relates to head and neck cancer and discuss the theory of cancer immunosurveillance and immune escape. A brief overview to key components in the tumor microenvironment is provided. Current research on cytokines as biomarkers, cancer stem cells, tumor antigens, and immunotherapeutic strategies is presented.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Immunology
- Immunotherapy
- Head and neck cancer
- Biomarkers
- Immune evasion
- Immune surveillance
- Monoclonal antibodies
6.1 Introduction
The immune system plays a key role in the development, establishment, and progression of head and neck cancer. A greater understanding of the important contribution of the dysregulation and evasion of the immune system in the development and evolution of head and neck cancers should lead to improved therapies and outcomes for patients. Head and neck cancer evades the host’s immune system on different levels: (1) manipulation of its own immunogenicity, (2) production of immunosuppressive molecules, and (3) promotion of immunomodulatory cell types. Through the tumor’s influence on the microenvironment, the immune system can be exploited to promote metastasis, angiogenesis, and growth. In this chapter, we review basic immunology as it relates to head and neck cancer and discuss the theory of cancer immunosurveillance and immune escape.
There has been a recent renaissance in the idea that nascent cancer cells are destroyed by the immune system before tumor formation can occur (termed immune surveillance). Derangements in the immune system or alterations in the transformed cells may allow immune escape that enables the cancer to become manifest. In the progress, the tumor interacts with the immune system in manifold ways: transcription factors such as NFκB (nuclear factor kappa-light-chain-enhancer of activated B cells) and STAT3 (signal transducers and activators of transcription), which are usually deregulated in tumor-promoting inflammatory states in response to cytokine stimuli, are aberrantly activated in tumor cells and are intensively studied as possible targets for therapeutic intervention. Tumors themselves produce cytokines such as transforming growth factor-beta (TGF-β), interleukin (IL)-6, and IL-10, which suppress cell-mediated antitumor immunity. In response to inflammatory stimuli, head and neck cancer cells also can express ligands which are involved in lymphocyte and dendritic cell migration. Expression of these ligands by tumor cells, such as CCR7 and CXCR4, constitutes immune exploitation of established signals intended for immune cells and has been associated with tumor invasion, metastasis, and cell survival, leading to treatment resistance. Another recently espoused theory is the idea that tumors are comprised of a heterogeneous cell population in the tumor microenvironment that includes a special subpopulation of cancer stem cells (CSC) that are able to recreate the entire tumor phenotype and potentially evade immune recognition. These cells appear to be more resistant to conventional chemotherapy and radiation and may not possess the same tumor antigen expression or T cell recognition as non-CSC.
In head and neck cancer patients, there appear to be global alterations in the functional state of the immune system, as evidenced by changes in serum cytokines, chemokines, and other immune-related biomarkers. We discuss investigations on the identification of serum biomarkers to monitor cancer progression, prognosis, treatment response, and relapse. Finally various immunotherapeutic strategies designed to utilize the immune system to stimulate elimination of cancer are described. These include cancer vaccines using tumor peptide antigens or viral, bacterial, and DNA-based vectors—as well as tumor antigen-specific monoclonal antibodies (mAb). The recent clinical efficacy of these FDA-approved mAb, including cetuximab (anti-EGFR) and bevacizumab (anti-VEGF), has stimulated research of immunological mechanisms that enhance antitumor activity.
6.2 Brief Overview of the Immune System
The immune system has traditionally been divided into two major arms: innate and adaptive immunity. This dichotomy is somewhat artificial since there is tremendous interaction between the two components. Innate immunity refers to the part of the immune system that provides antigen nonspecific, first-line protection. The effectors of innate immunity include natural killer (NK) cells, neutrophils, macrophages, dendritic cells (DC), and monocytes that attack/ingest extracellular debris or pathogens. Innate immunity also utilizes pattern recognition systems that recognize molecules that are not normally present in the human body: double-stranded RNA, bacterial cell wall components, lipopolysaccharides, and microbial membranes. These pattern recognition systems can be represented by enzymes like lysozyme, antimicrobial peptides (defensins), soluble factors (complement, C-reactive protein, mannose-binding lectin), and cell surface receptors (Toll-like receptors, scavenger receptors). Innate immunity is more static and nonspecific, showing less adaptation in magnitude or efficacy after repeated exposure to antigenic challenges. However, innate immune signals effectively trigger the adaptive immune system. DC and other antigen-presenting cells link the two systems. They ingest and process tumor antigens, after effectors of innate immunity have destroyed the tumor cell. DC then present these antigens to cytolytic and helper T lymphocytes, causing clonal expansion of antigen-specific T cells. Activation of the adaptive immune system (T lymphocytes) provides immunologic memory responses against these antigens. Thus, key effectors in tumor immunology are B cells, T cells, NK cells, and DC.
6.3 B Lymphocytes
Early in the field of immunology, humoral immunity was believed to be the primary effector mechanism; in 1948 plasma cells were identified as the source of antibodies. Plasma cells are one of the two endpoints for B cells, the other being the memory B cell. B cells can be activated via T cell-dependent or T cell-independent antigens. Tumor antigens are T cell-dependent antigens which require binding of the antigen to the B cell receptor and a secondary activation signal via CD40 on an activated helper T cell. It is well established that B cells in cancer patients are capable of recognizing and producing antibodies to tumor antigens [1, 2]. In head and neck cancer, circulating serum antibodies have been found against p53 [3], MUC1 [4], p40 [5], p73 [6], and HPV E6 and E7 [7]. However, levels of circulating antibody have not been correlated with clinical outcome other than high postoperative levels of anti-p53 antibody which have been correlated with poor prognosis [8]. Interestingly, it has been noted that there is an increased frequency of immunoglobulin (Ig) E subtype in head and neck cancer [2, 9]. The significance of this finding, if any, is unclear.
6.4 T Lymphocytes
T lymphocytes were defined in the early 1960s when mice were thymectomized in an attempt to prevent lymphoma. When the initial experiments in adult mice failed to have any effect, neonatal thymectomized mice were found to have profoundly decreased lymphocyte numbers and were unable to generate antibodies despite having plasma cells. Based on these data, Miller theorized that the thymus must be the source of a “helper” cell that is required to produce antibody [10–12]. In later experiments, CD8 T cells were identified as a primary effector of specific tumor/allograft rejection.
T lymphocytes are defined by the presence of T cell receptors (TCR) on their cell surface. TCR are part of the immunoglobulin superfamily and undergo germline DNA rearrangement to produce diversity much like immunoglobulin genes in B cells. TCR recognize tumor antigens which are short peptide fragments bound to or “presented by” major histocompatibility complexes (MHC). There are two main classes of MHC: MHC I molecules, found on the cell surface of all nucleated cells, and MHC II, found only on professional antigen-presenting cells such as macrophages and dendritic cells. MHC class I and II bind with peptides, which are derived from (tumor) proteins and “processed” within the cell. Hereafter, MHC molecules present these (tumor) peptides on the cell surface for recognition by T cells. The TCR can only recognize peptide antigens when presented by a particular self-MHC molecule, a phenomenon known as MHC restriction, which led to the Nobel Prize in 1996 to Doherty and Zinkernagel. Therefore, CD8 T cells can recognize syngeneic (self) but not allogeneic (from someone else) tumor cells. MHC I binding tumor peptides are usually 8–10 amino acids in length, are derived from endogenous proteins processed via the proteasome, and are presented to CD8 T cells [13]. MHC II peptides are longer (11–16 amino acids), are derived from exogenous proteins taken in by endocytosis, and are presented to CD4 T cells.
T lymphocytes are generally divided into CD4+ or CD8+ T cells. While it remains unclear how T cells are selected to become CD4 or CD8 cells, there are usually twice as many CD4 T cells as CD8 T cells released. Once antigen is encountered along with the appropriate costimulatory signals, T cells become activated and differentiated. CD4 T helper (TH) cells usually differentiate into one of two major subclasses, TH1 and TH2. This differentiation depends on the cytokine milieu in the environment at the time of activation. These two subsets of CD4 cells are differentiated by function and cytokine secretion profile. The TH1 subset is responsible for most cell-mediated immune functions such as activation of CD8 T cells, inflammation and delayed-type hypersensitivity, as well as production of complement-activating IgG antibodies. Macrophages or dendritic cells will produce IL-12 in response to intracellular pathogens. IL-12 along with interferon-gamma (IFN-γ) and IL-18 drive the TH1 response. TH1 cells secrete IL-2, IFN-γ, and tumor necrosis factor (TNF)-α and are considered to be the strongest antitumor subtype.
On the other hand, IL-4 drives a TH2 response [14]. The TH2 response stimulates B cells to produce IgM, IgE, and non-complement-activating IgG, as well as activating eosinophils, in response to parasitic invasion. TH2 T cells are strongly implicated in allergy and are considered to be tumor permissive. TH2 cells secrete granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-3, IL-4, IL-5, IL-10, and IL-13. More recently, other subsets of CD4 T cells have been identified. TH17 cells require TGF-β and IL-6 for differentiation and are defined by their production of IL-17. IL-17 is known to induce the production of several chemokines that attract proinflammatory cells, and IL-17 expression is greatly increased in autoimmune diseases [15]. The final subset of CD4 T cells is the regulatory T cell (Treg) that was originally defined as a CD4+CD25+Foxp3 (Forkhead box P3)+ T cell. Tregs are thought to be a reciprocal subtype to TH17 cells in that both are induced by TGF-β, but Tregs are immunosuppressive as opposed to TH17 cells which are proinflammatory. Tregs have recently been strongly correlated with disease status in SCCHN patients [16, 17].
6.5 Natural Killer Cells
NK cells were discovered in 1975 when experiments studying tumor lysis by lymphocytes from immunized animals found lysis that was independent of previous immunization or activation [18]. This was thought to be an artifact until the NK cell was isolated and given the name “natural killer” cell for its ability to kill tumors without previous activation. NK cells kill much in the same way as cytotoxic T cells, through the interaction Fas ligand on their surface with Fas on target cells inducing apoptotic cell death. They also constitutively possess perforin and granzyme granules and degranulate causing cytolysis. Unlike T cells that are self-MHC restricted and require self-MHC for activation, NK cells can be suppressed by the presence of self-MHC via killer cell immunoglobulin-like receptors (KIR) that can inhibit NK killing when bound by self-MHC [19]. These inhibitory signals dominate functional responses as to activating receptors, and therefore, presentation of self-MHC on the target’s surface is protective. Activation receptors on the NK cell include NKD2D and FcγIII receptor. NKD2D binds ligands produced by cells stressed by DNA damage or infection. FcγIII receptor is a high affinity receptor for IgG which provides a mechanism by which NK cells can recognize targets bound by antibody. Activating Fcγ receptors mediate antibody-dependent cell-mediated cytotoxicity (ADCC) by NK cells, macrophages, monocytes, neutrophils, and eosinophils.
6.6 Dendritic Cells
DC are antigen-presenting cells and as such are potent initiators of the immune response. They use several mechanisms for efficient antigen uptake, including phagocytosis, macropinocytosis, and adsorptive endocytosis. After uptake, antigen is shunted into lysosomes and degraded for presentation on MHC II. DC also possess B7 molecules on their surface that provide a necessary secondary activation signal to T cells after engagement of the MHC-peptide complex with the TCR. Because DC are such potent activators of T cells and initiators of adaptive immunity, they have been intensely studied as a possible therapeutic for cancer immunotherapy.
Another important process mediated by DC is cross-presentation of antigen derived from tumor cells or shed tumor products/vesicles. Exogenous antigen is processed via the exogenous pathway and presented to CD4 cells by DC via MHC II. However, DC are able to move exogenous antigen to the endogenous pathway and present these antigen to CD8 cells via MHC I. This surrogate presentation of exogenous antigen to the endogenous pathway is defined as cross-presentation. Cross-presentation serves a very important function because it allows DC to activate cytotoxic T cells against virally infected cells and tumor cells and has recently been harnessed in cancer vaccine trials.
6.7 Cancer Immunosurveillance and Immunoediting
The idea of immune control of malignant cells was first proposed by Paul Ehrlich in 1908, but it was not until the 1950s that greater understanding of the immune system gave rise to a formalized hypothesis. This “cancer immunosurveillance” hypothesis was introduced by Burnet and Thomas and stated that tumor cells must have recognizably different antigens than normal cells and therefore have the potential for immune clearance. Also at that time, the phenomenon of allograft rejection via cellular immunity was observed. Because grafting of allogeneic tissue is not a naturally occurring event, Thomas proposed that the actual primary function of cellular immunity was not to protect against allografts but rather to protect against tumors. Conflicting experimental results led many to abandon the idea of cancer immunosurveillance for several decades, until several key discoveries have led to a revival of the hypothesis. First was the discovery of the NK cell in the late 1970s which seemed to provide innate immune protection from tumor [20]. The discovery of IFN-γ and its proapoptotic effect on tumor growth gave additional support to the potential for immune clearance of cancer cells [21]. Mice lacking IFN-γ receptors produced more tumors with decreased latency after methylcholanthrene challenge, and addition of IFN-γ was protective against transplanted, spontaneous, and induced tumors in another experiment. Studies in mice lacking perforin, a key component of cytolytic granules in T cells and NK cells, recapitulated the results in IFN-γ receptor knockout mice with more frequent tumors and lower latency of formation [22]. Mice with genetically induced immunodeficiency were found to be more susceptible to both spontaneous and chemically induced tumors. In humans, epidemiologic data from AIDS patients demonstrate increased risk of lymphoma, Kaposi’s sarcoma, and virally induced carcinomas of the genitourinary tract. There also appears to be a higher risk of HPV-associated HNC in HIV+ patients [23]. These data confirm the unchallenged idea that immune protection from viral infections reduces risks of cancer associated with viruses.
But what about tumors without viral etiology? Data gathered from transplant patients who are immunosuppressed to avoid organ rejection demonstrate increased risk of many tumors with no known viral etiology such as lung, head and neck [24], pancreatic, endocrine, and colon cancer and melanoma [25]. The cancer immunosurveillance hypothesis has given rise to the theory of cancer immunoediting which represents the idea that immune surveillance of cancers provides selective pressure on tumor cells and negatively selects for cells that can evade the immune system. One study showed that many tumors grown in immunocompromised mice are rapidly cleared when injected into immunocompetent mice, whereas cancers from immunocompetent mice continue to grow when transplanted into other immunocompetent mice, indicating a qualitative difference in the cancer cells that was dependent on the immune environment [26]. The theory contends that successful tumor formation can occur only after the cancer has discovered a means by which it can evade the immune system.
6.8 Immune Escape and Immunosuppression in Head and Neck Cancer
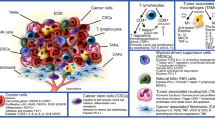
In order to establish effective immunotherapies, it is of utmost importance to understand the different pathways of the tumor’s scopes for immunoevasion. Two primary evasion means are distinguished: firstly, the cancer cells’ ability to reduce their innate immunogenicity and, secondly, their suppression of the immune response (Fig. 6.1). To date, various mechanisms through which the tumor cells achieve this immunoevasion have been identified, partly depending on the tumor’s characteristics (e.g., site, differentiation, and host). A key component for the immune system’s recognition of different or altered cells is the human leukocyte antigen (HLA) system. The HLA system is the human display of the MHC classes. As previously described, T cells interact with the MHC/HLA molecules. Tumor cells can reduce T cell-mediated recognition by altering HLA class I expression. It has been noted that some tumor cells have a complete loss of HLA expression due to defects in β2-microglobulin expression or function. Alternatively, chromosomal defects in the HLA-encoding genes themselves can cause selective loss of HLA expression. This process has been noted in approximately 50 % of head and neck squamous cell carcinomas [27] and was correlated with poor prognosis in esophageal squamous cell cancer [28] and laryngeal squamous cell cancer [29].

Tumor cell immune evasion and exploitation. Tumor cells secrete several small molecules and cytokines that depress NK, DC, and T cell function and induce immunosuppressive MDSC and regulatory T cells. MHC downregulation and defects in the antigen presentation machinery impairs T cell recognition. Fas ligand is expressed which kills T cells. Chemokine receptors aid in metastasis of the cancer cell to lymph nodes
On the other hand, cells with complete loss of HLA may evade immune response by T cell recognition but represent a strong trigger for NK cell activation as the absence of HLA removes a key inhibitory signal for NK cells. Therefore, tumor cells must employ multiple mechanisms to realize immunoevasion. One proposed explanation for the lack of NK cell killing is that cancer cells possess defects in their antigen presentation machinery (APM) for HLA molecules that are tumor antigen specific. Endogenous antigens are processed through the cytoplasmic immunoproteasome which consists of various subunits including low molecular weight proteasome (LMP) 2, LMP7, and LMP10. Antigenic peptides are transported to the endoplasmic reticulum by the transporter associated with antigen processing (TAP) where they are associated with HLA class I heavy chains by tapasin [30]. The observation that T cell recognition could be reconstituted with either exogenous peptide or upregulation of APM expression [31] confirms the biological significance of this immune escape mechanism. This reduces selectively tumor antigen-HLA peptide completely without reduction in overall surface HLA density. Thus, SCCHN cells that express HLA I and whole tumor antigen can evade T cell recognition through decreased expression of LMP2, TAP1, TAP2, and tapasin but still maintain nonspecific HLA I expression in order to avoid recognition by NK cells.
In addition to altered expression of HLA, SCCHN tumor cells express Fas ligand which can interact with Fas and transduce a powerful apoptosis signal to activated T cells allowing immune evasion [32] by eliminating tumor-infiltrating T lymphocytes.
Another important group of molecules that emerged in the focus of research is the group of immune checkpoint receptors (ICR). Being part of the immune system’s control mechanisms against overreactive functions in inflammatory response and thus limiting autoimmunity, this mechanism is exploited in the tumor microenvironment. Several receptors have been identified that are expressed on exhausted cells and/or show inhibitory regulation upon stimulation, including cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), lymphocyte activation gene-3 (LAG-3), T cell immunoglobulin and mucin protein-3 (TIM-3), B and T cell lymphocyte attenuator (BTLA), and programmed death-1 (PD-1). Its ligand PD-L1 (B7-H1, CD274) induces a loss of function of cytotoxic T cells (CTL) [33]; PD-L1 is upregulated in multiple tumor cell lines including HNSCC [34]. CTLA-4 is a member of the B7 receptor family expressed by CD4+, CD8+, and regulatory T cells (Treg) [17], which competes with CD28 to stimulatory ligands CD80 and CD86. LAG-3 is another receptor that was shown to enhance Treg function [35]. If TIM-3 is rather a marker or a mediator for immunosuppression is still investigated, studies correlated TIM-3 expression levels with poor clinical outcome [36]. BTLA and its ligand herpesvirus entry mediator (HVEM) are receptors from the immunoglobulin superfamily and were first receptors that demonstrated cross talk between TNF and Ig ligands [37]. It is mainly expressed on B lymphocytes and can induce inhibitory and stimulatory pathways depending on its ligands, although the inhibitory function was shown to be the dominant pathway in knockout mice [38].
As described, these pathways play an important role in tumor immune escape. In the recent years, tremendous advances in the understanding of these described mechanisms have been made, allowing further establishment of immunotherapies that we will outline at the end of this chapter.
6.9 The Establishment of a Cancer-Promoting Tumor Microenvironment
We discussed pathways for the reduction of immune response in direct cell-to-cell contact between tumor cells and two of the main effector cells, T and NK cells. But the points of action through which the tumor cells generate a tumor-promoting environment are much more versatile. They include and are realized through direct (tumor cells) and indirect (induced) cytokine secretion. Many of the cytokines play an important role in cancer formation and can be localized in the setting of chronic inflammation. The fact that some cancers arise at sites of chronic inflammation was first noted by Virchow over a century ago. Since then, chronic inflammatory states have been linked to a myriad of tumors: Helicobacter pylori infection and gastric cancer, inflammatory bowel disease and colon cancer, chronic irritation of the aerodigestive tract by tobacco/alcohol, and SCCHN. Studies of the tumor microenvironment demonstrate infiltration of inflammatory mediators and a complex milieu of cytokines including TGF-β, IL-6, IL-10, GM-CSF, IL-1β, IL-23, and TNF-α as well as chemokines, which are “chemotactic cytokines” that direct immune cell migration. More recent developments link many of those cytokines to the formation of suppressive immune cells like myeloid-derived suppressive cells (MDSC), regulatory T cells (Treg), tumor-associated macrophages (TAM), and their effectors, which are exploited and promoted by the tumor microenvironment.
6.10 Cytokines
Cytokines and other molecules that suppress immune function such as IL-10, TGF-β, IL-6, PGE2, VEGF, and GM-CSF are known to be produced by SCCHN cells. IL-10 reduces activation of cytotoxic T cells and has been correlated with advanced stage head and neck cancer [39]. TGF-β suppresses T cell and NK activation and is a key cytokine in the differentiation of regulator T cells [40]. TGF-β production is increased in preneoplastic oral cavity lesions and promotes angiogenesis and a protumorigenic microenvironment linking it to early tumor formation [41]. IL-6 signals via STAT3 to inhibit DC maturation, NK cell, T cell, neutrophil, and macrophage activation [42] and has been correlated with recurrence and survival in SCCHN [43]. Reduced DC numbers and function have been observed in this disease [44]. STAT3 is a transcription factor that is also involved in several other immunosuppressive pathways such as IL-10 signaling [45], suppression of dendritic cells [46], downregulation of IL-12 [47], and generation of regulatory T cells [48]. PGE2 is a prosurvival, proangiogenic molecule that is produced by many cancers including SCCHN [49, 50]. It is also a potent immunomodulator that decreases T cell proliferation, inhibits Th1 T cells, decreases B cell proliferation, and inhibits maturation and antigen presentation of DC [51]. VEGF, which is primarily thought of as a promoter of angiogenesis, is overexpressed in 90 % of SCCHN [52] and functions to increase the ratio of immature to mature DC in the tumor microenvironment which is thought to lead to T cell anergy [53]. GM-CSF, when produced in large quantities by tumors, recruit MDSC [54, 55] which have been identified in SCCHN.
6.11 Chemokines
Chemokines are a family of small heparin-binding cytokines that direct the movement and migration of leukocytes. There are four groups of chemokines based on the arrangement of cysteine residues near the N-terminus of the proteins: C, CC, CXC, and CX3C. The G-coupled transmembrane chemokine receptors are also divided into these four groups based on their cognate ligand [56]. SCCHN cells have aberrant expression of several chemokines. They overexpress CXCL1 which has been implicated in tumor angiogenesis, nodal metastasis, and leukocyte infiltration. CCL2 is also overexpressed in squamous cell cancer and is thought to have similar functions. CXCL5 is found in metastatic SCCHN and is involved in tumor migration and tumorigenesis. CXCL8, also found in metastatic SCCHN, promotes matrix metalloprotease secretion and subsequent extracellular matrix breakdown and tissue invasion.
Of the chemokine receptors, CXCR4 and CCR7 are of particular interest as these two receptors are overexpressed in malignant cells including SCCHN cells. Increased expression of CXCR4 and its ligand, CXCL12, is associated in SCCHN cells with nodal metastasis, tumor recurrence, and overall survival. Studies of CXCR4 activation have shown increased metastatic potential, induction of matrix metalloprotease and collagenase expression, decreased cell adhesion, and increased cell mobility. CCR7 appears to have similar biological actions. High CCR7 expression is clinically associated with tumor stage, lymphatic invasion, nodal metastasis, and poorer prognosis [57]. A study of chemokine receptor expression differences between primary and metastatic SCCHN cell lines found that only CCR7 was consistently upregulated in metastatic SCCHN [58]. CCR7 also provides tumor survival and invasion signals via the PI3 kinase signal transduction pathway [59]. These actions in tumor cells are similar to the action of CCR7 in dendritic and CD8+ cells where they mediate chemotaxis to lymph nodes and antiapoptotic signals and may explain the predilection of SCCHN to metastasize to lymph nodes where there is a high concentration of chemokines. The production of chemokines and their receptors by SCCHN tumor cells represents exploitation of the immune system to promote tumor survival and metastasis.
A key regulator of the inflammatory response in cancer is the transcription factor NFκB [60] which stimulates many cancer-promoting cytokines and chemokines in SCCHN [61]. NFκB sits downstream of several soluble factors including TNF-α, IL-1, and reactive oxygen species that are produced by macrophages and granulocytes that infiltrate tumor. Of interest in relation to SCCHN, NFκB activation can also be elicited by cigarette smoke condensate, betel nut extract, and EGFR signaling [62–64]. Activation of the NFκB pathway induces several tumor-promoting processes in SCCHN [65]. NFκB is traditionally thought of as a stress response transcription factor because it controls expression of several prosurvival genes such as mdm2, TRAF1, TRAF2, IAP, and Bcl-XL. These act as antiapoptotic signals for tumor cells and confer resistance to natural death pathways for aberrant cells. NFκB also promotes tumor cell proliferation and expansion through regulation of a key cell cycle modulator, cyclin D1. Angiogenesis is promoted by NFκB through VEGF production, and several cytokines including TNF-α, IL-1, IL-6, and IL-8 are induced causing a positive feedback loop. Tissue invasion is promoted by the upregulation of heparinase, matrix metalloprotease, and urokinase. It has also been suggested that NFκB mediates resistance to treatment with chemotherapy and radiation via regulation of growth arrest and DNA damage (GADD) and glutathione S-transferase [66]. The activation of NFκB by inflammatory immune mediators demonstrates yet another subversion and exploitation of the immune system by cancer to promote key aspects of tumor formation and progression.
6.12 Immune Mediators as Cancer Biomarkers
Because of the derangements in production of cytokines and other immunomodulatory molecules caused by cancer, there has been investigation into the possibility of using cytokine profiles as biomarkers. Biomarkers are of considerable interest because they could be useful in early detection of cancer, determination of prognosis, as a marker of treatment response, and selection of optimal treatment regimen. Cytokines as biomarkers have been investigated in SCCHN in several studies. An older study found that serum TNF-α was 100-fold higher in cancer patients than in disease-free controls [67]. A subsequent study linking serum TNF-α levels to cancer status was published, but that paper found IL-6 to be a more sensitive marker than TNF-α [68]. Another cytokine commonly cited in papers as a possible biomarker for detection of tumor is IL-8 which is elevated in recurrent or metastatic cancer [69]. In a study of over 300 subjects encompassing those with active disease, no evidence of disease and healthy smokers, 60 cytokines were measured and a panel of 25 including IL-8, IFN-α, IFN-γ, IL-1, and RANTES could correctly identify active disease with a sensitivity of 84.5 % and a specificity of 92 % [70]. This provided a proof of principle that the immune system may serve as a biosensor of malignancy and disease status. In another study, IL-6, IL-8, VEGF, and hepatocyte growth factor were elevated in cancer patients and decreases over treatment correlated with improved survival. Interestingly, elevated pretreatment VEGF was a good prognostic factor [71]. This is in contrast to a study in non-small cell lung cancer [72] and head and neck cancer (ASCO 2009 A6035) which demonstrated low pretreatment VEGF as a predictor of better treatment response and longer progression-free survival. A large study of 444 patients found that high pretreatment IL-6 is an independent predictor of poor prognosis [43].
6.13 Key Components of the Tumor Microenvironment: Myeloid-Derived Suppressor Cells, Regulatory T Cells, Tumor-Associated Macrophages, and Cancer Stem Cells
Myeloid-derived suppressor cells (MDSC) are a diverse family of myeloid origin with T cell suppressive functions that express surface molecules such as Gr1+(murine) CD11b+, CD33+, and CD34+ [73]. They are increased in almost all cancer patients and, indeed, were first characterized in SCCHN [55] where their link to VEGF and GM-CSF was discovered. In addition to VEGF and GM-CSF, MDSC are induced by IL-6, IL-1β, PGE2, and complement C5a. Initial studies in SCCHN found that MDSC inhibit IL-2 secretion by activated T cells which is a key step in T cell proliferation and escalation of cell-mediated immunity. Also, they deplete the tumor microenvironment of arginine and cysteine which are essential for T cell activation. MDSC produce nitric oxide and reactive oxygen species that catalyze the nitration of the TCR which inhibits TCR-MHC interactions and subsequent activation. Downregulation of the TCR ζ chain which also interferes with T cell activation is mediated by MDSC along [74] with downregulation of l-selectin which is important for migration of naïve T cells to lymph nodes. Data on the effect of MDSC on NK cells has been conflicting with reports of both enhancing as well as suppressive action on NK cells which may be a function of the heterogeneity of MDSC populations. MDSC also promote induction of Tregs via production of IL-10, TGF-β, and arginase [73]. Treatments such as antibody depletion, retinoic acid, gemcitabine, and STAT3 blockade diminish MDSC, restore immune surveillance, increase T cell activation, and improve efficacy of immunotherapy. The basal levels of MSDC increase with age and may contribute to increased tumor frequency and growth rate increase with age [75].
Though it was long suspected that a subset of T cells was immunosuppressive, the characterization occurred relatively recently when it was found that this subpopulation represented CD4+ cells that also expressed CD25 [76]. So far, four subtypes of regulatory T cells have been identified: naturally occurring thymus-derived CD4+CD25highFoxP3+Tregs, antigen-induced IL-10-dependent Tregs (Tr1), IL-4-dependent Tregs (Th3), and antigen-specific Tregs [16]. There is also a CD8+CD25+ variant which also appears to have immunosuppressive ability, but their biological significance is unclear and they are thought to be overshadowed by the much more abundant CD4+ Tregs [77]. Tregs cause anergy, apoptosis, and cell cycle arrest of activated T cells via production of IL-10, TGF-β, and direct cell-to-cell contact [78]. They also inhibit the action of dendritic cells, NK cells, and B cells [79]. In SCCHN patients, Tregs are increased in frequency in peripheral blood and among T cells infiltrating the tumor and draining lymph nodes resulting in an immunosuppressed state [17, 80, 81]. Also, Treg numbers are inversely proportional to DC and CD8+ T cell numbers in SCCHN [82, 83]. Treg frequency as a prognostic indicator is unclear as one study linked increased Tregs with better locoregional control [84] while another study found increased Tregs associated with early recurrence [85]. Additionally, Treg frequency is elevated in SCCHN patients after treatment, indicating that oncologic treatment increases Treg numbers [17].
A different cellular component in the tumor microenvironment is represented by macrophages. They can engage different phenotypic functions depending on environmental conditions. Two main subsets are distinguished. On the one hand, the M1 phenotype is being activated by IFNγ and Toll-like receptors (TLR), producing proinflammatory cytokines such as TNF-α with a T cell-stimulating effect resolving in a TH1 response. On the other hand, the alternatively activated macrophages (M2) force a TH2 response, with production of interleukins such as IL-4 and IL-13. Obviously, there is no black and white in a plastic cell population such as the macrophages, but there have been several studies linking the number of TAM in tumor to a worse clinical outcome and characterizing their phenotype closely associated to the M2 population. TAMs have been demonstrated to produce EGF, IL-6, and IL-10 and have been associated with angiogenesis, local tumor progression, and metastasis [86].
Additionally to these immunosuppressive cell types with origin from the immune system, a hypothesis about treatment-resistant and adoptive tumor cells—so-called cancer stem cells—that provide another challenge is being investigated. Recently, there has been growing interest in the cancer stem cell hypothesis. Heterogeneity in tumor cells has long been accepted, and this theory postulates the existence of a subpopulation of tumor cells that are pluripotent and able to effectively recapitulate the entire heterogeneous tumor when transferred to another site. They are known to be more resistant than other tumor cells to chemotherapy as well as radiation [87]. Several defining markers of these stem cells have been proposed. The first marker proposed was CD44 [88], a cell surface glycoprotein which binds hyaluronate but may also inhibit the action of the p53 tumor suppressor in cancer cells [89]. However, CD44 expression is abundant in normal epithelia, and its utility as a cancer stem cell marker is questionable [90]. Another proposed marker is aldehyde dehydrogenase 1 (ALDH1) which is found in many embryonic stem cells and was identified as the responsible protein in conferring resistance to chemotherapeutic agents in stem cells [91]. Because these cancer stem cells are able to reconstitute the entire tumor, many believe that, ultimately, it is treatment of this small population of resistant cells that determines the success or failure of oncologic therapy. If this is the case, it is important that these cells be addressed in any treatment regimen. Because ALDH1 is not highly expressed in normal tissues, its potential as a tumor antigen target has been recently explored [92].
These data indicate that SCCHN induces an immunosuppressed state via multiple potent mechanisms which is a barrier to effective cancer immunotherapy. They secrete immunosuppressive cytokines and molecules. Cytokine levels are aberrant in SCCHN patients indicating deregulation or dysregulation of cytokine pathways [93]. There is increased frequency of immunosuppressive regulatory immune cells, and there is a global dysfunction of almost every facet of the immune system in SCCHN patients.
6.14 Head and Neck Cancer Immunotherapy
As described, the tumor’s interactions with the immune system and the evasion strategies are manifold. Therefore, therapeutic approaches have to be versatile as well. They include vaccination therapies and monoclonal antibody (mAb)-based therapies for multiple, different targets. Playing a central role in immunology, vaccination approaches for tumor therapy have been developed in the past years.
There are several strategies for delivering tumor vaccines with each having inherent advantages and disadvantages. All methods depend on delivering an antigen to the host in an effort to elicit an adaptive cellular immune response to the tumor antigen. Most methods require the use of a specific known tumor antigen, but some can use entire tumor cells as part of the vaccine to activate the immune system against multiple unspecified and unknown tumor antigens.
DNA vaccines utilize delivery of naked DNA encoding a known tumor antigen to the patient. This DNA is taken up by cells, and the antigen is expressed for subsequent processing and presentation by DC. DNA vaccines are safe, inexpensive, and easy to deliver and do not induce the formation of neutralizing antibodies allowing repeated administration. However, they have low transfection efficiency and elicit a very weak immune response and therefore are often engineered to encode proteins that target DC or are given with adjuvant agents that increase DC activation. Currently in SCCHN, a DNA vaccine encoding an HPV-16 E6/E7 fusion protein is under development for HPV-positive SCCHN [94], and another vaccine encoding Hsp65 has been tested in a phase I trial [95] and demonstrated clinical response in 4 out of 14 patients with recurrent unresectable SCCHN.
Bacterial/viral vaccines can deliver tumor antigen and function as an immune adjuvant because the immune system responds to a perceived infection. They are very immunogenic, relatively inexpensive, and easy to manufacture but have the downsides of potential toxicity, preexisting neutralizing antibodies, or the formation of antibodies against the bacterial or viral vector limiting repeat dosing or effectiveness. Also, these tend to elicit a stronger humoral rather than cellular immune response which is less desirable. Several such vaccines are currently under development: HPV-16 E7 Listeria vaccine [96], Vaccinia-based E6/E7 vaccine [97], and a Vaccinia-based E2 [98].
Peptide vaccines consist of synthesized peptides that have been designed to correspond to an epitope on a tumor antigen that binds well to the cleft of an HLA molecule. They are similar to DNA vaccines in that they are safe and inexpensive with low immunogenicity but have the added drawback of being restricted to the HLA subclass for which they were designed. The popular HLA subclass used in vaccine design is HLA-A2 as this is the most common subclass found in Caucasians. Clinical trials are under way with a MAGE-A3/HPV-16 peptide (NCT00257738) and an LMP2 peptide for EBV-related nasopharyngeal carcinoma (NCT00078494).
To circumvent HLA restriction, whole proteins can be used as a vaccine. Whole proteins can be processed by the antigen-presenting cells and presented on self-MHC to cause activation of T cells. However, the vast majority of identified tumor antigen proteins are self-proteins, and therefore, the patient’s immune system is tolerant to these proteins. Therefore, there is tremendous difficulty in producing an effective immune response with protein vaccines.
Tumor cell vaccines are similar to whole protein vaccines in that they are not HLA restricted and specific epitopes need not be known for their use. Often the tumor cells are given with adjuvant agents or modified by viral infection to improve their immunogenicity. A Newcastle disease virus-infected tumor cell vaccine was found to induce a specific T cell response and [99] that correlated with better clinical outcome. These vaccines tend to be labor intensive because the tumor has to be isolated and processed before it can be used as a vaccine.
Dendritic cells are the most potent activators of antigen-specific T cells, and consequently, DC vaccines are the most widely studied cancer vaccine strategy. This is an extremely labor-intensive method in which dendritic cells are isolated from each patient and they are loaded with tumor antigen ex vivo. This loading can be in the form of peptides, proteins, DNA transfection, tumor cell lysates, apoptotic tumors, necrotic tumors, or cell fusion. After DC are loaded with tumor antigen, they undergo maturation and activation with various cytokine cocktails to prime them for presenting the tumor antigen to T cells. These DC are then introduced to the patients, usually into the tumor or into lymph nodes. Several DC-based vaccines are currently being developed for SCCHN: intratumoral injection of DC (NCT00492947), multivalent p53 DC vaccine [100], and lysyl oxidase-like-4 transfected DC [101]. Efforts to reverse the immunosuppression associated with cancer include stimulating cocktails of multiple cytokines delivered systemically to improve immune competence.
Besides cytokines, different drugs imply a strong stimulating potential for immunomodulation in anticancer therapy. A group that is investigated in clinical trials of HNSCC is the group of TLR agonists. TLR are pattern recognition receptors (PRR) that can recognize so-called pathogen-associated molecular patterns (PAMPs) from external pathogens such as viral RNA, bacterial DNA, or surface molecules. Therefore, they are closely associated to the innate immune response and stimulate upon activation various parts of the immune system, notably cells from myeloid origin. This effect is utilized in anticancer therapy. TLR agonists induce the maturation and cross-priming of dendritic cells (DC) and have been shown to induce NK cell-dependent lysis of tumor cells in combination with mAb such as antiepidermal growth factor receptor (EGFR) cetuximab [102]. Clinical trials of TLR agonists in combination with mAb are evolving and include HNSCC-specific trials like a neoadjuvant phase Ib trial (NCT02124850) or adjuvant trials in patients with locally advanced, recurrent, or metastatic HNSCC (NCT01836029, NCT01334177).
6.15 Monoclonal Antibody-Based Immunotherapy of SCCHN
Today the most widely used form of cancer immunotherapy is mAb therapy. Therein, the different targets are distinguished, aiming the previously discussed different mechanisms of tumor-promoting effects: Tumor antigen (TA)-targeted mAbs, cytokine-targeted mAbs, tumor necrosis factor receptor (TNFR)-family costimulatory targeted mAbs and immune checkpoint-targeted mAbs. Currently available mAbs that are circumstantially investigated in head and neck cancer are listed in Table 6.1. The most extensively studied of these is cetuximab, a mouse-human chimeric IgG1 antiepidermal growth factor receptor mAb [103]. EGFR is an attractive target in SCCHN because it is overexpressed in 80–90 % of SCCHN and leads to tumor cell proliferation, invasion, angiogenesis, tumor survival, and, consequently, poor survival and prognosis [104, 105].
It is becoming clear that anti-EGFR mAb mediate antigen-specific immune responses to targeted tumors (Fig. 6.2). There are two major mechanisms by which mAb can activate the immune system against a tumor target, direct killing via lytic immune cell (NK cell or monocytes) and complement fixation, or opsonization of tumor for phagocytosis and subsequent antigen processing. The latter would induce TA-specific cytotoxic T lymphocytes (CTL) to recognize and lyse tumor cells. One of the most direct methods by which antibodies can cause tumor lysis is via antibody-dependent cellular cytotoxicity (ADCC) mediated by NK cells and probably monocytes and neutrophils. Panitumumab and cetuximab both mediate ADCC [106], and the extent of ADCC is heavily influenced by genetic polymorphisms in FcγRIIIa, also known as CD16 [107]. Complement activation via the classical pathway is another major effector of humoral immunity and is activated by IgM, IgG1, IgG2, and IgG3. A combination of cetuximab and matuzumab can elicit complement-dependent cytotoxicity in vitro [108]. In addition to direct activation of NK cell lysis of tumor cells, TA-specific mAbs can elicit CD8+ T cell responses to tumor-derived antigens through interaction with FcγRs on antigen-presenting cells (APC). In human cells, there are three activating FcγRs, FcγRI, FcγRIIa, and FcγRIII, and one inhibiting FcγR, FcγRIIB [109] with FcγRIIa being the dominant receptor on APC. This antigen-specific T cell activation was noted in 78 % of patients treated with trastuzumab for breast cancer, and this activation seemed to correlate positively with clinical response [110]. Specific T cell activation has been demonstrated in a model using glioma and cetuximab [111], and it is likely that similar T cell activation also occurs in SCCHN patients treated with anti-EGFR mAbs [112].

Schematic representation of ADCC; the effector mAb has a constant fragment [Fc] that interacts with immune effector cells and a variable fragment [F(ab)] that is antigen (EGFR) specific. During cross-presentation, tumor antigens are degraded in the cytoplasm of dendritic cells (DC) and presented to T cells producing a cellular immune response [Reprinted from Lee S, Lopez-Albaitero A, Ferris RL. Immunotherapy of head and neck cancer using tumor antigen-specific monoclonal antibodies. Current Oncology Reports. 2009;11(2), 156–162. With permission from Springer Science + Business Media]
The mechanism for TA-specific T cell induction may actually be enhanced by ADCC and NK cell activation. In addition to their ability to mediate ADCC, activated NK cells, particularly CD56bright NK cells [113] have also been shown to secrete cytokines, such as IFN-γ, TNF-α, and chemokines, such as macrophage inflammatory protein-(MIP)-1α, MIP-1β, and RANTES, that inhibit tumor cell proliferation, enhance antigen presentation, and aid in the chemotaxis of T cells [107, 114]. Indeed, NK cells can interact with other innate immune cells that are present during the early phases of inflammatory responses [115]. This so-called NK cell-DC cross talk follows the recruitment of both NK cells and DC to sites of inflammation [116, 117], resulting in potent activating bidirectional signaling. NK cells in the presence of cytokines released by DC become activated, regulating both the quality and the intensity of innate immune responses. Also, activated NK cells release cytokines that favor DC maturation and select the most suitable DC for subsequent migration to lymph nodes and efficient T cell priming. In addition, IFN-γ secreting NK cells can be recruited directly to the lymph nodes to enhance T cell induction [117]. Elevated levels of the NK cell-derived chemokines IL-8, macrophage inflammatory protein-1, and RANTES have been detected within the sera of trastuzumab-responding cancer patients [114]. These NK cell factors could induce the chemotaxis of naive and activated T cells, as indicated by the correlation of their presence with the infiltration of tumor tissue by CD8+ CTL. These data suggest that NK cell cytokine and chemokine production may enhance DC cross-presentation and T cell induction, with the potential to spread it to other TA [118].
In order to face the immunosuppressing cytokines of the tumor microenvironment (NK cell cytolysis, induction of Treg [119]) that are associated with prognosis and clinical outcome [71], cytokine-targeted mAbs are developed. One of them is bevacizumab, which is a humanized IgG1 specific against VEGF-A (FDA approved for NSCLC and colon cancer). A phase II trial of a combination of bevacizumab and erlotinib in SCCHN demonstrated a response rate of 14.6 % and an overall mean survival of 6.8 months [52]. A phase II trial that investigated bevacizumab in combination with docetaxel and radiation in locally advanced HNSCC showed tolerability and effectiveness (3-year PFS 61.8 %) [120]. Ficlatuzumab is a humanized anti-hepatocyte growth factor (HGF) mAb that is currently in phase I trial in combination with cetuximab.
So far, the investigation of TNFR-targeting mAb in clinical trials for HNSCC is in phase I. Because of the important costimulatory pathways for immune cell activation, substances like CP-870,893 (Pfizer), an IgG2 CD40 agonist; OX40 mAb (AgonOx, Providence Health), an IgG2 OX40 agonist; or urelumab (Bristol-Myers Squibb), an IgG4 CD137 agonist, have been investigated in multiple anticancer trials [121].
Other strategies target specific inhibitory molecules. To reduce T cell anergy, anti-CTLA-4 antibodies are developed to block the inhibitory signal [94] alone or in combination with other mAb such as cetuximab (NCT01935921). The same accounts for the previously described PD-1 [95], and antagonistic antibodies to this protein have demonstrated efficacy in phase II trials [96]. Based on promising results of the use of PD-1 blocking monoclonal antibodies (mAb) in different cancer subsets (phase I-III), clinical trials of phase I and phase II have emerged for advanced recurrent or metastatic HNSCC [6]. A different group of receptors with a modulating effect on immune cells is the killer cell immunoglobulin-like receptors (KIRs). They interact with MHC I molecules and regulate immune response. Most of the receptors have a suppressing effect on the cytotoxicity. Anti-KIR antibodies remove the major inhibitory signal on NK cells. Ongoing trials are investigating an anti-KIR mAb in combination with the anti-CTLA-4 mAb ipilimumab (NCT01750580) or anti-PD-1 mAb nivolumab (NCT01714739).
6.16 Conclusion
Cancer immunology is a rapidly evolving field, and it is only recently that we have begun to understand the complex interaction between cancer and the host immune system. Tumor cells demonstrate several methods to exploit the immune system to help promote angiogenesis, derive prosurvival and proliferative signals, and induce metastasis and tumor progression. At the same time, cancers are able to cloak themselves from the immune system by self-modification and by immunosuppression of the host. Recent results from clinical trials show evidence for effective anticancer immunotherapies. Because of the manifold tumor evasion strategies and hence different response rates for treatments, combinational therapies urge into focus for cancer treatment. These insights and better understanding of the workings of the immune system have allowed the recent explosion of several promising immunotherapeutic agents that are currently in clinical use as well as under development.
References
Vlock DR, Schantz SP, Fisher SG, Savage HE, Carey TE, Wolf GT. Clinical correlates of circulating immune complexes and antibody reactivity in squamous cell carcinoma of the head and neck. The Department of Veterans Affairs Laryngeal Cancer Study Group. J Clin Oncol. 1993;11:2427–33.
Calenoff E, Cheever MA, Satam M, Dutra JC, Pelzer HJ, Kern RC, Hanson DG. Serum immunoglobulins specific for intracellular proteins of squamous cell carcinoma. Arch Otolaryngol Head Neck Surg. 1995;121:183–91.
Couch ME, Ferris RL, Brennan JA, Koch WM, Jaffee EM, Leibowitz MS, Nepom GT, Erlich HA, Sidransky D. Alteration of cellular and humoral immunity by mutant p53 protein and processed mutant peptide in head and neck cancer. Clin Cancer Res. 2007;13:7199–206.
Rabassa ME, Croce MV, Pereyra A, Segal-Eiras A. MUC1 expression and anti-MUC1 serum immune response in head and neck squamous cell carcinoma (HNSCC): a multivariate analysis. BMC Cancer. 2006;6:253.
Yamaguchi K, Patturajan M, Trink B, Usadel H, Koch W, Jen J, Sidransky D. Circulating antibodies to p40(AIS) in the sera of respiratory tract cancer patients. Int J Cancer. 2000;89:524–8.
Tominaga O, Unsal K, Zalcman G, Soussi T. Detection of p73 antibodies in patients with various types of cancer: immunological characterization. Br J Cancer. 2001;84:57–63.
Zumbach K, Hoffmann M, Kahn T, Bosch F, Gottschlich S, Gorogh T, Rudert H, Pawlita M. Antibodies against oncoproteins E6 and E7 of human papillomavirus types 16 and 18 in patients with head-and-neck squamous-cell carcinoma. Int J Cancer. 2000;85:815–8.
Shimada H, Shiratori T, Takeda A, Matsushita K, Okazumi S, Akutsu Y, Matsubara H, Nomura F, Ochiai T. Perioperative changes of serum p53 antibody titer is a predictor for survival in patients with esophageal squamous cell carcinoma. World J Surg. 2009;33:272–7.
Neuchrist C, Kornfehl J, Grasl M, Lassmann H, Kraft D, Ehrenberger K, Scheiner O. Distribution of immunoglobulins in squamous cell carcinoma of the head and neck. Int Arch Allergy Immunol. 1994;104:97–100.
Miller JF. Effect of neonatal thymectomy on the immunological responsiveness of the mouse. Proc Roy Soc B. 1962;156:2.
Miller JF, Mitchell GF. Cell to cell interaction in the immune response. I. Hemolysin-forming cells in neonatally thymectomized mice reconstituted with thymus or thoracic duct lymphocytes. J Exp Med. 1968;128:801–20.
Mitchell GF, Miller JF. Cell to cell interaction in the immune response. II. The source of hemolysin-forming cells in irradiated mice given bone marrow and thymus or thoracic duct lymphocytes. J Exp Med. 1968;128:821–37.
Masopust D, Vezys V, Wherry EJ, Ahmed R. A brief history of CD8 T cells. Eur J Immunol. 2007;37 Suppl 1:S103–10.
Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol. 1989;7:145–73.
Korn T, Oukka M, Kuchroo V, Bettelli E. Th17 cells: effector T cells with inflammatory properties. Semin Immunol. 2007;19:362–71.
Bergmann C, Strauss L, Wang Y, Szczepanski MJ, Lang S, Johnson JT, Whiteside TL. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: mechanisms of suppression and expansion in advanced disease. Clin Cancer Res. 2008;14:3706–15.
Strauss L, Bergmann C, Gooding W, Johnson JT, Whiteside TL. The frequency and suppressor function of CD4+CD25highFoxp3+ T cells in the circulation of patients with squamous cell carcinoma of the head and neck. Clin Cancer Res. 2007;13:6301–11.
Herberman RB, Nunn ME, Holden HT, Lavrin DH. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. II. Characterization of effector cells. Int J Cancer. 1975;16:230–9.
Miller JS. The biology of natural killer cells in cancer, infection, and pregnancy. Exp Hematol. 2001;29:1157–68.
Herberman RB, Holden HT. Natural cell-mediated immunity. Adv Cancer Res. 1978;27:305–77.
Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Cancer immunoediting: from immunosurveillance to tumor escape. Nat Immunol. 2002;3:991–8.
Russell JH, Ley TJ. Lymphocyte-mediated cytotoxicity. Annu Rev Immunol. 2002;20:323–70.
Gillison ML. Oropharyngeal cancer: a potential consequence of concomitant HPV and HIV infection. Curr Opin Oncol. 2009;21:439–44.
Jain A, Reyes J, Kashyap R, Rohal S, Abu-Elmagd K, Starzl T, Fung J. What have we learned about primary liver transplantation under tacrolimus immunosuppression? Long-term follow-up of the first 1000 patients. Ann Surg. 1999;230:441–8. discussion 448–449.
Birkeland SA, Storm HH, Lamm LU, Barlow L, Blohme I, Forsberg B, Eklund B, Fjeldborg O, Friedberg M, Frodin L, et al. Cancer risk after renal transplantation in the Nordic countries, 1964–1986. Int J Cancer. 1995;60:183–9.
Shankaran V, Ikeda H, Bruce AT, White JM, Swanson PE, Old LJ, Schreiber RD. IFNgamma and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature. 2001;410:1107–11.
Grandis JR, Falkner DM, Melhem MF, Gooding WE, Drenning SD, Morel PA. Human leukocyte antigen class I allelic and haplotype loss in squamous cell carcinoma of the head and neck: clinical and immunogenetic consequences. Clin Cancer Res. 2000;6:2794–802.
Mizukami Y, Kono K, Maruyama T, Watanabe M, Kawaguchi Y, Kamimura K, Fujii H. Downregulation of HLA Class I molecules in the tumour is associated with a poor prognosis in patients with oesophageal squamous cell carcinoma. Br J Cancer. 2008;99:1462–7.
Ogino T, Shigyo H, Ishii H, Katayama A, Miyokawa N, Harabuchi Y, Ferrone S. HLA class I antigen down-regulation in primary laryngeal squamous cell carcinoma lesions as a poor prognostic marker. Cancer Res. 2006;66:9281–9.
Ferris RL, Whiteside TL, Ferrone S. Immune escape associated with functional defects in antigen-processing machinery in head and neck cancer. Clin Cancer Res. 2006;12:3890–5.
Lopez-Albaitero A, Nayak JV, Ogino T, Machandia A, Gooding W, DeLeo AB, Ferrone S, Ferris RL. Role of antigen-processing machinery in the in vitro resistance of squamous cell carcinoma of the head and neck cells to recognition by CTL. J Immunol. 2006;176:3402–9.
Gastman BR, Atarshi Y, Reichert TE, Saito T, Balkir L, Rabinowich H, Whiteside TL. Fas ligand is expressed on human squamous cell carcinomas of the head and neck, and it promotes apoptosis of T lymphocytes. Cancer Res. 1999;59:5356–64.
Zou W, Chen L. Inhibitory B7-family molecules in the tumour microenvironment. Nat Rev Immunol. 2008;8:467–77.
Zandberg DP, Strome SE. The role of the PD-L1:PD-1 pathway in squamous cell carcinoma of the head and neck. Oral Oncol. 2014;50:627–32.
Sega EI, Leveson-Gower DB, Florek M, Schneidawind D, Luong RH, Negrin RS. Role of lymphocyte activation gene-3 (Lag-3) in conventional and regulatory T cell function in allogeneic transplantation. PLoS One. 2014;9, e86551.
Yang ZZ, Grote DM, Ziesmer SC, Niki T, Hirashima M, Novak AJ, Witzig TE, Ansell SM. IL-12 upregulates TIM-3 expression and induces T cell exhaustion in patients with follicular B cell non-Hodgkin lymphoma. J Clin Invest. 2012;122:1271–82.
Sedy JR, Gavrieli M, Potter KG, Hurchla MA, Lindsley RC, Hildner K, Scheu S, Pfeffer K, Ware CF, Murphy TL, Murphy KM. B and T lymphocyte attenuator regulates T cell activation through interaction with herpesvirus entry mediator. Nat Immunol. 2005;6:90–8.
Wang Y, Subudhi SK, Anders RA, Lo J, Sun Y, Blink S, Wang Y, Wang J, Liu X, Mink K, et al. The role of herpesvirus entry mediator as a negative regulator of T cell-mediated responses. J Clin Invest. 2005;115:711–7.
Jebreel A, Mistry D, Loke D, Dunn G, Hough V, Oliver K, Stafford N, Greenman J. Investigation of interleukin 10, 12 and 18 levels in patients with head and neck cancer. J Laryngol Otol. 2007;121:246–52.
Moutsopoulos NM, Wen J, Wahl SM. TGF-beta and tumors–an ill-fated alliance. Curr Opin Immunol. 2008;20:234–40.
Lu SL, Reh D, Li AG, Woods J, Corless CL, Kulesz-Martin M, Wang XJ. Overexpression of transforming growth factor beta1 in head and neck epithelia results in inflammation, angiogenesis, and epithelial hyperproliferation. Cancer Res. 2004;64:4405–10.
Cheng F, Wang HW, Cuenca A, Huang M, Ghansah T, Brayer J, Kerr WG, Takeda K, Akira S, Schoenberger SP, et al. A critical role for Stat3 signaling in immune tolerance. Immunity. 2003;19:425–36.
Duffy SA, Taylor JM, Terrell JE, Islam M, Li Y, Fowler KE, Wolf GT, Teknos TN. Interleukin-6 predicts recurrence and survival among head and neck cancer patients. Cancer. 2008;113:750–7.
Hoffmann TK, Muller-Berghaus J, Ferris RL, Johnson JT, Storkus WJ, Whiteside TL. Alterations in the frequency of dendritic cell subsets in the peripheral circulation of patients with squamous cell carcinomas of the head and neck. Clin Cancer Res. 2002;8:1787–93.
Murray PJ. STAT3-mediated anti-inflammatory signalling. Biochem Soc Trans. 2006;34:1028–31.
Sun Y, Chin YE, Weisiger E, Malter C, Tawara I, Toubai T, Gatza E, Mascagni P, Dinarello CA, Reddy P. Cutting edge: negative regulation of dendritic cells through acetylation of the nonhistone protein STAT-3. J Immunol. 2009;182:5899–903.
Kortylewski M, Xin H, Kujawski M, Lee H, Liu Y, Harris T, Drake C, Pardoll D, Yu H. Regulation of the IL-23 and IL-12 balance by Stat3 signaling in the tumor microenvironment. Cancer Cell. 2009;15:114–23.
Pallandre JR, Brillard E, Crehange G, Radlovic A, Remy-Martin JP, Saas P, Rohrlich PS, Pivot X, Ling X, Tiberghien P, Borg C. Role of STAT3 in CD4+CD25+FOXP3+ regulatory lymphocyte generation: implications in graft-versus-host disease and antitumor immunity. J Immunol. 2007;179:7593–604.
Snyderman CH, Milanovich M, Wagner RL, Johnson JT. Prognostic significance of prostaglandin E2 production in fresh tissues of head and neck cancer patients. Head Neck. 1995;17:108–13.
Camacho M, Leon X, Fernandez-Figueras MT, Quer M, Vila L. Prostaglandin E(2) pathway in head and neck squamous cell carcinoma. Head Neck. 2008;30:1175–81.
Harris SG, Padilla J, Koumas L, Ray D, Phipps RP. Prostaglandins as modulators of immunity. Trends Immunol. 2002;23:144–50.
Seiwert TY, Cohen EE. Targeting angiogenesis in head and neck cancer. Semin Oncol. 2008;35:274–85.
Johnson BF, Clay TM, Hobeika AC, Lyerly HK, Morse MA. Vascular endothelial growth factor and immunosuppression in cancer: current knowledge and potential for new therapy. Expert Opin Biol Ther. 2007;7:449–60.
Gabrilovich D. Mechanisms and functional significance of tumour-induced dendritic-cell defects. Nat Rev Immunol. 2004;4:941–52.
Pak AS, Wright MA, Matthews JP, Collins SL, Petruzzelli GJ, Young MR. Mechanisms of immune suppression in patients with head and neck cancer: presence of CD34(+) cells which suppress immune functions within cancers that secrete granulocyte-macrophage colony-stimulating factor. Clin Cancer Res. 1995;1:95–103.
Koizumi K, Hojo S, Akashi T, Yasumoto K, Saiki I. Chemokine receptors in cancer metastasis and cancer cell-derived chemokines in host immune response. Cancer Sci. 2007;98:1652–8.
Ding Y, Shimada Y, Maeda M, Kawabe A, Kaganoi J, Komoto I, Hashimoto Y, Miyake M, Hashida H, Imamura M. Association of CC chemokine receptor 7 with lymph node metastasis of esophageal squamous cell carcinoma. Clin Cancer Res. 2003;9:3406–12.
Wang J, Xi L, Hunt JL, Gooding W, Whiteside TL, Chen Z, Godfrey TE, Ferris RL. Expression pattern of chemokine receptor 6 (CCR6) and CCR7 in squamous cell carcinoma of the head and neck identifies a novel metastatic phenotype. Cancer Res. 2004;64:1861–6.
Wang J, Zhang X, Thomas SM, Grandis JR, Wells A, Chen ZG, Ferris RL. Chemokine receptor 7 activates phosphoinositide-3 kinase-mediated invasive and prosurvival pathways in head and neck cancer cells independent of EGFR. Oncogene. 2005;24:5897–904.
Karin M. Nuclear factor-kappaB in cancer development and progression. Nature. 2006;441:431–6.
Van Waes C, Yu M, Nottingham L, Karin M. Inhibitor-kappaB kinase in tumor promotion and suppression during progression of squamous cell carcinoma. Clin Cancer Res. 2007;13:4956–9.
Anto RJ, Mukhopadhyay A, Shishodia S, Gairola CG, Aggarwal BB. Cigarette smoke condensate activates nuclear transcription factor-kappaB through phosphorylation and degradation of IkappaB(alpha): correlation with induction of cyclooxygenase-2. Carcinogenesis. 2002;23:1511–8.
Lin SC, Lu SY, Lee SY, Lin CY, Chen CH, Chang KW. Areca (betel) nut extract activates mitogen-activated protein kinases and NF-kappaB in oral keratinocytes. Int J Cancer. 2005;116:526–35.
Bancroft CC, Chen Z, Yeh J, Sunwoo JB, Yeh NT, Jackson S, Jackson C, Van Waes C. Effects of pharmacologic antagonists of epidermal growth factor receptor, PI3K and MEK signal kinases on NF-kappaB and AP-1 activation and IL-8 and VEGF expression in human head and neck squamous cell carcinoma lines. Int J Cancer. 2002;99:538–48.
Ferris RL, Grandis JR. NF-kappaB gene signatures and p53 mutations in head and neck squamous cell carcinoma. Clin Cancer Res. 2007;13:5663–4.
Allen CT, Ricker JL, Chen Z, Van Waes C. Role of activated nuclear factor-kappaB in the pathogenesis and therapy of squamous cell carcinoma of the head and neck. Head Neck. 2007;29:959–71.
Soylu L, Ozcan C, Cetik F, Paydas S, Kiroglu M, Aydogan B, Sargin O, Ozsahinoglu C, Seyrek E. Serum levels of tumor necrosis factor in squamous cell carcinoma of the head and neck. Am J Otolaryngol. 1994;15:281–5.
Jablonska E, Piotrowski L, Grabowska Z. Serum Levels of IL-1b, IL-6, TNF-a, sTNF-RI and CRP in Patients with Oral Cavity Cancer. Pathol Oncol Res. 1997;3:126–9.
Gokhale AS, Haddad RI, Cavacini LA, Wirth L, Weeks L, Hallar M, Faucher J, Posner MR. Serum concentrations of interleukin-8, vascular endothelial growth factor, and epidermal growth factor receptor in patients with squamous cell cancer of the head and neck. Oral Oncol. 2005;41:70–6.
Linkov F, Lisovich A, Yurkovetsky Z, Marrangoni A, Velikokhatnaya L, Nolen B, Winans M, Bigbee W, Siegfried J, Lokshin A, Ferris RL. Early detection of head and neck cancer: development of a novel screening tool using multiplexed immunobead-based biomarker profiling. Cancer Epidemiol Biomarkers Prev. 2007;16:102–7.
Allen C, Duffy S, Teknos T, Islam M, Chen Z, Albert PS, Wolf G, Van Waes C. Nuclear factor-kappaB-related serum factors as longitudinal biomarkers of response and survival in advanced oropharyngeal carcinoma. Clin Cancer Res. 2007;13:3182–90.
Hanrahan EO, Ryan AJ, Mann H, Kennedy SJ, Langmuir P, Natale RB, Herbst RS, Johnson BE, Heymach JV. Baseline vascular endothelial growth factor concentration as a potential predictive marker of benefit from vandetanib in non-small cell lung cancer. Clin Cancer Res. 2009;15:3600–9.
Ostrand-Rosenberg S, Sinha P. Myeloid-derived suppressor cells: linking inflammation and cancer. J Immunol. 2009;182:4499–506.
Umansky V, Sevko A. Overcoming immunosuppression in the melanoma microenvironment induced by chronic inflammation. Cancer Immunol Immunother. 2012;61:275–82.
Grizzle WE, Xu X, Zhang S, Stockard CR, Liu C, Yu S, Wang J, Mountz JD, Zhang HG. Age-related increase of tumor susceptibility is associated with myeloid-derived suppressor cell mediated suppression of T cell cytotoxicity in recombinant inbred BXD12 mice. Mech Ageing Dev. 2007;128:672–80.
Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. 1995;155:1151–64.
Cosmi L, Liotta F, Lazzeri E, Francalanci M, Angeli R, Mazzinghi B, Santarlasci V, Manetti R, Vanini V, Romagnani P, et al. Human CD8+CD25+ thymocytes share phenotypic and functional features with CD4+CD25+ regulatory thymocytes. Blood. 2003;102:4107–14.
Alhamarneh O, Amarnath SM, Stafford ND, Greenman J. Regulatory T cells: what role do they play in antitumor immunity in patients with head and neck cancer? Head Neck. 2008;30:251–61.
Ralainirina N, Poli A, Michel T, Poos L, Andres E, Hentges F, Zimmer J. Control of NK cell functions by CD4+CD25+ regulatory T cells. J Leukoc Biol. 2007;81:144–53.
Strauss L, Bergmann C, Szczepanski M, Gooding W, Johnson JT, Whiteside TL. A unique subset of CD4+CD25highFoxp3+ T cells secreting interleukin-10 and transforming growth factor-beta1 mediates suppression in the tumor microenvironment. Clin Cancer Res. 2007;13:4345–54.
Strauss L, Bergmann C, Whiteside TL. Functional and phenotypic characteristics of CD4+CD25highFoxp3+ Treg clones obtained from peripheral blood of patients with cancer. Int J Cancer. 2007;121:2473–83.
Sakakura K, Chikamatsu K, Takahashi K, Whiteside TL, Furuya N. Maturation of circulating dendritic cells and imbalance of T-cell subsets in patients with squamous cell carcinoma of the head and neck. Cancer Immunol Immunother. 2006;55:151–9.
Chikamatsu K, Sakakura K, Whiteside TL, Furuya N. Relationships between regulatory T cells and CD8+ effector populations in patients with squamous cell carcinoma of the head and neck. Head Neck. 2007;29:120–7.
Badoual C, Hans S, Rodriguez J, Peyrard S, Klein C, Agueznay Nel H, Mosseri V, Laccourreye O, Bruneval P, Fridman WH, et al. Prognostic value of tumor-infiltrating CD4+ T-cell subpopulations in head and neck cancers. Clin Cancer Res. 2006;12:465–72.
Boucek J, Mrkvan T, Chovanec M, Kuchar M, Betka J, Boucek V, Hladikova M, Betka J, Eckschlager T, Rihova B. Regulatory T cells and their prognostic value for patients with squamous cell carcinoma of the head and neck. J Cell Mol Med. 2010;14:426–33.
Komohara Y, Jinushi M, Takeya M. Clinical significance of macrophage heterogeneity in human malignant tumors. Cancer Sci. 2014;105:1–8.
Tang C, Ang BT, Pervaiz S. Cancer stem cell: target for anti-cancer therapy. FASEB J. 2007;21:3777–85.
Prince ME, Sivanandan R, Kaczorowski A, Wolf GT, Kaplan MJ, Dalerba P, Weissman IL, Clarke MF, Ailles LE. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proc Natl Acad Sci U S A. 2007;104:973–8.
Godar S, Ince TA, Bell GW, Feldser D, Donaher JL, Bergh J, Liu A, Miu K, Watnick RS, Reinhardt F, et al. Growth-inhibitory and tumor-suppressive functions of p53 depend on its repression of CD44 expression. Cell. 2008;134:62–73.
Mack B, Gires O. CD44s and CD44v6 expression in head and neck epithelia. PLoS One. 2008;3, e3360.
Chen YC, Chen YW, Hsu HS, Tseng LM, Huang PI, Lu KH, Chen DT, Tai LK, Yung MC, Chang SC, et al. Aldehyde dehydrogenase 1 is a putative marker for cancer stem cells in head and neck squamous cancer. Biochem Biophys Res Commun. 2009;385:307–13.
Visus C, Ito D, Amoscato A, Maciejewska-Franczak M, Abdelsalem A, Dhir R, Shin DM, Donnenberg VS, Whiteside TL, DeLeo AB. Identification of human aldehyde dehydrogenase 1 family member A1 as a novel CD8+ T-cell-defined tumor antigen in squamous cell carcinoma of the head and neck. Cancer Res. 2007;67:10538–45.
Lathers DM, Young MR. Increased aberrance of cytokine expression in plasma of patients with more advanced squamous cell carcinoma of the head and neck. Cytokine. 2004;25:220–8.
Yan J, Reichenbach DK, Corbitt N, Hokey DA, Ramanathan MP, McKinney KA, Weiner DB, Sewell D. Induction of antitumor immunity in vivo following delivery of a novel HPV-16 DNA vaccine encoding an E6/E7 fusion antigen. Vaccine. 2009;27:431–40.
Victora GD, Socorro-Silva A, Volsi EC, Abdallah K, Lima FD, Smith RB, Moyses RA, Zarate-Blades CR, Michaluart P, Silva CL, et al. Immune response to vaccination with DNA-Hsp65 in a phase I clinical trial with head and neck cancer patients. Cancer Gene Ther. 2009;16:598–608.
Sewell DA, Pan ZK, Paterson Y. Listeria-based HPV-16 E7 vaccines limit autochthonous tumor growth in a transgenic mouse model for HPV-16 transformed tumors. Vaccine. 2008;26:5315–20.
Davidson EJ, Faulkner RL, Sehr P, Pawlita M, Smyth LJ, Burt DJ, Tomlinson AE, Hickling J, Kitchener HC, Stern PL. Effect of TA-CIN (HPV 16 L2E6E7) booster immunisation in vulval intraepithelial neoplasia patients previously vaccinated with TA-HPV (vaccinia virus encoding HPV 16/18 E6E7). Vaccine. 2004;22:2722–9.
Albarran YCA, de la Garza A, Cruz Quiroz BJ, Vazquez Zea E, Diaz Estrada I, Mendez Fuentez E, Lopez Contreras M, Andrade-Manzano A, Padilla S, Varela AR, Rosales R. MVA E2 recombinant vaccine in the treatment of human papillomavirus infection in men presenting intraurethral flat condyloma: a phase I/II study. BioDrugs. 2007;21:47–59.
Karcher J, Dyckhoff G, Beckhove P, Reisser C, Brysch M, Ziouta Y, Helmke BH, Weidauer H, Schirrmacher V, Herold-Mende C. Antitumor vaccination in patients with head and neck squamous cell carcinomas with autologous virus-modified tumor cells. Cancer Res. 2004;64:8057–61.
Schuler PJ, Harasymczuk M, Visus C, Deleo A, Trivedi S, Lei Y, Argiris A, Gooding W, Butterfield LH, Whiteside TL, Ferris RL. Phase I dendritic cell p53 peptide vaccine for head and neck cancer. Clin Cancer Res. 2014;20:2433–44.
Weise JB, Csiszar K, Gottschlich S, Hoffmann M, Schmidt A, Weingartz U, Adamzik I, Heiser A, Kabelitz D, Ambrosch P, Gorogh T. Vaccination strategy to target lysyl oxidase-like 4 in dendritic cell based immunotherapy for head and neck cancer. Int J Oncol. 2008;32:317–22.
Stephenson RM, Lim CM, Matthews M, Dietsch G, Hershberg R, Ferris RL. TLR8 stimulation enhances cetuximab-mediated natural killer cell lysis of head and neck cancer cells and dendritic cell cross-priming of EGFR-specific CD8+ T cells. Cancer Immunol Immunother. 2013;62:1347–57.
Yewale C, Baradia D, Vhora I, Patil S, Misra A. Epidermal growth factor receptor targeting in cancer: a review of trends and strategies. Biomaterials. 2013;34:8690–707.
Rubin Grandis J, Melhem MF, Gooding WE, Day R, Holst VA, Wagener MM, Drenning SD, Tweardy DJ. Levels of TGF-alpha and EGFR protein in head and neck squamous cell carcinoma and patient survival. J Natl Cancer Inst. 1998;90:824–32.
Bauman JE, Ferris RL. Integrating novel therapeutic monoclonal antibodies into the management of head and neck cancer. Cancer. 2014;120:624–32.
Lopez-Albaitero A, Ferris RL. Immune activation by epidermal growth factor receptor specific monoclonal antibody therapy for head and neck cancer. Arch Otolaryngol Head Neck Surg. 2007;133:1277–81.
Lopez-Albaitero A, Lee SC, Morgan S, Grandis JR, Gooding WE, Ferrone S, Ferris RL. Role of polymorphic Fc gamma receptor IIIa and EGFR expression level in cetuximab mediated, NK cell dependent in vitro cytotoxicity of head and neck squamous cell carcinoma cells. Cancer Immunol Immunother. 2009;58:1853–64.
Dechant M, Weisner W, Berger S, Peipp M, Beyer T, Schneider-Merck T, Lammerts van Bueren JJ, Bleeker WK, Parren PW, van de Winkel JG, Valerius T. Complement-dependent tumor cell lysis triggered by combinations of epidermal growth factor receptor antibodies. Cancer Res. 2008;68:4998–5003.
Dhodapkar MV, Dhodapkar KM, Li Z. Role of chaperones and FcgammaR in immunogenic death. Curr Opin Immunol. 2008;20:512–7.
Taylor C, Hershman D, Shah N, Suciu-Foca N, Petrylak DP, Taub R, Vahdat L, Cheng B, Pegram M, Knutson KL, Clynes R. Augmented HER-2 specific immunity during treatment with trastuzumab and chemotherapy. Clin Cancer Res. 2007;13:5133–43.
Banerjee D, Matthews P, Matayeva E, Kaufman JL, Steinman RM, Dhodapkar KM. Enhanced T-cell responses to glioma cells coated with the anti-EGF receptor antibody and targeted to activating FcgammaRs on human dendritic cells. J Immunother. 2008;31:113–20.
Lee SC, Srivastava RM, Lopez-Albaitero A, Ferrone S, Ferris RL. Natural killer (NK): dendritic cell (DC) cross talk induced by therapeutic monoclonal antibody triggers tumor antigen-specific T cell immunity. Immunol Res. 2011;50:248–54.
Cooper MA, Fehniger TA, Fuchs A, Colonna M, Caligiuri MA. NK cell and DC interactions. Trends Immunol. 2004;25:47–52.
Roda JM, Joshi T, Butchar JP, McAlees JW, Lehman A, Tridandapani S, Carson III WE. The activation of natural killer cell effector functions by cetuximab-coated, epidermal growth factor receptor positive tumor cells is enhanced by cytokines. Clin Cancer Res. 2007;13:6419–28.
Mailliard RB, Son YI, Redlinger R, Coates PT, Giermasz A, Morel PA, Storkus WJ, Kalinski P. Dendritic cells mediate NK cell help for Th1 and CTL responses: two-signal requirement for the induction of NK cell helper function. J Immunol. 2003;171:2366–73.
Lucas M, Schachterle W, Oberle K, Aichele P, Diefenbach A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity. 2007;26:503–17.
Moretta L, Ferlazzo G, Bottino C, Vitale M, Pende D, Mingari MC, Moretta A. Effector and regulatory events during natural killer-dendritic cell interactions. Immunol Rev. 2006;214:219–28.
El-Shami K, Tirosh B, Bar-Haim E, Carmon L, Vadai E, Fridkin M, Feldman M, Eisenbach L. MHC class I-restricted epitope spreading in the context of tumor rejection following vaccination with a single immunodominant CTL epitope. Eur J Immunol. 1999;29:3295–301.
Lee JC, Lee KM, Kim DW, Heo DS. Elevated TGF-beta1 secretion and down-modulation of NKG2D underlies impaired NK cytotoxicity in cancer patients. J Immunol. 2004;172:7335–40.
Yao M, Galanopoulos N, Lavertu P, Fu P, Gibson M, Argiris A, Rezaee R, Zender C, Wasman J, Machtay M, Savvides P. A phase II study of bevacizumab in combination with docetaxel and radiation in locally advanced squamous cell carcinoma of the head and neck. Head Neck. 2015;37(11):1665–71.
Melero I, Hirschhorn-Cymerman D, Morales-Kastresana A, Sanmamed MF, Wolchok JD. Agonist antibodies to TNFR molecules that costimulate T and NK cells. Clin Cancer Res. 2013;19:1044–53.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Kansy, B.A., Lee, S.C., Ferris, R.L. (2016). Immunology of Head and Neck Cancer. In: Bernier, J. (eds) Head and Neck Cancer. Springer, Cham. https://doi.org/10.1007/978-3-319-27601-4_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-27601-4_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-27599-4
Online ISBN: 978-3-319-27601-4
eBook Packages: MedicineMedicine (R0)