
Abstract
Lipopolysaccharide (LPS, endotoxin) is a major component of the outer membrane of gram negative bacteria. It is an activator of humoral and cellular responses in humans with potential use as adjuvant in vaccine technology. Importantly, LPS has a large capacity to induce Th1-type responses and stimulate cytotoxic T lymphocytes, which are poorly obtained by standard adjuvants but required for specific immune stimulatory therapies. In contrast, LPS possess an extreme toxicity that limit its clinical use in humans. Alteration of its chemical structure led the generation of LPS-based derivatives with reduced toxicity but retaining adjuvant properties. Monophosphoryl lipid A (MPLA) has been the most successful LPS-based adjuvant, currently incorporated in approved vaccine preparations and extensively used in vaccine trials and preclinical studies. Novel designed structures, analogous to LPS and generated by chemical synthesis, can offer lower production cost and lesser heterogenic formulations than MPLA and, in addition, be even most suitable for specific immune therapies. Thus, LPS-based structures are valuable contributions as adjuvants in human vaccinology and open new possibilities to existing demands for specific therapies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
The development of subunit vaccines has improved the safety of human vaccine prophylaxis but requires strong adjuvants. Alum (diverse aluminum salts) is the most used adjuvant with an acceptable profile of side effects and induction of optimal protection against many human pathogens. However, alum is not suitable against certain pathogens or for new vaccine therapies like cancer or allergy. Lipopolysaccharide is a component of the outer membrane of gram-negative bacteria largely studied as adjuvants by their inherent ability to stimulate immune responses. In contrast, LPS induces inacceptable toxic effects in humans.
The generation of new detoxified LPS species with higher adjuvant characteristics than alum and acceptable toxicity has opened new perspectives to the vaccination. In this book chapter, the basic aspects of LPS structure, toxicity, and activation of the immune system are first introduced. Next, adjuvant characteristics of alum and corresponding drawbacks are briefly cited. Followed, the most promising detoxified LPS molecules and adjuvant characteristics are further discussed with special emphasis in current advances. A final overview section summarized the most relevant points.
1 LPS Structure and Biological Activity
Lipopolysaccharide is a component of the external leaflet of the outer membrane of gram-negative bacteria. It is a complex glycolipid formed by three domains, a fatty acid-rich domain (lipid A), an oligosaccharide domain (core), and a repeating oligosaccharide domain (O-antigen) [1]. Figure 33.1 represents the typical LPS organization.

General chemical structure of LPS of gram-negative bacteria. The LPS structure of Escherichia coli is depicted. LPS consists of three regions: lipid A, core, and O-antigen. The chemical structure of the lipid A of E. coli that displays the maximal immunostimulatory or endotoxic activities in humans (the topic of this work) is further detailed (black colored). Additionally, those substituents that possess the lipid A of Salmonella minnesota used to obtain MPL are blue colored. Residues of core and O-antigen region are schematized and abbreviated as KDO 2-keto-3-deoxyoctonoic acid, Hep d-glycero-d -manno-heptose, Glu d-glucose, Gal d-galactose, GluNAc N-acetyl-glucosamine, and GalNAc N-acetyl-galactosamine. The length of the O-antigen depends on the number of repeating units. Additional substituents or modifications can be found in nature, but they are not shown for clarity
The lipid A domain is a β-1, 6-linked d-glucosamine disaccharide linked to variable number of ester- and amide-linked 3-hydroxy fatty acids and phosphate groups (Fig. 33.1). Its architecture is highly conserved, but different microorganisms may present variations in the number and length of the fatty acid side chains, the presence of terminal phosphate residues, and associated modifications. The core domain is a branched oligosaccharide region formed by nine or ten sugars, and its composition is more variable between species than lipid A. Finally, the O-antigen, if present, is the most variable of the tree domains and consists of up to 50 repeating oligosaccharide units formed of 2–8 monosaccharide moieties. In addition, certain modifying enzymes can alter the composition of LPS contributing to increase the LPS heterogeneity [2–5].
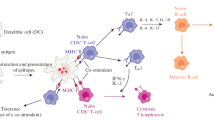
Toll-like receptors (TLR)s belong to a family of receptors that recognize a broad diversity of specific but conserved structures of pathogen microorganisms [6]. Immediately after stimulation, TLRs initiated the activation of immune defense mechanisms. Toll-like receptor 4 (TLR4), in complex with the glycoprotein MD-2, constitutes the LPS receptor [7, 8]. TLR4 is a membrane-spanning protein present on antigen-presenting cells (APC) (macrophages and dendritic cells) and epithelial cells of humans. Its stimulation requires the cooperation of associated molecules, like the LPS-binding protein and CD14 that facilitate LPS transfer to the receptor. TLR4 stimulation induces the formation of intracellular protein complexes that leads to the activation of intracellular signaling cascades [9, 10]. These reactions trigger the biosynthesis and secretion of diverse proinflammatory cytokines (IL-1, IL-8, IL-12, TNFα, and IFNγ) and the production of co-stimulatory molecules [11] that finally activates humoral and cellular responses including activation of the complement system [12, 13], activation of macrophages [14, 15] B and T cells, and enhancement of cellular cooperation [11]. Consequently, this response is beneficial for the control of local infections. In contrast, high LPS dose, specially released to the blood system during sepsis, leads to large secretion of cytokines and inflammatory mediators with severe [16, 17] and/or fatal consequences [18, 19]. Table 33.1 summarizes the beneficial and harmful effects of LPS in humans, but for further details see revision [20, 21]. In summary, LPS is a strong activator of the immune system (adjuvant) but also a highly toxic substance (endotoxin).
The lipid A region is the major responsible of the TLR4 stimulation. Variations in its structure, mainly regarding the number and length of acyl acid chains, and the charge are crucial in this regard [22]. The hexa-acylated E. coli lipid A (canonical LPS structure and depicted in Fig. 33.1) with fatty acids of 12–14 carbons and two phosphate residues is the maximal stimulator of human TLR4 (hTLR4) [23, 24]. In contrast, the tetra-acylated lipid IVa with fatty acids of 18–16 carbons and a phosphate residue, an intermediate in the biosynthetic pathway of lipid A, does not stimulate hTLR4 (canonical hTLR4 antagonist) [25].
2 Lipid A Analogous Structures and Its Role as Adjuvants
Vaccine based on infectious attenuated or inactivated whole pathogens contains a large variety of target structures for TLRs and, subsequently, promotes strong and long protection. However, they generate a large variety of side effects even with fatal consequences [26, 27]. Vaccines based on one or certain purified components (subunit vaccines) show acceptable safely but a poor immunogenicity and require additional immune stimulators (adjuvants). Alum refers to several aluminum salts and is the most used adjuvant. It is safe and elicits predominantly a Th2-type antibody response that shows to be effective in a large variety of vaccines [28]. However, alum hardly promotes Th1-type antibody responses [29]. Adjuvants that favor Th1 or more balanced Th1/Th2 responses are required to induce optimal immune protection against certain pathogens [30] or diseases as cancer [31] or allergy [32]. Apart from that, alum poorly stimulates mucosal immunity. Mucosa tissues are the first line of defense against many pathogens and the ecological niche of commensal and opportunistic microorganisms, for example, Neisseria meningitidis.
Therefore, mucosal immunity is considered the gold therapy to evade pathogen colonization and confer herd immunity against certain particular pathogens. Vaccine adjuvants that target mucosal immunization must promote a large series of biological and complex activities as Th17 cell development, APC proliferation, and IgA production [33, 34]. In this regard, several substances have been extensively studied as bacterial toxins or CpG, among others [34], but, till date, no available approved adjuvant exists (with the exception of MPL, to be discussed next). Alternatively to alum, three additional adjuvants were licensed: MF59, AS03, and RC-529. MF59 is an oil in water emulsion with low oil content, and it is included in an approved influenza vaccine [35]. Although MF59 induces a more balanced Th1/Th2 response than alum, it shows partial efficacy, often requiring the coadministration of Th1 enhancers. ASO3 and RC-529 contain LPS-based substances to be discussed later.
LPS has attracted large attention as adjuvant by its high capacity to induce Th1-type responses against coadministrated antigens. One of the most relevant factors involved in the development of this response is IL-12. Note that LPS is a stimulator of this interleukin. Interestingly, TLR4 receptors are tactically present at mucosa surfaces; therefore, it would be expected that TLR4 agonists can promote immune responses at local and distal mucosal sites. In the past decades, several strategies were followed to reduce its extreme toxicity without altering this inherent capacity; variation of the LPS composition, in particular lipid A, by chemical treatments and chemical synthesis of lipid A analogues is a good example (see detailed revision [36]). As a result, a diverse lipid A species was generated, although only few exhibited the desirable properties. Next, the most relevant substances and clinical applications are further discussed.
The chemical hydrolysis of the LPS of S. minnesota which contains a lipid A with seven acyl chains and three phosphate groups as depicted in Fig. 33.1 (with blue-colored substituent), generated one of the most successful LPS-based adjuvants, the monophosphoryl lipid A (MPL). This derivate structure is a six-acyl side chain lipid A with one phosphoryl group [37] (see Fig. 33.2). MPL demonstrated to be less toxic than the parent (0.1% of toxicity) [38] with a toxic side effect profile comparable to alum [39, 40] while retaining the stimulatory properties of LPS. At present, MPL is adjuvant of approved vaccine preparations for humans in Europe (human papillomavirus (Cervarix) [33, 34] and pollen allergy (Pollinex Quattro) [41, 42]) and Australia (hepatitis B virus (Fendrix) [43]), and it has been used extensively in human vaccine trials for several infectious diseases like malaria [44], tuberculosis, [45, 46] or tumor growth [47].

Chemical structure of relevant LPS-based adjuvants
Because MPL is highly hydrophobic and generates aggregates in aqueous solution that may considerably affect the TLR4 activation, it is often formulated in combination with alum or other delivery systems [48]. These combinations, together with other factors (accompanying antigen or administration route), can alter its adjuvant action. For example, in aqueous formulations MPL promotes antibody production, while in oil in water emulsions, it better stimulates T-cell responses. In contrast, MPL combined with other delivery systems is a strong stimulator of cytotoxic T lymphocyte proliferation. Delivery systems can also modify their biological properties. Liposomes are spherical vesicles formed by phospholipid bilayers extensively used to deliver antigens in its native conformation. Incorporation of MPL into liposomes reduced the residual toxicity of MPL but retained intact its adjuvant potential [49]; this effect was also observed with other detoxified LPS species [50]. Therefore, liposomal MPL formulations were extensively studied in human trials for different indications as malaria [49], pneumococcal disease, [51] or genital herpes type 2 [52] and in experimental animals for Streptococcus pyogenes infections [53] or toxin neutralization of E. coli [54]. Finally, it is significant to notice MPL’s success in providing mucosal immunity after mucosal [55] and intramuscular [56, 57] administration; MPL formulations for mucosal vaccines have been extensively explored for the treatment of different diseases; genital herpes [58] and mucosal leishmaniasis [59] are good examples.
One of the main drawbacks attributed to MPL is the heterogeneity of lipid A congeners generated during its production with subsequent purification cost. A solution to this problem could be the generation of synthetic lipid A analogues. Chemical synthesis generates pure and defined structures that reduce production cost. Like natural LPS species, these structures interact with hTLR4. Therefore, numerous analogues with variation in acyl chain length and positions, phosphate groups, or the backbone unit were generated and their biological activity further analyzed. Till date, the most suitable molecules in vaccine development are RC-529, E6020, GLA, and ONO-4007 (see chemical structure in Fig. 33.2).
RC-529 is a synthetic analogue of MPL composed by a monosaccharide backbone with six fatty acyl chains [38]. It is a very attractive adjuvant. Like MPL, it is well tolerated and effective during clinical trials [38] but with a lower production cost. In fact, its use was approved in Argentina in a hepatitis B vaccine. It is often in combination with different delivery systems to enhance its solubility or improve its delivery without affecting immune stimulatory capacities [60]. In addition, several studies performed in experimental animals indicate that RC-529 is an efficient mucosal adjuvant against pathogens that lack effective vaccine therapy. For example, it elicited bactericidal antibodies after intranasal immunization with the Streptococcus pneumoniae protein PppA [61] and the meningogoccal protein P2086, [60] and it promoted high antibody titers in macaques at the nasal and genital mucosa against an HIV peptide immunogen [62]. Similarly, it reduced nasal colonization of nontypable Haemophilus influenzae and Moraxella catarrhalis in mice that were immunized via nasal with recombinant proteins, [63] and it conferred significant protection against lethal influenza challenge [64].
E6020 is an hexa-acylated acyclic backbone [65, 66] with higher biological activities than alum [65, 66] or MPL [65] and with a reduced toxicity [67]. Its simple structure allows the production of high-purity material than other synthetic TLR4 agonists [65]. Various works showed its high capacity to shift immune responses towards a Th1 profile when combined with conventional vaccines [65, 67, 68]. Generation of this immune profile is especially relevant in cancer vaccines. Indeed, E6020 in combination with a monoclonal antibody (trastuzumab) enhanced significantly protection against tumor growth in animal models [69].
GLA is a hexa-acyl synthetic lipid A derivative composed of a disaccharide backbone with a single phosphate group. Results show that it has even more powerful adjuvant abilities than MPL [70, 71], and it exhibited a good safety profile in animals and in Phase I trials [72]. MPL has a strong but not overwhelming ability to promote Th1 responses. Interestingly, GLA exhibits a strong ability to shift antigen-specific immune responses towards Th1 type [73, 74]; hence, it is being proposed as a better alternative to MPL to confer adequate protection against certain pathogens. In fact, significant protection in animals of experimentation was reported against Toxoplasma gondii, [75] Mycobacterium tuberculosis, [76, 77] or influenza virus [72].
ONO-4007 is a tri-acylated acyclic sulphonated backbone. This molecule induced tumor and metastases regression in animal models [78, 79]. This property was due to its strong ability to stimulate secretion of tumor necrosis factor (TNF-α) by macrophages [80, 81]. Studies in rodents showed remarkable but selective efficacy against TNF-α-sensitive tumors, which improved in combination with other antitumor therapies [82]. Unfortunately, only a primed state induction of TNF-α was detected in human cells [83]. Phase I clinical studies revealed a limited capacity and the antitumor studies were not continued [79]. In contrast, the molecule exhibits anti-leishmanial [84] and anti-allergy activities [85].
3 Conclusions
Although LPS was long known as an immune stimulatory substance with potential adjuvant use, the large variety of unacceptable toxic effects drastically restricted its clinical use. However, the finding that MPL was safe and retained the desirable adjuvant properties of LPS opened new possibilities to treat pathogen diseases. In contrast to previous adjuvants, LPS-based adjuvants offer new benefits from their ability to enhance Th1-type responses and stimulate cytotoxic T lymphocytes. This activity is essential to confer protection against many pathogens and to develop prophylaxis therapies against other diseases as cancer or allergy. Indeed, this is supported by the efficacy of MPL in available vaccines whereas standard adjuvants failed to provide protection. Additionally, the high adjuvant capacity of LPS-based adjuvants has considerable and obvious benefits in mucosa protection, faster activation of protection, reduction of booster doses, functional immunization in elderly, or preparation of polyvalent vaccine formulations. Certain drawbacks were attributed to MPL, e.g., elevate production cost and possible activation/enhancement of TLR4-related autoimmune diseases. Synthetic lipid A analogues with similar biological activity like MPL have demonstrated considerable reduction of the production cost. In regard to the activation TLR4 autoimmune diseases, accumulated data till date from immunization in humans provides further evidence of safety. In summary, LPS-based adjuvants improve the current vaccination therapies and open possibilities to solve their existing challenges.
References
Rietschel, E.T., et al.: Bacterial endotoxins: chemical structure, biological activity and role in septicaemia. Scand. J. Infect. Dis. Suppl. 31, 8–21 (1982)
Guo, L., et al.: Lipid A acylation and bacterial resistance against vertebrate antimicrobial peptides. Cell 95, 189–198 (1998)
Gibbons, H.S., Lin, S., Cotter, R.J., Raetz, C.R.: Oxygen requirement for the biosynthesis of the S-2-hydroxymyristate moiety in Salmonella typhimurium lipid A. Function of LpxO, A new Fe2+/alpha-ketoglutarate-dependent dioxygenase homologue. J. Biol. Chem. 275, 32940–32949 (2000)
Reynolds, C.M., et al.: An outer membrane enzyme encoded by Salmonella typhimurium lpxR that removes the 3’-acyloxyacyl moiety of lipid A. J. Biol. Chem. 281, 21974–21987 (2006)
Trent, M.S., Pabich, W., Raetz, C.R., Miller, S.I.: A PhoP/PhoQ-induced Lipase (PagL) that catalyzes 3-O-deacylation of lipid A precursors in membranes of Salmonella typhimurium. J. Biol. Chem. 276, 9083–9092 (2001)
Kawai, T., Akira, S.: Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 34, 637–650 (2011)
Palsson-McDermott, E.M., O’Neill, L.A.: Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 113, 153–162 (2004)
Trinchieri, G., Sher, A.: Cooperation of Toll-like receptor signals in innate immune defence. Nat. Rev. Immunol. 7, 179–190 (2007)
Kawai, T., Akira, S.: TLR signaling. Cell Death Differ. 13, 816–825 (2006)
Miggin, S.M., O’Neill, L.A.: New insights into the regulation of TLR signaling. J. Leukoc. Biol. 80, 220–226 (2006)
Alexander, C., Rietschel, E.T.: Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 7, 167–202 (2001)
Morrison, D.C., Kline, L.F.: Activation of the classical and properdin pathways of complement by bacterial lipopolysaccharides (LPS). J. Immunol. 118, 362–368 (1977)
Cooper, N.R., Morrison, D.C.: Binding and activation of the first component of human complement by the lipid A region of lipopolysaccharides. J. Immunol. 120, 1862–1868 (1978)
Conti, P., et al.: Activation of human natural killer cells by lipopolysaccharide and generation of interleukin-1 alpha, beta, tumour necrosis factor and interleukin-6. Effect of IL-1 receptor antagonist. Immunology 73, 450–456 (1991)
Kobayashi, M., et al.: Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 170, 827–845 (1989)
Cinel, I., Dellinger, R.P.: Advances in pathogenesis and management of sepsis. Curr. Opin. Infect. Dis. 20, 345–352 (2007)
Annane, D., Bellissant, E., Cavaillon, J.M.: Septic shock. Lancet 365(63–78) (2005)
Vincent, J.L., et al.: Sepsis in European intensive care units: results of the SOAP study. Crit. Care Med. 34, 344–353 (2006)
Martin, G.S., Mannino, D.M., Eaton, S., Moss, M.: The epidemiology of sepsis in the United States from 1979 through 2000. N. Engl. J. Med. 348, 1546–1554 (2003)
Kotani, S., Takada, H.: Structural requirements of lipid A for endotoxicity and other biological activities – an overview. Adv. Exp. Med. Biol. 256, 13–43 (1990)
Nowotny, A.: Molecular aspects of endotoxic reactions. Bacteriol. Rev. 33, 72–98 (1969)
Miller, S.I., Ernst, R.K., Bader, M.W.: LPS, TLR4 and infectious disease diversity. Nat. Rev. Microbiol. 3, 36–46 (2005)
Golenbock, D.T., Hampton, R.Y., Qureshi, N., Takayama, K., Raetz, C.R.: Lipid A-like molecules that antagonize the effects of endotoxins on human monocytes. J. Biol. Chem. 266, 19490–19498 (1991)
Kotani, S., et al.: Synthetic lipid A with endotoxic and related biological activities comparable to those of a natural lipid A from an Escherichia coli re-mutant. Infect. Immun. 49, 225–237 (1985)
Saitoh, S., et al.: Lipid A antagonist, lipid IVa, is distinct from lipid A in interaction with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced TLR4 oligomerization. Int. Immunol. 16, 961–969 (2004)
David, S., Vermeer-de Bondt, P.E., van der Maas, N.A.: Reactogenicity of infant whole cell pertussis combination vaccine compared with acellular pertussis vaccines with or without simultaneous pneumococcal vaccine in the Netherlands. Vaccine 26, 5883–5887 (2008)
Gustafsson, L., Hallander, H.O., Olin, P., Reizenstein, E., Storsaeter, J.: A controlled trial of a two-component acellular, a five-component acellular, and a whole-cell pertussis vaccine. N. Engl. J. Med. 334, 349–355 (1996)
Brewer, J.M., et al.: Aluminium hydroxide adjuvant initiates strong antigen-specific Th2 responses in the absence of IL-4- or IL-13-mediated signaling. J. Immunol. 163, 6448–6454 (1999)
Dubensky Jr., T.W., Reed, S.G.: Adjuvants for cancer vaccines. Semin. Immunol. 22, 155–161 (2010)
Soghoian, D.Z., Streeck, H.: Cytolytic CD4(+) T cells in viral immunity. Expert Rev. Vaccines 9, 1453–1463 (2010)
Ziegler, A., et al.: EpCAM, a human tumor-associated antigen promotes Th2 development and tumor immune evasion. Blood 113, 3494–3502 (2009)
Akkoc, T., Akdis, M., Akdis, C.A.: Update in the mechanisms of allergen-specific immunotheraphy. Allergy Asthma Immunol. Res. 3, 11–20 (2011)
Chen, K., Cerutti, A.: Vaccination strategies to promote mucosal antibody responses. Immunity 33, 479–491 (2010)
Lawson, L.B., Norton, E.B., Clements, J.D.: Defending the mucosa: adjuvant and carrier formulations for mucosal immunity. Curr. Opin. Immunol. 23, 414–420 (2011)
O’Hagan, D.T.: MF59 is a safe and potent vaccine adjuvant that enhances protection against influenza virus infection. Expert Rev. Vaccines 6, 699–710 (2007)
Arenas, J.: The role of bacterial lipopolysaccharides as immune modulator in vaccine and drug development. Endocr. Metab. Immune Disord. Drug Targets 12(3), 221–235 (2012)
Baldridge, J.R., Crane, R.T.: Monophosphoryl lipid A (MPL) formulations for the next generation of vaccines. Methods 19, 103–107 (1999)
Evans, J.T., et al.: Enhancement of antigen-specific immunity via the TLR4 ligands MPL adjuvant and Ribi.529. Expert Rev. Vaccines 2, 219–229 (2003)
Ulrich, J.T., Myers, K.R.: Monophosphoryl lipid A as an adjuvant. Past experiences and new directions. Pharm. Biotechnol. 6, 495–524 (1995)
Vajdy, M., Singh, M.: The role of adjuvants in the development of mucosal vaccines. Expert Opin. Biol. Ther. 5, 953–965 (2005)
Drachenberg, K.J., Wheeler, A.W., Stuebner, P., Horak, F.: A well-tolerated grass pollen-specific allergy vaccine containing a novel adjuvant, monophosphoryl lipid A, reduces allergic symptoms after only four preseasonal injections. Allergy 56, 498–505 (2001)
Drachenberg, K.J., Heinzkill, M., Urban, E., Woroniecki, S.R.: Efficacy and tolerability of short-term specific immunotherapy with pollen allergoids adjuvanted by monophosphoryl lipid A (MPL) for children and adolescents. Allergol. Immunopathol. (Madr.) 31, 270–277 (2003)
Kundi, M.: New hepatitis B vaccine formulated with an improved adjuvant system. Expert Rev. Vaccines 6, 133–140 (2007)
Aide, P., et al.: Four year immunogenicity of the RTS, S/AS02(A) malaria vaccine in Mozambican children during a phase IIb trial. Vaccine 29, 6059–6067 (2011)
Von, E.K., et al.: The candidate tuberculosis vaccine Mtb72F/AS02A: Tolerability and immunogenicity in humans. Hum. Vaccin. 5, 475–482 (2009)
Polhemus, M.E., et al.: Evaluation of RTS, S/AS02A and RTS, S/AS01B in adults in a high malaria transmission area. PLoS One 4, e6465 (2009)
Brichard, V.G., Lejeune, D.: Cancer immunotherapy targeting tumour-specific antigens: towards a new therapy for minimal residual disease. Expert Opin. Biol. Ther. 8, 951–968 (2008)
Garcon, N., Chomez, P., Van, M.M.: GlaxoSmithKline Adjuvant Systems in vaccines: concepts, achievements and perspectives. Expert Rev. Vaccines 6, 723–739 (2007)
Fries, L.F., et al.: Liposomal malaria vaccine in humans: a safe and potent adjuvant strategy. Proc. Natl. Acad. Sci. U. S. A. 89, 358–362 (1992)
Arenas, J., et al.: Coincorporation of LpxL1 and PagL mutant lipopolysaccharides into liposomes with Neisseria meningitidis opacity protein: influence on endotoxic and adjuvant activity. Clin. Vaccine Immunol. 17, 487–495 (2010)
Vernacchio, L., et al.: Effect of monophosphoryl lipid A (MPL) on T-helper cells when administered as an adjuvant with pneumocococcal-CRM197 conjugate vaccine in healthy toddlers. Vaccine 20, 3658–3667 (2002)
Olson, K., et al.: Liposomal gD ectodomain (gD1-306) vaccine protects against HSV2 genital or rectal infection of female and male mice. Vaccine 28, 548–560 (2009)
Hall, M.A., et al.: Intranasal immunization with multivalent group A streptococcal vaccines protects mice against intranasal challenge infections. Infect. Immun. 72, 2507–2512 (2004)
Tana, W.S., Isogai, E., Oguma, K.: Induction of intestinal IgA and IgG antibodies preventing adhesion of verotoxin-producing Escherichia coli to Caco-2 cells by oral immunization with liposomes. Lett. Appl. Microbiol. 36, 135–139 (2003)
Baldridge, J.R., Yorgensen, Y., Ward, J.R., Ulrich, J.T.: Monophosphoryl lipid A enhances mucosal and systemic immunity to vaccine antigens following intranasal administration. Vaccine 18, 2416–2425 (2000)
Kidon, M.I., Shechter, E., Toubi, E.: Vaccination against human papilloma virus and cervical cancer. Harefuah 150, 33–36 (2011). 68
Labadie, J.: Postlicensure safety evaluation of human papilloma virus vaccines. Int. J. Risk Saf. Med. 23, 103–112 (2011)
Morello, C.S., Levinson, M.S., Kraynyak, K.A., Spector, D.H.: Immunization with herpes simplex virus 2 (HSV-2) genes plus inactivated HSV-2 is highly protective against acute and recurrent HSV-2 disease. J. Virol. 85, 3461–3472 (2011)
Llanos-Cuentas, A., et al.: A clinical trial to evaluate the safety and immunogenicity of the LEISH-F1 + MPL-SE vaccine when used in combination with sodium stibogluconate for the treatment of mucosal leishmaniasis. Vaccine 28, 7427–7435 (2010)
Zhu, D., Barniak, V., Zhang, Y., Green, B., Zlotnick, G.: Intranasal immunization of mice with recombinant lipidated P2086 protein reduces nasal colonization of group B Neisseria meningitidis. Vaccine 24, 5420–5425 (2006)
Green, B.A., et al.: PppA, a surface-exposed protein of Streptococcus pneumoniae, elicits cross-reactive antibodies that reduce colonization in a murine intranasal immunization and challenge model. Infect. Immun. 73, 981–989 (2005)
Egan, M.A., et al.: A comparative evaluation of nasal and parenteral vaccine adjuvants to elicit systemic and mucosal HIV-1 peptide-specific humoral immune responses in cynomolgus macaques. Vaccine 22, 3774–3788 (2004)
Mason, K.W., et al.: Reduction of nasal colonization of nontypeable Haemophilus influenzae following intranasal immunization with rLP4/rLP6/UspA2 proteins combined with aqueous formulation of RC529. Vaccine 22, 3449–3456 (2004)
Baldridge, J.R., et al.: Immunostimulatory activity of aminoalkyl glucosaminide 4-phosphates (AGPs): induction of protective innate immune responses by RC-524 and RC-529. J. Endotoxin Res. 8, 453–458 (2002)
Ishizaka, S.T., Hawkins, L.D.: E6020: a synthetic Toll-like receptor 4 agonist as a vaccine adjuvant. Expert Rev. Vaccines 6, 773–784 (2007)
Morefield, G.L., Hawkins, L.D., Ishizaka, S.T., Kissner, T.L., Ulrich, R.G.: Synthetic Toll-like receptor 4 agonist enhances vaccine efficacy in an experimental model of toxic shock syndrome. Clin. Vaccine Immunol. 14, 1499–1504 (2007)
Przetak, M., et al.: Novel synthetic LPS receptor agonists boost systemic and mucosal antibody responses in mice. Vaccine 21, 961–970 (2003)
Baudner, B.C., et al.: MF59 emulsion is an effective delivery system for a synthetic TLR4 agonist (E6020). Pharm. Res. 26, 1477–1485 (2009)
Wang, S., et al.: Effective antibody therapy induces host-protective antitumor immunity that is augmented by TLR4 agonist treatment. Cancer Immunol. Immunother. 61, 49–61 (2012)
Baldwin, S.L., et al.: Intradermal immunization improves protective efficacy of a novel TB vaccine candidate. Vaccine 27, 3063–3071 (2009)
Bertholet, S., et al.: Optimized subunit vaccine protects against experimental leishmaniasis. Vaccine 27, 7036–7045 (2009)
Coler, R.N., et al.: A synthetic adjuvant to enhance and expand immune responses to influenza vaccines. PLoS One 5, e13677 (2010)
Xiao, L., et al.: A TLR4 agonist synergizes with dendritic cell-directed lentiviral vectors for inducing antigen-specific immune responses. Vaccine 30, 2570–2581 (2012)
Pantel, A., et al.: A new synthetic TLR4 agonist, GLA, allows dendritic cells targeted with antigen to elicit Th1 T-cell immunity in vivo. Eur. J. Immunol. 42, 101–109 (2012)
Cong, H., et al.: Toxoplasma gondii HLA-B*0702-restricted GRA7(20-28) peptide with adjuvants and a universal helper T cell epitope elicits CD8(+) T cells producing interferon-gamma and reduces parasite burden in HLA-B*0702 mice. Hum. Immunol. 73, 1–10 (2012)
Baldwin, S.L., et al.: The importance of adjuvant formulation in the development of a tuberculosis vaccine. J. Immunol. 188, 2189–2197 (2012)
Windish, H.P., et al.: Protection of mice from Mycobacterium tuberculosis by ID87/GLA-SE, a novel tuberculosis subunit vaccine candidate. Vaccine 29, 7842–7848 (2011)
Ueda, H., Yamazaki, M.: Induction of tumor necrosis factor in a murine tumor by systemic administration of a novel synthetic lipid A analogue, ONO-4007. J. Immunother. 20, 65–69 (1997)
Matsumoto, N., Aze, Y., Akimoto, A., Fujita, T.: Restoration of immune responses in tumor-bearing mice by ONO-4007, an antitumor lipid A derivative. Immunopharmacology 36, 69–78 (1997)
Kuramitsu, Y., et al.: A new synthetic lipid A analog, ONO-4007, stimulates the production of tumor necrosis factor-alpha in tumor tissues, resulting in the rejection of transplanted rat hepatoma cells. Anticancer Drugs 8, 500–508 (1997)
Matsumoto, N., Oida, H., Aze, Y., Akimoto, A., Fujita, T.: Intratumoral tumor necrosis factor induction and tumor growth suppression by ONO-4007, a low-toxicity lipid A analog. Anticancer Res. 18, 4283–4289 (1998)
Inagawa, H., et al.: Mechanisms by which chemotherapeutic agents augment the antitumor effects of tumor necrosis factor: involvement of the pattern shift of cytokines from Th2 to Th1 in tumor lesions. Anticancer Res. 18, 3957–3964 (1998)
Matsumoto, N., Aze, Y., Akimoto, A., Fujita, T.: ONO-4007, an antitumor lipid A analog, induces tumor necrosis factor-alpha production by human monocytes only under primed state: different effects of ONO-4007 and lipopolysaccharide on cytokine production. J. Pharmacol. Exp. Ther. 284, 189–195 (1998)
Khan, M.A., et al.: Inhibition of intracellular proliferation of Leishmania parasites in vitro and suppression of skin lesion development in BALB/c mice by a novel lipid A analog (ONO-4007). Am. J. Trop. Med. Hyg. 67, 184–190 (2002)
Iio, J., et al.: Lipid A analogue, ONO-4007, inhibits IgE response and antigen-induced eosinophilic recruitment into airways in BALB/c mice. Int. Arch. Allergy Immunol. 127, 217–225 (2002)
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Arenas, J. (2014). Bacterial Lipopolysaccharide as Adjuvants. In: Giese, M. (eds) Molecular Vaccines. Springer, Cham. https://doi.org/10.1007/978-3-319-00978-0_7
Download citation
DOI: https://doi.org/10.1007/978-3-319-00978-0_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-00977-3
Online ISBN: 978-3-319-00978-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)