
Abstract
The trigeminal nerve is the fifth and thickest cranial nerve and not only it is responsible for facial sensation and motor functioning, but it is also responsible for chemosensory perception. On top of innervating the skin of the face, the trigeminal nerve also innervates the mucosa of the nose and mouth. Here, chemosensory perception starts with the activation of different receptors, the most important and best known ones being the transient receptor potential (GlossaryTerm
TRP
) channels which give rise to sensations such as burning, warmth, coolness, coldness, and pain. From the mucosa, the trigeminal chemosensory information is conveyed through the trigeminal ganglion to the thalamic nuclei in the brain stem; from here fibers project to both, the somatosensory cortex and chemosensory areas of the brain.Most odors stimulate the trigeminal system, in addition to the olfactory system, especially in higher concentrations. However, overlaps between both sensory systems are not limited to the stimulus level, as they interact with each other on peripheral (mucosa) and central (brain) levels. As a consequence, subjects with a lacking olfactory system show lower trigeminal sensitivity and subjects with a lack of trigeminal sensitivity show lower olfactory activations.
Different techniques are available to assess the state and the functionality of the trigeminal system. Such techniques include behavioral assessment by testing participants lacking a sense of smell – in order to exclude olfactory interference – as well as administrating different types of stimuli (pure odorants, pure trigeminal, or a mix of both). More objective measures include electrophysiological methods that evaluate the peripheral and central activations via the negative mucosal potentials (NMPs) and the trigeminal event-related potentials (GlossaryTerm
ERP
s), respectively, as well as functional magnetic resonance imaging, and to a lesser extent, positron emission tomography (GlossaryTermPET
).Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others



Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
The trigeminal nerve is responsible for facial sensations (touch and pain ) and to a considerably lesser degree to motor functioning (masticating). In addition, it also provides chemosensory information from the oral and nasal mucosa, such as the perception of piquancy when eating something spicy. Although a limited amount of research has focused on the chemosensory part of the trigeminal system, this chapter aims at providing an overview over key knowledge on the trigeminal system. Specifically, the anatomy, the different pathways, the various methods of studying the system, the interactions between the olfactory and the trigeminal system as well as the impact of the loss of one system on the other are discussed.
1 General Neuroanatomy of the Trigeminal System
The trigeminal nerve is the fifth cranial nerve (CN V). It is the largest of the 12 cranial nerves; its name stems from the fact that it has three major branches (tri=three; geminus=twin; hence triplets). These three branches are the ophthalmic nerve (CN V1), the maxillary nerve (CN V2), and the mandibulary nerve (CN V3). The trigeminal nerve is primarily a sensory nerve with some motor functions.
1.1 Trigeminal Ganglion, Trigeminal Nuclei
Sensory information from the three branches converges on the trigeminal ganglion (Gasserian ganglion, semilunar ganglion); here, the cell bodies of the incoming sensory fibers are located. The trigeminal ganglion has analogous features as the dorsal root ganglia of the spinal cord. Although the cell bodies of the neurons are located in the trigeminal ganglion, there are no synapses; thus the neurons are called pseudo-unipolar. Somatotopic information from the three branches is preserved [46.1], which means that the relative location of the three branches maintains the same location within the trigeminal ganglion. To illustrate, the branches from the ophthalmic nerve will be placed above the branches from the mandibular nerve as their layout on the human face. From here, afferent fibers enter the brain stem in the angle between pons and the middle cerebellar peduncle [46.2]. In the brain stem, they travel to the trigeminal nuclei, which are arranged from the mesencephalon (mesencephalic nucleus) to pons (main sensory trigeminal nucleus) and the rostral portions of the spinal cord (spinal trigeminal nuclei). It is important to note that the fibers are still arranged somatotopically [46.3]. See Fig. 46.1 for an overview over the trigeminal pathway.


The trigeminal pathway. A: Fibers of the trigeminal nerve (neurons) are located throughout the epithelium of the nasal cavity (gray). B: Cell bodies of the trigeminal neuron are located in the trigeminal ganglion. Nevertheless, no information is transferred between neurons at this stage. C: The first (1) relay station is located in the brainstem (blue). D: The second (2) relay station is located in the thalamus (green). E: Third-order neurons (3) project from the thalamus to the somatosensory regions of the cortex (orange). Note that somatotopic information is preserved throughout the pathway. Purple: cerebellum; light orange: corpus callosum
1.2 Central NervousProcessing Structures
The first synaptic relay is found in the trigeminal nucleus, a structure located in the brainstem . Here the information is transmitted to neurons projecting to the second relay station in the thalamus which is a regulator for consciousness and alertness. Specifically, trigeminal information travels to lateral (ventrobasal) and central (centromedial and parafascicular) thalamic nuclei. From here, the third neuronal projection sends the information mainly to the somatosensory cortex which represents the brain area responsible for sensations, such as touch, pain, or heat. In humans, the ophthalmic regions are represented in the inferior portion of the postcentral gyrus, whereas information from the mandibular nerve is processed in more superior regions in the central sulcus [46.4, 46.5]. Consequently, the relative layout of the three trigeminal branches is preserved in all portions of the trigeminal pathway, from the trigeminal ganglion to the somatosensory cortex.
2 Chemosensory Trigeminal Neuroanatomy
In addition to the facial skin, different branches of CN V innervate the mucosa of nose and mouth. Specifically, the ethmoidal nerves from the ophtalmic nerve and the nasal and some alveolar branches from the maxillary nerve innervate the nasal mucosa; further, other branches of the maxillary nerve innervate the oral mucosa together with the lingual nerve, the buccal nerve, and the alveolar nerves from the mandibular nerve. In the mucosa, the nerve endings of the trigeminal nerve are capable to detect chemosensory information in addition to somatosensory information, via specific chemoreceptors.
2.1 Peripheral Structures, Receptors
On receptor levels, chemical stimuli activate specific receptors of the trigeminal nerve, most of them being ion channels belonging to the subfamily of transient receptor potential (GlossaryTerm
TRP
) receptors, which have been discovered only relatively recently and are discussed in detail in Chap. 34 of this book. One of their main characteristics is that they are activated by both, a certain temperature range and chemical stimuli, such as capsaicin, menthol, camphor, and many more.In addition to the TRP channels, there is also evidence for non-TRP receptors. These receptors are activated by nicotine (nicotinic acetylcholine receptors) [46.6] and acids (proton-gated ion channels) [46.10, 46.7, 46.8, 46.9]. Further, in addition to these receptors on free trigeminal nerve endings solitary chemoreceptor cells have been described in the nasal cavity, although not yet in humans [46.11]. These cells are activated by bitter substances via specific receptors and reach the surface of the nasal epithelium and form synaptic contacts with trigeminal afferent nerve fibers. They may add to the repertoire of compounds that can activate the intranasal trigeminal system.
2.2 Central Nervous Processing Structures
Chemosensory information shares the processing units with this classical somatosensory pathway. However, chemosensory trigeminal stimuli activate brain regions in addition to these somatosensory structures such as the insula, the orbitofrontal cortex, and the piriform cortex [46.12, 46.13, 46.14]. Usually, these additional areas are considered being olfactory and gustatory regions. In order to include trigeminal processing, the term chemosensory areas has been put forward to describe the ensemble of these regions. While most trigeminal stimuli also evoke odorous sensations, that is, have a smell [46.15], it is unclear whether the activation of the chemosensory areas occurs via the olfactory pathway – stimulation of olfactory receptors by the trigeminal stimulus leads to the activation of olfactory nerves – or whether there is a direct and chemosensory-specific connection between the somatosensory pathway and the chemosensory processing areas, with a hitherto unknown exact neuroanatomy. The latter hypothesis is supported by the fact that even the odorless trigeminal stimulus carbon dioxide leads to the activation of chemosensory brain areas [46.12]. In fact, there are some collaterals of the trigeminal nerve that end within the olfactory epithelium; some even re-enter the central nervous system and terminate within the olfactory bulb [46.16]. While the exact function of these fibers is unclear, they may provide one of the links by which the trigeminal and the olfactory system interact.
3 Trigeminal Perception
Compared to the olfactory nerve, chemoreception within the trigeminal system is relatively unspecific. Still, the sensations arising from trigeminal chemosensation are well beyond simple pain perception as trigeminal stimulation may lead to such diverse perceptions as cooling, burning, stinging, or tingling. The TRP channel receptors play a key role in trigeminal perception.
3.1 Receptors and Perception
The first TRP receptor to be discovered and described in detail is the TRPV1 receptor. It is activated by noxious heat above \({\mathrm{43}}\,{\mathrm{{}^{\circ}\mathrm{C}}}\) [46.17] as well as a variety of chemical stimuli, including capsaicin (the spicy ingredient of hot peppers ) [46.17], eugenol (the key compound of essential oil of cloves) [46.36], and acids [46.33]. Stimulation of TRPV1 receptors leads to the perception of a tingling perception, which in higher intensities becomes sharp, burning , and painful [46.44]. There are several other TRPV such as TRPV3. This receptor is activated at lower temperatures starting from \({\mathrm{39}}\,{\mathrm{{}^{\circ}\mathrm{C}}}\) [46.45, 46.46]. This receptor is also chemosensitive as different compounds, such as thymol (a main component of spices such as oregano and thyme) [46.24, 46.26], act as agonists. Activation of TRPV3 is associated with the sensation of warmth [46.26].
On the other side of the physiological temperature range, we find the TRPM8 receptor. This particular receptor is activated by cool temperatures starting from \({\mathrm{39}}\,{\mathrm{{}^{\circ}\mathrm{C}}}\) [46.27], as well as chemical substances such as menthol (main component of peppermint) [46.27], and eucalyptol (eucalyptus) [46.47]. Stimulation of this receptor provides the sensation of cooling without being particularly painful. Further toward the range of noxious cold is the TRPA1 channel, which is activated by cold temperatures below \({\mathrm{17}}\,{\mathrm{{}^{\circ}\mathrm{C}}}\) [46.25, 46.33]. Again, this receptor is chemosensitive as compounds such as allyl thioisocyanate (mustard oil) [46.25, 46.33] serve as agonists. TRPA1 stimulation gives rise to a dull, painful sensation. See Fig. 46.2 for an overview over the temperature ranges of these four receptors. It is interesting to note that many of the stimuli are ingredients of spices and herbs, such as hot pepper, cloves, oregano, thyme, eucalyptus, peppermint, mustard, etc. See Table 46.1 for an overview of the different stimuli that activate the four TRP receptors mentioned in this chapter.


Receptive range of four intranasal TRP receptors. The green area indicates the physiological temperature in the nasopharynx, that is, the temperature to which the nose conditions the inhaled air (after [46.49])
When humans breathe air at room temperature normally, inhaled air is heated within the nasal cavity to an average of 28-\({\mathrm{32}}\,{\mathrm{{}^{\circ}\mathrm{C}}}\) [46.48, 46.49]. This temperature lies between the temperature ranges of TRPM8/TRPA1 on one hand and TRPV3/TRPV1 on the other hand. Thus, none of these receptors should be activated when we breathe clean air at room temperature. Upon stronger inhalation of air at room temperature, however, the TRPM8 receptor may be activated, since intranasal temperature drops [46.49]. Further, changes of the air temperature induce receptor potentials and, as a consequence, the perception of cold, cool, warm, or hot. Similarly, the presence of one of the multiple trigeminal stimuli may induce receptor potentials evoking according sensations even in the absence of a real temperature change. In fact, the inhalation of menthol does lead to the perception of coolness and, by analogy, the sensation of an increased nasal patency. Objective measures of nasal air temperature and nasal patency however show that menthol inhalations do not affect any of both measures [46.48].
We do not yet know how different trigeminal receptors are distributed within the nasal epithelium. In this context, it is interesting to note that three different trigeminal stimuli, which most likely bind to different receptors, evoked comparable activation patterns at different sites within the nasal epithelium [46.50]. Therefore, different areas of the nasal cavity seem to exhibit distinct concentrations of receptors, but the ratios between different receptors seem to be stable throughout the nasal cavity.
4 Assessment of the Trigeminal System
Different methods are available to assess the sensitivity of trigeminal system in humans in vivo. One challenge a researcher faces is the fact that olfactory and trigeminal system are intimately related. Specifically, most trigeminal stimuli activate the olfactory system in addition to the trigeminal system, and they do so at lower concentrations [46.51]. Therefore, although the intensity of the trigeminal sensation is usually stronger than the olfactory one, there usually is some form of interference from the olfactory input. Several methods have been used to overcome this problem, each of which has its own advantages and disadvantages.
4.1 Behavioral Methods
Different methods are available to assess sensitivity in the trigeminal system with behavioral measures. In order to avoid the effects of olfactory interference, one could either test participants with anosmia, i. e., lack of olfactory perceptions, or focus on tests which rely more or less exclusively on trigeminal sensations.
4.1.1 Behavioral Testing in Anosmia
Anosmia, acquired or congenital lack of olfactory function is a relatively common finding as 1 in 20 is affected in the general population [46.52, 46.53]. Of course, persons with anosmia do not perceive the olfactory component of mixed olfactory-trigeminal stimuli; thus minimizing olfactory interference. In fact, persons with anosmia can only detect mixed olfactory-trigeminal stimuli by virtue of the test compound’s trigeminal component. This can be used to assess the trigeminal impact of a substance, since the percentage of subjects with anosmia who detect a stimulus is correlated with the stimulus’ trigeminal impact [46.15, 46.54]. The typical method to assess olfactory thresholds in persons with normal olfactory function is to ask participants to detect an odorous substance in different concentrations amongst several odorless blanks. If the same stimulus with the same paradigm is used in subjects with anosmia the resulting threshold will be based on the trigeminal sensation [46.55, 46.56], thus providing a method to assess trigeminal thresholds.
However, one has to be careful when generalizing the findings from persons with anosmia to the general population. We know that anosmia is associated with a reduced trigeminal sensitivity [46.57, 46.58, 46.59, 46.60]. The difference in trigeminal sensitivity between subjects with anosmia and participants with a normal sense of smell may be small, and therefore not be detected when investigating only a small number of subjects [46.55, 46.56, 46.61], but is a clear limiting factor of this approach.
4.1.2 Tests Depending on Trigeminal Stimulation
In addition to testing subjects who cannot perceive olfactory information, olfactory interference can be excluded in those tests, which depend solely on trigeminal perception. One exemplary method is the lateralization task, in which subjects are asked to identify the nostril to which a stimulus has been delivered (lateralization task). This task is based on the fact that humans are unable to localize pure odorants [46.62, 46.63], but can do so for stimuli which activate the trigeminal system, mixed olfactory/trigeminal stimuli. Depending on the trigeminal impact [46.44], such stimuli can be localized with high accuracy [46.51, 46.62, 46.64, 46.65, 46.66, 46.67]. Using this fact allows for two different tasks: first, one can determine the sensitivity of the trigeminal system in a given population or compare sensitivity between two groups of subjects [46.66, 46.68]; second, one can determine the trigeminal potency of different chemical compounds [46.44, 46.56]. Thus, by using the lateralization test, one can determine the threshold for a given substance, by using a staircase procedure [46.51]. Such a procedure is often used as an experimental design to assess the thresholds by lowering or elevating the task difficulty until we find the minimally intense stimuli to which the participant would respond accurately. Alternatively, a single concentration of a test compound with strong trigeminal impact may be used in a semiquantitative screening protocol. Here, the sum of correct identifications is used for further statistical analyses. The latter is mostly used in a clinical setting. The lateralization test can be cumbersome, since it is important to keep a relatively long inter stimulus interval of at least 40 s in order to avoid habituation [46.69]. An alternative method is to register response times in a lateralization task [46.70], which has the advantage to provide parametric data with fewer trials and thus shorter test times.
As an alternative, some groups take advantage of the fact that the cornea and conjunctiva of the eye are also innervated by the trigeminal nerve. These epithelia, in fact, allow for the perception of painful sensations, but do not respond to pure olfactory stimuli. Thus, vapor-phase chemical stimuli which evoke the sensations of burning or stinging in the cornea can be used to assess trigeminal irritation thresholds in the eye. However, one has to be careful to avoid co-stimulation of the nose and thus the olfactory system. Corneal thresholds can be used as estimates of trigeminal sensitivity since irritation thresholds obtained in the eye and nose are significantly correlated [46.71].
Another approach is to instruct subjects with a normal olfactory function to distinguish between olfactory and trigeminal sensations. Therefore, after being trained to focus on trigeminal sensations subjects learn to disregard the simultaneous olfactory sensation. Typically, subjects receive instructions such as have you felt any sensation like burning, stinging, cooling, or tickling? By using this method, one can assess trigeminal perception thresholds [46.58, 46.72, 46.73, 46.74]. However, one has to keep in mind that olfactory interference will occur and can unconsciously affect subjects’ responses, which limits the application of this method.
Another method rules out olfactory interference, by using pure trigeminal stimuli, stimuli which exclusively activate the trigeminal system. However, a possible concomitant olfactory stimulation is very difficult to exclude. Only few stimuli are available in this category; they include CO2 or capsaicin. Here it is important to note that CO2 acts as a trigeminal stimulus only in very high concentrations \((> {\mathrm{100000}}\,{\mathrm{ppm}})\) [46.75], which may be dangerous for participants’ safety. Thus, CO2 as a nasal trigeminal stimulus can safely be employed only as brief \((<{\mathrm{3}}\,{\mathrm{s}})\) stimulus, or alternatively with mouth breathing and velopalatine closure (isolation of the nasal cavity).
4.2 Electrophysiological Methods
In contrast to behavioral techniques, electrophysiological measures rely less on participants’ response and collaboration. They therefore provide a more objective assessment of trigeminal sensitivity. Still, the problem of olfactory interference is difficult to avoid.
4.2.1 Event-Related Potentials
Trigeminal ERP are electroencephalography (GlossaryTerm
EEG
)-derived polyphasic signals obtained at the surface of the scalp [46.76] due to the activation of cortical neurons that generate electromagnetic fields. In other words, trigeminal ERP are a central nervous representation of the processing of trigeminally mediated sensations. The EEG is a noisy signal containing activity from many cortical neurons; therefore ERP have to be extracted from the background activity; this can be done by averaging responses to single stimuli reducing the random background noise [46.77]. Again, single stimulations have to be separated by a relatively long inter-stimulus interval of at least 30-40 s to avoid effects of habituation [46.69]. Further, in order to obtain meaningful averages with an acceptable signal-to-noise ratio, at least 10 single responses have to be recorded resulting in ERP sessions of 45 min to 2 h. This requires subjects’ vigilance to be stabilized by simple tasks, such as a tracking task on a computer screen [46.78]. The main advantage of ERP is the very high temporal resolution in the range of milliseconds. However, this comes with a relatively poor spatial resolution. Further, ERP are prone to artifacts; therefore successful ERP recording requires an olfactometer, a device which allows for the delivery of stimuli with:-
1.
A sharp onset
-
2.
Exactly defined duration
-
3.
Without concomitant mechanical co-stimulation
-
4.
Without concomitant thermal co-stimulation [46.76].
The nomenclature of the trigeminal ERP responses follows that of other sensory domains; a small first positive peak (P1) typically occurs at latencies later than 200 ms, followed by a first major negative peak (N1; approximately 400 ms), and the late positive complex (P2 or P2/P3; approximately 650 ms) [46.77, 46.78] (Fig. 46.3 for an overview). Largest responses are obtained from central and parietal electrodes; measures of interest are usually amplitudes and latencies of the major peaks. ERP are mostly used in a research setting, since they require a relatively high effort in terms of cost and time. Further, the discussion on olfactory interference also applies to trigeminal ERP: since most trigeminal stimuli also activate the olfactory system, trigeminal ERP will in most cases also contain signals from olfactory processing areas.


Procedure of trigeminal ERP recording: (a) Single EEG recordings in response to a 200 ms stimulus (shaded area indicates stimulus). Note that response to the stimulus is overlapped by random brain activity. In order to extract specific response, several trials are recorded (t1 to t n ). (b) EEG response are averaged. (c) Event-related potential is visible after averaging. The most important peaks, N1 and P2 are indicated by crosshair. For each peak one can analyze latency (blue; N1 and P2), and amplitude (red; baseline to peak amplitude: N1 and P2; peak-to-peak amplitude: N1P2)
4.2.2 Negative Mucosal Potential
Another electrophysiological procedure is relatively free from olfactory interference: the NMP is measured on the level of the nasal mucosa and thus the periphery of the trigeminal system [46.79, 46.80, 46.81, 46.82, 46.83]. The NMP is the summating receptor potentials of chemoreceptors of the trigeminal nerve [46.84] and thus an electrophysiological correlate of trigeminal activation of the nasal respiratory epithelium [46.79, 46.80, 46.81, 46.82, 46.83]. As such it is independent from olfactory stimulation. NMPs are recorded by means of an electrode placed on the respiratory mucosa [46.85]; electrode placement should be done under endoscopical control [46.86]. Again, the signal is averaged, but due to the lower background noise, fewer recordings are needed to obtain a meaningful NMP. In fact, one single recording may be enough for interpretation. Similar to ERP, NMP recording requires an olfactometer for stimulus presentation to assure a sharp stimulus onset and exactly defined duration and to avoid concomitant mechanical and thermal co-stimulation. NMP consist of a slow negative wave with a latency of approximately 1000 ms [46.87]. The largest NMP indicating the highest sensitivity is observed at the nasal septum, lowest on the nasal floor and the olfactory cleft [46.50, 46.88]. See Fig. 46.4 for an overview over electrophysiological recording sites.


Electrophysiological recording sites: event-related potentials are EEG derived signals and recorded from the surface of the skull (red), central responses; negative mucosal potentials are mucosa-generated responses and recorded from the epithelium (blue), peripheral responses
4.3 Brain Imaging
Over the last three decades, several techniques have been made available to neuroscientists, which allow for the investigation of brain structure and brain function in vivo. The most commonly used technique is functional magnetic resonance imaging (GlossaryTerm
MRI
), which measures the ratio of oxygenated hemoglobin/deoxygenated hemoglobin in the brain. From this one can infer the activation level of different brain areas (neurovascular coupling). Specifically, researchers can observe which areas of the brain are activated by different tasks. Functional MRI permits the investigation of brain activation with a relatively high spatial resolution. Still, a voxel – the 3D analog of a pixel – typically has the size of \({\mathrm{27}}\,{\mathrm{mm^{3}}}\) (3 mm × 3 mm × 3 mm); thus each voxel contains the information of the average activation of ten thousands of neurons. Functional MRI has been used to explore sensory processing; in some studies the trigeminal system has been examined with this technique. Functional MRI has shown that chemosensory trigeminal stimulation leads to the activation patterns which only partly overlap with somatosensory stimuli (brainstem [46.14], thalamus [46.13], SI/SII [46.89], anterior cingulate [46.14]). However, chemosensory trigeminal stimuli also activate olfactory regions, piriform, orbitofrontal, and insular cortex [46.12, 46.14, 46.90]. It is worth mentioning that especially orbitofrontal areas are susceptible to image distortion in MRI due to the proximity of the nasal sinuses making the examination of this chemosensory area somewhat difficult.Other brain imaging techniques can be used to investigate the trigeminal system, but have so far only been used in a few studies. One of these techniques, PET assesses the concentration of radioactive markers. Importantly, it allows for imaging of the orbitofrontal areas of the brain free of distortion; however, this comes with a weaker spatial and temporal resolution compared to functional MRI. PET-based experiments reported additional activation in the amygdala, claustrum, and lateral hypothalamus due to trigeminal chemosensory stimulation [46.91].
5 Interactions Between Olfaction and Trigeminal System
As mentioned earlier, most stimuli activate the trigeminal system as well as the olfactory system [46.51, 46.92, 46.93]. Additionally, psychophysical and electrophysiological findings report that these two systems interact [46.93], by suppression and enhancing each other [46.92, 46.94, 46.95, 46.96, 46.97, 46.98]. It is hypothesized that this interaction occurs at three levels:
-
1.
On the stimulus level
-
2.
In the periphery (mucosa)
-
3.
On central levels (brain; Fig. 46.5) [46.92].
Fig. 46.5 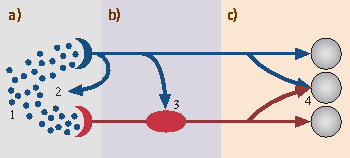
Sites of interaction between the trigeminal system (blue) and the olfactory system (red). (a) Mucosal level: 1. The same molecules can stimulate both, the olfactory system and the trigeminal system; 2. Trigeminal reflexes may alter nasal patency. (b) Olfactory bulb: 3. Some trigeminal nerve endings terminate in the olfactory bulb. (c) Central nervous system: 4. The trigeminal system and the olfactory system share central processing units


In order to understand this interaction, it is necessary to evaluate both sensory systems separately. However the fact that they are usually stimulated together raises a serious methodological challenge. One possibility to overcome this problem is to investigate the trigeminal function in subjects without a working sense of smell [46.93].
5.1 Stimuli
Subjects with anosmia are able to detect a large number of volatile chemicals despite the lack of functionality of the olfactory nerve, suggesting that stimuli perception was processed by the trigeminal nerve [46.15]. Moreover, patients with olfactory dysfunction also demonstrate lower trigeminal sensitivity which highlights the interdependence of the two systems [46.100, 46.99]. Additionally, healthy subjects have higher trigeminal thresholds compared to olfactory ones suggesting that it takes a higher concentration of an odorant to stimulate the trigeminal nerve as compare to the olfactory nerve [46.51]. Thus, most odorants also activate the trigeminal system. It should be noted however that although most odorants activate both, the trigeminal and the olfactory nerves, there are a few exceptions. For instance, CO2 activates the trigeminal system with little or no concomitant olfactory stimulation [46.101, 46.102, 46.103]. On the other hand only a few odorants activate the olfactory nerve selectively with little activation of the trigeminal nerve [46.15] such as vanillin [46.104] decanoic acid [46.15], hydrogen sulfide \(\mathrm{(H_{2}S)}\) [46.104] and phenylethyl alcohol (GlossaryTerm
PEA
) [46.51, 46.63].Further, most irritants that stimulate the trigeminal system can activate the olfactory system as well. This has been shown indirectly, as trigeminal stimulation activates regions typically involved in the olfactory processing such as piriform cortex [46.105, 46.12], the anterior orbitofrontal cortex [46.106, 46.107, 46.108, 46.12], rostral insula [46.109, 46.12], and the superior temporal gyrus [46.110, 46.12]. In addition, CO2, which is usually considered to be a selective trigeminal stimulus, recruited the olfactory pathways in low concentrations [46.111]. Moreover, roughly 10 % of piriform cortex neurons responded to both, olfactory and trigeminal stimuli, further highlighting the overlap between the two systems. Altogether, these studies show that both sensory systems exhibit a large extent of promiscuity already on the level of the stimuli.
5.2 Indirect Interaction
On top of an interaction at the level of the stimuli, there is also an indirect interaction between the trigeminal and the olfactory systems. For instance, trigeminal reflexes trigger alteration of nasal patency and respiration as well as changes in the mucus covering the epithelium as a result of the stimulation of glands and secretory cells [46.112]. Additionally, electrophysiological studies indicate that odors can modify, via local axon reflex, the spontaneous activity of olfactory receptor cells which triggers the release of different peptides as well as analgesic effects from trigeminal fibers innervating the olfactory epithelium resulting in a modification of odor perception [46.113, 46.114, 46.115]. Putting together, these studies indicate that other substances can influence the olfactory and the trigeminal systems [46.93].
5.3 Peripheral Interaction
In addition to a complete loss of olfaction, subjects with anosmia also have a lower trigeminal function, when assessed with behavioral measures [46.100, 46.103, 46.116, 46.117, 46.93]. On peripheral levels however, subjects with anosmia exhibit larger NMP, which indicated increased peripheral susceptibility [46.68, 46.87]. Thus, a model with mixed sensory adaptation/compensation was put forward to explain the interaction between the two systems at a peripheral level. Some neuroanatomical studies provide underpinnings for these interactions. For example, axons of the trigeminal nerve re-enter the central nervous system (GlossaryTerm
CNS
) and terminate in the glomerular layer of the olfactory bulb [46.118, 46.16]. They may be activated by the lateral excitatory network within the olfactory glomeruli [46.119] of a working olfactory system, which is associated with a reduced peripheral responsiveness [46.74]. In the case of olfactory loss, however, this excitatory network is no longer activated; as a consequence, the intrabulbar trigeminal collaterals become disinhibited, resulting in an increased activation of the trigeminal system at peripheral levels [46.93]. In other words, the lack of a functioning olfactory system leads to the disinhibition of the intrabulbar trigeminal collaterals which results in higher NMP activations. This type of mechanism is also found in the gustatory system in which input from the chorda tympani inhibits that of the glossopharyngeal nerve. Damage to the chorda tympani abolishes this inhibition which increases the input from areas innervated by the glossopharyngeal nerve [46.120].5.4 Central Interaction
Studies on patients with anosmia also revealed interactions of the two systems at central levels. When evaluating such interaction at central levels, electrophysiological measures are particularly useful. Trigeminal event-related potentials (GlossaryTerm
tERP
s) are electrophysiological responses generated by the cortex. Compared to controls, subjects with acquired anosmia exhibited smaller tERPs [46.87], whereas those with congenital anosmia presented similar activations as controls [46.68]. Moreover, low concentrations of isopentyl acetate (an odor with trigeminal components), triggered odor-like responses in the piriform cortex of mice whereas at high concentrations triggered trigeminal-like responses providing support that some neurons of the piriform cortex can be modulated to respond to both types of stimuli [46.121]. Additionally, the duration of olfactory dysfunction and tERP amplitudes are positively correlated [46.59] suggesting that anosmia-triggered reduction of trigeminal sensitivity could improve over time. Therefore, these studies advocate that on one hand, a fully functional trigeminal system relies on the functionality of the olfactory system [46.122, 46.123, 46.58, 46.59, 46.60, 46.68], and on the other hand, longer the period of time of olfactory loss, better the adaptive mechanisms. FMRI studies provide further evidence as to the key areas of central interaction between olfactory and trigeminal stimulation [46.12, 46.90]. While each sensory system has unique central processing areas, they share a considerable amount of central processing areas. For example, activation of brainstem, ventrolateral posterior thalamus, anterior cingulate, precentral gyrus, and somatosensory areas are commonly observed following trigeminal stimulation, but not after exposure to odors [46.14]. In turn, activation of amygdala and the ventral putamen is a typical feature of olfactory but not of nonpainful trigeminal stimulation [46.124]. However, both trigeminal and olfactory stimulation activate piriform cortex, the orbitofrontal cortex and the rostral insula [46.124, 46.14]. Thus, these are the prime candidates on the quest for the areas of interaction between the trigeminal and the olfactory system. In fact, patients with anosmia exhibited, when compared to controls, less activations in, amongst other regions, the orbitofrontal cortex and the insula, but also primary somatosensory cortex [46.89]. This may help explaining smaller tERP amplitudes and higher trigeminal thresholds in this patient group [46.93]. In healthy subjects a mixture of CO2 (trigeminal stimuli) and PEA (pure odorant) led to higher activations than the sum of activation of CO2 and PEA presented independently, in both, chemosensory areas (orbitofrontal cortex) and multisensory integration centers (intraparietal sulcus) [46.125].Putting together, these imaging studies suggest a great overlap between the two chemical senses highlighting the intimate connections between the two systems.
6 Clinical Aspects of the Trigeminal System
Different factors may affect trigeminal sensitivity. The most prominent amongst them is aging as older people consistently exhibit lower trigeminal sensitivity [46.123, 46.83]. Additionally, different diseases and olfactory dysfunction also impact the trigeminal function [46.59]. As previously mentioned, the olfactory and trigeminal systems interact with one another at different levels and therefore examining a clinical population with olfactory and trigeminal dysfunction could shed some light on the co-dependence of the two systems.
6.1 Olfactory Dysfunction and Trigeminal Perception
Hyposmia (a reduced sense of smell) and anosmia are relatively common in the normal population. It is estimated that around 15 % of the population exhibit hyposmia and 5 % suffer from anosmia [46.52, 46.53]. Generally, typical causes of hyposmia and anosmia include sinunasal disease, head trauma, upper respiratory tract infections, or neurodegenerative diseases whereas congenital anosmia occurs only in around 2 % of those with anosmia [46.126, 46.127, 46.128]. Anosmia and hyposmia are associated with lower trigeminal sensitivity when tested with several different techniques. Specifically, subjects with anosmia exhibit less changes in their breathing pattern when inhaling airborne chemicals [46.122, 46.129], have lower intensity ratings of trigeminal stimuli [46.60], exhibit higher thresholds to irritants [46.130, 46.58], and have fewer correct answers on lateralization tests [46.123, 46.62]. Lower trigeminal chemosensitivity seems to be a general feature of acquired anosmia and independent of its etiology [46.123], while congenital anosmia is somewhat different from acquired anosmia. As a matter of fact, congenital anosmia is associated with similar behavioral measures of trigeminal sensitivity as controls [46.68]. Interestingly, somatosensory measures (such as responsiveness to touch) are not affected by olfactory dysfunction [46.57]. Further, peripheral susceptibility seems to be larger in patients with anosmia (congenital or acquired), in sharp contrast to the findings from central and behavioral measures [46.93].
The interactions between anosmia and the trigeminal system seem not be stable over time: the duration of the olfactory loss and the functionality of the trigeminal system are correlated as the latter gets better over time [46.87]. Further, those 10-30 % [46.131] of cases of anosmia which show recovery of olfactory function actually exhibit even larger peripheral responses than those without recovery [46.87] suggesting additional complex mechanisms taking place. Puttting together, these studies show that acquired anosmia is associated with lower trigeminal sensitivity, while anosmia in general is associated with increased peripheral responsiveness in the trigeminal system.
6.2 Trigeminal Dysfunction and Olfactory Perception
As previously mentioned, the trigeminal nerve is the thickest of the cranial nerve which makes it more resistant to trauma compared to the olfactory nerve [46.87]. Consequently, cases in which the trigeminal nerve is damaged are very scarce. In one experiment, local anaesthesia of the middle nasal meatus resulted in an elevation in detection thresholds of n-butanol suggesting that blocking the trigeminal system results in a reduced olfactory function [46.132]. In another report, two subjects with a total unilateral destruction of the trigeminal nerve due to the removal of an acoustic neuroma, reported lower odor intensity ratings compared to controls with normal trigeminal function [46.133]. In another case study, a women who, as a result of a meningioma, had a loss of trigeminal function on her left side, exhibited lower olfactory function on the affected side [46.103]. Specifically, NMPs responses, tERPs responses to \(\mathrm{H_{2}S}\) (pure odorant) as well as to CO2 (trigeminal irritant) were reduced on the left side compared to the right side. Additionally, pure odorant detection threshold was 64 times higher on the left side compare to the right side. Puttting together, a damaged trigeminal system may have an impact on olfactory perception highlighting once more the interdependence of the two systems at different levels of odor processing.
Abbreviations
- CNS:
-
central nervous system
- EEG:
-
electroencephalography
- ERP:
-
event-related potential
- MRI:
-
magnetic resonance imaging
- PEA:
-
phenylethylalcohol
- PET:
-
positron emission tomography
- tERP:
-
trigeminal event-related potential
- TRP:
-
transient receptor potential
References
D. Borsook, A.F. DaSilva, A. Ploghaus, L. Becerra: Specific and somatotopic functional magnetic resonance imaging activation in the trigeminal ganglion by brush and noxious heat, J. Neurosci. 23, 7897–7903 (2003)
A. Waldeyer, A. Mayet, D. Keyserling: Anatomie des Menschen (Walter de Gruyter, Berlin, New York 1993)
W. Firbas, H. Gruber, R. Mayr, M. Tschabitscher: Neuroanatomie (Wilhelm Maudrich, Wien, München, Berlin 1988)
D. Borsook, R. Burstein, L. Becerra: Functional imaging of the human trigeminal system: Opportunities for new insights into pain processing in health and disease, J. Neurobiol. 61, 107–125 (2004)
A.F. DaSilva, L. Becerra, N. Makris, A.M. Strassman, R.G. Gonzalez, N. Geatrakis, D. Borsook: Somatotopic activation in the human trigeminal pain pathway, J. Neurosci. 22, 8183–8192 (2002)
N. Thuerauf, M. Kaegler, R. Dietz, A. Barocka, G. Kobal: Dose-dependent stereoselective activation of the trigeminal sensory system by nicotine in man, Psychopharmacology (Berl) 142, 236–243 (1999)
R. Waldmann, G. Champigny, F. Bassilana, C. Heurteaux, M. Lazdunski: A proton-gated cation channel involved in acid-sensing, Nature 386, 173–177 (1997)
C.C. Chen, S. England, A.N. Akopian, J.N. Wood: A sensory neuron-specific, proton-gated ion channel, Proc. Natl. Acad. Sci. USA 95, 10240–10245 (1998)
J.R. de Weille, F. Bassilana, M. Lazdunski, R. Waldmann: Identification, functional expression and chromosomal localisation of a sustained human proton-gated cation channel, FEBS Letter 433, 257–260 (1998)
K. Babinski, K.T. Le, P. Seguela: Molecular cloning and regional distribution of a human proton receptor subunit with biphasic functional properties, J. Neurochem. 72, 51–57 (1999)
T.E. Finger, B. Böttger, A. Hansen, K.T. Anderson, H. Alimohammadi, W.L. Silver: Solitary chemoreceptor cells in the nasal cavity serve as sentinels of respiration, Proc. Natl. Acad. Sci. USA 100, 8981–8986 (2003)
J.A. Boyle, M. Heinke, J. Gerber, J. Frasnelli, T. Hummel: Cerebral activation to intranasal chemosensory trigeminal stimulation, Chem. Senses 32, 343–353 (2007)
E. Iannilli, C. Del Gratta, J.C. Gerber, G.L. Romani, T. Hummel: Trigeminal activation using chemical, electrical, and mechanical stimuli, Pain 139, 376–388 (2008)
J. Albrecht, R. Kopietz, J. Frasnelli, M. Wiesmann, T. Hummel, J.N. Lundstörm: The neuronal correlates of intranasal trigeminal function-an ALE meta-analysis of human functional brain imaging data, Brain Res. Rev. 62, 183–196 (2010)
R.L. Doty, W.P.E. Brugger, P.C. Jurs, M.A. Orndorff, P.J. Snyder, L.D. Lowry: Intranasal trigeminal stimulation from odorous volatiles: Psychometric responses from anosmic and normal humans, Physiol. Behav. 20, 175–185 (1978)
M.L. Schaefer, B. Bottger, W.L. Silver, T.E. Finger: Trigeminal collaterals in the nasal epithelium and olfactory bulb: A potential route for direct modulation of olfactory information by trigeminal stimuli, J. Comp. Neurol. 444, 221–226 (2002)
M.J. Caterina, M.A. Schumacher, M. Tominaga, T.A. Rosen, J.D. Levine, D. Julius: The capsaicin receptor: A heat-activated ion channel in the pain pathway, Nature 389, 816–824 (1997)
P.M. Richards, E.C. Johnson, W.L. Silver: Four irritating odorants target the trigeminal chemoreceptor TRPA1, Chemosens. Percept. 3, 190–199 (2010)
W.L. Silver, T.R. Clapp, L.M. Stone, S.C. Kinnamon: TRPV1 receptors and nasal trigeminal chemesthesis, Chem. Senses 31, 807–812 (2006)
D.M. Bautista, P. Movahed, A. Hinman, H.E. Axelsson, O. Sterner, E.D. Högestätt, D. Julius, S.E. Jordt, P.M. Zygmunt: Pungent products from garlic activate the sensory ion channel TRPA1, Proc. Natl. Acad. Sci. USA 102, 12248–12252 (2005)
M.M. Salas, K.M. Hargreaves, A.N. Akopian: TRPA1-mediated responses in trigeminal sensory neurons: Interaction between TRPA1 and TRPV1, Eur. J. Neurosci. 29, 1568–1578 (2009)
L.J. Macpherson, B.H. Geierstanger, V. Viswanath, M. Bandell, S.R. Eid, S. Huang, A. Patapoutian: The pungency of garlic: Activation of TRPA1 and TRPV1 in response to allicin, Curr. Biol. 15, 929–934 (2005)
C. Saunders, W.Y. Li, T.D. Patel, J.A. Muday, W.L. Silver: Dissecting the role of TRPV1 in detecting multiple trigeminal irritants in three behavioral assays for sensory irritation, F1000Res. (2013), eCollection 2013 doi:10.12688/f1000research.2-74.v1
A.K. Vogt-Eisele, K. Weber, M.A. Sherkheli, G. Vielhaber, J. Panten, G. Gisselmann, H. Hatt: Monoterpenoid agonists of TRPV3, Br. J. Pharmacol. 151, 530–540 (2007)
M. Bandell, G.M. Story, S.W. Hwang, V. Viswanath, S.R. Eid, M.J. Petrus, T.J. Earley, A. Patapoutian: Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin, Neuron 41, 849–857 (2004)
H. Xu, M. Delling, J.C. Jun, D.E. Clapham: Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels, Nat. Neurosci. 9, 628–635 (2006)
D.D. McKemy, W.M. Neuhausser, D. Julius: Identification of a cold receptor reveals a general role for TRP channels in thermosensation, Nature 416, 52–58 (2002)
L.J. Macpherson, S.W. Hwang, T. Miyamoto, A.E. Dubin, A. Patapoutian, G.M. Story: More than cool: Promiscuous relationships of menthol and other sensory compounds, Mol. Cell Neurosci. 32, 335–343 (2006)
H.X. Xu, N.T. Blair, D.E. Clapham: Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism, J. Neurosci. 25, 8924–8937 (2005)
T. Selescu, A.C. Ciobanu, C. Dobre, G. Reid, A. Babes: Camphor activates and sensitizes transient receptor potential melastatin 8 (TRPM8) to cooling and icilin, Chem. Senses 38, 563–575 (2013)
M.A. Sherkheli, H. Benecke, J.F. Doerner, O. Kletke, A.K. Vogt-Eisele, G. Gisselmann, H. Hatt: Monoterpenoids induce agonist-specific desensitization of transient receptor potential vanilloid-3 (TRPV3) ion channels, J. Pharm. Pharm. Sci. 12, 116–128 (2009)
A. Moqrich, S.W. Hwang, T.J. Earley, M.J. Petrus, A.N. Murray, K.S. Spencer, M. Andahazy, G.M. Story, A. Patapoutian: Impaired thermosensation in mice lacking TRPV3, a heat and camphor sensor in the skin, Science 307, 1468–1472 (2005)
S.E. Jordt, D.M. Bautista, H.H. Chuang, D.D. McKemy, P.M. Zygmunt, E.D. Högestätt, I.D. Meng, D. Julius: Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1, Nature 427, 260–265 (2004)
A.N. Akopian, N.B. Ruparel, N.A. Jeske, A. Patwardhan, K.M. Hargreaves: Role of ionotropic cannabinoid receptors in peripheral antinociception and antihyperalgesia, Trends Pharmacol. Sci. 30, 79–84 (2009)
S.C. Stotz, J. Vriens, D. Martyn, J. Clardy, D.E. Clapham: Citral sensing by transient [corrected] receptor potential channels in dorsal root ganglion neurons, PLoS ONE 3, e2082 (2008)
B.H. Yang, Z.G. Piao, Y.B. Kim, C.H. Lee, J.K. Lee, K. Park, J.S. Kim, S.B. Oh: Activation of vanilloid receptor 1 (VR1) by eugenol, J. Dent. Res. 82, 781–785 (2003)
C.R. McNamara, J. Mandel-Brehm, D.M. Bautista, J. Siemens, K.L. Deranian, M. Zhao, N.J. Hayward, J.A. Chong, D. Julius, M.M. Moran, C.M. Fanger: TRPA1 mediates formalin-induced pain, Proc. Natl. Acad. Sci. USA 104, 13525–13530 (2007)
L.J. Macpherson, B. Xiao, K.Y. Kwan, M.J. Petrus, A.E. Dubin, S. Hwang, B. Cravatt, D.P. Corey, A. Patapoutian: An ion channel essential for sensing chemical damage, J. Neurosci. 27, 11412–11415 (2007)
M. Lubbert, J. Kyereme, N. Schobel, L. Beltran, C.H. Wetzel, H. Hatt: Transient receptor potential channels encode volatile chemicals sensed by rat trigeminal ganglion neurons, PLoS ONE 8, e77998 (2013)
M. Trevisani, J. Siemens, S. Materazzi, D.M. Bautista, R. Nassini, B. Campi, N. Imamachi, E. Andrè, R. Patacchini, G.S. Cottrell, R. Gatti, A.I. Basbaum, N.W. Bunnett, D. Julius, P. Geppetti: 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1, Proc. Natl. Acad. Sci. USA 104, 13519–13524 (2007)
A. Moussaieff, N. Rimmerman, T. Bregman, A. Straiker, C.C. Felder, S. Shoham, Y. Kashman, S.M. Huang, H. Lee, E. Shohami, K. Mackie, M.J. Caterina, J.M. Walker, E. Fride, R. Mechoulam: Incensole acetate, an incense component, elicits psychoactivity by activating TRPV3 channels in the brain, FASEB Journal 22, 3024–3034 (2008)
Y. Karashima, N. Damann, J. Prenen, K. Talavera, A. Segal, T. Voets, B. Nilius: Bimodal action of menthol on the transient receptor potential channel TRPA1, J. Neurosci. 27, 9874–9884 (2007)
G.P. Ahern, X.B. Wang, R.L. Miyares: Polyamines are potent ligands for the capsaicin receptor TRPV1, J. Biol. Chem. 281, 8991–8995 (2006)
J. Frasnelli, J. Albrecht, B. Bryant, J.N. Lundstrom: Perception of specific trigeminal chemosensory agonists, Neuroscience 189, 377–383 (2011)
G.D. Smith, J. Gunthorpe, R.E. Kelsell, P.D. Hayes, P. Reilly, P. Facer, J.E. Wright, J.C. Jerman, J.P. Walhin, L. Ooi, J. Egerton, K.J. Charles, D. Smart, A.D. Randall, P. Anand, J.B. Davis: TRPV3 is a temperature-sensitive vanilloid receptor-like protein, Nature 418, 186–190 (2002)
A.M. Peier, A.J. Reeve, D.A. Andersson, A. Moqrich, T.J. Earley, A.C. Hergarden, G.M. Story, S. Colley, J.B. Hogenesch, P. McIntyre, S. Bevan, A. Patapoutian: A heat-sensitive TRP channel expressed in keratinocytes, Science 296, 2046–2049 (2002)
H.J. Behrendt, T. Germann, C. Gillen, H. Hatt, R. Jostock: Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay, Br. J. Pharmacol. 141, 737–745 (2004)
J. Lindemann, E. Tsakiropoulou, M.O. Scheithauer, I. Konstantinidis, K.M. Wiesmiller: Impact of menthol inhalation on nasal mucosal temperature and nasal patency, Am. J. Rhinol. 22, 402–405 (2008)
P. Rouadi, F.M. Baroody, D. Abbott, E. Naureckas, J. Solway, R.M. Naclerio: A technique to measure the ability of the human nose to warm and humidify air, J. Appl. Physiol. 87(1999), 400–406 (1985)
M. Scheibe, C. van Thriel, T. Hummel: Responses to trigeminal irritants at different locations of the human nasal mucosa, Laryngoscope 118, 152–155 (2008)
C.J. Wysocki, B.J. Cowart, T. Radil: Nasal trigeminal chemosensitivity across the adult life span, Percept. Psychophys. 65, 115–122 (2003)
A. Bramerson, L. Johansson, L. Ek, S. Nordin, M. Bende: Prevalence of olfactory dysfunction: The skovde population-based study, Laryngoscope 114, 733–737 (2004)
B.N. Landis, C.G. Konnerth, T. Hummel: A study on the frequency of olfactory dysfunction, Laryngoscope 114, 1764–1769 (2004)
J. Frasnelli, T. Hummel, J. Berg, G. Huang, R.L. Doty: Intranasal localizability of odorants: Influence of stimulus volume, Chem. Senses 36, 405–410 (2011)
J.E. Cometto-Muniz, W.S. Cain, M.H. Abraham, R. Kumarsingh: Trigeminal and olfactory chemosensory impact of selected terpenes, Pharmacol. Biochem. Behav. 60, 765–770 (1998)
J.E. Cometto-Muniz, W.S. Cain: Trigeminal and olfactory sensitivity: Comparison of modalities and methods of measurement, Int. Arch. Occup. Environ. Health 71, 105–110 (1998)
J. Frasnelli, B. Schuster, T. Zahnert, T. Hummel: Chemosensory specific reduction of trigeminal sensitivity in subjects with olfactory dysfunction, Neuroscience 142, 541–546 (2006)
H. Gudziol, M. Schubert, T. Hummel: Decreased trigeminal sensitivity in anosmia, J. Otorhinolaryngol. Relat. Spec. 63, 72–75 (2001)
T. Hummel, S. Barz, J. Lotsch, S. Roscher, B. Kettenmann, G. Kobal: Loss of olfactory function leads to a decrease of trigeminal sensitivity, Chem. Senses 21, 75–79 (1996)
M. Kendal-Reed, J.C. Walker, W.T. Morgan, M. LaMacchio, R.W. Lutz: Human responses to propionic acid. I. Quantification of within- and between-participant variation in perception by normosmics and anosmics, Chem. Senses 23, 71–82 (1998)
L. Cui, W.J. Evans: Olfactory event-related potentials to amyl acetate in congenital anosmia, Electroenceph. Clin. Neurophysiol. 102, 303–306 (1997)
G. Kobal, S. Van Toller, T. Hummel: Is there directional smelling?, Experientia 45, 130–132 (1989)
J. Frasnelli, G. Charbonneau, O. Collignon, F. Lepore: Odor localization and sniffing, Chem. Senses 34, 139–144 (2009)
E. von Skramlik: Über die Lokalisation der Empfindungen bei den niederen Sinnen, Z. Sinnesphysiol. 56, 69 (1924)
G. von Békésy: Olfactory analogue to directional hearing, J. Appl. Physiol. 19, 369–373 (1964)
T. Hummel, T. Futschik, J. Frasnelli, K.B. Huttenbrink: Effects of olfactory function, age, and gender on trigeminally mediated sensations: A study based on the lateralization of chemosensory stimuli, Toxicol. Lett. 140–141, 273–280 (2003)
J. Porter, T. Anand, B. Johnson, R.M. Khan, N. Sobel: Brain mechanisms for extracting spatial information from smell, Neuron 47, 581–592 (2005)
J. Frasnelli, B. Schuster, T. Hummel: Subjects with congenital anosmia have larger peripheral but similar central trigeminal responses, Cereb. Cortex 17, 370–377 (2007)
T. Hummel, G. Kobal: Chemosensory event-related potentials to trigeminal stimuli change in relation to the interval between repetitive stimulation of the nasal mucosa, Eur. Arch. Otorhinolaryngol. 256, 16–21 (1999)
L. Keita, J. Frasnelli, V. La Buissonniere-Ariza, F. Lepore: Response times and response accuracy for odor localization and identification, Neuroscience 238, 82–86 (2013)
J.E. Cometto-Muniz, W.S. Cain, H.K. Hudnell: Agonistic effects of airborne chemicals in mixtures: Odor, nasal pungency, and eye irritation, Percept. Psychophys. 59, 665–674 (1997)
J. Frasnelli, T. Hummel: Intranasal trigeminal threshold in healthy subjects, Environ. Toxicol. Pharmacol. 9, 575–580 (2005)
M.A. Smeets, P.J. Bulsing, S. Van Rooden, R. Steinmann, J.A. De Ru, N.W. Ogink, C. van Thriel, P.H. Dalton: Odor and irritation thresholds for ammonia: A comparison between static and dynamic olfactometry, Chem. Senses 32, 11–20 (2007)
P. Dalton, D. Dilks, T. Hummel: Effects of long-term exposure to volatile irritants on sensory thresholds, negative mucosal potentials, and event-related potentials, Behav. Neurosci. 120, 180–187 (2006)
D. Shusterman, J. Balmes: Measurement of nasal irritant sensitivity to pulsed carbon dioxide: A pilot study, Arch. Environ. Health 52, 334–340 (1997)
G. Kobal: Elektrophysiologische Untersuchungen des menschlichen Geruchssinns (Thieme, Stuttgart 1981)
T. Hummel, G. Kobal: Chemosensory evoked potentials. In: Chemical Signals in Vertebrates VI, ed. by R.L. Doty, D. Müller-Schwarze (Plenum, New York 1992) pp. 565–569
J. Frasnelli, J. Lotsch, T. Hummel: Event-related potentials to intranasal trigeminal stimuli change in relation to stimulus concentration and stimulus duration, J. Clin. Neurophysiol. 20, 80–86 (2003)
G. Kobal: Pain-related electrical potentials of the human nasal mucosa elicited by chemical stimulation, Pain 22, 151–163 (1985)
J. Lötsch, T. Hummel, H.G. Kraetsch, G. Kobal: The negative mucosal potential: Separating central and peripheral effects of NSAIDs in man, Eur. J. Clin. Pharmacol. 52, 359–364 (1997)
N. Thürauf, T. Hummel, B. Kettenmann, G. Kobal: Nociceptive and reflexive responses recorded from the human nasal mucosa, Brain Res. 629, 293–299 (1993)
N. Thürauf, I. Friedel, C. Hummel, G. Kobal: The mucosal potential elicited by noxious chemical stimuli: Is it a peripheral nociceptive event, Neurosci. Lett. 128, 297–300 (1991)
J. Frasnelli, T. Hummel: Age-related decline of intranasal trigeminal sensitivity: Is it a peripheral event?, Brain Res. 987, 201–206 (2003)
T. Hummel, C. Schiessl, J. Wendler, G. Kobal: Peripheral electrophysiological responses decrease in response to repetitive painful stimulation of the human nasal mucosa, Neurosci. Lett. 212, 37–40 (1996)
D. Ottoson: Analysis of the electrical activity of the olfactory epithelium, Acta Physiol. Scand. 35, 1–83 (1956)
D.A. Leopold, T. Hummel, J.E. Schwob, S.C. Hong, M. Knecht, G. Kobal: Anterior distribution of human olfactory epithelium, Laryngoscope 110, 417–421 (2000)
J. Frasnelli, B. Schuster, T. Hummel: Interactions between olfaction and the trigeminal system: What can be learned from olfactory loss, Cereb. Cortex 17, 2268–2275 (2007)
M. Scheibe, T. Zahnert, T. Hummel: Topographical differences in the trigeminal sensitivity of the human nasal mucosa, Neuroreport 17, 1417–1420 (2006)
E. Iannilli, J. Gerber, J. Frasnelli, T. Hummel: Intranasal trigeminal function in subjects with and without an intact sense of smell, Brain Res. 1139, 235–244 (2007)
T. Hummel, R.L. Doty, D.M. Yousem: Functional MRI of intranasal chemosensory trigeminal activation, Chem. Senses 30, i205–i206 (2005)
I. Savic, B. Gulyas, H. Berglund: Odorant differentiated pattern of cerebral activation: Comparison of acetone and vanillin, Hum. Brain Mapp. 17, 17–27 (2002)
T. Hummel, A. Livermore: Intranasal chemosensory function of the trigeminal nerve and aspects of its relation to olfaction, Int. Arch. Occup. Environ. Health 75, 305–313 (2002)
J. Frasnelli, T. Hummel: Interactions between the chemical senses: Trigeminal function in patients with olfactory loss, Int. J. Psychophysiol. 65, 177–181 (2007)
W.S. Cain, C.L. Murphy: Interaction between chemoreceptive modalities of odour and irritation, Nature 284, 255–257 (1980)
L. Cashion, A. Livermore, T. Hummel: Odour suppression in binary mixtures, Biol. Psychol. 73, 288–297 (2006)
T. Hummel, A. Livermore, C. Hummel, G. Kobal: Chemosensory event-related potentials in man: Relation to olfactory and painful sensations elicited by nicotine, Electroencephalogr. Clin. Neurophysiol. 84, 192–195 (1992)
A. Livermore, T. Hummel: The influence of training on chemosensory event-related potentials and interactions between the olfactory and trigeminal systems, Chem. Senses 29, 41–51 (2004)
L. Jacquot, J. Monnin, G. Brand: Influence of nasal trigeminal stimuli on olfactory sensitivity, C. R. Biol. 327, 305–311 (2004)
C.J. Wysocki, B.J. Cowart, E. Varga: Nasal-trigeminal sensitivity in normal aging and clinical populations, Chem. Senses 22, 826 (1997)
J. Frasnelli, B. Schuster, T. Hummel: Olfactory dysfunction affects thresholds to trigeminal chemosensory sensations, Neurosci. Lett. 468, 259–263 (2010)
W.S. Cain: Olfaction and the common chemical sense: Some psychophysical contrasts, Sens. Process. 1, 57–67 (1976)
E.L. Coates: Olfactory CO(2) chemoreceptors, Respir. Physiol. 129, 219–229 (2001)
A. Husner, J. Frasnelli, A. Welge-Lussen, G. Reiss, T. Zahnert, T. Hummel: Loss of trigeminal sensitivity reduces olfactory function, Laryngoscope 116, 1520–1522 (2006)
B. Kettenmann, C. Hummel, H. Stefan, G. Kobal: Multiple olfactory activity in the human neocortex identified by magnetic source imaging, Chem. Senses 22, 493–502 (1997)
R.J. Zatorre, M. Jones-Gotman, A.C. Evans, E. Meyer: Functional localization and lateralization of human olfactory cortex, Nature 360, 339–340 (1992)
E.T. Rolls, H.D. Critchley, A. Treves: Representation of olfactory information in the primate orbitofrontal cortex, J. Neurophysiol. 75, 1982–1996 (1996)
D.A. Kareken, M. Sabri, A.J. Radnovich, E. Claus, B. Foresman, D. Hector, G.D. Hutchins: Olfactory system activation from sniffing: Effects in piriform and orbitofrontal cortex, Neuroimage 22, 456–465 (2004)
D.H. Zald, J.V. Pardo: Emotion, olfaction, and the human amygdala: Amygdala activation during aversive olfactory stimulation, Proc. Natl. Acad. Sci. USA 15, 4119–4124 (1997)
I. Savic, B. Gulyas, M. Larsson, P. Roland: Olfactory functions are mediated by parallel and hierarchical processing, Neuron 26, 735–745 (2000)
B. Kettenmann, V. Jousmaki, K. Portin, R. Salmelin, G. Kobal, R. Hari: Odorants activate the human superior temporal sulcus, Neurosci. Lett. 203, 143–145 (1996)
Q. Chevy, E. Klingler: Odorless trigeminal stimulus CO\({}_{2}\) triggers response in the olfactory cortex, J. Neurosci. 34, 341–342 (2014)
T.E. Finger, M.L. Getchell, T.V. Getchell, J.C. Kinnamon: Affector and effector functions of peptidergic innervation of the nasal cavity. In: Chemical Senses: Irritation, ed. by B.G. Green, J.R. Mason, M.R. Kare (Marcel Dekker, New York 1990) pp. 1–20
G. Brand: Olfactory/trigeminal interactions in nasal chemoreception, Neurosci. Biobehav. Rev. 30, 908–917 (2006)
J.F. Bouvet, J.C. Delaleu, A. Holley: The activity of olfactory receptor cells is affected by acetylcholine and substance P, Neurosci. Res. 5, 214–223 (1988)
I. Kratskin, T. Hummel, L. Hastings, R. Doty: 3-Methylindole alters both olfactory and trigeminal nasal mucosal potentials in rats, Neuroreport 11, 2195–2197 (2000)
G. Kobal, C. Hummel: Cerebral chemosensory evoked potentials elicited by chemical stimulation of the human olfactory and respiratory nasal mucosa, Electroencephalogr. Clin. Neurophysiol. 71, 241–250 (1988)
T. Hummel, E. Iannilli, J. Frasnelli, J. Boyle, J. Gerber: Central processing of trigeminal activation in humans, Ann. NY Acad. Sci. 1170, 190–195 (2009)
T.E. Finger, B. Bottger: Peripheral peptidergic fibers of the trigeminal nerve in the olfactory bulb of the rat, J. Comp. Neurol. 334, 117–124 (1993)
J.M. Christie, G.L. Westbrook: Lateral excitation within the olfactory bulb, J. Neurosci. 26, 2269–2277 (2006)
B.P. Halpern, L.M. Nelson: Bulbar gustatory responses to anterior and to posterior tongue stimulation in the rat, Am. J. Physiol. 209, 105–110 (1965)
K.S. Carlson, C.Z. Xia, D.W. Wesson: Encoding and representation of intranasal CO\({}_{2}\) in the mouse olfactory cortex, J. Neurosci. 33, 13873–13881 (2013)
J.C. Walker, M. Kendal-Reed, S.B. Hall, W.T. Morgan, V.V. Polyakov, R.W. Lutz: Human responses to propionic acid. II. Quantification of breathing responses and their relationship to perception, Chem. Senses 26, 351–358 (2001)
T. Hummel, T. Futschik, J. Frasnelli, K.B. Huttenbrink: Effects of olfactory function, age, and gender, on trigeminally mediated sensations: A study based on the lateralization of chemosensory stimuli, Toxicol. Lett. 140, 273–280 (2003)
J. Seubert, J. Freiherr, J. Djordjevic, J.N. Lundstrom: Statistical localization of human olfactory cortex, Neuroimage 66C, 333–342 (2012)
J.A. Boyle, J. Frasnelli, J. Gerber, M. Heinke, T. Hummel: Cross-modal integration of intranasal stimuli: A functional magnetic resonance imaging study, Neuroscience 149, 223–231 (2007)
A.F. Temmel, C. Quint, B. Schickinger-Fischer, L. Klimek, E. Stoller, T. Hummel: Characteristics of olfactory disorders in relation to major causes of olfactory loss, Arch. Otolaryngol. Head Neck Surg. 128, 635–641 (2002)
H.W. Berendse, M.M. Ponsen: Detection of preclinical Parkinson’s disease along the olfactory tract, J. Neural Transm. Suppl. 70, 321–325 (2006)
A. Mackay-Sim, A.N. Johnston, C. Owen, T.H. Burne: Olfactory ability in the healthy population: Reassessing presbyosmia, Chem. Senses 31, 763–771 (2006)
J.C. Walker, M. Kendal-Reed, M.J. Utell, W.S. Cain: Human breathing and eye blink rate responses to airborne chemicals, Environ. Health Perspect. 109(Suppl 4), 507–512 (2001)
R.L. Doty: Intranasal trigeminal detection of chemical vapors by humans, Physiol. Behav. 14, 855–859 (1975)
J. Reden, A. Mueller, C. Mueller, I. Konstantinidis, J. Frasnelli, B.N. Landis, T. Hummel: Recovery of olfactory function following closed head injury or infections of the upper respiratory tract, Arch. Otolaryngol. Head Neck Surg. 132, 265–269 (2006)
A. Welge-Lussen, C. Wille, B. Renner, G. Kobal: Anesthesia affects olfaction and chemosensory event-related potentials, Clin. Neurophysiol. 115, 1384–1391 (2004)
W.S. Cain: Contribution of the trigeminal nerve to perceived odor magnitude, Ann. NY Acad. Sci. 237, 28–34 (1974)
Acknowledgements
We thank Cristian Petrescu for help with the illustrations. JF is supported by the Research Centre of Sacré-Coeur Hospital in Montréal, the University of Québec in Trois-Rivières, the Fonds de Recherche du Québec - Santé, and the Natural Sciences and Engineering Research Council of Canada; SM is supported by a scholarship from NSERC.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Frasnelli, J., Manescu, S. (2017). The Intranasal Trigeminal System. In: Buettner, A. (eds) Springer Handbook of Odor. Springer Handbooks. Springer, Cham. https://doi.org/10.1007/978-3-319-26932-0_46
Download citation
DOI: https://doi.org/10.1007/978-3-319-26932-0_46
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-26930-6
Online ISBN: 978-3-319-26932-0
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)