
Abstract
In the Upper Danube basin, plant growth and biomass production strongly influence water, carbon and nitrogen fluxes. The model component Biological is part of the simulation system DANUBIA and calculates plant growth and biomass production for various vegetation categories (e.g. grassland, winter wheat, sugar beet and maize). An ecohydrological model approach is needed to account for the interactions of water, carbon and nitrogen fluxes in the soil-plant-atmosphere system. Meteorological and pedological model input data are provided by dynamically coupled DANUBIA model components. When analysing global change effects on agroecosystems, it is crucial to consider agricultural decisions such as type of use and management. Biological uses this information as input data from the coupled Farming actor model. In turn, the modelled biomass production and yield data are needed by Farming for the selection of crops to be cultivated. For the model validation analysis, measured and modelled values were compared for several test fields covering a wide range of meteorological and pedological conditions. On the district scale, agricultural statistics were used for validation. Maps of biomass production for 1 year are shown for selected field crops and managed grassland in the Upper Danube basin. Within DANUBIA, spatial and temporal changes in plant growth and biomass production for the past, present and future are assessed at the regional scale.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords

1 Introduction
Plant growth and biomass production govern to a large extent the water and matter regime within the Upper Danube drainage basin. Especially in agricultural ecosystems, biomass production and the expected yield are crucial for decisions on land use and management. The Biological model component simulates the effects of changed environmental conditions and land use on plant growth and biomass production for seminatural and agricultural ecosystems as well as their effects on the fluxes of water, carbon (C) and nitrogen (N).
Calculations of CO2 uptake and transpiration (see Chap. 36) form the basis for modelling plant growth and biomass production. When analysing the effects of changed environmental conditions on plant growth and biomass production, it is necessary to consider numerous other processes. Thus, for example, biomass production and also yield are determined not only by the phenological pattern but also by the distribution of assimilated C and incorporated N among the different plant organs (allocation).
Climate change therefore leads to adjustments in agricultural decisions such as type of use and management (e.g. sowing date, fertiliser plan). The close interdependency of natural and socioeconomic processes is implemented by coupling the Biological model component and the Farming actor model (see Chap. 40). The management decisions modelled by Farming influence biomass production. The resulting yield in turn affects the selection of crops to be cultivated.
2 Data Processing
See Chap. 36.
3 Model Documentation
Biological models the water and nutrient fluxes of the vegetation in seminatural and agricultural ecosystems within DANUBIA. A variety of interactions determine the fluxes of water, C and N in the soil-plant-atmosphere system. Thus, for example, photosynthesis is influenced by the concentration of nitrogen in the leaves. Higher concentrations of nitrogen promote photosynthesis. Increased uptake of carbon boosts biomass production, which raises nitrogen demand. Nitrogen uptake is coupled to water uptake and depends on root length density (root length per unit soil volume). An ecohydrological model approach is needed to account for these interdependencies. In this way, the response of plants to meteorological conditions and to the availability of water and nitrogen is depicted.
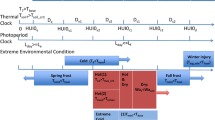
Biological combines the two process-based plant growth models Crop Environment Resource Synthesis (CERES) (Jones and Kiniry 1986) and Genotype-by-Environment interaction on CROp growth Simulator (GECROS) (Yin and van Laar 2005). The key processes modelled in Biological are presented in Fig. 37.1. Uptake of water and N is modelled according to CERES approaches. In contrast, the following processes are modelled using GECROS concepts: photosynthesis and transpiration (see Chap. 36), allocation of C and N, respiration (CO2 release), N demand, N fixation in legumes, leaf area development and senescence (leaf and root decay).

Modelled processes in Biological
The formation of root biomass and its decay are calculated according to GECROS. The development of root length density as a determining variable for water and N uptake is modelled according to CERES. Water and N uptake and root length density are modelled soil layer specific. Soil water, nitrate and ammonium content, which are dynamically modelled by Soil and Soil Nitrogen Transformation (SNT) (see Chaps. 24 and 38), are used as input along with field capacity, wilting point, saturated water content and bulk density, all of which are soil texture specific. N uptake is differentiated as either nitrate or ammonium uptake.
The GECROS phenology model was expanded to model phenological development according to Streck et al. (2003) in order to include vernalisation in addition to temperature and day length. Vernalisation is a key factor for winter wheat and winter rapeseed especially in the context of climate change. Moreover, modelling of germination and emergence (the appearance of sprouts or leaves at the ground surface) was complemented according to CERES. This approach ensures that an adjustment of the sowing date (on the part of the Farming actor model) as a result of a predicted climate change is adequately depicted by the model.
Dynamic modelling of the C and N allocation is needed in order to predict future plant growth and biomass production. Therefore, allocation is calculated based on the functional equilibrium approach according to GECROS. Thus, during times of insufficient water or N supply, root biomass production is increased, and correspondingly, with low available radiation, production of above-ground biomass is promoted. Modelling of leaf area development and senescence is physiologically based, according to GECROS. Leaf area is calculated based on the dynamic allocation of C and N in the leaves. Senescence is triggered when the concentration of N in the leaves or roots drops to a critical value.
All process descriptions in Biological are equally applicable for all vegetation categories. A specific differentiation is achieved by assigning the considered plant to functional groups (e.g. winter or summer crop) as well as by plant-specific parameterisation of the model input variables. Biological implements plant growth for a total of 22 different vegetation categories. Process-based growth models have been implemented for winter wheat (a representative for winter cereals), spring barley (a representative for summer cereals), winter rapeseed (for oil seeds), peas (for legumes), maize, potato, sugar beet and managed grassland. The yields for the other vegetation categories are calculated from empirically derived regression equations based on the modelled yields for plants with comparable phenology and management. For example, spring wheat yield is derived from the value modelled for spring barley. The regression equations were calculated from long-term statistics for agricultural districts (BLSD 2008 and SLBW 2008). For forests, only transpiration and net assimilation are modelled based on a predefined leaf area index (LAI) development.
Validation of the model results was carried out independent of scale. Measured and modelled values were compared for several test fields that cover a wide range of meteorological and pedological conditions. The measured data consist of LAI (leaf area per unit ground area) as well as biomass and C and N content of the individual plant organs, all for different plant types. Validation data are taken in part from field measurements by the working group (e.g. Lenz 2007; Lenz-Wiedemann et al. 2010) and in part from the literature (Kenter 2003 and McVoy et al. 1995). On the district scale, agricultural statistics were used for validation. In addition to the central task of modelling transpiration, Biological also serves within DANUBIA to provide harvest yields to the Farming actor model. The model results relating to the interaction between soil and plants (root growth, uptake of water and N, release of organic C and N back to the soil) are used by Soil and SNT. Results relating to the interaction between the atmosphere and vegetation (LAI, leaf temperature, plant height) are imported by RadiationBalance and Surface.
4 Results
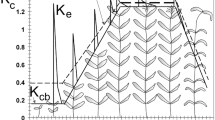
The comparison of measured and modelled biomass and leaf area development is shown in Fig. 37.2 using the example of a sugar beet field. At the beginning of growth, the model marginally overestimates the photosynthetically active leaf area, while later deviations are greater. The mean square error between the modelled and measured LAI is 0.54 m2/m2. Measured values and model results for green leaf biomass agree well at some points of time but show significant deviations at others. This results in a mean square error of 147 g/m2. The modelled biomass of dead brown leaves (44 g/m2) is almost exactly equivalent to the last measured value (46 g/m2). The mean square error for the beet body is 86 g/m2. A corresponding validation with comparable results was carried out for six additional sugar beet fields, three spring barley fields, two maize fields, three winter wheat fields and two potato fields (Lenz 2007).

Comparison of measured and modelled biomass (in g dry matter per m2) and leaf area development (m2 green leaf area per m2 ground surface) for sugar beet (2005 in Feienberg, measurements taken by the authors) (With permission from Lenz-Wiedemann et al. 2010)
Map 37.1 shows the spatial distribution of biomass production in 1995 for the Upper Danube basin. The model results are presented for selected field crops (total biomass without roots) and managed grassland (biomass at cutting date). The influence of soil characteristics on biomass production can easily be traced on the maps. Thus, the intensively cultivated regions on loess soils in the Dungau area show high biomass production, while the regions less favourable to agriculture in the Allgäu exhibit lower biomass production.
References
Bavarian State Ministry of Food, Agriculture and Forestry (ed) (1996) Bayerischer Agrarbericht, Munich
BLSD (2008) Datenbank GENESIS online. Bayerisches Landesamt für Statistik und Datenverarbeitung, München
Jones CA, Kiniry JR (eds) (1986) CERES-Maize. A simulation model of maize growth and development. Texas A&M University Press, College Station
Kenter C (2003) Ertragsbildung von Zuckerrüben in Abhängigkeit von der Witterung. PhD thesis, Georg-August-University, Göttingen
KTBL Association for Technology and Structures in Agriculture (2000/01) Taschenbuch Landwirtschaft. Landwirtschaftsverlag GmbH, Münster
Lenz VIS (2007) A process-based crop growth model for assessing Global Change effects on biomass production and water demand. A component of the integrative Global Change decision support system DANUBIA. PhD thesis, University of Cologne
Lenz-Wiedemann VIS, Klar CW, Schneider K (2010) Development and test of a crop growth model for application within a Global Change decision support system. Ecol Model 221:314–329, Elsevier
McVoy CW, Kersebaum KC, Arning M, Kleeberg P, Othmer H, Schröder U (1995) A data set from north Germany for the validation of agroecosystem models: documentation and evaluation. Ecol Model 81:265–300
SLBW (2008) Struktur und Regionaldatenbank. Statistisches Landesamt Baden-Württemberg, Stuttgart
Streck NA, Weiss A, Baenziger PS (2003) A generalized vernalization function for winter wheat. Agron J 95:155–159
Yin X, van Laar H (2005) Crop systems dynamics. An ecophysiological simulation model for genotype-by-environment interactions. Wageningen Academic Publishers, Wageningen
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Lenz-Wiedemann, V.I.S., Reichenau, T.G., Klar, C.W., Schneider, K. (2016). Plant Growth and Biomass Production. In: Mauser, W., Prasch, M. (eds) Regional Assessment of Global Change Impacts. Springer, Cham. https://doi.org/10.1007/978-3-319-16751-0_37
Download citation
DOI: https://doi.org/10.1007/978-3-319-16751-0_37
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-16750-3
Online ISBN: 978-3-319-16751-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)