
Abstract
We measured soil properties, vertical accretion, and nutrient (organic C, N, and P) accumulation across a range of habitats to evaluate spatial variability of soil properties and processes of alluvial floodplain wetlands of the Altamaha River, Georgia, USA. The habitats vary in elevation and distance from the river channel, creating differences in the depth and duration of inundation. Habitats closer to the river had lower bulk density and higher total P than habitats further removed. 137Cs and 210Pb accretion rates were also greater at sites closer to the channel. Mineral sediment deposition and nutrient accumulation were greater in sloughs closer to the channel and lower in elevation relative to other habitats. We found distance to be a significant predictor of mineral soil properties across the floodplain. Bulk density increased whereas TP and silt content decreased with distance from the river channel. 137Cs accretion, P accumulation, and mineral sediment deposition also decreased with distance from the main channel. Elevation was not a significant predictor of soil properties or processes measured. Long-term (100 year) sediment accumulation rates based on 210Pb were significantly higher than 50-year rate of sedimentation based on 137Cs, perhaps as the result of greater land clearing for agriculture and lack of best management practices in the southeastern USA prior to 1950. Distance from the main channel is the driving force behind the spatial variability of soil properties and processes measured; however, slough habitats closest to the channel and lowest in elevation relative to other habitats maintain distinct vegetation patterns and are hotspots for N, P, and sediment accumulation. Characterization of soil properties and processes of alluvial floodplain forests and other wetlands should take into consideration microtopographic and spatial variation across the wetland.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
4.1 Introduction
Freshwater wetland soils are important sinks for sediment, organic carbon (C), nitrogen (N), and phosphorus (P) (Brinson et al. 1981a, b; Burke 1975; Craft and Casey 2000; Hopkinson 1992; Martin and Hartman 1987; Mausbach and Richardson 1994; Naiman and Décamps 1997), and a wetland’s ability to trap sediment and accumulate nutrients depends on its connectivity to other systems (Mitsch and Goselink 2000). Wetlands such as riparian and alluvial floodplain forests are hydrologically open systems, with larger catchment areas and higher anthropogenic inputs (e.g., nutrients and sediment) than hydrologically closed systems, such as depressional wetlands (Craft and Casey 2000), and as a result, alluvial floodplain wetlands act as nutrient and sediment sinks, reducing sediment deposition and eutrophication downstream in receiving water bodies (Noe and Hupp 2005).
The alluvial floodplain forest floor is a heterogeneous mix of hummocks, hollows, and other microtopographic features that are defined by differences in hydroperiod and soil type (Wharton 1978). The small-scale differences in elevation between microtopographic sites have important implications for ecosystem structure and function (Courtwright and Findlay 2011). Microsites are associated with the maintenance of species diversity in both upland and wetland forest ecosystems (Ehrenfeld 1994). Microtopography influences the frequency and duration of flooding (Rheinhardt 1992) and creates variation in biogeochemical cycling (Darke and Walbridge 2000). Microtopographic relief results in adjacent areas of aerobic and anaerobic soil, which facilitate nutrient cycling (Bridgham et al. 2001; Mosier et al. 2009) including carbon sequestration and N cycling and removal (Wolf et al. 2011). Higher rates of litterfall production and phosphorus and cation (K, Mg) circulation have been found on lower, wetter sites in the floodplain relative to higher, drier sites (Schilling and Lockaby 2005). Studies of alluvial floodplain forest soils to date have focused on the effects of topographic heterogeneity and microtopography on plant communities, net primary production (NPP), and nutrient cycling (Ehrenfeld 1994; Schilling and Lockaby 2005, 2006; Courtwright and Findlay 2011; Wolf et al. 2011), but none have investigated relationships between microtopography and sediment deposition, carbon sequestration, or nutrient (N and P) accumulation.
Using feldspar marker layers, Noe and Hupp (2005) found that sediment and nutrient (C, N, P) accumulation was greater in hydraulically connected floodplain wetlands than in floodplains (Virginia-Maryland-Delaware, USA) where the river was channelized and levees emplaced. Kroes et al. (2007) investigated sediment accretion using the same methods in one of the same rivers (Pokomoke MD). They reported no difference in accretion (mm/year) across the floodplain. Percent organic matter increased with distance from the river, suggesting that mineral sediment deposition decreases with distance from the channel (Kroes et al. 2007). In tidal marshes, sedimentation and nutrient accumulation tend to be greater adjacent to the tidal creek as compared to interior areas (Craft 2007; Loomis and Craft 2010; Craft 2012). Also in tidal marshes, studies indicate that sedimentation is a function of depth and duration of tidal inundation (Morris et al. 2002) and distance from the creek (Temmerman et al. 2005). However, direct measurements are lacking for nontidal alluvial forested wetlands.
We measured soil accretion and nutrient and sediment accumulation across four microtopographic habitats spanning the gradient from the river to upland, in an alluvial floodplain forest along the Altamaha River, Georgia, USA. The four habitats vary in elevation and distance from the river and presumably differ in depth, duration, and frequency of inundation. We hypothesize that sediment deposition and phosphorus accumulation will decrease with distance from the main channel and with elevation in the floodplain. We also hypothesize that carbon sequestration and nitrogen accumulation will be greatest in the areas of lowest elevation.
4.2 Methods
4.2.1 Site Description
The study site is located on the Altamaha River in Griffin Ridge Wildlife Management Area, Long Co., southeastern Georgia (31°68′ N, 81°82′ W) (Fig. 4.1). The Altamaha is a red water river high in sediment and mineral nutrients derived from the oxidized clay soils of the piedmont, which are reddish in color (Conner et al. 2007). The Altamaha drains nearly 23 million hectares of the piedmont and coastal plain, with an average discharge of 166.3 m3 s−1 and sediment yield of 57.1 kg ha−1year−1 (Schilling and Lockaby 2006) (Altamaha River #02226000 1939-2009; USGS 2010).

Map of Griffin Ridge Wildlife Management Area in relation to the Piedmont Region and Georgia Coast
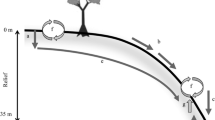
Four habitats were sampled along a cross-sectional gradient of the floodplain: levee, slough, floodplain, and upper slough. Each habitat differs in elevation and distance from the main river channel and hence in the degree of inundation and alluvial inputs from floodwaters (Fig. 4.2). Vegetation is dominated by Platanus occidentalis L. (sycamore) and Betula nigra L. (river birch) along the levee; Nyssa ogeche Bartr. ex Marsh. (Ogeechee gum), Nyssa aquatic L. (water tupelo), Taxodium distichum L. (bald cypress), and Taxodium ascendens Brongn. (pond cypress) in the sloughs; and Fraxinus pennsylvanica Marsh. (green ash), Acer rubrum L. (red maple), and Quercus spp. (oak) in floodplain locations (Craft pers. obs.). Soils consist of Tawcaw series (somewhat poorly drained, fine, kaolinitic, thermic Fluvaquentic Dystrudepts) in the higher-lying areas (e.g., levee and floodplain) with a Chastain series (poorly drained, fine, mixed, semiactive, acid, thermic Fluvaquentic Endoaquepts) in the sloughs and lower-lying positions (USDA 2010). Sample collection sites were located in areas close to the river and within the floodplain. However, Griffin Ridge, a dry, elevated sand ridge that straddles the floodplain, was excluded from sampling due to soils and vegetation not characteristic of wetland areas.

Schematic of the four landscape positions sampled relative to typical spring peak floodwater levels
4.2.2 Soil Sampling and Analysis
Soil cores, 8.5 cm diameter by 60 cm deep, were collected from each habitat. Two cores each were collected from the slough and upper slough habitats, while three cores each were collected from the levee and floodplain habitats (Fig. 4.2.). Cores were sectioned in the field with the top 30 cm separated into 2-cm increments and the bottom 30 cm into 5-cm increments. Each increment was air dried, weighed for bulk density, and then ground and sieved through a 2-mm mesh screen. A subsample was oven-dried at 85 °C, for determination of moisture content, organic carbon, total nitrogen, and total phosphorus. Bulk density was calculated from the dry weight per unit volume for each depth increment after correcting for moisture content with the oven-dried subsample (Blake and Hartge 1986). Organic C and N were determined using a PerkinElmer 2400 CHN analyzer (PerkinElmer Corporation, Waltham, MA). Analysis of an in-house standard (% C = 6.1, % N = 0.365) returned average recovery rates of 103 % and 102 % for C and N, respectively. Total P was determined by colorimetric analysis following digestion with HNO3–HClO4 (Sommers and Nelson 1972). Analysis of estuarine sediment (NIST #1646a) returned an average recovery rate of 88 % for total P. Percent sand, silt, clay, and organic matter (0–10 cm) were determined by the hydrometer method (Gee and Bauder 1986).
Soil accretion was determined using ground and sieved soil increments packed into 50 mm-diameter by 9 mm-deep petri dishes which were analyzed for 137Cs and 210Pb by gamma analysis of the 661.62 keV and 46.5 keV photopeaks, respectively (Craft and Casey 2000). The 137Cs maximum in each core corresponds with the peak fallout from ground thermonuclear weapons testing, which occurred in 1964. 210Pb is a naturally occurring radionuclide (half-life of 22 years) that can be used to estimate soil accretion during the past 100–150 years (Craft and Casey 2000). The constant activity model was used to determine accretion using the distribution of excess 210Pb (total 210Pb minus background 210Pb) (Oldfield and Appleby 1984; Schelske et al. 1988). Accumulation of organic C, N, and P was calculated using the 137Cs and 210Pb accretion rates, bulk density, and nutrient concentrations (C, N, and P) down to the increment containing the peak 137Cs and excess 210Pb activities, respectively, for each core.
4.2.3 GIS/LiDAR Distance and Elevation Calculation
Elevations of the four habitats were estimated through interactive examination of a LiDAR dataset of the area (Photo Science and Fugro EarthData, Inc. 2010). A 10 m-resolution surface model was generated by using NOAA’s Digital Coast Data Access Viewer to filter the average, last return points from the 2010 Coastal Georgia Topographic LiDAR dataset for the study area (NOAA 2011). The dataset was queried using ArcGIS 10 Desktop software (ESRI 2011) for 10 m areas along the transect to estimate the elevations (relative to NAVD88) for the primary topographic features noted in the field (levee, sloughs, and floodplain).
4.2.4 Statistical Analysis
Bulk soil properties, accretion, and accumulation were analyzed using a one-way analysis of variance with habitat as the main effects variable. Student’s t-test was used to test for differences in 137Cs versus 210Pb accretion and accumulation. Tests of normality and homoscedasticity revealed that the data were normally distributed and estimate a common variance. Regression analysis was used to test the effects of distance from main channel and elevation on soil properties, accretion, and accumulation (SAS 1996). All tests of significance were conducted at α = 0.05.
4.3 Results
Bulk soil properties (0–30 cm) varied among habitats across the Altamaha River floodplain. Bulk density was greater at sites furthest from the river, in the upper floodplain and upper slough (1.2 g cm−3), and lowest in the slough (0.75 g cm−3) (p < 0.02, F = 7.5) (Table 4.1). There was no difference in percent organic carbon or total nitrogen among habitats. Total P, however, was higher in the slough (530 μg/g) than in the upper slough and floodplain (120 μg g−1) (p < 0.05, F = 7.1) (Table 4.1). Soils (0–10 cm) were composed predominantly of sand particles (63–82 %) with lesser amounts of silt (10–24 %), clay (3–7 %), and organic matter (5–10 %) (Fig. 4.3). There was no significant difference in particle size fractions among habitats.

Particle-size distribution and organic matter content (0–10 cm) of soils across the floodplain of the Altamaha River
137Cs profiles revealed interpretable peaks, corresponding to 1964, of the maximum 137Cs fallout from aboveground nuclear weapons tests. 210Pb also exhibited interpretable total and excess 210Pb concentrations, decreasing with depth corresponding to its exponential decay with time (Fig. 4.4). 137Cs soil accretion ranged from 0.2 to 3.3 mm year−1 and was greater in the slough than in the floodplain and upper slough (p < 0.01, F = 12.2) (Fig. 4.5). 137Cs organic C sequestration ranged from 7 to 61 g m−2 year−1 (Fig. 4.5). Total N and P accumulation ranged from 0.4 to 5.4 g N m−2 year−1 and 0.04 to 1.15 g P m−2 year−1. As with accretion, N (p < 0.05, F = 6.4) and P (p < 0.01, F = 20.9) accumulation were significantly greater in the slough than the upper slough and floodplain (P only). Sediment deposition on the levee and in the slough also was greater relative to habitats (floodplain, upper slough) far removed from the main channel (p < 0.01, F = 12.2) (Fig. 4.6).

Representative 137Cs and 210Pb profiles for 1 L levee site

Mean 137Cs- and 210Pb-based soil accretion (±1 S.E.) across the floodplain of the Altamaha River. Different letters indicate significant differences (p < 0.05) according to ANOVA analysis

Mean (a) carbon sequestration and accumulation of (b) nitrogen, (c) phosphorus, and (d) mineral sediment (±1 S.E.) across the floodplain of the Altamaha River. Different letters indicate significant differences (p < 0.05) according to ANOVA analysis
Like 137Cs, 210Pb accretion was significantly greater in the slough (p < 0.05, F = 5) than in the upper slough and the levee as well. C sequestration ranged from 34 to 68 g C m−2 year−1, while N and P accumulation ranged from 2.7 to 7.8 g N m−2 year−1 and 0.41 to 1.5 g P m−2 year−1. The slough also exhibited greater P accumulation than other habitats (p < 0.05, F = 7.2). Likewise, N accumulation was greater in the slough than the levee and upper slough habitats (p < 0.05, F = 6.5). No differences were found in 210Pb sediment deposition (1,170–2,760 g m−2 year−1) across the floodplain.
Mean rates of 210Pb soil accretion (2.5 ± 0.6 mm year−1) were consistently but not significantly higher than accretion based on 137Cs (1.6 ± 0.6 mm/year). However, 210Pb accumulation of mineral sediment (1,870 ± 300 g m−2 year−1) was significantly greater than sedimentation rates based on 137Cs (1,030 ± 250 g m−2 year−1) (p < 0.05). Like accretion, 210Pb accumulation of organic C, N, and P was consistently higher than rates based on 137Cs but not significantly so.
Multiple regression analysis revealed that soil properties and processes associated with mineral fraction were significantly related to distance from the main channel. Bulk density increased with distance from the main channel (r 2 = 0.78, p = 0.002), whereas silt content (r 2 = 0.43, p = 0.05), 137Cs-based soil accretion (r 2 = 0.68, p = 0.006), P accumulation (r 2 = 0.69, p = 0.006), and mineral sediment deposition (r 2 = 0.75, p = 0.002) decreased with distance from the channel, though 210Pb accretion and accumulation rates were not predicted by distance. We found no relationship between soil elevation and surface soil properties or processes.
4.4 Discussion
4.4.1 Spatial Patterns
Microtopography is known to affect wetland plant productivity and diversity and biogeochemical processes, yet little research has investigated its effect on sedimentation and nutrient accumulation processes. It has been shown that small variations in elevation can affect wetland structure and function, such as nutrient flow and redox conditions (Courtwright and Findlay 2011), diversity of plant communities (Ehrenfeld 1994), and biogeochemical cycling (Rheinhardt 1992). Surprisingly, in our study, we observed no relationship between elevation and soil properties or processes at our study site. This may be attributed in part to the small change in elevation relative to distance among our four habitats (≈1 m.).
Shilling and Lockaby (2005) investigated vegetation and soil properties within floodplains on the Satilla and Altamaha rivers, GA, and found lower bulk density at lower, wetter habitats relative to higher, drier habitats. We also found lower bulk density in the slough than in higher, drier habitats, but we observed no relationship between bulk density and elevation in our study. In the Shilling and Lockaby (2005) study, bulk density ranged from 0.9 g cm−3 in wetter habitats to 1.3 g cm−3 in intermediate habitats, as compared to bulk density of 0.8 g cm−3 in the slough and 1.2 g cm−3 in the floodplain measured in our study. Shilling and Lockaby (2005) also found that habitats intermediate between their wet and dry sites had significantly more percent soil organic C, but they observed no difference in soil N among their habitats. We observed no difference in percent organic C or N among our habitats. Using a high-precision elevation model, Drouin et al. (2011) also found that alluvial wetlands in the St. Lawrence River basin, Vermont, much like ours, that are separated from the channel by a natural levee with a parallel slough exhibit no significant difference in percent organic C produced by differences in hydroperiod in sloughs (1.5–1.75 %) versus higher elevation habitats (0.75–1.0 %). Soil type is another factor that creates variation in microtopography, producing areas differing in water-holding capacity and mineral composition which lead to distinct plant communities (Wharton 1978). Our floodplain forest exhibits little difference in soil texture (sand, silt, clay, and OM) among habitats, which may further explain a lack of significant differences in C and N accumulation between habitats, as the accumulation of these elements is highly associated with variation in soil organic matter (Brunet and Astin 1997; Noe and Hupp 2005).
Connectivity to the river channel is also known to affect soil processes, notably sediment and P deposition. For example, Noe and Hupp (2005), measuring accretion over feldspar clay marker horizons, found that floodplains that were more connected to a water source had greater amounts of mineral sediment and P accumulation. We also found that habitats closer to the main channel had higher sediment deposition and P accumulation than those far removed from the channel (Fig. 4.5). Noe and Hupp (2005) also found that more connected systems had higher rates of organic C and N accumulation, but we did not observe this in our study. Inorganic P is often sorbed to mineral sediments, and it has been shown that P concentrations and the rates of P accumulation are strongly correlated with rates of sediment deposition and that floodplains capturing more sediment will capture more P, with higher P concentrations and accumulation rates close to the main channel and decreasing with distance from the channel (Noe and Hupp 2005; Kroes et al. 2007). We observed this in our study as well, as the positions closest to the main river channel had higher concentrations and accumulation of P and mineral sediment that decreased with distance from the main channel. Distance was a significant predictor of mineral sediment processes, but not for organic C or N, which are driven more by in situ organic matter production and deposition rather than sediment transport.
4.4.2 Temporal Patterns
210Pb sediment accumulation was significantly higher than rates based on 137Cs, which suggests greater sediment deposition in the floodplain in the past 100 years than in the past 50 years. A similar pattern was observed in Georgia floodplain wetlands that drain to the Gulf of Mexico, which was attributed to historical changes in land use and demographic trends in the watershed, and the more recent implementation of BMPs reducing erosion and sediment runoff (Craft and Casey 2000). Georgia and the southeastern states saw the greatest erosive land use (ELU) during the period from 1860 to 1920 (Trimble 1974). It was commonplace in agriculture at that time to grow crops in an area until the land was deemed unproductive, then abandon it, and leave it bare (Trimble 1974). In the time after the Civil War, there was an increase in tenant farmers who were also more prone to poor land-use practices than landowners. This severe abuse of the land promoted rapid soil loss that filled streams and severely altered water flows in the area. Streams topped their banks changing bottomland fields to bottomland swamps, pushing cultivation to upland areas, and in the process creating more erosion (Trimble 1974).
In the twentieth century, ELU decreased greatly over all of the Southern Piedmont, where the headwaters of the Altamaha River are located, and was low everywhere by 1967. The greatest decreases of ELU in the area occurred in regions including the Piedmont areas of Georgia (Trimble 1974). During this period, there was a large decrease in harvested cropland across the Southern Piedmont. The abandonment of land deemed unsuitable for crops, the effects of the boll weevil making cotton cultivation difficult and expensive, a drop in cotton prices, modern technology allowing for fewer acres of corn and oats necessary, and government controls on crop acreage all contributed to the decrease in farm acreage (Trimble 1974). This was also paired with an increase in land conservation as abandoned agricultural fields were allowed, or promoted, to convert back into forest. Practices such as crop rotation, contour plowing, and terracing were also widely implemented, and these new practices contributed to a large decrease in erosion across the Piedmont area (Trimble 1974). These land-use changes along with a shift in agricultural practices reduced the sediment load into streams and wetlands, accounting for greater sediment input in the years before their implementation.
Our limited sample size (n = 10 cores across the floodplain) limits the conclusions we can draw. However, our findings of differences in mineral soil properties (and lack of differences in organic soil properties) are supported by other studies (Schilling and Lockaby 2005; Noe and Hupp 2005; Franklin et al. 2009; Drouin et al. 2011). Furthermore, our measured rates of C sequestration and N, P, and sediment accumulation are within the range of other published studies of nutrient accumulation and sediment deposition in floodplain wetlands (Table 4.2).
In conclusion, distance from the main river channel was a significant predictor of mineral soil properties. Bulk density increased with distance from the channel, and processes such as P accumulation and sediment deposition decreased with distance. Elevation was not a strong predictor of soil properties or processes across the floodplain. Sloughs, especially those nearest to the main channel, which are the lowest sites within our floodplain, were hotspots of sediment and nutrient accumulation as evidenced by greater soil accretion, as well as greater N and P concentrations and accumulation, and mineral sediment deposition. In recent years, there has been increasing emphasis on quantifying soil properties and processes at a landscape scale, especially C stocks, C sequestration, and nutrient removal. This study adds to a body of literature which suggests a high degree of small-scale heterogeneity across alluvial landscapes, leading to spatial variability in soil properties and processes. Thus, scaling measurements from a single habitat across a heterogeneous alluvial floodplain wetland may over- or underestimate nutrient accumulation and retention.
In order to accurately inventory soil C and nutrient stocks, studies of soil properties and processes of alluvial floodplains and other wetlands should account for spatial variability, especially distance, across the wetland. Similarly, it is important to recognize temporal variability in sedimentary process (e.g., the measured difference in long-term 210Pb versus 50-year 137Cs rates) that relates to land-use change when attempting to project soil process like C sequestration and nutrient accumulation by these ecosystems in the future.
References
Blake, G. R., & Hartge, K. H. (1986). Bulk density. In A. Klute (Ed.), Methods of soil analysis. Part 1. Physical and mineralogical methods (Agronomy monograph 9 2nd ed., pp. 363–375). Madison: ASA and SSSA.
Bridgham, S., Johnston, C. A., Schubauer-Berigan, J. P., & Weishampel, P. (2001). Phosphorus sorption dynamics in soils and coupling with surface and porewater in riverine wetlands. Soil Science Society of America Journal, 65, 577–588.
Brinson, M. M., Swift, R. C., Plantico, C., & Barclay, J. S. (1981a). Riparian ecosystems: Their ecology and status. U. S. Fish and Wildlife Service, Biological Services Program, Washington, DC, USA. FWS/OBS-81/17.
Brinson, M. M., Lugo, A. E., & Brown, S. (1981b). Primary productivity, decomposition and consumer activity in fresh water wetlands. Annual Review of Ecological Systems, 12, 123–161.
Brown, S. L. (1978). A comparison of cypress ecosystems in the landscape of Florida. PhD dissertation. Gainesville, FL, USA: University of Florida.
Brunet, R. C., & Astin, K. B. (1997). Spatio-temporal variations in sediment nutrient levels: The River Adour. Landscape Ecology, 12, 171–184.
Burke, W. (1975). Fertilizer and other chemical losses in drainage water from blanket bog. Irish Journal of Agricultural Research, 14, 163–178.
Conner, W. H., Doyle, T. W., & Krauss, K. R. (2007). Ecology of tidal forested wetlands of the Southeastern United States. New York: Springer.
Courtwright, J., & Findlay, S. E. G. (2011). Effects of microtopography on hydrology, physicochemistry, and vegetation in a tidal swamp of the Hudson River. Wetlands, 31, 239–249.
Craft, C. B. (2007). Freshwater input structures soil properties, vertical accretion and nutrient accumulation of Georgia and U.S. tidal marshes. Limnology and Oceanography, 52(3), 1220–1230.
Craft, C. B. (2012). Tidal freshwater forest accretion does not keep pace with sea level rise. Global Change Biology, 18, 3615–3623.
Craft, C. B., & Casey, W. P. (2000). Sediment and nutrient accumulation in floodplain and depressional freshwater wetlands of Georgia, USA. Wetlands, 20, 323–332.
Darke, A. K., & Walbridge, M. R. (2000). Al and Fe biogeochemistry in a floodplain forest: Implications for P retention. Biogeochemistry, 51, 1–32.
Drouin, A., Saint-Laurent, D., Lavoie, L., & Oullet, C. (2011). High-precision elevation model to evaluate the spatial distribution of soil organic carbon in active floodplains. Wetlands, 31, 1151–1164.
Ehrenfeld, J. G. (1994). Microtopography and vegetation in Atlantic white cedar swamps: The effects of natural disturbances. Canadian Journal of Botany, 73, 474–484.
ESRI (Environmental Systems Resource Institute). (2011). ArcMap 9.3. Redlands: ESRI.
Franklin, S. B., Kupfer, J. A., Pezeshki, S. R., Gentry, R., & Smith, R. D. (2009). Complex effects of channelization and levee construction on western Tennessee floodplain forest function. Wetlands, 29(2), 451–464.
Gee, G. W., & Bauder, J. W. (1986). Particle-size analysis. In A. Klute (Ed.), Methods of soil analysis, part 1 (pp. 383–411). Madison: American Society of Agronomy.
Hopkinson, C. S. (1992). A comparison of ecosystem dynamics in fresh water wetlands. Estuaries, 15, 549–562.
Johnston, C. A., Bubenzer, G. D., Lee, G. B., Madison, F. W., & McHenry, J. R. (1984). Nutrient trapping by sediment deposition in a seasonally flooded lakeside wetland. Journal of Environmental Quality, 13, 283–290.
Kleiss, B. A. (1996). Sediment retention in a bottomland hardwood wetland in eastern Arkansas. Wetlands, 16, 321–333.
Kroes, D. E., Hupp, C. R., & Noe, G. B. (2007). Sediment, nutrient, and vegetation trends along the tidal, forested Pocomoke River, Maryland. In W. H. Conner, T. W. Doyle, & K. W. Krauss (Eds.), Ecology of tidal freshwater forested wetlands of the southeastern United States (pp. 113–137). New York: Springer.
Kuenzler, E. J., Mulholland, P. J., Yarbro, L. A., Smock, L. A. (1980). Distributions and budgets of carbon, phosphorus, iron and manganese in a floodplain swamp ecosystem. Water Resources Research Institute of the University of North Carolina, Raleigh, NC, USA. Report no. 157.
Loomis, M. J., & Craft, C. B. (2010). Carbon sequestration and nutrient (nitrogen, phosphorus) accumulation in river-dominated tidal marshes, Georgia, USA. Soil Science Society of America Journal, 74(3), 1028–1036.
Martin, D. B., & Hartman, W. A. (1987). Correlations between selected trace elements and organic matter and texture in sediments of northern prairie wetlands. Journal of the Association of Official Analytical Chemists, 70, 916–919.
Mausbach, M. J., & Richardson, J. L. (1994). Biogeochemical processes in hydric soil formation. Current Topics and Wetland Biogeochemistry, 1, 68–127.
Mitsch, W. J., & Goselink, J. G. (2000). The value of wetlands: Importance of scale and landscape setting. Ecological Economics, 35, 25–33.
Mitsch, W. J., Dorge, C. L., & Wiemhoff, J. R. (1979). Ecosystem dynamics and a phosphorus budget of an alluvial cypress swamp in southern Illinois. Ecology, 60, 1116–1124.
Morris, J. T., Sundareshwar, P. V., Nietch, C. T., Kjerfve, B., & Cahoon, D. R. (2002). Response of coastal wetlands to rising sea level. Ecological Applications, 83, 2869–2877.
Mosier, K., Ahn, C., & Noe, G. (2009). The influence of microtopography on soil nutrients in created mitigation wetlands. Restoration Ecology, 17, 641–651.
Naiman, R. J., & Décamps, H. (1997). Ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 28, 621–658.
National Oceanic and Atmospheric Administration’s Digital Coast Data Access Viewer. http://www.csc.noaa.gov/dataviewer. 18 Nov 2011.
Noe, G. B., & Hupp, C. R. (2005). Carbon, nitrogen, and phosphorus accumulation in floodplains of Atlantic Coastal Plain Rivers, USA. Ecological Applications, 15(4), 1178–1190.
Oldfield, F., & Appleby, P. G. (1984). Empirical testing of 210Pb models for dating lake sediments. In E. Y. Haworth & J. W. G. Lund (Eds.), Lake sediments and environmental history (pp. 93–124). Minneapolis: University of Minnesota Press.
Photo Science and Fugro EarthData, Inc. (2010). Coastal Georgia Elevation Project Lidar Data, 1:2,400 scale, digital LiDAR data.
Rheinhardt, R. (1992). A multivariate analysis of vegetation patterns in tidal freshwater swamps of Lower Chesapeake Bay, USA. Bulletin of the Torrey Botanical Club, 119, 192–207.
SAS Institute. (1996). SAS user’s guide: Statistics. Cary: SAS Institute, Inc.
Schelske, C. L., Robbins, J. A., Gardner, W. D., Conley, D. J., & Bourbonniere, R. A. (1988). Sediment record of biogeochemical responses to anthropogenic perturbations of nutrient cycles in Lake Ontario. Canadian Journal of Fisheries and Aquatic Sciences, 45, 1291–1303.
Schilling, E. B., & Lockaby, B. G. (2005). Microsite influences on productivity and nutrient circulation within two southeastern floodplain forests. Soil Sciences Society of America Journal, 69, 1185–1195.
Schilling, E. B., & Lockaby, B. G. (2006). Relationships between productivity and nutrient circulation within two contrasting southeastern U.S. floodplain forests. Wetlands, 26, 181–192.
Sommers, L. E., & Nelson, D. W. (1972). Determination of total phosphorus in soils: A rapid perchloric acid digestion procedure. Soil Science Society of America Journal, 36, 902–904.
Temmerman, S., Bouma, T. J., Govers, G., & Lauwaet, D. (2005). Flow paths of water and sediment in a tidal marsh: Relations with marsh development stage and tidal inundation height. Estuaries, 28, 338–352.
Trimble, S. W. (1974). Man-induced soil erosion on the Southern Piedmont 1700–1970. Soil Conservation Society of America. Milwaukee, WI: University of Wisconsin, Department of Geography.
USDA (U.S. Department of Agriculture). (2010). Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. http://websoilsurvey.usda.gov/. Last accessed 12 July 2010.
USGS (U.S. Geological Survey). (2010). Georgia real-time stream flow data. [Online] http://waterdata.usgs.gov/ga/nwis/current?type = flowgroup_key = basin_cd. Last accessed 12July2010. USGS, Reston, VA.
Wharton, C. H. (1978). Natural environments of Georgia. Geologic and Water Resources Division and Resource Planning Section, Office of Planning and Research, Georgia Department of Natural Resources, Atlanta, GA.
Wolf, K. L., Ahn, C. W., & Noe, G. B. (2011). Microtopography enhances nitrogen cycling and removal in created mitigation wetlands. Ecological Engineering, 37, 1398–1406.
Acknowledgments
We thank Nathan Knowles for his help in sample collection, Sarah Sutton for her help with sample analysis and input into the methods section, Jeff Ehman for his help acquiring the GIS/LiDAR elevation and distance datasets, and Anya Hopple and Anne Altor for edits to early versions of the manuscript. This research was supported by the US Department of Energy through grant #TUL-563-07/08 and the National Science Foundation grant #OCE-9982133 to the Georgia Coastal Ecosystems Long-Term Ecological Research Program. This is contribution number 1037 from the University of Georgia Marine Institute.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Bannister, J.M., Herbert, E.R., Craft, C.B. (2015). Spatial Variability in Sedimentation, Carbon Sequestration, and Nutrient Accumulation in an Alluvial Floodplain Forest. In: Vymazal, J. (eds) The Role of Natural and Constructed Wetlands in Nutrient Cycling and Retention on the Landscape. Springer, Cham. https://doi.org/10.1007/978-3-319-08177-9_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-08177-9_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-08176-2
Online ISBN: 978-3-319-08177-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)