
Abstract
The radially symmetric body of starfish has major implications on their nervous system including eyes and vision. All the up to 50 arms are structurally identical, and most examined species have a small compound eye basally on the terminal tube foot of each arm. The 20–300 ommatidia of the compound eyes are lens-less but hold approximately 100 photoreceptors with outer segments made of a combination of microvilli and a modified cilium. The eyes support image forming vision but of low spatial resolution and extremely low temporal resolution with flicker fusion frequencies ≤1 Hz. Starfish are color-blind, and vision seems to be based on a single rhabdomeric opsin although many other types of opsins are expressed in their eyes. Starfish also possess extraocular photoreceptors, but little is known about their identity and function. Not many visually guided behaviors are known from starfish so far, but habitat recognition is well documented in a couple of tropical species. More behavioral data are urgently needed, but interestingly, recent data suggest that at least in some situations vision is integrated with olfaction and rheotaxis forming a sensory hierarchy, where olfaction is dominating. Such processing and integration putatively take place in the central nervous system. The eyes are direct extensions of the radial nerve, which constitute the major part of the CNS of starfish and other echinoderms. In general, the echinoderm CNS is enigmatic and the functionality is at best speculative. Here we present new data showing differentiations of the radial nerve along the length of the arms and differences in radial nerve structure between eye-possessing and eyeless species.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Compound eyes
- Ommatidia
- Low resolution vision
- Radial nerve
- Sea star
- Radial symmetry
- Echinoderm
- Temporal resolution
- Habitat recognition
4.1 Introduction
Echinoderms are remarkable in many ways. Despite being nested within Deuterostomia, most of the adult animals display penta-radial symmetry, which comes about through a truly astonishing metamorphosis of their larvae rivaling in complexity the metamorphosis of holometabolic insects (Lascalli 2000). This has major impact on the organization of the organs in the body, including the nervous system. Echinoderms are in general large animals, but many are semi-sessile with a rather simple body structure where, for example, the excretion and the blood-vascular systems are strongly reduced (Chia and Koss 1994). Most species also lack strong abilities to osmoregulate, and they are thus often restricted to areas of high salinity oceanic water. In these areas, though, they are often dominating the benthos, and echinoderms are found in all seas from the intertidal zone down to the deepest parts of the abyssal plains below 8000 m.
Asteroidea, starfish, is one of the most species-rich classes of echinoderms counting close to 2000 extent species (www.marinespecies.org/asteroidea). Most are specialized predators and display the most active behavior of all echinoderms even though the measured maximum walking speed is only approx. 80 cm/min (Mueller et al. 2011). In coastal areas, many species like Asterias rubens feed on the abundance of mussels often found in the area, whereas deep-sea starfish often prey on gorgonian corals or tiny planktonic or benthic crustaceans. The latter are prey for members of the specialized brisingid family, which are sit-and-wait predators putatively catching their crustacean prey in similar ways as cnidarian polyps using their 1000s of pedicellaria, instead of cnidocytes, to capture their planktonic prey (Zhang et al. 2019). Starfish are not exclusively predators, though; some are scavengers, whereas others are suspension feeders and a large number of species are detritivorous like the iconic blue starfish Linckia laevigata (Mueller et al. 2011). There are no doubts that starfish play major ecological roles in most marine habitats sometimes even at the level of single species. This is the case for the corallivorous species complex Acanthaster planci. Found all over the tropics, they specialize in feeding on scleractinian corals (Fig. 4.1). They occasionally occur in major outbreaks causing great damage to the coral reefs they inhabit and devour, and these outbreaks have increased in frequency over the last 50 years. It is estimated that approx. 20% of the coral decline globally is a result of these outbreaks (De’ath and Moran 1998; Fabricius et al. 2000; Moran 1986). For this reason, they have been intensively studied and become a model species for understanding starfish ecology and reproductive biology and lately also starfish neurobiology and sensory ecology including vision (Hall et al. 2017; Lowe et al. 2018; Lucas 2013; Motti et al. 2018; Petie et al. 2016b).

The model starfish Acanthaster planci. (a) A. planci is a many-armed species with up to 24 arms each carrying a compound eye at the tip. The specimen here is an 8 cm juvenile seen from its aboral side clearly demonstrating the characteristic radial symmetry of starfish. The many spines have given it the trivial name crown-of-thorns starfish. (b) As late juveniles and adults, A. planci feeds almost exclusively on scleractinian corals, and here it is eating a staghorn coral. The white areas of the coral are areas already eaten by the starfish. It is estimated that 20% of all coral decline on the Great Barrier Reef in Australia is caused by outbreaks of A. planci
Starfish are clearly radially symmetric with a central disk from which a number of evenly distributed arms project to the sides (Fig. 4.1). Most species of starfish have 5 arms, but in some species, like the Antarctic Labidiaster annulatus, there can be as many as 50 arms. There is no known differentiation between the arms in a specimen, the arms are identical containing the same elements, and several species can reproduce asexually by autotomizing an arm, which will afterward regenerate the rest of the body (Clements et al. 2019). Mainly due to this unorthodox organization of the body, starfish and other echinoderms are often considered to lack a central nervous system (CNS) as a central brain is not present (Clark et al. 2019). However, this is a far too simplistic view on the echinoderm nervous system. Detailed morphological studies have clearly demonstrated that they do possess a CNS but without a central brain. It consists of a ring nerve encircling the mouth opening and branching off a number of radial nerve cords, in starfish one projecting into each of the arms (Mashanov et al. 2006, 2009). Running in parallel with the CNS are the major parts of the water vascular system, unique to echinoderms, where the functional parts are the tube feet running in rows on the outside of the animals (Ullrich-Lüter et al. 2011). The tube feet serve a multitude of functions including locomotion, prey capture, excretion, respiration, and sensing. Almost all sensing in tube feet takes place in their distal end, normally forming an attachment disc, and here a high number of putative mechano- and chemoreceptors are found (Moore and Thorndyke 1993; Ullrich-Lüter et al. 2011). Accordingly, several behavioral studies have indicated that starfish, for a large part, are guided by rheotaxis and olfaction (Castilla and Crisp 1973; Valentincic 1975). Uniquely within echinoderms, most starfish possess an additional sensory system on the terminal tube foot, which is the first tube foot to develop during the metamorphosis and the only unpaired tube foot. This tube foot carries a compound eye basally on the oral side (Fig. 4.2) (Jourdain 1865). The eye constitutes the distal-most part of the radial nerve cord and is thus embedded in the CNS similar to the vertebrate eye, but since there is an eye on each arm tip, starfish eyes are also clear examples of a distributed visual system with between 5 and 50 eyes evenly distributed along the periphery of the body (Garm 2017).

Diversity of starfish eyes. Most studied starfish have a compound eye on the terminal tube foot (TTF) of each arm. Here the TTF is seen for Marthasterias glacialis (a), Pentaceraster mammillatus (b), Asterina sp. (c), Acanthaster planci (d), and Culcita novaeguineae (e). The red screening pigment of the ommatidia is clearly seen on all species (arrow in c). Note the very large eyes on P. mammillatus and C. novaeguineae. Following the TTF are a number of smaller tube feet lacking the disk (asterisk), and they are believed to be the center of olfaction in starfish often called sensory tube feet
4.2 The Starfish Eyes
The starfish compound eye is sometimes called the optic cushion referring to the cushion shape with the ommatidia evenly distributed along the surface. Depending on species, there may be from fewer than 20 to more than 300 ommatidia with each of them clearly distinguishable as a ring of bright red screening pigment (Fig. 4.3). The chemical nature of this screening pigment is still unknown. It has been proven behaviorally that the eyes form true images and that the animal uses this image information (see later for details) (Petie et al. 2016a). Each ommatidium contains a large number of photoreceptors, but since the outer segments of the receptors are intermingled, it suggests that they sample light from the same area in space, thus collectively forming one separate part of the image (like a pixel in a digital image). In most cases, the ommatidia are round with a round pupil, but in some deep-sea species, the ommatidia are strongly elongated along the oral-aboral axis putatively gaining sensitivity without losing resolution along the horizon (Fig. 4.3) (Birk et al. 2018). So far, Hippasteria phrygiana is the examined species with the most ommatidia per eye (up to 320) (Birk et al. 2018), but preliminary data suggests that other species like Culcita novaeguineae and Pentaceraster mammillatus may have even more (Fig. 4.2).

Morphology of the starfish eye. (a, d, g) Eyes of Acanthaster planci, Pteraster pulvillus, and Novodinia americana, respectively. Note the round ommatidia in A. planci and N. americana and the vertically elongated ommatidia in the semi deep-sea species P. pulvillus. (b, e, h) Longitudinal sections of an ommatidium from each species. There are no lenses, and the inner part is filled with the outer segments (OS) of the photoreceptors surrounded by screening pigment (SP) mainly from pigment cells. (c, f, i) TEM micrographs of framed areas in (b, e, h) respectively. Note the rather loose arrangement of the photosensitive membranes in A. planci (c) and the dense packing for the deep-sea species N. americana (i). Scale bar in (b) also fits (e) and (h); scale bar in (c) also fits (f) and (i). (Modified after Birk et al., 2018)
The structure of the ommatidia is similar across all examined species. They are built by the same two cell types: pigmented photoreceptors and pigment cells. There seem to be no other cells involved in the eyes even though they are covered by an epithelium, which has been suggested to have an optical function (Penn and Alexander 1980). In most examined species, the epithelium is a monolayer of flat electron lucent cells, which rules out optical functions (Birk et al. 2018; Garm and Nilsson 2014). A fully developed ommatidium from an eye of L. laevigata or A. planci has approximately 100 pigment cells and a little fewer photoreceptors. The pigment cells are about 10 μm wide and 15–20 μm long with the apical part filled with pigment granules. The photoreceptors are slimmer but longer with their photosensitive outer segment alone being 10–30 μm long (Garm and Nilsson 2014). Interestingly, the outer segments of starfish eyes are made of a combination of microvilli and a modified cilium (Petie et al. 2016b), which might result in starfish photoreceptors having a different transduction mechanism than the normal ciliary and rhabdomeric photoreceptors. This is supported by structurally similar receptors in chiton larvae expressing xenopsins, a recently discovered family of opsins (Vöcking et al. 2017). In shallow water species, the photosensitive membranes are loosely packed compared to the eyes from deep-sea starfish, which seem to have increased their sensitivity by packing the membranes tighter (Fig. 4.3).
The starfish eye grows with the size of the animal. Contrary to many other animals where the eyes grow allometrically relative to the rest of the body, there is a close to linear growth in eyes of A. planci. As a few-month-old juveniles (3–5 cm in diameter), they have only 20–30 ommatidia in each eye, but as adults (40–50 cm in diameter), they have between 280 and 300 ommatidia in each eye (Korsvig-Nielsen et al. 2019; Petie et al. 2016b). The new ommatidia are added both in the periphery of the eye and in between the existing ommatidia, and this results in more acute vision as the animals grow, while the visual field of the eyes stays the same. Interestingly, the size of each ommatidium is the same in juveniles and adults indicating that they compromise spatial resolution as juveniles but not sensitivity (Korsvig-Nielsen et al. 2019). The terminal tube foot including the compound eye is formed during the larval metamorphosis, but here there are as few as four to five ommatidia. The visual capacity and functional significance of these tiny eyes are yet to be tested.
4.2.1 Low Pass Filtering in Starfish Eyes
As mentioned above, the highest number of ommatidia found in a starfish eye is just above 300, and this was for the North Atlantic species H. phygiana (Birk et al. 2018). Since the number of ommatidia putatively equals the number of resolved areas (pixels) in the formed image, it is obvious that the obtained spatial information is low. The maximum spatial resolution has been estimated in a few species through measurements of the interommatidial angles, and they are in the range of 7–17° (Birk et al. 2018; Garm and Nilsson 2014), which is comparable to cubomedusan and small insect eyes (Nilsson et al. 2005). Interestingly, the deep-sea species, Novodinia americana, is one of those with the highest resolution, which is putatively because they need to resolve bioluminescent patterns (Birk et al. 2018). Still, the relative low spatial resolution in all examined starfish results in low pass filtering – low spatial frequencies (large objects) are seen and high spatial frequencies (small objects) are filtered away. This again means that small and distant objects will not be seen whereas large relatively nearby objects will. In the only two species, L. laevigata and A. planci, where there is behavioral evidence for which part of the visual environment they see, it turns out to be the major structure in their habitat, the coral boulders (see below for details on visually guided behaviors) (Petie et al. 2016b; Sigl et al. 2016).
Low pass filtering is a central theme of starfish vision perhaps best seen in the temporal resolution. Again the initial data came from the model species A. planci, and it was found that this species have the lowest temporal resolution of all animal eyes examined so far (Petie et al. 2016b). The temporal resolution is measured as the flicker fusion frequency (fff) – the frequency where a sinusoidal change in the light intensity is no longer registered by the photoreceptors but seen as a constant stimulation with the average intensity. Depending on the absolute intensity, this typically varies between 20 and 55 Hz for the human eye, whereas the fff of A. planci is as low as 0.5–0.6 Hz (Fig. 4.4a). This means that changes happening faster than every other second are not seen! Such extreme low pass temporal filtering results in even slow moving objects causing severe image blur, and only stationary large objects – like the coral boulders – are seen by the animals. Importantly, there is also a match with the self-motion of the starfish. With such a low temporal resolution, any fast movement of the eye bearing structures will also result in problems with image blur, but with maximum speeds of 60 cm/min, this is no problem for A. planci (Mueller et al. 2011). Importantly, the fff of a given eye is influenced not only by the intensity of light changes but also by the temperature of the photoreceptors. The warmer the eyes, the faster they are (Fritsches et al. 2005), and since A. planci is a tropical animal, this could mean that cold water starfish have even lower temporal resolution. This is not the case, though, since our previously unpublished data generated with the same protocol as for A. planci shows that A. rubens kept at 10 °C have a fff of approx. 1 Hz (Fig. 4.4c). This strongly indicates that it is not physiological constrains that set the fff in starfish photoreceptors but rather selective pressure on having low pass temporal filtering. The putative explanation is that in combination with the low pass spatial filter, it works as match filtering. Such filtering ensures that only large stationary objects are seen, whereas small moving objects, which are normally not important for the starfish, are removed from the visual input minimizing the need for processing power in the CNS.

Temporal resolution and spectral sensitivity. (a) Flicker fusion frequency (fff) curve for Acanthaster planci. There is a steep decline in the power of the response with increasing frequency, and the fff at 0.1 (broken line) is reached at approx. 0.6 Hz. (b) Spectral sensitivity curve for A. planci. The best fit using the least squares method is for an opsin peaking at 472 nm (red curve). Note that the curve is narrower than the opsin absorption curve indicating spectral filtering is taking place. (c) Flicker fusion frequency (fff) curve for Asterias rubens. Here there is also a steep decline in the power of the response with increasing frequency, and the fff at 0.1 (broken line) is reached at approx. 0.9 Hz. (d) Spectral sensitivity curve for A. rubens. The best fit is for an opsin peaking at 484 nm (red curve). A and B are modified after Petie et al., 2016b. Error bars in (c) and (d) indicate SD, n = 8 in (c) and 9 in (d)
4.2.2 Opsins and Spectral Sensitivity
All available data suggest that vision in starfish is based on opsins as the visual pigment – similar to all other examined animal eyes (Land and Nilsson 2012). Molecular examinations of Asterias rubens and Patiria miniata identified six and ten different opsins, respectively, but no data on expression patterns are available for these species (D’Aniello et al. 2015). Tissue specific transcriptomic data also found ten different opsins belonging to seven opsin families in A. planci with many of them highly expressed in the eye bearing terminal tube foot. Still, the only rhabdomeric opsin found in the sequences was by far the highest expressed opsin in the area of the eyes, which suggests that it is the visual pigment of the photoreceptors (Lowe et al. 2018). If true, this would be the first deuterostome eyes with photoreceptors utilizing rhabdomeric opsins. A chaopsin was also highly expressed in the eyes, but the functions of this enigmatic opsin family remains unknown. As mentioned earlier, a recently discovered opsin family, xenopsins, has been found in chiton photoreceptors structurally similar to starfish photoreceptors, but xenopsins were not found in A. planci and have so far not been found outside Prostomia (Döring et al. 2020).
Electrophysiological work performing electroretinograms (ERGs) supports the presence of a single opsin in the photoreceptors (Garm and Nilsson 2014; Petie et al. 2016b). In A. planci, L. laevigata, and A. rubens, the spectral sensitivity has a single peak in the blue part of the visual spectrum with λ-max = 472nm, 452nm, and 484nm, respectively. In the two first species, the obtained spectral sensitivity curves are narrower than the modeled absorption curve of an opsin (Garm and Nilsson 2014; Govardovskii et al. 2000; Petie et al. 2016b) (Fig. 4.4b). The suggested explanation here is that there is an external filtering happening putatively removing the damaging UV light present at high intensities in the shallow water tropical habitat of these two species (Garm and Nilsson 2014; Petie et al. 2016b). The spectral sensitivity peak in the deep blue part of the spectrum for these two species is matching the most abundant wavelengths in the clear ocean water they live in (McFarland and Munz 1975), thereby optimizing the contrast to objects in the water. The spectral curve of A. rubens has a much better match with a modeled opsin curve indicating that spectral filtering does not take place in this species (Fig, 4d). The peak sensitivity is also shifted a bit to about 485 nm, and both these differences match A. rubens living in temperate waters with a higher organic content, which removes most of the UV light and green-shifts the color of the water (Lythgoe 1979).
Behavioral experiments with blinded animals (eye-ablated) and specimens of eyeless species show that they still respond to light stimuli, which must then be controlled by yet unidentified extraocular photoreceptors (see later this chapter). This is well in line with data from members of other echinoderm classes such as sea urchins and brittle stars (Lesser et al. 2011; Sumner-Rooney et al. 2020; Ullrich-Lüter et al. 2013) (see also Chapter 3 this volume). Highly interesting, in both sea urchins and brittle stars there is recent evidence that these extraocular photoreceptors provide the animals with spatial vision although of very low resolution (Kirwan et al. 2018; Sumner-Rooney et al. 2020). It is currently unknown which of the opsins are involved in starfish extraocular photoreceptors, but in both sea urchins and brittle stars, it is a rhabdomeric opsin. What all the other opsins are doing in starfish is still unknown. There are no experimental or expression data to resolve this, but from their structure and through comparison with other systems, it has been suggested that some might be photoisomerases while others serve different physiological functions (Lowe et al. 2018).
4.3 Behavioral Repertoire of Starfish
Even though most echinoderms appear to be semi-sessile at first glance, they turn out to have a rather sophisticated behavioral repertoire, which is especially true for starfish. For over 100 years, starfish behavior has been examined with a focus on their foraging (Fenchel 1965; Kalmus 1929; Scheibling 1981) and reproduction (Boivin et al. 1986; Hamel and Mercier 1994). One of the functionally significant results from these studies is that there is not a leading arm per say – all arms take turn leading the animal pinpointing the uniqueness of their radial symmetric organization (Pearse et al. 1987). Most starfish are predators, many with a preference for bivalve prey, and their foraging has been shown to be at least partly olfactory guided but often in combination with negative rheotaxis (walking against the current) (Dale 1997). When it comes to reproduction, they are typically broadcast spawners, but little is known about the sensory cues behind the gamete release (Hamel and Mercier 1994). As all other animals, starfish are most likely multimodal in their behavioral control, and so far, chemoreception, mechanoreception, and photoreception have been documented (see [20], for review). The main sensory organs are the tube feet, and especially the young distal-most tube feet on each arm seem to be at play here. They lack the attachment disc and have no apparent mechanical function but are instead stretched out in front and above the arm (Sloan 1980). As mentioned earlier, this is also where the compound eyes are found: at the base of the unpaired terminal tube foot. Even though many behavioral studies have shown that starfish are olfactory guided, it is highly likely that control of most (if not all) behaviors involves multiple sensory input. Here we present some of the first evidence of multimodal behavior and reveal the included sensory hierarchy (see Sect. 4.5 on “Multimodal Control of Behavior” below).
4.4 Light Guided Behaviors
4.4.1 Shadow Response and Extraocular Photoreception
Extraocular photoreception seems to be common in echinoderms and has been studied in detail in the sea urchins Strongylocentrotus purpuratus and Diadema africanum (Kirwan et al. 2018; Ullrich-Lüter et al. 2011) and in brittle stars (see Chapter 3 this volume). Opsin expressing cells were found several places in S. purpuratus including at the base of the tube feet where they were suggested to support directional vision using the crust for directional screening (Ullrich-Lüter et al. 2011). Behavioral evidence from D. africanum supports such a system in sea urchins including proper image formation even though the spatial resolution is one of the lowest ever measured (30–70°) (Kirwan et al. 2018). Extraocular photoreception has also been suggested from starfish including from molecular data (Lowe et al. 2018), but little is known about the functional significance it may have in these animals.
In a previously unpublished behavioral study, we examined the shadow response of the sensory tube feet in adult specimens of four species of North Atlantic starfish: three with prominent eyes, L. laevigata (n = 3), Marthasterias glacialis (n = 5), and Crossaster papposus (n = 4), and the burrowing eyeless Astropecten irregularis (n = 2). We had previously observed that some starfish would retract the distal-most sensory tube feet when subjected to a passing shadow putatively as a predator avoidance response, and we tested the sensory basis behind this behavior. In room light, the arm tip including the sensory tube feet of a single arm was initially illuminated with a handheld torch for 1 min after which light was turned off. For L. laevigata, M. glacialis, and C. papposus, we tested both intact and eye-ablated specimens (number of test animals is the same as listed above in both cases). When illuminated, both the intact and eye-ablated animals of all species had the distal-most tube feet extended; typically moving them slowly from side to side (Fig. 4.5). The tube feet of L. laevigata were notably shorter/less extended than those of the other species (compare Fig. 4.5a, e, and i), which correlates with its coral habitat containing several fish species known to attack starfish including Chaetodon sp. (Cowan et al. 2017). The intact and eye-ablated L. laevigata and the eyeless A. irregularis all reacted in a similar way to the light-off stimulus mimicking a shadow passing the arm. With a latency of 0.7 +/−0.2 s for L. laevigata and 1.9 +/−0.6 s for A. irregularis, they rapidly withdraw all of the distal-most tube feet (Fig. 4.5). Interestingly, A. irregularis did not retract the terminal tube foot (Fig. 4.5j), which is of unknown function. This response to a pure light stimulation proved that it is governed by photoreceptors and since both the eye-ablated L. laevigata and the eyeless A. irregularis responded extraocular photoreceptors are involved. The whereabouts of these photoreceptors are still unknown, but a probable location is in the tube feet similar to what has been found in sea urchins (Ullrich-Lüter et al. 2011). Neither the intact nor the eye-ablated specimens of the last two species, C. papposus and M. glacialis, showed any response to the light-off stimulus (Fig. 4.5e–h, M. glacialis not shown), stressing the diversified response pattern. Despite the species differences, our results showed that similar to other echinoderms, some species of starfish do not rely on photoreception in their eyes alone but also utilize extraocular photoreceptors in their light guided behaviors.

Light-off response in the distal-most tube feet. (a–d) Both intact and eye-ablated Linckia laevigata respond to a shadow mimic (light-off) by retracting the distal-most black tube feet (arrows) showing that this behavior is at least partly guided by extraocular photoreceptors. (e–h) Neither intact nor eye-ablated Crossaster papposus responded to light-off and kept the distal-most tube feet extended at all times. (i–j) The eyeless Astropecten irregularis also respond to light-off by retracting the distal-most tube feet (arrows) adding further evidence to the presence of extraocular photoreceptors. Note that the unpaired terminal tube foot is not retracted (asterisk)
4.4.2 Visually Guided Habitat Detection: Proof of Image Forming Eyes
Some of the first experiments testing light guided behaviors in starfish were done with Asterias amurensis a little over 50 years ago, and this species was found to be negative phototactic (Yoshida and Ohtsuki 1968). New data support such behavior and that at least some starfish use vision for negative phototaxis and move toward dark objects in their habitat (Garm and Nilsson 2014; Korsvig-Nielsen et al. 2019; Petie et al. 2016b). A limited number of species have been tested, and most knowledge about the visually guided behavior again comes from A. planci, but data from L. laevigata are also available (Garm and Nilsson 2014). The visually guided behavior of these two species has been studied in situ at tropical coral reefs, as well as in behavioral arenas where the visual environment could be controlled in detail and where no directional olfactory or mechanosensory stimuli were present (Garm and Nilsson 2014; Petie et al. 2016a, 2016b; Sigl et al. 2016). The results from these combined experiments clearly show that both species are attracted to dark structures, which in their natural habitat are large coral boulders. Interestingly, the results from the behavioral arena showed that vision alone is sufficient to accomplish this behavior. The results from the natural habitat are somewhat divergent. When tested close to the reef in weak non-laminar currents, eye-ablated A. planci and L. laevigata with assumed intact chemo- and mechanoreception walked randomly, whereas sham operated animals walked toward the coral reef, indicating that vision is required to accomplish the behavior (Fig. 4.6). Another study did indicate, though, that under stronger semi-laminar current conditions, blinded animals can navigate toward the reef possibly using rheotaxis in combination with olfaction (Sigl et al. 2016). Our previously unpublished data from the North Atlantic species M. glacialis support the inclusion of rheotactic information, since they were only attracted to a 37° large dark visual stimulus in the presence of a semi-laminar current (see Sect. 4.5 on “Multimodal control of Behavior” for details).

Visually guided navigation in Acanthaster planci. (a) The visual scene toward and away from the coral habitat, when first spectrally filtered (blue opsin) and then spatially filtered (8 degrees resolution) to match the properties of vision in A. planci. Note that the coral reef is still visible as a dark area rising from the ocean floor. (b, c) Results from in situ behavioral experiments with A. planci showing that sham operated animals (vision intact, but a couple of locomotory tube feet removed) are able to detect the habitat whereas blinded animals (terminal tube foot with eye removed but other senses intact) walk randomly. There is 1 min between the dots; the white dots indicate the end positions. (Modified after Garm, 2017)
The results from the behavioral arena are well in line with the low spatial resolution in A. planci determined from eye morphology, since they were not attracted to dark visual stimuli until they took up a visual angle of at least 14° (Fig. 4.7a) (Petie et al. 2016a). The results also supported the lower resolution in juveniles as they were not attracted until the stimuli had a visual angle of 27° or more (Korsvig-Nielsen et al. 2019). Being attracted to dark objects/areas can be accomplished without proper image formation, though, using simple directional intensity measurements. Which mechanism lies behind the negative phototaxis has been tested through behavioral experiments for A. planci (Petie et al. 2016b). In the behavioral arena, the starfish were presented with a black and white square on a mid-intensity gray background, which means that just measuring the light intensity from a given area will not reveal the stimulus since it has the same average intensity as the gray background. Still, the adult A. planci showed clear directional walking and attraction when the initial angular height of the square was 32° or above (Fig. 4.7b). Importantly, they did not contact the center of the square but the black half of it (Petie et al. 2016a). These were the first direct behavioral proof of image forming eyes in starfish earlier suggested from the eye morphology. In contrast, juvenile A. planci were not able to discriminate the black and white stimulus from the gray background, even though they showed a strong tendency to be either attracted or repelled (Korsvig-Nielsen et al. 2019). This emphasizes that starfish vision changes (improves) with age and/or size of the animal, a phenomenon seen several places in the animal kingdom (Land and Nilsson 2012).

Negative phototaxis is Acanthaster planci. (a) When presented with a series of black circles on the wall of the behavioral arena, A. planci ignores these visual stimuli when smaller than 14° in visual angle. If larger than 14°, A. planci displays negative phototaxis and is attracted to the circles. (b) When presented with visual stimuli detectable only using spatial resolution (black and white square on average intensity gray background), A. planci again displays negative phototaxis but not until the vertical part of the black stimulus is 32° in visual angle or larger. Note that the animals contact the stimuli on the left side (at approx. 340°), which is the black part. The center of the stimulus is at 0°, and the icons to the left indicate visual stimuli and background intensity. Numbers above circles indicate the angular size of the stimulus. Black dots on the circles = individual mean headings, central arrows = the mean vector, broken lines = 95% confidence interval of the mean vector, and P = result of Raleigh test. (Modified after Petie et al., 2016a)
4.4.3 Eye Movements and Active Vision
An important part of the visual ecology of many animals is the ability to actively control the visual input through a variety of eye movements (Land 2019). Four functional types of eye movements exist: setting the gaze direction relative to own body position; compensation for self or object motion, thereby stabilizing the gaze; scanning movements; and fixational eye movements counteracting adaptation. Highly interesting, three of the four types of eye movements seem to be present in A. planci (Beer et al. 2016).
The individual eyes of A. planci have a broad horizontal visual field of about 100°, but it only spans approx. 20° vertically. Still, the terminal tube foot holding the eye is placed on a bendable nob on the arm tip allowing changes of the vertical part of the gaze direction. It was found that on the arms leading at the given time, the gaze direction was centered a few degrees above the horizon, which matches the desire to detect large coral boulders, their habitat, on the seafloor. The arms trailing at the given time, on the other hand, were held flat along the substrate but with the nob at the tip of the arm bent upward and the eye gazing approx. 45° above horizon possibly looking out for predators (Beer et al. 2016). Further, when traversing an obstacle movement of the nob would counteract 60–70% of the arm movements, thereby stabilizing the gaze direction (Beer et al. 2016). Lastly, A. planci raises and lowers the leading arms in a rhythmic way, which results in the eyes changing the vertical part of their gaze direction about 6° in a little more than 2 sec. This matches the temporal and spatial resolution and is, thus, putatively used as fixational eye movements refreshing the image on the retina and enhanced horizontal contrast lines without compromising spatial resolution (Beer et al. 2016; Petie et al. 2016b).
Even though active vision through eye movements in starfish has only been examined for A. planci so far, we predict that it is a common feature of starfish vision, since most visual tasks will benefit from this. The presence of these advanced aspects of vision stresses that starfish are highly dependent on visual information but also that complex neuronal circuitry is present providing the feedback needed to control the movements.
4.4.4 Other Starfish Behaviors Putatively Involving Vision
There are major gaps in our knowledge of starfish behavioral repertoires, and when it comes to which senses control the known behaviors, our knowledge is also limited (Garm 2017). As illustrated above, surprisingly few controlled experiments have tested their visually guided behaviors when considering that their eyes were discovered more than 200 years ago. To fully appreciate the starfish visual ecology, new hypothesis driven behavioral experiments are warranted. One of the highly interesting aspects of starfish ecology, which has only been studied in little details, is the bioluminescence found in some deep-sea species (Birk et al. 2018; Henning 1974). Especially members of the order Brisingida should be studied further, since they combine the ability to emit light with prominent eyes and relatively high spatial resolution compared to other starfish (Birk et al. 2018). A plausible explanation for this combination could be that they use the bioluminescence for visual communication in the darkness of the deep sea where finding a mate is often problematic. If true, this would take starfish vision and visually guided behaviors to a new level of complexity. The shallow water Antarctic species Odontaster validus does include photoreception in its reproductive behavior, but likely only extraocular photoreception measuring day length to coordinate gamete release (Pearse and Bosch 2002; Pearse et al. 1986). Several nonluminescent deep-sea starfish also have prominent eyes (Birk et al. 2018), and since many of them are predators, they might use vision to detect bioluminescent prey such as deep-sea corals. Vision could also be involved in prey detection in shallow water species, but considering the known spatial and temporal resolution, this would have to be rather large and stationary prey items. All these testable hypotheses are awaiting experimental proof, but several other hypotheses and ideas will undoubtedly emerge once more species and behaviors have been examined.
4.5 Multimodal Control of Behavior
It has been shown within the latest years that starfish can use a combination of rheotaxis, olfaction, and true vision to navigate the ocean floor (Petie et al. 2016b; Sigl and Laforsch 2016; Sigl et al. 2016). In a previously unpublished study, we investigated this further especially to gain information on the potential sensory hierarchy of such multimodal behavior. In a behavioral arena, M. glacialis was presented with either a rheotactic stimulus (semi-laminar flow, n = 12), visual stimulus (black area on white background, n = 12), olfactory stimulus (prey scent) in a semi-laminar flow (n = 13), combined rheotactic and visual stimuli (diverging 110°, n = 14), or combined olfactory, rheotactic, and visual stimuli (diverging 110°, n = 10). In the arena, M. glacialis showed a clear need for multimodal sensory input and walked randomly if only a single stimulus was present (Fig. 4.8). Interestingly, they also displayed a sensory hierarchy and were attracted to the visual stimulus when an odorless semi-laminar flow was present, and this is irrespective of the direction to the black area relative to the current. When a prey scent was added to the flow, this combined stimulus became attractive and overruled the visual input (Fig. 4.8). This supports that M. glacialis is mainly olfactory guided but also that other senses, like vision, take over if no reliable olfactory cue is present. This novel data are in contrast to A. planci where one study found that vision is not only necessary but also sufficient to navigate to their habitat in low current situation (Petie et al. 2016a). When a stronger directional current is present but with no visual cues, A. planci seems to use rheotaxis to find the coral boulders putatively including olfactory cues (Sigl et al. 2016). There is, thus, a high chance that starfish with ecologies differing from that of M. glacialis and A. planci will combine their senses in other ways with different hierarchal orders possibly also including other senses than vision, olfaction, and rheotaxis.

Sensory hierarchy in Marthasterias glacialis. (a) When only presented with a visual stimulus (black circle at 0°), M. glacialis walks randomly in the behavioral arena. (b) When presented with both a visual stimulus and a semi-laminar flow (large black arrow indicates flow direction), M. glacialis showed negative phototaxis and walked toward the black visual cue. (c) When a chemical stimulus (prey odor) was added to the semi-laminar flow (red arrow), M. glacialis abandoned negative phototaxis and displayed an axial response with either positive or negative chemotaxis. Central black arrow = mean vector of all individual headings (blue dots), r = length of mean vector, pr = result of Raleigh test, broken lines = 95% confidence interval, N = number of test animals
4.6 Processing of the Visual Information
Due to their secondary radial symmetry, adult starfish have no cephalization with a single body region, where nervous tissue and most senses are concentrated. Nonetheless, detailed morphological and physiological studies have proven that these animals still possess a central nervous system (CNS) or “brain” – it is just organized differently as what could be called a dispersed CNS. Besides the recognition of a dispersed CNS, though, we are still in the very early days of experimentally testing its functionality.
When a starfish explores the seafloor, they have no preset leading arm and all arms take turns leading the animal (Pearse et al. 1987). This is likely one of the reasons for the senses in starfish, including the eyes, being dispersed with a repetition on each arm. As every arm around the body is equally receptive to environmental stimuli, the sensory processing and integration seemingly need to follow the same pattern. Starfish do have a well-defined CNS including a nerve ring encircling the mouth, but the major part of the CNS is the radial nerve cords (RNC), dispersed with a repetition in every arm (Fig. 4.9b), and this is putatively where the visual information is processed. The nerve ring appears to function at least in part as a means of communication between the RNCs, which is backed by our unpublished results from A. planci. When bisecting the nerve ring on opposite sides of the animals, they became quiescent at first. After about 10 min, the animals started moving, but it was obvious that the two halves of the animals could not communicate and coordinate locomotion. On several occasions, the result was the starfish pulling itself apart with the two halves afterward walking “normally” around the tank (Petie and Garm, unpublished results).

The starfish central nervous system (CNS). (a) A schematic drawing of a cross section through the radial nerve cord (RNC) in starfish – not to scale. Arrowheads show neural connection between the ectoneural and hyponeural area. Aggregations of giant neurons (g) are drawn as seen in Acanthaster planci. Three other types of neurons are indicated: N1 is a neuron located in the neuroepithelium in the ectoneural area, N2 is a neuron with the cell body located in the neuropil, and N3 is a putative sensory neuron. (b) Schematic drawing of the starfish central nervous system comprising a RNC in each arm connected to a circumoral nerve ring (NR). CT = connective tissue, HC = hyponeural canal, HS = haemal sinus, NP = neuropil, S = supporting cells, W = water vascular system
4.6.1 Structure of the Starfish CNS
The echinoderm CNS consists of a hyponeural area and an ectoneural area, and in starfish, the entire CNS (nerve ring and RNC) has this division, which is not the case for all echinoderm classes (Engle 2013; Mashanov et al. 2006; Märkel and Röser 1991; Viehweg et al. 1998; Zueva et al. 2018). The ectoneural area makes up the oral side part of the RNC and has by far the most neurons. The hyponeural area overlays the aboral surface of the ectoneural area (Viehweg et al. 1998) (Fig. 4.9). It was initially thought that the two areas were completely separated by a sheath of connective tissue including a basal lamina, but recent data from several echinoderm classes (Holothuroidea, Ophiuroidea, and Asteroidea) have found neural bridges in the connective tissue connecting the two areas (Mashanov et al. 2006; Zueva et al. 2018) (Fig. 4.9a). The starfish CNS lies orally to the water vascular and the haemal systems. In the arms, the connective tissue surrounding the haemal sinus continues orally and separates the hyponeural area in two also forming a two parted hyponeural canal (Fig. 4.9a). Little is known about the function of this canal, but it might be involved in maintenance of the RNC removing waste products and supplying nutrients. In other echinoderm classes, an additional ectoneural canal is present, and oddly enough in echinoids it is open to the external environment, giving access for seawater to enter (Märkel and Röser 1991).
The CNS is connected by peripheral nerves to different areas of the body. Peripheral nerves from the RNC in starfish innervates the spines, the spine-free zone of the interambulacrum, and all the tube feet including the distal-most sensory tube foot with the eye (Formery et al. 2021). Little is known from starfish, but in brittle stars, the peripheral nerves can originate from either the ectoneural area, the hyponeural area, or a mix of both (Zueva et al. 2018). Across echinoderms, the size of the hyponeural system is correlated with the amount of muscle tissue (Mashanov et al. 2016), and it is believed to be involved in motor control only (Cobb 1995; Cobb and Stubbs 1981). This is questioned by the presence of putative sensory cilia extending from the neuroepithelium and into the hyponeural canal (Fig. 4.9a), which has been observed in several echinoderm species (Cobb 1987; Mashanov et al. 2006; Viehweg et al. 1998). Based on their ultrastructure and position inside the canals, these cilia have been proposed to be proprioceptors or chemoreceptors. Still, the ectoneural system comprises by the far the most neurons of which many are sensory neurons (Cobb 1987) and the ectoneural area is the only part of the RNC that continues all the way to the distal-most tube foot at the tip of the arm where it directly contacts the compound eye (Fig. 4.10) (Moss et al. 1998). In total, morphological evidence strongly suggests that visual processing happens in the ectoneural part of each RNS.

Nervous system in the eye of Asterias rubens visualized with anti-tubulin immunostaining. (a) The eye (encircled by broken white line) sits as a direct extension of the radial nerve cord (RNC). Nerves from the ommatidia (o) collect beneath the eye (arrowhead) and contact the RNC. (b) Close-up of a couple of ommatidia and the nerve beneath the eye. Broken line indicates an ommatidium; the dark part is where the screening pigment is situated. Arrowhead points at nerve connecting to the eye
4.6.2 The Ectoneural Part of the RNC
The space between the neuroepithelia is filled with neurites (the existing morphological data does not allow us to differentiate between axons and dendrites) and cell bodies of neurons, and this neuropil is partially divided into separate compartments by processes from the supporting cells (see below). In both the hypo- and the ectoneural area, most of the neurites run parallel with the RNC, but some run transversally (Fig. 4.11c). Synapses are omnipresent in the ectoneural part of the RNC as indicated by the putative synaptic vesicles, which again supports that this is an area of information processing. There are three types of synaptic vesicles all 100–150 nm in diameter – electron lucent vesicles, electron dense vesicles, and dense-cored vesicles – and in some neurites, more than one type of vesicle is present (Fig. 4.11d).

TEM micrographs showing the organization of the distal part of the radial nerve cord (RNC) in Acanthaster planci. (a) Most of the approx. 80-μm-thick RNC is filled with ectoneural (e) neuropil (NP) occasionally transversed by fiber bundles (FB) from the supporting cells. Note that toward the hyponeural part, there is an aggregation of giant neurons (GN). (b) The FB extend from the neuropil to the basal lamina (BL) where they attach with hemidesmosomes (H). (c) Most neurons run parallel with the longitudinal axis of the RNC (cross-sectioned here), but some transverse the neuropil area (TN). (d) The neuropil displays a high number of synapses as indicated by putative synaptic vesicles. Different types of vesicles are seen: electron lucent vesicles (LV), dense vesicles (DV), and dense-cored vesicle (DCV). G = glycocalyx, NE = neuroepithelium, HN = hyponeural area, CT = connective tissue, CF = collagen fiber
Most neurites found in the ectoneural part of the starfish RNC are quite small typically between 100 and 600 nm in diameter depending on the species (Fig. 4.11). This small diameter is a general echinoderm trait, and it has made it hard to accomplish any form of electrophysiological work on the CNS. To our knowledge, the only published recordings from echinoderm CNS so far come from the larger (giant) neurons of brittle stars (Cobb and Moore 1989). These neurons are 10–20 μm in diameter and are of two types based on the electrophysiological data. The largest neurons alerted the whole animal, and their activity resulted in a “freezing” state of the body with the arms stiffening. The other group of giant neurons was active during behaviors such as escape and feeding. Importantly, the work showed that depending on the type of stimuli, the sensory integration happened either throughout the entire CNS or locally in a particular RNC (Cobb and Moore 1989). Whether this is a general echinoderm feature or specific to brittle stars is not known though.
An open question about the functional organization of the starfish RNC is if there are regional specializations where a certain type of information is processed. To look for such regions, we compared the morphology of the basal (close to the nerve ring), middle, and distal part of the RNC of three species: A. planci (n = 3), A. rubens (n = 3), and A. irregularis (n = 3) (Fig 4.12). Not surprisingly, we found a correlation between the absolute number of neurites and the size of the animal, with the largest species, A. planci, displaying the most and the small A. irregularis the least neurites. There was an interesting difference between species, though, in that the two eye carrying species, A. planci and A. rubens, displayed very similar patterns. They had a close to linear decline in the number of neurites from the basal to the distal part of the RNC (Fig. 4.12d, g). In the eyeless A. irregularis, on the other hand, the ectoneural area in the distal part of the RNC seemed swollen and had the highest number of neurites (Fig. 4.12a–c). We do not know the exact number of neurons as the neurites can be branches of dendrites and/or axons, but it is likely significantly less than the millions of neurites we count. Still, there is putatively a correlation between the relative number of neurites and the amount of processing indicating a processing hotspot distally in the RNC of A. irregularis. This came as a surprise since A. rubens and A. planci have the same putative chemosensory and tactile tube feet distally on the arm as A. irregularis, and adding visual information thus appears to reduce the need for processing power. A part of the explanation could be that most of the visual information is processed already in the distal-most tube foot carrying the eye. This tube foot does hold a specialized nervous system laying in direct extension of the RNC as seen by our antibody stains (Fig. 4.10). Further, when compared to the eyeless distal tube feet in sea cucumber and sea urchin, the eye carrying starfish tube foot has a much denser nerve plexus with circumferential and longitudinal condensation (Formery et al. 2021).

Distribution of neurites along the radial nerve cord (RNC). (a–c) In Astropecten irregularis, the highest number of neurites in the RNC is found in the distal part (c), and the middle part (b) has the lowest number. (d–f) Asterias rubens in general has a higher number of neurites in the RNC than A. irregularis, and the highest number is here found in the basal part (e). The distal part has the lowest number (f). (g–i) Following its larger size, Acanthaster planci has the most neurites in the RNC of the three species (more than 4 million basally), and as in A. rubens, the highest number is found basally (i) and the lowest distally close to the eye (j). Arrowhead shows aggregation of giant neurons, which appear as lighter areas of the neuropil. E = ectoneural area, Hy = hyponeural area. Error bars in (a, d) and (g) indicate SD
As in the RNC of brittle stars, we also found giant neurons in the ectoneural part of the starfish RNC, though somewhat smaller (approx. 3–6 μm in diameter). In both A. planci and A. rubens, the number of giant neurons increases toward the distal part of the RNC (Fig. 4.12h, i). Furthermore, in the middle and distal parts of the RNC, the giant neurons were concentrated in subsystems in the central area, which is the area contacting the distal-most tube foot and the eye (Fig. 4.12f). As this pattern was not found in A. irregularis, it could indicate that the giant neurons play a part in the visual processing and integration. Still, we are only just beginning to understand the functional organization of the starfish CNS, and particularly electrophysiological data is warranted before any conclusions can be made about where and how the visual processing happens.
4.6.3 Supporting Cells
Both the ectoneural and hyponeural area have a neuroepithelium that surrounds the neuropil with intermingled supporting cells. The supporting cells have a fiber bundle in the cell body extending into a basal process and for the bipolar cells an additional apical process. The processes run through the neuropil area, where the apical process runs into the neuroepithelium and the basal process toward the basal lamina where they attach with hemidesmosomes (Figs. 4.9 and 4.11). We found no differences between A. planci, A. rubens, and A. irregularis, and the organization is similar in other echinoderms (Mashanov et al. 2010).
The function of the supporting cells is not fully understood, but they seem important for several processes in the RNS and thus potentially also for the visual processing. Bargmann and Behrens first suggested that the supporting cells in A. rubens are glial-like cells, but this was subsequently disputed (Bargmann and Behrens 1963; Cobb 1995; Cobb and Stubbs 1981). However, it has been shown in both starfish and sea cucumbers that they produce a material similar to Reissner’s substance found in secretory glial cells in chordates. They also share morphological features with the radial glia of chordates found in the embryonic CNS, which for some chordates persist into adulthood (Mashanov et al. 2009; Viehweg et al. 1998). Additionally, the supporting cells of holothurians play important roles in both RNC regeneration after injury and in nervous tissue growth in general. During regeneration, the supporting cells near the lesion lose the processes and dedifferentiate. The processes are phagocytosed by the surrounding cells, and the remaining cell body becomes highly proliferative and an important source of new supporting cells and neurons (Mashanov et al. 2008, 2015).
Even if the supporting cells in some aspect resemble glial cell, they do not encircle the neurons as seen in chordates, and it is thus hard to imagine that they have the same function supporting the neurotransmission (Hartline and Colman 2007). Cnidarians also appear to lack glial sheaths in their CNS, and this lack allows neurons running in parallel in the hydrozoan nerve ring to enhance the signal by exciting each other, a phenomenon known as piggybacking (Mackie 2004). Whether this also happens in the starfish RNC is currently unknown.
4.6.4 Specializations in A. planci: Neural Bulbs on the RNC
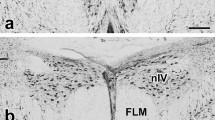
It was recently discovered that A. planci has bulbous structures protruding from the ectoneural surface of the RNC (Smith 2018) (Fig. 4.13). These structures have so far not been observed in other species of echinoderms. The neural bulbs are found every approx. 50 μm along the entire length of the RNC, and they are up to 170 μm long. The function of these neural bulbs is still not completely understood, but they seem to be integrated parts of the RNCs and thus potentially involved in sensory processing. The center of the neural bulb is mostly filled with neurites connected to the RNC, but they are slightly larger than the average RNC neurites, 0.5–1.1 μm in diameter (Fig. 4.13d). The neuroepithelium of the ectoneural area also covers the neural bulb, and a high number of putative mechanosensory cilia with a collar of microvilli are found in this part of the neuroepithelium (Fig. 4.13b). Besides these putative sensory cells, the neuroepithelium of the bulbs contains many secretory cells (Smith 2018) (Fig 4.13c).

Structure of the neural bulbs on the RNC of Acanthaster planci. (a) Cross section of the RNC showing two neural bulbs (NB). (b) Putative sensory cell in the NB epithelium displaying a ciliary rootlet (C) and a collar of microvilli (M) around the cilium. (c) Secretory cell with vesicles (SV) filled with granular material (G) secreted into the mucus layer on the outside of the NB epithelium. (d) The center of the neural bulb is filled with neurites (N) and fiber bundles (FB) crossing the bulb in different directions. ML = mucus layer, NP = neuropil, NE = neuroepithelium, Nu = nucleus, V = vacuole
Interestingly, juveniles of A. planci do not have the neural bulbs, and their emergence seems correlated with a shift in diet. Juveniles feed on algae, but when they reach a size of 5–8 cm in diameter, they start feeding on hard corals and this is when the bulbs start to appear. It has thus been suggested that the sensory system of the bulbs along with the high secretory activity is used in counteractions against the cnidocytes and other defense mechanisms of the corals (Smith 2018).
4.7 Concluding Remarks
Most starfish have a prominent compound eye at the tip of each arm, and recent evidence suggests that they are not as strictly olfactory guided as previously suggested (Dale 1997; Garm and Nilsson 2014). Even in the deep sea, the eyes persist in many species indicating that detection of bioluminescence plays an important role also for some starfish. Their visual capacity seems to be closely linked to their ecology and behavioral needs, with low pass spatial and temporal filtering removing most except for large and stationary objects from sight. The behavioral evidence points out negative phototaxis and habitat detection as important visually guided behaviors but also that vision is part of sensory hierarchy that can be dominated by olfaction. A great enigma when it comes to starfish vision is where and how the visual information is processed. The radial symmetry of starfish results in a CNS where a central brain with distinct sensory specific centers is missing. Instead the evidence suggests that each eye carrying arm has its own processing center, the radial nerve cord (RNC). Here we have presented some of the first data on the structure of starfish RNC and shown how it differs between eye carrying and eyeless species, but the interpretation still suffers from a complete lack of physiological data.
Interestingly, the echinoderm CNS resembles the cnidarian CNS especially what is found in Cubomedusae. The cnidarian CNS is also radially symmetric, and a major part is a nerve ring encircling the mouth (Garm et al. 2006; Mackie 2004). Furthermore, Cubomedusae have four sensory structures called rhopalia, which each holds an additional part of the CNS, the rhopalial nervous system (RNS), connected to the ring nerve. The rhopalia also carries a set of six eyes each, and the visual information processing putatively happens in the RNS (Bielecki et al. 2013; Garm and Mori 2009; Nilsson et al. 2005). This arrangement has a strong resemblance to starfish with each rhopalium putatively paralleling an arm. The combined physiological, behavioral, and modeling data from cubomedusae shows that behavioral control is not accomplished through a collaboration between the four rhopalia but rather as a competition. At any given time, the rhopalium receiving the strongest stimulation overrules the others through a resetting mechanism and becomes the sole control unit (Satterlie and Nolen 2001; Satterlie and Spencer 1979; Stöckl et al. 2011). Whether the same is the case for starfish is still to be tested, but it is currently the most likely hypothesis, and, thus, starfish are in many ways a collection of semi-independent arms!
References
Bargmann W, Behrens B (1963) Über den Feinbau des Nervensystems des Seesterns (Asterias rubens L.). Zeitschrift für Zellforschung und Mikroskopische Anatomie 59:746–770
Beer S, Wentzel C, Petie R, Garm A (2016) Active control of the visual field in the starfish Acanthaster planci. Vision Res 127:28–34
Bielecki J, Nachman G, Garm A (2013) Swim pacemaker response to bath applied neurotransmitters in the cubozoan Tripedalia cystophora. J Comp Physiol A 199:785–795
Birk M, Blicher ME, Garm A (2018) Deep-sea starfish from the Arctic have well developed eyes in the dark. Proc R Soc London B Biol Sci 285:20172743. https://doi.org/10.1098/rspb.2017.2743
Boivin Y, Larrivée D, Himmelman JH (1986) Reproductive cycle of the subarctic brooding asteroid Leptasterias polaris. Mar Biol 92:329–337
Castilla J, Crisp D (1973) Responses of Asterias rubens to water currents and their modification by certain environmental factors. Netherlands J Sea Res 7:171–190
Chia F, Koss R (1994) Asteroidea. In: Harrison FW, Chia F-S (eds) Echinodermata, vol 14, pp 169–246
Clark EG, Kanauchi D, Kano T, Aonuma H, Briggs DEG, Ishiguro A (2019) The function of the ophiuroid nerve ring: how a decentralized nervous system controls coordinated locomotion. J Exp Biol 222:jeb192104. https://doi.org/10.1242/jeb.192104
Clements M, Wolfe K, Schwartz K, Bryne M (2019) Forever fissiparous: asexual propagation and stable demography in a tropical and geographically isolated asterinid sea star. Mar Biol 166. https://doi.org/10.1007/s00227-019-3518-0
Cobb JLS (1987) Neurobiology of the echinodermata. In: Nervous systems in invertebrates, Springer, Boston, pp 483–525
Cobb JLS (1995) The nervous systems of Echinodermata: recent results and new approaches. In: The nervous systems of invertebrates: an evolutionary and comparative approach, Birkhäuser, Basel, pp 407–424
Cobb JLS, Moore A (1989) Studies on the integration of sensory information by the nervous system of the brittlestar Ophiura ophiura. Mar Freshw Behav Physiol 14:211–222
Cobb JLS, Stubbs TR (1981) The giant neurone system in ophiuroids I. The general morphology of the radial nerve cords and circumoral ring. Cell Tiss Res 219:197–207
Cowan Z-L, Pratchett M, Messmer V, Ling S (2017) Known predators of crown-of-thorns starfish (Acanthaster spp.) and their role in mitigating, if not preventing, population outbreaks. Diversity 9. https://doi.org/10.3390/d9010007
Dale J (1997) Chemosensory search behavior in the starfish Asterias forbesi. Biol Bull 193:210–212
D'Aniello S, Delroisse J, Valero-Garcia A, Lowe EK, Byrne M, Cannon JT, Halanych KM, Elphick MR, Mallefet J, Kaul-Strehlow S et al (2015) Opsin evolution in the Ambulacraria. Mar Genom 24:177–183
De'ath G, Moran PJ (1998) Factors affecting the behaviour of crown-of-thorns starfish (Acanthaster planci L.) on the Great Barrier Reef 2: feeding preferences. J Exp Mar Biol Ecol 220:107–126
Döring CC, Kumer S, Tumu SC, Kourtesis I, Hausen H (2020) The visual pigment xenopsin is widespread in protostome eyes and impacts the view on eye evolution. eLife 9:e55193. https://doi.org/10.7554/eLife.55193
Engle S (2013) Ultrastructure and development of the body cavities in Antedon bifida (Pennant, 1777) (Comatulida, Crinoida), vol. PhD
Fabricius KE, Okaji K, De'ath G (2000) Three lines of evidence to link outbreaks of the crown-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral reefs 29:593–605
Fenchel T (1965) Feeding biology of the sea star Luidia sarsi Düben and Koren. Ophelia 2:223–236
Formery L, Orange F, Formery A, Yaguchi S, Lowe CJ, Schubert M, Croce JC (2021) Neural anatomy of echinoid early juveniles and comparison of nervous system organization in echinoderms. J Comp Neurol 529:1135–1156
Fritsches KA, Brill RW, Warrant EJ (2005) Warm eyes provide superior vision in swordfishes. Curr Biol 15:55–58
Garm A (2017) Sensory biology of starfish - with emphasis on recent discoveries in their visual ecology. Integrat Comp Biol. https://doi.org/10.1093/icb/icx086
Garm A, Mori S (2009) Multiple photoreceptor systems control the swim pacemaker activity in box jellyfish. J Exp Biol 212:3951–3960
Garm A, Nilsson DE (2014) Visual navigation in starfish: first evidence for use of vision and eyes in starfish. Proc R Soc London B Biol Sci 281:20133011. https://doi.org/10.1098/rspb.2013.3011
Garm A, Ekström P, Boudes M, Nilsson DE (2006) Rhopalia are integrated parts of the central nervous system in box jellyfish. Cell Tiss Res 325:333–343
Govardovskii VI, Fyhrquist N, Reuter T, Kuzmin DG, Donner K (2000) In search of the visual pigment template. Vis Neurosci 17:509–528
Hall M, Kocot KM, Baughman KW, Fernandez-Valverde SL, Gauthier MEA, Hatleberg WL, Krishman A, McDougall C, Motti CA, Shoguchi E et al (2017) The crown-of-thorns starfish genome as a guide for biocontrol of this coral reef pest. Nature 544:231–234
Hamel J, Mercier A (1994) Influence of environmental factors on prespawnign behaviour, spawning and developmental biology of the brooding starfish Leptasterias Polaris. In: David B, Guille A, Feral J, Roux M (eds) Echinoderms through times. Balkema, Rotterdam, p 310
Hartline DK, Colman DR (2007) Rapid conduction and the evolution of giant axons and myelinated fibers. Curr Biol 17:R29–R35
Henning PJ (1974) New observations on the bioluminescence of echinoderms. L Zool London 172:401–418
Jourdain S (1865) On the eyes of Asteracanthion rubens, Müll. & Trosch. (Uraster rubens, Forbes). Annals Magazine Nat His 15:238–240
Kalmus H (1929) Versuche fiber die Bewegungen der Seesterne, besonders von Asterina gibbosa. Zool Vergleich Phsiol 9:703–733
Kirwan JD, Bok MJ, Smolka J, Foster JJ, Hermandez JC, Nilsson DE (2018) The sea urchin Diadema africanum uses low resolution vision to find shelter and deter enemies. J Exp Biol 221. https://doi.org/10.1242/jeb.176271
Korsvig-Nielsen C, Hall M, Motti CA, Garm A (2019) Eyes and negative phototaxis in juvenile crown-of-thorns starfish, Acanthaster species complex. Biol Open 8. https://doi.org/10.1242/bio.041814
Land MF (2019) Eye movements in man and other animals. Vision Res 192:1–7
Land MF, Nilsson DE (2012) Animal eyes. Oxford University Press, Oxford
Lascalli T (2000) Larval budding, metamorphosis, and the evolution of life-history patterns in echinoderms. Invert Biol 119:234–241
Lesser MP, Carleton KL, Böttger SA, Barry TM, Walker CW (2011) Sea urchin tube feet are photosensory organs that express a rhabdomeric-like opsin and PAX6. Proc R Soc London B Biol Sci 278:3371–3379
Lowe EK, Garm A, Ullrich-Lüter EM, Cuomo C, Arnone MI (2018) The crowns have eyes: Multiple opsins found in the eyes of the Crown-of-Thorns Starfish Acanthaster planci including the first r-opsin utilized by a deuterostome eye. BMC Evol Biol Accepted for publication
Lucas JS (2013) Crown-of-thorns starfish. Curr Biol 23:945–946
Lythgoe JN (1979) The ecology of vision. Clarendon Press, Oxford
Mackie GO (2004) Central neural circuitry in the jellyfish Aglantha: a model “simple nervous system”. Neuro-Signals 13:5–19
Märkel K, Röser U (1991) Ultrastructure and organization of the epineural canal and the nerve cord in sea urchins (Echinodermata, Echinoida). Zoomorphol 110:267–279x
Mashanov VS, Zueva OR, Heinzeller T, Dolmatov IY (2006) Ultrastructure of the circumoral nerve ring and the radial nerve cords in holothurians (Echinodermata). Zoomorphol 125:27–38
Mashanov VS, Zueva OR, Heinzeller T (2008) Regeneration of the radial nerve cord in a holothurian: a promising new model system for studying post-traumatic recovery in the adult nervous system. Tiss Cell 40:351–375
Mashanov VS, Zueva OR, Heinzeller T, Aschauser B, Neumann WW, Grondoda JM, Cifuentes M, Garcia-Arrarás JE (2009) The central nervous system of sea cucumbers (Echinodermata: Holothuroidea) shows positive immunostaining for a chordate glial secretion. Front Zool 6. https://doi.org/10.1186/1742-9994-6-11
Mashanov VS, Zueva OR, Garcia-Arrarás JE (2010) Organization of glial cells in the adult sea cucumber central nervous system. Glia 58:1581–1593
Mashanov VS, Zueva OR, Garcia-Arrarás JE (2015) Myc regulates programmed cell death and radial glia dedifferentiation after neural injury in an echinoderm. BMC Dev Biol 15:1–9
Mashanov VS, Zueva OR, Rubilar T, Epherra L, Garcia-Arrarás JE (2016) Echinodermata. In: Schmidt-Rhaesa A, Harzsch S, Purschke G (eds) Structure and Evolution of Invertebrate Nervous Systems. Oxford University Press, Oxford
McFarland WN, Munz FW (1975) Part II The photic environment of clear tropical seas during the day. Vision Res 15:1063–1070
Moore S, Thorndyke MC (1993) Immunocytochemical mapping of the novel echinoderm neuropeptide SALMFamide 1 (S1) in the starfish Asterias rubens. Cell Tiss Res 274:605–618
Moran PJ (1986) The Acanthaster phenomenon. Oceanograph Mar Biol Annu Rev 24:379–480
Moss C, Jackie Hunter A, Thorndyke MC (1998) Patterns of bromodeoxyuridine incorporation and neuropeptide immunoreactivity during arm regeneration in the starfish Asterias rubens. Philos Trans R Soc London 353:421–436
Motti CA, Bose U, Roberts RE, McDougall C, Smith MK, Hall M, Cummins SF (2018) Chemical ecology of chemosensation in Asteroidea: insights towards management strategies of pest species. J Chem Ecol 44:147–177
Mueller B, Bos AR, Graf G, Gumanao GS (2011) Size-specific locomotion rate and movement pattern of four common Indo-Pacific sea stars (Echinodermata; Asteroidea). Aquat Biol 12:157–164
Nilsson DE, Gislén L, Coates MM, Skogh C, Garm A (2005) Advanced optics in a jellyfish eye. Nature 435:201–205
Pearse JS, Bosch I (2002) Photoperiodic regulation of gametogenesis in the Antarctic sea star Odontaster validus Koehler: evidence for a circannual rhythm modulated by light. Invert Repro Dev 41:73–81
Pearse JS, Eernisse DT, Pearse VB, Bauchamp KA (1986) Photoperiodic regulation of gametogenesis in sea stars, with evidence for an annual calendar independent of fixed daylength. American Zool 26:417–431
Pearse V, Pearse J, Buchsbaum M, Buchsbaum R (1987) Living invertebrates. Blackwell Science Inc.
Penn PE, Alexander CG (1980) Fine structure of the optic cusion in the asteroid Nepanthia belcheri. Mar Biol 58:251–256
Petie R, Garm A, Hall M (2016a) Crown-of-thorns starfish have true image forming vision. Front Zool 13:41. https://doi.org/10.1186/s12983-016-0174-9
Petie R, Hall M, Hyldahl M, Garm A (2016b) Visual orientation by the crown-of-thorns starfish (Acanthaster planci). Coral reefs 35:1139–1150. https://doi.org/10.1007/s00338-016-1478-0
Satterlie RA, Nolen TG (2001) Why do cubomedusae have only four swim pacemakers? J Exp Biol 204:1413–1419
Satterlie RA, Spencer AN (1979) Swimming control in a cubomedusan jellyfish. Nature 231:141–142
Scheibling RE (1981) Optimal foraging movements of Oreaster reticulatus (L.) (Echinodermata: Asteroidea). J Exp Mar Biol Ecol 51:173–185
Sigl R, Laforsch C (2016) The influence of water currents on movement patterns on sand in the crown-of-thorns seastar (Acanthaster cf. solaris). Diversity 8:25. https://doi.org/10.3390/d8040025
Sigl R, Streibl S, Laforsch C (2016) The role of vision for navigation in the crown-of-thorns seastar Acanthaster planci. Sci Rep 6:30834. https://doi.org/10.1038/srep30834
Sloan NA (1980) The arm curling and terminal tube-foot response of the ateroid Crossaster papposus. J Nat Hist 14:469–482
Smith MK (2018) Molecular investigations of the neural system in the Crown-of-Thorns Seastar, Acanthaster planci species-complex, vol. PhD. University of Queensland
Stöckl AL, Petie R, Nilsson DE (2011) Setting the pace: new insights into central pattern generator interactions in box jellyfish swimming. PLoS ONE 6:1–11
Sumner-Rooney L, Kirwan JD, Lowe EK, Ullrich-Lüter EM (2020) Extraocular vision in a brittle star Is mediated by chromatophore movement in response to ambient light. Curr Biol 30:319–327
Ullrich-Lüter EM, Dupont S, Arboleda E, Hausen H, Arnone MI (2011) Unique system of photoreceptors in sea urchin tube feet. In: Proceedings of the National Academy of Sciences USA, vol 108, pp 8367–8372
Ullrich-Lüter EM, D'Aniello S, Arnone MI (2013) C-opsin Expressing Photoreceptors in Echinoderms. Integrat Comp Biol 53:27–38
Valentincic T (1975) Amino-acid chemoreception and other releasing factors in the feeding response of the sea star Marthasterias glacialis (D.). In: Barnes H (ed) Proceedings from the ninth European marine biological symposium. University of Aberdeen press, Aberdeen, pp 693–705
Viehweg J, Khoury M, Heinzeller T, Mashanova D, Mashanov VS (1998) Secretory radial glia in the ectoneural system of the sea star Asterias rubens (Echinodermata). Acta Zool 92:119–131
Vöcking O, Kourtesis I, Tumu SC, Hausen H (2017) Co-expression of xenopsin and rhabdomeric opsin in photoreceptors bearing microvilli and cilia. eLife 6:e23435
Yoshida M, Ohtsuki H (1968) The phototactic behavior of the starfish Asterias amurensis Lütken. Biol Bull, pp 516–532
Zhang R, Wang C, Zhou Y, Zhang H (2019) Morphology and molecular phylogeny of two new species in genus Freyastera (Asteroidea: Brisingida: Freyellidae), with a revised key to close species and ecological remarks. Deep-Sea Res 1:154. https://doi.org/10.1016/j.dsr.2019.103163
Zueva OR, Khoury M, Heinzeller T, Mashanova D, Mashanov VS (2018) The complex simplicity of the brittle star nervous system. Front Zool 15:1–26
Acknowledgments
The authors appreciate the input from the editors and the anonymous reviewer. We would also like to acknowledge our coworkers from the results presented here: Dr. Ronald Petie, Prof. Dan-E Nilsson, Prof. Mike Hall, Dr. Martin F. Blicher, Mrs. Marie-Helene Birk, Mrs. Sabrina Beer, and Mrs. Camilla Wentzel.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Garm, A., Sundberg, D., Korsvig-Nielsen, C.E. (2023). Dispersed Vision in Starfish: A Collection of Semi-independent Arms. In: Buschbeck, E., Bok, M. (eds) Distributed Vision. Springer Series in Vision Research. Springer, Cham. https://doi.org/10.1007/978-3-031-23216-9_4
Download citation
DOI: https://doi.org/10.1007/978-3-031-23216-9_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-23215-2
Online ISBN: 978-3-031-23216-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)