
Abstract
In oxygenic photosynthesis, photosystem I (PSI) drives electron transfer from plastocyanin or cytochrome c to ferredoxin. PSI forms a core complex that consists of 11–14 subunits and associates more than one hundred cofactors such as chlorophyll a, carotenes, naphthoquinones, lipids, and iron-sulfur (Fe-S) clusters. PSI complex is embedded in the thylakoid membranes and harvests light using antenna pigments, transfers excitons from antenna to reaction center, where photochemical reaction occurs, and stabilizes the charge separation to allow for oxidation of plastocyanin or cytochrome c and reduction of ferredoxin. PSI core complex forms a trimer in cyanobacteria but associates light-harvesting complexes I (LHCI) to form a PSI-LHCI supercomplex in plants and algae. The crystal structures of PSI core trimer and PSI-LHCI supercomplex have revealed that the subunits and cofactors of PSI are organized to optimize the efficiency of light harvesting and photochemical reaction. However it remains elusive how these components of PSI are integrated into a functional structure in photosynthetic organisms.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Thermophilic Cyanobacterium
- Antenna Chlorophyll
- Exciton Transfer
- Redox Component
- Cyclic Electron Transfer
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Oxygenic photosynthesis by plants, algae, and cyanobacteria produces carbohydrates from CO2 and H2O using light energy and generates O2 as a by-product. Photosystem II (PSII) generates strong oxidants that split H2O into O2, H+, and electrons, whereas photosystem I (PSI ) produces strong reductants. Two photosystems operate in series to drive oxygenic photosynthetic electron transfer from H2O to nicotinamide adenine dinucleotide phosphate (NADP+) to produce NADPH. This linear electron transfer reactions concomitantly generate proton motive force, which is used to synthesize ATP from ADP and phosphate (Pi) by ATP synthase. NADPH and ATP are used to assimilate CO2 (Fig. 1). Light energy is first absorbed by photosynthetic pigments such as chlorophylls and carotenoids , and the resulting excitons are transferred to reaction centers (RCs) where photochemical reaction causes charge separation . A series of electron donors and acceptors in RCs are engaged in the charge separation and its stabilization.

Electron transfer system of oxygenic photosynthesis. Four multiprotein complexes (PSI, PSII, Cyt b 6 f, and ATP synthase) are localized in the thylakoid membranes. PQ pool is present in the hydrophobic region of the thylakoid membranes and Pc, FNR, and Fd are located on the peripheral region of the thylakoid membranes. Electron transfer reaction is driven by PSI and PSII using light energy. Proton translocation across the thylakoid membranes is coupled to the electron transfer reactions. The resulting ΔpH across and membrane potential are used to produce ATP from ADP and Pi (inorganic phosphate) by ATP synthase
PSI accepts electrons from PSII through plastoquinone (PQ), cytochrome b 6 f (Cyt b 6 f) complex, and plastocyanin (Pc) or cytochrome c (Cyt c), and donates electrons to NADP+ through ferredoxin (Fd) and ferredoxin-NADP oxidoreductase (FNR) (Fig. 1). PSI core complex consists of 11–14 subunits and a number of cofactors such as chlorophyll a (Chla), carotenes, naphthoquinones, lipids, and iron-sulfur (Fe-S) clusters and is embedded in lipid bilayer of the thylakoid membranes as a large intrinsic complex. Thus PSI is a multi-protein complex that harvests light energy, causes photochemical reaction, and stabilizes the charge separation. In addition to the linear electron transfer, PSI is engaged in driving the cyclic electron transfer by which Fd reduced by PSI transfers an electron back to PSI through PQ, Cyt b 6 f, and Pc (or Cyt c) to produce ATP without generating NADPH. This cyclic electron transfer plays important roles not only in producing ATP to optimize ATP/NADPH ratio but also in protecting PSI from excess light energy. The reductants generated by PSI are also used for assimilation of nitrogen and sulfur as well as for providing reducing power for redox regulation of enzymes in chloroplasts. Thus PSI plays multiple and pivotal functions in oxygenic photosynthesis.
2 PSI Forms a Chlorophyll-Protein Complex
The components of PSI are precisely organized in a chlorophyll-protein complex to carry out efficient light harvesting and photochemical reactions. PSI complex, which is embedded in the thylakoid membranes , can be solubilized using a mild detergent and can be biochemically purified without significantly affecting its structure and activity. Figure 2 shows the subunit composition of the PSI preparation purified from the thylakoid extracts of the green alga Chlamydomonas reinhardtii. SDS-polyacrylamide gel electrophoresis clearly revealed that the purified PSI complex contains a number of PSI subunits. The diffuse bands around 70 kDa are two homologous RC subunits encoded by psaA and psaB genes. In addition to these RC subunits, the PSI complex contains several small subunits that are encoded by psaC, psaD, psaE, psaF, psaG, psaH, psaK, and psaL genes. It was also confirmed by immunoblotting analyses using anti-PsaI and anti-PsaJ antibodies that the PSI complex also contains very small subunits (<5 kDa) encoded by psaI and psaJ genes. As summarized in Table 1, the chloroplast genes (psaA, psaB, psaC, psaI and psaJ) encode four subunits while the nuclear genes coded for the other subunits. Localization of PsaO in PSI complex has not yet been determined because this subunit is weakly bound to and is easily dissociated from PSI complex during solubilization and purification. However chemical cross-linking experiments revealed that PsaO interacts with PsaL [1]. In addition to the PSI subunits present in the PSI core complex, the PSI complex from Chlamydomonas contains nine subunits (Lhca1-9) of light-harvesting complex I (LHCI ) [2]. Because their sizes are rather similar, some of them are not clearly separated on SDS-polyacrylamide gel. The PSI complex from Chlamydomonas is thus designated as a PSI-LHCI supercomplex. It is of note that plant PSI also forms PSI-LHCI supercomplex but the number of LHCI is four as will be described below. In summary, Chlamydomonas PSI-LHCI supercomplex has a larger structure of about 700 kDa with 40 transmembrane helices.

Subunit composition of PSI-LHCI supercomplex purified from the green alga Chlamydomonas reinhardtii. The purified PSI-LHCI supercomplex from the thylakoid extracts was dissociated into polypeptides and the polypeptides were separated by SDS-polyacrylamide gel electrophoresis. The separated polypeptides were stained. Two large homologous polypeptides, PsaA and PsaB are separated around 70 kDa and several smaller polypeptides (PsaD, PsaF, PsaL, PsaH, PsaE, PsaG, PsaC, and PsaK) are resolved. Very small polypeptides, PsaI and PsaJ, are not observed under this condition. In addition to PSI polypeptides, nine LHCI polypeptides (Lhca1-9) are separated. Since some Lhca polypeptides are closely separated, some of them are not detected individually
PSI core complex has a structure conserved among cyanobacteria, algae, and plants and is composed of PsaA, PsaB, PsaC, PsaD, PsaE, PsaF, PsaI, PsaJ, PsaK, and PsaL and associates cofactors necessary to harvest light energy, charge separation and its stabilization in common [3]. However, there are some minor differences in subunit composition between cyanobacteria and algae/plants; cyanobacterial PSI core complex additionally contains PsaM and PsaX (PsaX is found only in thermophilic cyanobacteria) while algae and plats contain PsaG, PsaH, PsaN, and PsaO (PsaP is found only in plants). It is of note that cyanobacterial PSI core complex forms a trimer [4], whereas in algae and plants PSI core complex is a monomeric and forms a PSI-LHCI supercomplex [5].
3 Subunit Structure of Photosystem I Core Complex
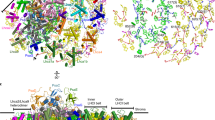
The crystal structures of PSI core trimer from the thermophilic cyanobacteria Thermosynechococcus elongatus and PSI-LHCI supercomplex from Pisus sativum (pea) have been determined at 2.5 and 2.8 Å resolutions, respectively [4, 6, 7]. The subunit structure and cofactor organization are highly conserved between these two core complexes (Fig. 3a, b). The structure of the cyanobacterial core complex viewed from the stromal side shows that two homologous RC subunits, PsaA and PsaB, each with 11 transmembrane helices, form a heterodimer (Fig. 3a). On the surface of the stromal side, three subunits, PsaC, PsaD, and PsaE, are clustered to constitute the docking site for Fd (Fig. 3a). On a hydrophobic side of the RC, three intrinsic subunits, PsaI, PsaL, and PsaM, are intimately associated with one another (Fig. 3a, c). PsaL and Ca2+ are located at the center of PSI core trimer in cyanobacteria and are required for the stabilization of the trimer. In plants, PsaM is absent but PsaH is located in close proximity of PsaL (Fig. 3b). The presence of PsaH in plants and algae prevents the PSI core complex from the trimerization. Instead PsaL and PsaH are involved in reversible association of light-harvesting complexes II (LHCIIs) with PSI-LHCI supercomplex during the state transitions by which LHCIIs are shuttled between PSI and PSII complexes in order to balance excitation energy distribution. It is inferred that Chla molecules bound to PsaL and PsaH may be involved in exciton transfer from LHCII to PSI RC [6]. On the other hydrophobic side of PsaI and PsaL, three intrinsic subunits, PsaF, PsaJ, and PsaX, are located (Fig. 3a, d). In plants, PsaX is absent but four LHCIs (Lhca1-4) are present on the side of PsaF and PsaJ. PsaK is attached to PsaA in cyanobacteria while PsaG is additionally associated with PsaB at a pseudo-symmetric position with respect to PsaK in plants and algae. These two subunits are homologous and are required for LHCI binding to PSI core complex in plants and algae [8]. In plants and algae, PsaF has two extended α-helices on the lumenal side of the PSI RC subunits, which are involved in Pc or Cyt c docking and are required for the efficient electron transfer from Pc or Cyt c to P700 [5].

Subunit organization of PSI core complex and PSI-LHCI supercomplex. a The subunit structure viewed from stromal side of cyanobacterial PSI core complex (PDB 1JB0). The structure of the monomeric PSI core is shown. b The subunit structure viewed from stromal side of plant PSI-LHCI supercomplex (PDB 2WSC). In addition to PSI core complex, four LHCI complexes are attached on the PsaF and PsaJ side of the PSI core complex. c The subunit structure viewed from PsaL/PsaI/PsaM side of the cyanobacterial PSI core complex. d The subunit structure viewed from PsaF/PsaJ/PsaX side of the cyanobacterial PSI core complex
4 Electron Transfer Reactions
The RC of PSI contains one electron donor and five electron acceptors involved in photochemical reactions (Fig. 4). The initial charge separation occurs between the primary electron donor, P700 , and an intermediate electron acceptor, A0 , resulting in the radical pair, P700+/A0 −. The reduced A0 subsequently donates the electron to another electron acceptor, A1. A1 − successively donates the electron to the three 4Fe-4S clusters, FX , FA , and FB . The resulting charge separation P700+/F −B is significantly stable (t1/2 of the back reaction from F −B to P700+ is about 100 ms) so that P700+ can oxidize Pc or Cyt c and F −B can reduce Fd. Finally the redox components of PSI return to the original redox state and carries out another round of photochemical reaction.

Electron donor and acceptors of PSI. The primary electron donor, P700, and the three electron acceptors, A0, A1, and FX, are localized on PsaA/PsaB heterodimer. The initial charge separation occurs at t1/2 of 20–30 ps and the electron transfer from A1 to FX occurs at t1/2 of 100 and 700 ns. The terminal electron acceptors, FA and FB, are located on PsaC. Pc (Cyt c) reduces P700+, whereas Fd is reduced by F −B . The back reaction from F −B to P700+ occurs at t1/2 of about 100 ms
The crystal structure of PSI of the thermophilic cyanobacterium, T. elongatus, has also revealed the detailed organization of the redox components [4]. Figure 5 shows the configuration of the redox components in PSI core complex. Six Chla and two naphthoquinone molecules are located pseudo-symmetrically on the heterodimer of PsaA and PsaB. It is of note that two branches of redox components from P700 to Fx (A and B branches) are present. The structure shows that P700 is Chla heterodimer; one is Chla epimer (Chla′) coordinated by His-676 of PsaA and the other is Chla coordinated by His-656 of PsaB. The Chla molecules of the second pair, which are designated as accessory chlorophylls, are located between P700 and A0. They are coordinated by water molecules that are hydrogen-bonded to Asn-591 of PsaB and Asn-604 of PsaA, respectively. Biophysical measurements failed to detect a transient radical pair between P700 and the accessory chlorophyll probably because the lifetime of the reduced state of the accessory chlorophyll is too short to detect. The Chla molecules of the third pair, which are coordinated by Met-688 of PsaA and Met-668 of PsaB, respectively, correspond to A0. Naphthoquinone (phylloquinone) molecules of the fourth pair, which correspond to A1, are hydrogen-bonded to NH of backbone of Leu-722 on PsaA and Leu-706 of PsaB, respectively. Additionally their quinone planes interact with indole rings of Trp-697 of PsaA and Trp-677 of PsaB, respectively. The chemical species of A1 is naphthoquinone in the cyanobacterial and plant crystal structures. However three types of naphthoquinone have been reported to be present in PSI core complex depending on photosynthetic organisms (Fig. 6). A1 is phylloquinone in higher plants, some algae, and some cyanobacteria. However, A1 is 5′-monohydroxyphylloquinone or menaquinone-4 in some algae and some cyanobacteria [9]. It is not yet clear whether the difference of the naphthoquinone has any physiological significance.

The configuration of the PSI redox components. Chla dimer, P700, and a pair of accessory chlorophylls, A0, and A1 form two branches of electron transfer chain (PDB 1JB0). The two branches are connected at FX, and subsequently FA and FB successively accept electrons. Two His residues on PsaA and PsaB coordinate chlorophylls of P700. Two Trp residues on PsaA and PsaB are located between P700 and Pc (or Cyt c) binding site to facilitate electron transfer reaction from Pc (or Cyt c) to P700. Only chlorin ring of Chla is shown

Chemical structures of the three naphthoquinones
Site-directed mutagenesis to replace His-676 of PsaA or His 656 of PsaA and biophysical characterization of the resulting mutants suggested that spin density on the P700+ radical is unevenly distributed between the Chla pair; more density is detected on Chla coordinated by His-656 of PsaB [10, 11]. However, the two branches of the redox components are functional [12]. Kinetic measurements revealed that the reoxidation of A1 - by FX has two phases of t1/2 = 13 ns and t1/2 = 160 ns, suggesting that the two phylloquinone molecules are involved in the electron transfer from A1 to FX [13]. Site-directed mutagenesis to replace the Trp residue interacting with one of phylloquinone molecules revealed that the replacement of Trp residue of PsaB affected the fast phase while the replacement of Trp residue of PsaA affected the slow phase [14]. FX is coordinated between PsaA and PsaB by four cysteine residues; Cys-575 and Cys-584 of PsaA and Cys-561 and Cys-570 of PsaB coordinate the 4Fe-4S cluster. Two 4Fe-4S clusters, FA and FB, are located on PsaC. Two Trp residues on PsaA and PsaB, respectively, are located between P700 and the docking site for Pc and Cyt c, and facilitate the efficient electron transfer from Pc or Cyt c to P700 [15].
5 Antenna Pigments in PSI Core and LHCI Complexes
The cyanobacterial PSI core complex associates 96 Chla molecules; six Chla molecules are required for the electron transfer reaction, whereas 90 Chla molecules are involved in light harvesting as antenna chlorophylls. 79 Chla molecules are bound to the PSI RC heterodimer; PsaA and PsaB associate 40 and 39 Chla molecules, respectively, whereas the small subunits such as PsaJ, PsaK, PsaL, PsaM and PsaX, and a lipid, phosphatidylglycerol (PG), coordinate to Mg2+ of 11 Chla molecules.
The arrangements of the 21 transmembrane helices of the PSI RC subunits (PsaA and PsaB) and the PSII core complex (PsbA, PsbB, PsbC, and PsbD) are well conserved. The domains consisting of six helices (a-f) of the N-termini of PsaA and PsaB (peripheral parts), which correspond to the six helices of CP47 and CP43, bind 27 Chla molecules. By contrast, the domains consisting of five helices (g-k) of the C-termini of PsaA and PsaB (central parts), which correspond to the five helices of PsbA (D1) and PsbB (D2) of PSII RC, associate 13 and 12 Chla molecules, respectively (Fig. 7). The antenna chlorophylls are not distributed uniformly so that the distances among adjacent Chla molecules are close enough for efficient exciton transfer. Most Chla molecules are coordinated to imidazole group of His residues, or oxygen atoms of the side chain of Asp, Gln, Glu, and Tyr residues, of peptide bonds, or of water molecules [4]. For example, the peripheral parts of PsaA and PsaB associate 18 Chla molecules, which forms layer-like structures at the stromal and lumenal sides. However the antenna chlorophylls are separated from the six chlorophylls involved in the electron transfer except for the two Chla molecules on PsaA and PsaB, which are located in the proximity of A0 on PsaA and PsaB, respectively. It is proposed that excitons are transferred from the antenna pigments to RC mainly through these Chla molecules.

The arrangement of 22 transmembrane helices of PsaA and PsaB. a Localization of 11 transmembrane helices (a-k) of PsaA and PsaB are shown. Six transmembrane helices of N-termini of PsaA and PsaB (a-f) are shown in green [4]. These domains form peripheral parts of PSI RC and function as light harvesting. Five transmembrane helices of C-termini of PsaA and PsaB (g-k) are shown in magenta and purple. These domains are interlocked to constitute central part involved in both light harvesting and photochemical reaction. b Cross-section model of 11 transmembrane helices of PsaA and PsaB. Two domains consisting of helices a-f of PsaA and PsaB and two domains consisting of helices g-k of PsaA and PsaB are shown. P700 is located between two k helices of PsaA and PsaB whereas FX is located between two interhelical loops connecting h and i helices of PsaA and PsaB. The number of Chla associated with the four domains is indicated
One of the interesting features of the antenna chlorophylls of PSI complex is the presence of chlorophylls that absorb light at longer wavelengths than that of P700, which are designated as ‘red chlorophylls ’ or ‘red-shifted chlorophylls’. One trimer at the lumenal side of PsaB and three dimers of Chla at the central parts of PsaA and PsaB are tentatively assigned as red chlorophylls [4]. The three Chla molecules forming trimer are stacked like a staircase, whereas the chlorin planes of the three dimers are nearly parallel.
β-carotene molecules play roles in light harvesting and protecting RCs from photodamage by quenching excited Chla triplet states generated by photochemical reactions. Most of β-carotene molecules are deeply inserted in the PSI complex although many Chla molecules are present near the stromal or lumenal surface [1, 4, 7].
Recent progresses in the determination of the structure of PSI-LHCI have revealed detailed structure of four LHCIs (Lhca1-4) stably associated to PSI core complex in plant [5–7, 16]. PSI-LHCI associates 155 Chla, 12 Chlb, 28 β-carotene, 5 lutein, and 4 violaxanthin molecules. The four LHCIs associate 45 Chla, 12 Chlb, 4 β-carotene, 5 lutein, and 4 violaxanthin molecules [7]. Although the structures of the four LHCIs are well conserved, the distribution of Chlb is not uniform among the four LHCIs as summarized in Table 2. Lhca2 and Lhca4, which are located in the center of the LHCI tetramer, associate more Chlb molecules than Lhca1 and Lhca3. Each LHCI associates one molecule of β-carotene, lutein, and violaxanthin in addition to Chla and Chlb. However one additional lutein molecule is present between Lhca1 and Lhca4. As shown in Fig. 3b, Lhca1, Lhca4, Lhca2, and Lhca3 form a tetramer on one side of PSI core complex (PsaF and PsaJ side), which is located between PsaG and PsaK. PsaG and PsaK, which are homologous and are located pseudo-symmetrically, are involved in stabilization of the LHCI tetramer.
Two main pathways for exciton transfer from LHCI to PSI core are proposed on the basis of the structure of PSI-LHCI supercomplex. PsaG and PsaK bind Chla molecules and intimately associate with Lhca1 and Lhca3, respectively. It is inferred that excitons on the LHCI tetramer are first moved to Lhca1 or Lhca3, and are then transferred to PSI core antenna through Chla molecules associated with PsaG and PsaK, respectively.
6 Biogenesis of Photosystem I Complex
The subunits and cofactors of PSI core complex in cyanobacteria and PSI-LHCI supercomplex in plants and algae are precisely organized as already described above. Thus these components need to be synthesized on demand and the synthesized components must be properly integrated into a functional complex. It is unlikely that this assembly process proceeds spontaneously. Accordingly it is expected that an assembly apparatus for PSI complex, which assists integration of the PSI components, should be present [17, 18]. However it remains elusive how and by which factors the constituent components are integrated into PSI complexes. However several factors that are necessary for proper assembly of PSI complex have been reported [17, 18]. Among them, the chloroplast-encoded Ycf3 and Ycf4 proteins are essential factors for PSI biogenesis in the green alga Chlamydomonas, because the knockout mutants of Ycf3 or Ycf4 accumulate no PSI complex [19]. Biochemical isolation and characterization of Ycf4 protein revealed that Ycf4 is part of a large complex, on which newly synthesized PSI subunits are assembled into a PSI core subcomplex. Subsequently the other small PSI subunits are successively integrated into the subcomplex. In the case of PSI-LHCI assembly, the integration of PsaG and PsaK are one of the latest steps in the PSI biogenesis [8]. However more detailed assembly process remain to be addressed.
7 Conclusion
The PSI complexes of cyanobacteria, algae, and plants have functionality for efficient light harvesting, charge separation, and electron transfer to oxidize Pc or Cyt c and reduce Fd. 11-14 subunits of PSI core complex and a number of cofactors involved in charge separation and electron transfer as well as in light harvesting are precisely organized in large chlorophyll-protein complexes. The determined structures of PSI core trimer and PSI-LHCI supercomplex have revealed the detailed structure responsible for the conversion of labile light energy into stable redox energy at quantum yield of nearly 1.0.
References
Jensen PE et al (2004) The PSI-O subunit of plant photosystem I is involved in balancing the excitation pressure between the two photosystems. J Biol Chem 279(23):24212–24217
Takahashi Y et al (2004) Comparison of the subunit compositions of the PSI-LHCI supercomplex and the LHCI in the green alga Chlamydomonas reinhardtii. Biochemistry 43(24):7816–7823
Nelson N, Yocum CF (2006) Structure and function of photosystems I and II. Annu Rev Plant Biol 57:521–565
Jordan P et al (2001) Three-dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 411(6840):909–917
Ben-Shem A, Frolow F, Nelson N (2003) Crystal structure of plant photosystem I. Nature 426(6967):630–635
Mazor Y, Borovikova A, Nelson N (2015) The structure of plant photosystem I super-complex at 2.8 A resolution, vol 4. Elife
Qin X et al (2015) Photosynthesis. structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 348(6238):989–995
Ozawa S, Onishi T, Takahashi Y (2010) Identification and characterization of an assembly intermediate subcomplex of photosystem I in the green alga Chlamydomonas reinhardtii. J Biol Chem 285(26):20072–20079
Ozawa SI et al (2012) 5′-Monohydroxyphylloquinone is the dominant naphthoquinone of PSI in the green alga Chlamydomonas reinhardtii. Plant Cell Physiol 53(1):237–243
Redding K et al (1998) A systematic survey of conserved histidines in the core subunits of photosystem I by site-directed mutagenesis reveals the likely axial ligands of P700. EMBO J 17(1):50–60
Webber AN et al (1996) Site-directed mutations affecting the spectroscopic characteristics and midpoint potential of the primary donor in photosystem I. Biochemistry 35(39):12857–12863
Brettel K (1997) Electron transfer and arrangement of the redox cofactors in photosystem I. Biochimica Et Biophysica Acta-Bioenergetics 1318(3):322–373
Joliot P, Joliot A (1999) In vivo analysis of the electron transfer within photosystem I: are the two phylloquinones involved? Biochemistry 38(34):11130–11136
Guergova-Kuras M et al (2001) Evidence for two active branches for electron transfer in photosystem I. Proc Natl Acad Sci U S A 98(8):4437–4442
Sommer F et al (2004) The hydrophobic recognition site formed by residues PsaA-Trp(651) and PsaB-Trp(627) of photosystem I in Chlamydomonas reinhardtii confers distinct selectivity for binding of plastocyanin and cytochrome c(6). J Biol Chem 279(19):20009–20017
Amunts A, Drory O, Nelson N (2007) The structure of a plant photosystem I supercomplex at 3.4 A resolution. Nature 447(7140):58–63
Minagawa J, Takahashi Y (2004) Structure, function and assembly of Photosystem II and its light-harvesting proteins. Photosynth Res 82(3):241–263
Schottler MA, Albus CA, Bock R (2011) Photosystem I: Its biogenesis and function in higher plants. J Plant Physiol 168(12):1452–1461
Boudreau E et al (1997) The chloroplast ycf3 and ycf4 open reading frames of Chlamydomonas reinhardtii are required for the accumulation of the photosystem I complex. EMBO J 16(20):6095–6104
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Takahashi, Y. (2016). Recent Understanding on Photosystem I. In: Sugiyama, M., Fujii, K., Nakamura, S. (eds) Solar to Chemical Energy Conversion. Lecture Notes in Energy, vol 32. Springer, Cham. https://doi.org/10.1007/978-3-319-25400-5_24
Download citation
DOI: https://doi.org/10.1007/978-3-319-25400-5_24
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-25398-5
Online ISBN: 978-3-319-25400-5
eBook Packages: EnergyEnergy (R0)