
Abstract
This article reports the use of evolutionary game theory to understand the role of various factors underpinning the decision to confront in the competition for resources. One factor was the intrinsic rate of confrontation an organism would display in absence of context sensitive factors. Two other factors were responsive to the environment and two others to the health status of the organism. Factors were implemented as genes that determine the rate at which confrontation or cooperation would be selected. Organisms were evolving in environments of different levels of reward and punishment. At each cycle they would be paired with another organism and decide whether to confront or cooperate. We used a genetic algorithm to simulate the evolution of the gene pool over 500 cycles. The main finding is that the baseline rate of confrontation is responsive to the conditions in the environment. Our results also indicate that the decision to confront or cooperate depends not only upon the immediate competitive conditions (reward and punishment) in which organisms evolve but is also sensitive to the state of the organisms.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others


Keywords
1 Evolution of Cooperation and Confrontation
1.1 A Competition for Resources
Natural selection is the force that shapes organisms and the variety of their traits [1]. Competition for resources sharpens these traits making them increasingly adaptive in stable environments. Often, however, competition is indirect. Trees for example try to outgrow each other to capture more sunlight than the neighboring trees. But for many species, including homo sapiens, competition is direct. The bulk of research indicates that cooperation has emerged to increase our chances of survival [2,3,4]. Along with it the necessary increase in cognition has been the main driver behind the brain’s expansion [5], in particular the frontal cortices. Yet it is also clear that confrontation has not been eliminated. The passing on of successful genes to the next generation is often dependent upon the continuous confrontation with other members of the same species. There is ample evidence that confrontation has been part of human evolution as indicated by the numerous prehistoric sites documenting intentional (collective) violence [6,7,8]. Much of human history illustrates this principle at both the individual and group level. The classic example being Thucydides explaining the reasons of war between Sparta and Athens [9]. The rise of one power was challenging the domination by another and thus basically threatening to reduce its resources. In the 21st century the question of accessing resources has not changed and it is still the driver behind individual and group conflicts. Violence is often triggered when individuals estimate that the minimum number of resources necessary for survival will not be reached. As recently as 2018 the yellow vest movement in France was triggered by an increase in petrol taxes [10]. Suddenly a group formed from the collection of individual interests that was ready to confront the government. Throughout recorded history, and long before it, confrontations for resources have taken place. Investigating the factors that dictate the decision to confront or cooperate is thus essential to understanding violence between and within groups.
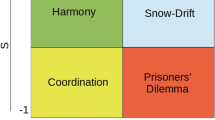
A question that the biological sciences faced in the 20th century was to explain how selfish organisms come to cooperate. A naïve interpretation of using confrontation as a means to acquire more resources would seem to suggest the conclusion that animals necessarily benefit from being aggressive. However, the gains from aggression are only valid as long as the competitor does not retaliate. Retaliation leads to a potential cost that might endanger the aggressor’s life. The balance between benefit and cost is axiomatic for determining the conditions underpinning cooperation or confrontation. It is at this stage that the theory of games started to play a crucial role in the investigation of cooperation in animals and humans. The paradigm has been widely used to explore and formalize human decision making in several academic disciplines. In its most basic setting, the so-called game refers to two individuals, A and B, facing a situation that involves two options, for example cooperating or confronting. Each individual makes a decision independently but is aware of the potential outcomes and that the same options are available to the other individual. Traditionally the game is represented as a matrix of choices, see Table 1.
The two-number vector in each cell represents the outcome for each of the two organisms. In the example showed in Table 1 we have the following outcomes. If both individuals decide to cooperate, they equally share the 6 points, each individual getting 3 points. If one individual decides to confront and the other one to cooperate, then the aggressor is rewarded with all the six points and the cooperative individual gains nothing. In cases where both decide to confront then a fight ensues, and they only get 1 point each. The objective of each individual is to maximize their gain. The choice or combination of choices that an individual will adopt to maximize gain is termed the strategy of that individual. Decisions, such as the one presented in Table 1, have largely been used to explore the conditions for cooperation [11]. One of the best-known thought experiments is the prisoner’s dilemma where the outcomes are penalties rather than gains. The paradigm has been instrumental in demonstrating that selfishness can drive individuals to make suboptimal decisions [12]. It is later work by Robert Axelrod that made a significant progress in our understanding of cooperation [12, 13]. Axelrod noted that the prisoner’s dilemma, like many other experiments within the paradigm, requires only one decision but in many real-life conditions individuals repeatedly interact with the same people, whether at home or at work, so it would be of interest to see how the strategy evolves when decisions are repeated. In such case, constant confrontation by both parties leads to penalties in the long run and thus systematically confronting is not a viable strategy. Axelrod and Hamilton have tested the efficiency of various strategies and demonstrated that the best one was a strategy consisting of offering cooperation in the first instance and then mirroring the behavior of the opponent. Since Axelrod’s pioneering works, the evolutionary game theory paradigm has been immensely successful in answering questions in numerous disciplines interested in cooperation and/or confrontation [14,15,16]. The present paper aims to investigate the decision to confront or cooperate with the same approach but will introduce further refinements into the modelling of the evolutionary process.
In this paper we use evolutionary genetic algorithms to investigate how living conditions affect confrontational rates in a virtual population of agents. The main manipulation is the use of different genes to code for different factors playing a role in the decision to confront. This approach permits estimating the relative importance and responsiveness of each component that plays a role in the choice of a behavioral strategy. The first and main trait we implement as genes is the inclination to confront. Though confrontational reactions are often triggered by environmental stimuli, the literature also suggests that confrontation has a genetic component [17]. The choice to confront is a multidimensional decision with many of its underpinning factors such as aggression and impulsivity including a genetic component [18]. Some combination of psychological traits, such as psychopathy, involve behavioral strategies incorporating confrontation as an option within their behavioral repertoire, making confrontations an adaptive strategy for humans. In this context, although the inclination to confront possibly results from the Gene-Environment interaction we will be considering it in this study as an inherited trait. The second manipulation will be a set of genes coding for our reactivity to the environment. How humans react to different conditions of gain and loss has been largely studied in behavioral economics [19]. It has been demonstrated that individuals tend to be risk tolerant in the domain of losses. For example, they usually prefer a 50% chance of losing £120 rather than losing £60 for sure. Individuals are also risk avoidant in the domain of gain so they would prefer to gain £60 for sure rather than a 50% chance of winning £120. When individuals face a situation where the outcome could be either positive or negative (so-called ‘mixed gambles’) they tend to be more risk tolerant. These responses result from evolutionary processes that promote the conservation of one’s own assets. Third, the last manipulation are genes that code for sensitivity to health status. Individuals that are weakened will tend to take less risk and avoid engaging in a confrontation as compared to those with a high health status who might be more inclined to confront.
In this context, the simulations carried out in this study aim to establish how the confrontation rate, defined as the number of confrontations per hundred decisions, varies as a function of the outcome. The second aim is to integrate the responsivity of moderating factors to our model of confrontation and cooperation. In the present paper we have implemented an evolutionary version of the experimental paradigm used in game theory to investigate how various factors underpinning the choice of confronting or cooperating are affected by different levels of reward and punishment.
2 Modelling the Evolution of Confrontation
2.1 Introduction
Our formal, simplified version of natural selection simulates an ecosystem of 1000 organisms. As the ecosystem is stable it systematically generates the same amount of food and thus can maintain the same number of organisms; thus, organisms that have disappeared get replaced before the next round of decisions. Each organism had initially 100 points of health. As the healthiest individuals (i.e., the top 10%) were selected for procreating, each organism was fighting in each cycle to potentially increase its health. Organisms that were paired had to decide whether to confront or cooperate. The combined decision of the two individuals in a pair created four potential outcomes that re-created the four conditions of game theory, see Table 2. The main difference between the original study by Axelrod and Hamiton [13] and the current implementation is that the decision is probabilistic, based upon the tendency of the individual to confront, rather than being stable over time and predictable. The probability to confront is δ and to cooperate is (1-δ).
We manipulated two variables, reward and the punishment, to evaluate how the different genes determining the value of δ vary. Reward and punishment were varied at each integer value between 1 and 100; creating 10,000 conditions (100 × 100). For each condition the rate of confrontation was initially equally distributed over the population. After 500 cycles of decisions, we recoded the gene profiles, the number of survivors in the 500th generation and their average health. We expected the genetic profile of the 500th generation to differ drastically from the equiprobable distribution used to define populations at cycle 0. The model was implemented in Python 3.7.
2.2 The Organisms
Individuals were defined as instances of a class organism defined by 3 properties. The first property, health was a score that varied from 0 to 100. Organisms start with Γ = 100, when the organism reaches a health of Γ = 0 it dies. The second property was age, noted τ, and was set as a counter of the number of cycles the organism has survived. Aging was implemented as a loss in health that is proportional to the number of cycles past the 25th cycle. For each cycle beyond 25, the organism would lose one more health point at that cycle. The third and most important property was genotype. Genotype was defined as 9 genes that coded five traits determining the probability of confrontation as reviewed above. The first trait was the natural propensity of organisms to use aggression to get resources. This tendency was coded by five genes that had an additive effect. Each of the genes had 5 alleles (A, B, C, D, & E) that coded for 5 different levels of confrontation (0%, 5%, 10%, 15%, & 20%), the genotype can thus vary from 0% (i.e., AAAAA) to 100% (i.e., EEEEE) and can take any value between that is a multiple of 5%, for example AABCA is 15% (0 + 0 + 5 + 10 + 0). The five genes together were setting the probability that an organism would respond to a decision by choosing to confront. This trait defined by a combination of five genes, termed confrontational propensity (CP), implements the natural variance in the inclination to confront.
The other four genes implemented the context sensitive modulation of the natural tendency to undertake confrontation or cooperation. Each of the other four genes uses the same five-letter coding (A, B, C, D, & E) and thus can modulate the intrinsic level of risk by up to 20%. Two modulation genes were responsive to the type of situation organisms were facing. In line with the literature reviewed above, one gene (termed risk sensitive one (RS1) was coding for whether the decision was in the domain of gains. When the decision to confront led to a positive payoff the gene RS1 increased the rate of cooperation. In agreement with the literature showing an increase in risk taking in mixed gambles a gene termed risk sensitive two (RS2) was increasing confrontation rates when the decision to confront led to a negative payoff. The two other context-dependent genes were responsive to the health status of the organism. One gene, termed health sensitive one (HS1), was increasing confrontation rates when the potential gain would reach maximum health. The second health-sensitive gene (HS2) gene was activated when the potential loss would lead to the death of the organism. HS2 was thus increasing cooperation.
Three criteria were applied to select the parameters we used in our simulations. The first criterion is the exploratory nature of our study. We wanted to estimate the degree of fit between the theoretical predictions of evolution and the implementation of our paradigm. To this end we decided to cover as wide a range of environmental conditions as possible, even if these are not likely to happen in nature. The second criterion is the limit in computational power. Our choice for the rate of random mutations is much higher than the rate established for human genes. Implementing a rate that is similar to mutation rates in real genes would have the effect of increasing the demand in computational resources without changing the results in the long run. Even if our working hypothesis deserves empirical testing, we considered that setting a high random rate of mutation is a reasonable choice for an exploratory study. Third, in some cases the choice was arbitrary due to the lack of evidence of a well-established value. It is not possible to establish in the human population the proportion of individuals that contribute most to the gene pool of the next generation. Our choice of 10% of the population constituting the elite reflects the fact that, in most species close to humans, genes are passed by a restricted sample of the population at each generation. Hence, even if the exact proportion of what constitutes the elite is arbitrary to some degree it does implement a natural process. The same logic applies to the choice of five genes for implementing the individuals’ propensity to use confrontation. It is clear that more than five genes enter the equation of determining the will to use aggression. It is also clear that an individual’s propensity to use aggression is also largely determined by life experience. By using five genes we implemented the fact that multiple genes are involved and provided a ground to explore the influence of high variance on aggression while limiting the computational demand for resources.
At initialization the program generates the 1000 organisms and their genotypes. At this initial state the allele distribution of each gene would allow the population to have an equiprobable distribution in each trait.
2.3 The Survival Cycle
The survival cycle is constituted by all the events and processes that occur between two pairings of the population, including the changes in health and generation of new organisms. It was implemented in five steps.
In step 1 individuals were paired randomly thus creating 500 situations. The following three parameters were calculated as a function of the level of reward and punishment.
-
The payoff of the decision to confront.
-
The gain associated with the decision to cooperate.
-
The cost associated with a fight.
In step 2 the decision was made individually by each organism as whether to confront or cooperate. First, the probability to confront of the organism was computed on the basis of the five genes CP, RS1, RS2, HS1, and HS2 with δ the probability of confronting, or confrontation rate, being determined as showed in Eq. 1.
Each gene that is sensitive to a condition was activated if relevant and consequently modified the probability of being confrontational. That is,
-
if payoff(confront) > 0 then RS1 was activated.
-
if payoff(confront) < 0 then RS2 was activated.
-
if health + gain > 100 then HS1 was activated.
-
if health – cost < 0 then HS2 was activated.
For each individual, the decision was made by comparing the value of a random variable X [0,1] with equiprobable distribution, to their genetically determined value of δ. When X < δ, the individual cooperates; and when X > δ, the individual confronts the opponent.
The third step was the encounter, where the paired organisms confront or cooperate, and the reward and punishment are allocated to their health points.
The fourth step consisted of determining which individuals have survived the cycle. Any individual that would have a health Γ ≤ 0 was dead and any individual with Γ > 0 was alive for the next cycle. Individuals surviving one cycle were rewarded by one age point. The 10% of survivors with the highest health points were considered the elite who provide the genetic source for reproduction.
The fifth step consisted of generating organisms to bring the ecosystem back to its original capacity. As above, 90% of the new organisms were generated from the individuals with the highest health status. The genotype of the new individual was determined as follows: the genotype from one random individual of the elite was selected and copied. Each of the 9 genes of the parent would then be submitted to a 1% risk of being the target of a mutation. The genetic profile of the remaining 10% of the new individuals was random.
3 Results
This section reports the results of the 10,000 simulations. After evolving for 500 generations, the genes coding for confrontation rates expressed phenotypic effects that varied greatly as a function of the environmental conditions. The section is organized around each of the traits.
3.1 Genes Determining Confrontational Propensity (CP)
The mean confrontational propensity expressed by the five genes sensitive to reward and punishment is presented in Fig. 1. Visual inspection suggests that the genes were highly sensitive to the magnitude of punishment.

Mean confrontational propensity per condition of reward and punishment.
A linear regression accounts for the relationship between reward and punishment on the one hand and confrontational propensity on the other: The multiple regression provides the following Eq. 2:
The model is significant and accounts for 70% of variance in the gene phenotypic effect, r = .87, F(2,9997) = 11736.908, p < .001. Figure 2 shows the degree of fit between model CP and the mean value of CP.

Relationship between the values predicted by Model CP and mean values of the phenotypic effect of CP genes.
3.2 Gene Sensitive to Gain (RS1)
Gene RS1 was activated when the sum of the payoffs of the decision to confront is superior to zero. Figure 3 shows the phenotypic effects of gene RS1 as a function of the payoff.
A regression on the positive payoff shows that the evolution of the phenotypic effects of gene RS1 varies as a function of the payoff making the organisms less confrontational when a sure reward can be secured. The model, reported in Eq. 3, accounts for 31% of the variance, r = .554, F(1,8348) = 3704.593, p < .001.

Mean phenotypic effect of gene RS1 as a function of the anticipated payoff for a confrontation.
3.3 Gene Sensitive to Loss (RS2)
Gene RS2 could upregulate the confrontation rate by up to 20% in response to negative payoffs. Figure 4 shows the relationship between the payoff of deciding to confront and the mean phenotypic effect of gene RS2. The data have been fit with a linear model, see Eq. 4, yielding a significant relationship that accounts for 17% of the variance, r = .419, F(1,1615) = 342.985, p < .001.

Mean phenotypic effect of gene RS2 as a function of the anticipated payoff for a confrontation.
3.4 Gene Responsive to Health (HS1)
Gene HS1 was responsive to situations where the gain in health from a confrontation is less than what the organism has to gain to reach maximum health. As the 10000 simulations yielded different values on the two markers of health (i.e., number of survivors, and average health of the surviving population) we proceeded by binning the results per 10 percentile and calculated the regression model on the mean value per bin. The resulting model, reported in Eq. 5, is highly significant and explains 88% of variance, r = .94, F(2,97) = 367.168, p < .001 and is clearly indicative of a linear trend, see Fig. 5.

Plot of the multilinear model linking the number of survivors at generation 500 and the health of the population to the mean phenotypic effect of gene HS1.
3.5 Gene Responsive to Death (HS2)
Gene HS2 was responsive to the opposite situation and downregulated confrontational levels when the organism was facing death. The model was calculated following the same procedure used for HS1 (see Eq. 6). It yielded a significant, but marginal, effect explaining 13% of variance, r = .386, F(2,97) = 8.48, p < .001, see Fig. 6.
Further investigation indicates that gene HS2 was highly sensitive to loss but could lead to either an increase or decrease in the confrontational rate.

Plot of the multilinear model linking the number of survivors and the health of the population to the mean phenotypic effect of gene HS2.
Ad hoc analysis indicates that the evolution of gene HS2 had high sensitivity to high penalty conditions, but the response of the gene (see Fig. 7) suggests a bifurcation in its evolution.

Mean value of HS2 per condition of reward and penalty.
4 Discussion
The simulations reported above have yielded a number of important findings. First is the fact that a combination of genes sensitive to reward and punishment are determining the confrontation rate to adapt the organisms to different environments. Our results indicate a linear relationship between loss (reward - punishment) and confrontation rate. That the environmental conditions and the phenotypic effect of the CP genes are related by a linear relationship reflects the simplified environment that is used to run the simulations. Modelling of more genes and implementing the interaction between genes might bring to light more complex relationships. What our results nevertheless show is that organisms with no ability to project themselves into the future, such as with working memory, develop an adaptive level of confrontation. In doing so, our results strengthen the huge amount of research that indicates that aggression and the rate of confrontation (and by extension of cooperation) evolved well before humans appeared. The confrontation rate, like any other trait submitted to evolutionary pressures will mechanically be modified over generations if the conditions of reward and punishment change, and it can be an automated reaction that does not require conscious calculation.
Second our results corroborate the idea that confrontation derives from multiple factors that are acting at different levels. Some of the genes were directly responsive to the payoff associated with confronting but others were related to the health status of the organism. The genes modelled here constitute a simplification of reality but show that even in this simplified environment organisms develop a subtle response to variations in their competitive conditions. Our results argue against explanations based on solely social factors which have been put forth in social science and argue for a gene-environment interaction. It is the triggering of genes in specific conditions that might promote ancestrally acquired behaviors. Our results indicate that the willingness to confront stems from complex dynamics that involve the genetic background and the environment modulating the level of risk that people are willing to take. Our results suggest that individual differences in confrontation are likely to be determined by patterns of genes that have coevolved. Some of these genes are not directly involved in evaluating the outcome of the confrontation. Confrontation is thus not necessarily a primal (aggressive) response but is generated through complex dynamics.
The evolutionary simulations we conducted have two limits that are worth bearing in mind when interpreting the results. First, we note that in many instances where the reward is high the organisms did not depart significantly from the initial rate of confrontation. The low selection pressure would partly explain this result and calls for more research into the topic. In natural conditions if rewards are high and punishment is low organisms will increase in numbers up to the point where they have to compete for resources. These dynamics have not been implemented in the present version and should constitute the focus of future research. Second, an important point that stands at the crossroads between the limits of the work and its novelty is the fact that the organisms simulated were not conscious. There was no planning ahead (e.g., avoiding potential confrontations) and the organisms were only reacting to situations. How consciousness influences decisions to confront was beyond the scope of this work but represent the next step in understanding the conditions that promote confrontation or cooperation. Future research should implement a rudimentary form of working memory and model the ability of organisms to make decisions based on an understanding of their future.
In conclusion, our study found that it is possible to simulate the impact of genes on the decision to confront or cooperate. Future iterations of this work may shed light on our understanding of how resource competition can lead to conflict and the potential dynamics of those conflicts.
References
Darwin, C.: On the origin of species. Penguin (1859)
Rand, D.G., Nowak, M.A.: Human cooperation. Trends Cogn. Sci. 17, 413–425 (2013)
Trivers, R.L.: The evolution of reciprocal altruism. Q. Rev. Biol. 46, 35–57 (1971)
Clutton-Brock, T.: Cooperation between non-kin in animal societies. Nature 462, 51–57 (2009)
Dunbar, R.I.: The social brain hypothesis. Evolutionary Anthropology: Issues, News, and Reviews 6, 178–190 (1998)
Armit, I., Knüsel, C., Robb, J., Schulting, R.: Warfare and violence in prehistoric Europe: an introduction. War and sacrifice, pp. 1–11. Brill (2007)
Churchill, S.E., Franciscus, R.G., McKean-Peraza, H.A., Daniel, J.A., Warren, B.R.: Shanidar 3 Neandertal rib puncture wound and paleolithic weaponry. J. Hum. Evol. 57, 163–178 (2009)
Walker, P.L.: Cranial injuries as evidence of violence in prehistoric southern California. Am. J. Phys. Anthropol. 80, 313–323 (1989)
Thucydides: History of the peloponesian war. (1972)
Jetten, J., Mols, F., Selvanathan, H.P.: How economic inequality fuels the rise and persistence of the Yellow Vest movement. International Review of Social Psychology 33, 1–12 (2020)
Smith, J.M.: Evolution and the Theory of Games. Cambridge University Press (1982)
Axelrod, R.: Effective choice in the prisoner’s dilemma. J. Conflict Resolut. 24, 3–25 (1980)
Axelrod, R., Hamilton, W.D.: The evolution of cooperation. Science 211, 1390–1396 (1981)
Nowak, M.A.: Five rules for the evolution of cooperation. Science 314, 1560–1563 (2006)
Friedman, D.: Evolutionary games in economics. Econometrica: Journal of the Econometric Society 637–666 (1991)
Choi, J.-K., Bowles, S.: The coevolution of parochial altruism and war. Science 318, 636–640 (2007)
Pavlov, K.A., Chistiakov, D.A., Chekhonin, V.P.: Genetic determinants of aggression and impulsivity in humans. J. Appl. Genet. 53, 61–82 (2012)
Allen, J.J., Anderson, C.A., Bushman, B.J.: The general aggression model. Curr. Opin. Psychol. 19, 75–80 (2018)
Kahneman, D., Tversky, A.: Prospect theory: An analysis of decision under risk. Handbook of the fundamentals of financial decision making: Part I, pp. 99–127. World Scientific (2013)
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this paper

Cite this paper
Chassy, P., Cole, J., Brennan, C. (2022). An Evolutionary Game Theory Model of the Decision to Confront. In: Bramer, M., Stahl, F. (eds) Artificial Intelligence XXXIX. SGAI-AI 2022. Lecture Notes in Computer Science(), vol 13652. Springer, Cham. https://doi.org/10.1007/978-3-031-21441-7_12
Download citation
DOI: https://doi.org/10.1007/978-3-031-21441-7_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-21440-0
Online ISBN: 978-3-031-21441-7
eBook Packages: Computer ScienceComputer Science (R0)