
Abstract
Biological materials such as skins, bones, teeth or seashells boast remarkable structures and mechanisms, many of them unmatched by engineering materials. In these materials, fracture toughness is key to achieve high strength, reliability, robustness, damage tolerance and notch performance, and to fulfil critical structural functions in the organism. In this chapter, we review and discuss some of the main strategies found in biological materials to resist the propagation of cracks and to reach high toughness. We discuss six major groups of natural materials through specific examples: a uniaxial fiber composite (tendon), a laminated composite (fish scales), a natural elastomer (skin), a mineralized brick and mortar composite (nacre), three-dimensional mineralized cross plies (conch shells, tooth enamel) and a complex hierarchical material (bone). The composition, architecture, mechanics of deformation and fracture, and overall performance is reviewed for each of these materials. The chapter concludes with a summary of the broad strategies deployed in biological materials to manage damage and prevent crack propagation. These lessons are now inspiring the next generation of structural materials.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

1 Introduction
Improving the performance of materials is critical to technological advances as demonstrated through history (bronze age, iron age...) and by modern technology (aerospace, biotechnology, computing) (Ashby 2010). In particular, improving the strength and toughness of materials by manipulating their composition and structure of structural materials has been a focus for thousands of years. As early as the 3\({\text {rd}}\) millennium BC, builders in the Indus valley incorporated straw into mud bricks as fiber reinforcements to increase their fracture toughness, strength and reliability (Lemmen and Khan 2012; Binici et al. 2007). Modern material engineering deals with critical issues related to resistance to fatigue cracks, stable crack propagation, toughening mechanisms, damage tolerance, notch performance and reliability (Ashby 2010; Lawn 1993; Anderson 2005). When exploited and optimized systematically, these concepts have enabled strong, tough and light materials for aerospace, transportation, construction, and energy applications. Despite these impressive advances, nature is well ahead of engineers in making materials (Fratzl and Weinkamer 2007; Barthelat 2015). The materials of trees, skeletons, teeth or protective shells are subject to stringent mechanical requirements. Just like many of our engineering materials, they must fulfil a variety of functions that include structural support, transfer of static, dynamic or cyclic forces, and protection against impact. Nature mainly uses four elements (C, H, O, N), a few building blocks (amino acids, polysaccharides, biominerals), and does not use metals for structural functions except for a few rare exceptions (Lichtenegger et al. 2003). In addition, natural materials are generally processed and fabricated in ambient, “normal” conditions, a severe limitation which contrasts with the extreme temperatures, pressures and chemistries that are common in metallurgy, and ceramic or polymer making. In terms of absolute strength and toughness, biological materials are inferior to our modern steels, largely because of constraints in raw materials. When these limitations in raw materials and processing conditions are taken into account, it is clear that nature is way ahead of engineers and materials scientists in terms of making “more with less”.
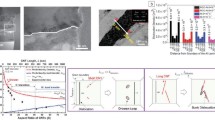
Figure 1 shows an Ashby material property map of toughness versus stiffness for a variety of biological materials. The five biological materials that will be discussed in more details in this chapter are highlighted in red on the chart (a sixth material—tendon—is so tough that no data on fracture toughness is available). Biological materials span 4–5 orders of magnitudes in terms of toughness and stiffness, despite of the narrow range of composition range described above. Soft proteinaceous materials such as skin are soft but extremely tough, while at the other extreme biominerals (calcite, calcium phosphate) are very stiff but also very brittle. Between these two extremes one finds a large number of biological materials with intermediate stiffness and toughness, and interestingly most of these materials display high combinations of these properties. For example, bone, teeth (dentine, enamel) and mollusc shell incorporate proteins and minerals and achieve simultaneous toughness and stiffness. The chart also highlights that nature “amplifies” the properties of raw materials to levels which are not seen yet in engineering materials. For example, nacre from mollusk shell is made of 95% vol. of aragonite (a brittle mineral which is similar to calcite), but its architecture is so well adapted to resist fracture that it is three orders of magnitude tougher (in energy terms).

Material property chart for modulus-toughness in biological materials (adapted from (?)). Guidelines show the best materials to resist large displacement, static force and impact. The specific materials discussed in this chapter (skin, fish scales, bone and nacre) are highlighted
For comparison the properties of a steel alloy (4340 steel: E \(\sim \) 200 GPa, \(J_c\) \(\sim \) 10 kJ/m\(^2\)) are also shown on the chart. The strongest and toughness of biological material cannot match our engineering materials, largely because of constraints in raw materials and processing.However, the “amplification” of toughness and stiffness that hard biological achieve from their fragile ingredients (biominerals, proteins) is not matched to this day by any engineering materials. Nature is developing these original solutions through natural selection and evolution, and while whether the structure of these materials is “optimum” is debatable, there is no question that their properties—including fracture toughness—are remarkably well adapted to their function. The toughness \(J_C\) is the energy required to generate new fracture areas in the material. Alternately, \(J_C\) can be interpreted as the amount of mechanical energy the material can absorb without fracturing, and therefore \(J_C\) can be used as a measure of the impact resistance of the material. By this measure, skin, fish scales, and bone antlers are the most adapted biological materials, which correlates with the function of these materials to resist impact loading without fracturing. In terms of resisting static forces and static stresses, the critical stress-intensity factor provides a better measure of resistance to crack propagation. This criterion maximizes \(\displaystyle (EJ_C)^{1/2}\) on the chart, and nacre, conch shell, and bone are the materials which are the most adapted to resist static forces without fracturing. Finally, we examine the materials which can withstand large deformations without fracture, in other words those that maximize \(\displaystyle \sigma _s/E\), where \(\sigma _s\) is strength. These materials therefore maximize \(\displaystyle (J_C/E)^{1/2}\), and in this case natural elastomers such as skin are clearly the best biological materials to resist large deformations without fracturing. While considering structure-property relationships in biological materials, it is important to also consider function, since the structure and properties of biological materials have evolved for specific functions.
The mechanical properties of biological materials have been of sustained interest for a long time, mainly for construction materials but also for biomechanics. For example the mechanical properties of wood have been of interest for a long time because wood is a construction material (Forest Products 1974). The mechanical properties of bone, cartilage, tendon, skin, and other connective tissues in the human body have also been of interest for biomedical applications, for example to guide optimum therapies, surgical treatments or rehabilitation strategies in orthopedics (Fung 2004).
Over last three decades there has been a renewed and vigorous effort towards understanding the mechanics of biological materials using the modern tools of materials science and mechanics. These powerful approaches let us probe the structure and mechanics of biological materials down to the nanoscale (Rabiei et al. 2010) (Gupta et al. 2005). Powerful computer models can now predict how a slight genetic mutation of the collagen structure at the molecular scale translates into crippling bone diseases (Gautieri et al. 2009). This “biological material science” (Meyers et al. 2008) is now largely motivated by bioinspiration and biomimetics (Barthelat 2007; Vincent et al. 2006). This transfer of technology from biology to engineering requires a precise understanding of structure-properties relationships and property-relationships in natural materials (Meyers et al. 2008), and this effort also includes fracture mechanics: Nature has evolved interesting architectures and mechanisms to prevent and control the propagation of cracks, and duplicating some of these ideas is a powerful approach that could overcome the inherent brittleness of glasses and ceramics (Wegst et al. 2015; Barthelat 2007; Espinosa et al. 2009), thereby expanding their range of applications.
2 Some General Construction Rules for Biological Materials
The range of compositions in biological materials is remarkably narrow considering the breadth of mechanical properties that can be achieved. In contrast with engineering materials, which make full use of most of the elements in the periodic tables, only four are prominent in nature: carbon, hydrogen, oxygen and nitrogen (C, H, O, N) account for 95% of the mass of biological organisms on earth. Proteins and polysaccharides are the “building blocks” of animals and plants. For example, proteins represent about 17% wt. of the human body, where they perform a wide variety of functions. The molecular building blocks for proteins are amino acids, small organic compounds with an amino group, a carboxyl (acid) group, and a radical group (R-group). The R-group varies in composition and size depending on the amino acid, and only 21 amino acids are found in nature. Sequences of amino acids are assembled by condensation, which can be interpreted as polymerization where the amino acids form long polymeric chains with a strong covalent backbone. The process of transcription of DNA into RNA, and translation into a protein chain, is the fundamental fabrication mechanisms for proteins. This process is precise and governs the exact sequence of amino acids along the protein chain (also called primary structure) with high fidelity. Once the primary structure is formed, side groups along the proteins promote the formation of hydrogen bonds which can induce the folding of the protein chain into specific configurations. The exact sequence and size of the side groups will produce repeatable and robust folding patterns which lead to complex three-dimensional configurations called secondary structures. Common secondary structures in structural proteins are coiling (alpha helix) and crystallization (beta sheets). In terms of structural behavior, proteins can be interpreted as a precise coiling of a strong covalent backbone which is stabilized by weaker hydrogen bonds. Under the action of a mechanical pull, individual proteins can unfold as the hydrogen bonds are broken, which produces large elongations. Structural proteins such as keratin, elastin, and collagen are critical for the stiffness, deformability, strength and toughness of skin, nails, tendons, and bones. To illustrate the construction of a structural protein, we take the example of collagen, which represents about 25% wt. of all proteins in the human body. Collagen is critical for the mechanical performance of bones, tendons, skin, and eye cornea. Collagen is therefore a “Universal protein” found across various structural tissues in the human body, and also across the animal kingdom (all mammals, fish scales, anemones, sea cucumbers). Collagen is made of up to 20 different amino acids, the exact composition varying across about 20 types. The main collagen types are type I (in most human tissues), type II (cartilage), type III (in blood vessels and repair sites). Here we focus the discussion on collagen type I. Proteinaceous collagen chains assemble into tropocollagen, which consists of three left-handed alpha chains twisted in a right-handed triple helix stabilized by hydrogen bonds. Alternating between a left and right handed assembly is critical for the stability of the molecule, just like alternating the twisting direction in multi-stranded ropes is important to prevent the unraveling of the strands. Every third R-group along these chains is glycine, the smallest residue in amino acids. This residue faces the inner side of the coil, making the tropocollagen helix very tight. Individual tropocollagen molecules are 300 nm long for a diameter of about 1.5 nm. They have a head and a tail with distinct functionalities, and they tend to assemble so “head” bonds assemble with a “tail”. This feature leads to the self-assemblies of tropocollagen molecules into three-dimensional periodic bundles called fibrils (Fratzl et al. 1998), which contain a periodic gaps and a 67 nm periodicity along the fibril. The resulting fibril is 20–200 nm thick, and can reach tens of millimeters in length. This fiber is large enough to be mechanically isolated, handled and tested in tension, using MEMS based actuators (Eppell et al. 2006) or capacitor-based devices (Poissant and Barthelat 2012). These tests reveal a relatively stiff fiber (\(E= 1\,\text {GPa}\)), which is strong (strength >200 MPa) yet deformable (strain at failure >10 %). As depicted on Fig. 2, this formidable fiber serves as a basis to form random networks (skin), uniaxial composites (tendons, ligaments) or crossplies (fish scales). To add stiffness, these proteins may be mineralized, as seen in mineralized tendons (low mineralization), bone, nacre, and enamel (high mineralization). At the molecular scale, individual collagen molecules (tropocollagen) bond via coordinated hydrogen bonds (Buehler 2006) and self-assemble into fibrils (Fig. 3a). Specific covalent crosslinks at the ends of the collagen molecules (telopeptide regions) provide cohesion and mechanical stability to the fibrils, and govern complex unraveling nano-mechanisms as the fibril is stretched (Uzel and Buehler 2011). While cellulose is the main structural protein for many animals (mammals, reptile, fish), polysaccharides are the main construction materials for insects (chitin) and plants (cellulose). These fiber-like materials follow the same broad construction rules of collagen: strong molecular backbones, weaker intermolecular interaction, self-assembly into fibrils and fibers.

Overview of the construction of collagenous tissues, which is typical of biological materials: a cells produce a sequence of amino acid that reflects DNA’s blueprint; b these primary structures assemble into more complex, 3D “secondary” structures: protocollagen molecule, collagen fibrils, collagen fibers; c these fibers are deposited in various ways to form biological tissue with a broad range of structural properties

The structure and mechanics of tendon: a Hierarchical bundles of collagen molecules, fibrils and fibers (Fratzl 2003); b Sinusoidal crimps (Scanning electron micrograph, Herod et al. 2016); c the fibrils in tendon are extremely long and may span the entire length of the tissue (Scanning electron micrograph, (Provenzano and Vanderby 2006); d typical tensile stress-stain curve of tendon (Haraldsson et al. 2005); e notch performance of tendon, adapted from Ker (2007) (tendon is so tough that crack propagation through its section is not possible); f schematic showing the mechanisms of crack blunting and delamination
3 A Uniaxial Fiber Composite: Tendon
The simplest way to arrange fibers and to make the most of their high performance in tension is to arrange them in parallel bundles, as in tendons. Tendons are highly specialized tissues whose function is to carry and transfer tensile forces between muscle and bone. Tendons must therefore be stiff in order to accurately and rapidly convert muscle action into skeletal motion, and strong in order to carry high tensile forces without failing. Tendons are also used as mechanical energy storage that, for example, improves the efficiency of running. They are made of collagen type I (60–90% dry weight), elastin (5%) and other extra-collagenous proteins (Shen et al. 2008). Collagen fibrils bundle into parallel fibers or fascicles about 50–300 \(\upmu \) in diameter (Fig. 3a). The fibrils are held by a much softer extra-collagenous matrix, whose shear modulus is three orders of magnitude lower than the modulus of the fibers (Ker 2007). At rest, the fibers are not completely straight and display wavy patterns called crimps (Fig. 3b). Mechanical models and in-situ X-ray images suggest that the collagen fibrils have a finite length, which enables them to glide on one another when tensile forces are applied on the tendon (Puxkandl et al. 2002). However, direct observations on tendons show individual fibrils with no apparent ends, which suggests that fibers in fact span the entire length of the tendon (Provenzano and Vanderby 2006; Svensson et al. 2017) (Fig. 3a).
Figure 3d shows the tensile response of tendon (tensile stress-strain curve Haraldsson et al. 2005). The “toe” and “heel” region of low stress corresponds to an initial regime where the crimps in the collagen fibers are straightened. This process requires little tensile force, but once the crimps are straightened the material is much stiffer. In the linear region, the fibers are straight and aligned along the direction of pulling, and the strong and stiff covalent backbones of the collagen molecules carry an increasing amount of tensile stress. He linear region is therefore relatively stiff (\(E= \)1–2 GPa). At tensile stress of about 50–100 MPa the material reaches its maximum carrying capability and strength. The tendon softens because of defibrillation and progressive rupture of the fibers (Fung et al. 2009). In uniaxial tension, tendons are among the stiffest and strongest non-mineralized biological materials. Figure 3e shows a tendon containing a deep cut and subjected to tension (Ker 2007). The initial deformability of tendon blunts the tip of the cut, turning the sharp slit into a rounded notch with the direct effect of reducing the stress concentration. In addition, since the shear modulus of the fascicle interfaces is much lower that the fibers, large shear deformations take place along these interfaces. The interfaces then fail in shear and channel “delamination” mode II cracks along the direction of pulling. This mechanism blunts the crack further into a Cook-Gordon type configuration (Cook et al. 1964), further preventing propagation into the fibers. The delamination of the fibers has also two effects: (i) behind the crack tip a large volume of material becomes stress free (Fig. 3f); (ii) In front of the crack tip, the tensile stresses become uniformly distributed. In the laboratory test, the mode II crack reach the grips of the loading machine before the ligament fails in tension (Ker 2007). This powerful mechanism therefore turns tendon into a notch insensitive material. The cut decreases the strength because of the reduction in nominal cross section, but the stress concentration at the tip of the cut is completely suppressed. Instead, the delaminating crack proceeds into the grips of the machine. In this configuration it is therefore impossible to propagate a crack in model I across the fibers, and therefore estimates for the fracture toughness of tendon are not available (Szczesny et al. 2015). Interestingly, the construction and mechanisms of tendon are identical to the design guidelines for modern fiber reinforced engineering composites: High concentration of stiff and strong fibers in a weaker matrix to provide high combinations of stiffness and strength. The strength of the interface between matrix and fibers is critical: The interfaces must be strong enough to provide cohesion to the material, yet weak enough to interact with propagating cracks in order to trigger powerful toughening mechanisms (Zok 2006) (Cook-Gordon blunting mechanism, crack deflection, crack bridging). The construction of tendon is perfectly adapted to carry large tensile forces, even in the presence of partial tears or local damage. However, it is a highly specialized tissue that can provide stiffness and stress along only one direction (tendons are very weak in the transverse direction). The next two examples discuss collagen fibrils in crossplies (fish scales) or random networks (skin). These different architectures produce different mechanisms and properties, but when stress concentrations are present fibers align locally, generating a “tendon-like” barriers to crack propagation.
4 A Natural Composite Laminate: Fish Scale
Tendons are rope-like tissues specialized in carrying tensile forces along a single direction. For other structural elements where biaxial stiffness and strength are required, crossply architectures are more adapted. In crossplies, the alignment of the fibers is uniform only within one layer (or “ply”) of the material, and adjacent layers have a rotated arrangement. Simple cross plies have a (0–90) degree arrangement, while more complex crossplies such as the Bouligand structure of arthropod cuticles (the hard shell of insects and crustaceans) only vary by a few degrees from one layer to the next (Raabe et al. 2005). The immediate benefit of cross plies is that the tensile strength and stiffness of individual fibers are available along different pulling direction within the plane of the plies. In effect, crossplies increase the isotropy of fibrous material, the main drawback being that along each of these directions the stiffness and the strength is reduced compared to the uniaxial composite (Nikolov et al. 2010). Cross plies are well adapted for hydrostatic skeletons, which are essentially pressurized reservoirs: the body of sea anemones, soft-shelled turtles (Scheyer et al. 2007), and human annulus fibrosus (?). In arthropod shells (cuticles) and fish scale, crossplies are well adapted to resist multiaxial stresses from sharp contacts (impacts, attacks from predators). For example, localized surface forces generate flexural stresses in the shell, which must be resisted along direction in the plane of the shell.

The hierarchical structure of typical teleost fish scales (shown here for a striped bass Zhu et al. 2012)
The structure and mechanics of natural scaled skin, and more particularly fish scales, have recently been the subject of several studies (Yang et al. 2013a, b; Bruet et al. 2008; Ikoma et al. 2003; Garrano et al. 2012; Lin et al. 2011; Meyers et al. 2012; Zhu et al. 2013; Browning et al. 2013; Zimmermann et al. 2013; Vernerey and Barthelat 2010). Figure 4 shows the hierarchical features of fish scales. At the macroscopic level, the scales are staggered and cover most of the body of the fish, providing a continuous barrier from penetration combined with flexural compliance. At the mesoscale level, individual scales are polygonal thin plates composed of type-I collagen fibrils partially mineralized with hydroxyapatite (16–59% mineral content in weight (Ikoma et al. 2003; Liu et al. 2008; Torres et al. 2008; Schonborner et al. 1979; Seshaiya et al. 1963). The outer layer of the scale is significantly more mineralized and often referred to as the “bony layer”. Bony and collagen layers have approximately the same thickness (\( {\sim } 100 \upmu \text {m} \)). These layers are both cross-ply layered composites, each ply being made of parallel collagen fibrils rotated across layers by angles that can vary across species (Bigi et al. 2001; Meunier 1984; Zylberberg et al. 1988; Meunier and Castanet 1982; Meunier 1981). The scales of striped bass, consist of a basal layer formed of 20–25 plies about 4–5 \(\upmu \)m thick each (Fig. 4d), where the collagen fibrils are rotated by 90 \(^\circ \)C from one ply to the next (Fig. 4e, f). Natural scaled skins have remarkable mechanical properties: compliance, resistance to penetration, lightweight, and ultra-thin structure (Yang et al. 2013a). Tensile tests on natural teleost fish scales confirmed the scale as a stiff, strong, and tough material with extensive inelastic deformation and energy dissipation including pullout, defibrillation, sliding and ply rotation (Ikoma et al. 2003; Garrano et al. 2012; Lin et al. 2011; Zhu et al. 2012; Zimmermann et al. 2013). Fracture toughness is a critical property to resist puncture or lacerations from predators or collisions with other fish or obstacles. Early tests revealed that fish scales are so tough that they could not be fractured, even after immersion in liquid nitrogen (Currey 1999). Figure 5a,b show a set of tensile test results on plain and notched scales. The curves have a bell shape, with an initial linear response followed by large tensile strains and tremendous energy absorption. As expected, the notched samples were weaker than the intact samples because of the presence of the notch and the associated reduction in the nominal cross section (i.e. minimum load bearing cross section). However, when the nominal stresses are calculated, the stress strain curves for the notched and intact scales are nearly identical. Individual fish scales from Morone saxatilis are therefore notch insensitive. At the early stage of loading, the bony and collagen layer delaminated because of the mismatch between their mechanical properties (Zhu et al. 2012) (Fig. 5b). Further increase of loading resulted in the fracture of the bony layer while the collagen layer was still deforming, with extensive defibrillation of the collagen cross plies up to the ultimate failure at around 0.8 mm displacement. The notched sample also showed crack blunting (Fig. 5b, 3c), a potent toughening mechanism for metals and polymeric materials. Instrumented fracture tests on individual fish scales are extremely difficult because of their small size and very high toughness. Recently a new miniature setup was used to measure the toughness of the scales along three crack propagation orientations (Dastjerdi and Barthelat 2015). Crack propagation was always stable, and the results confirmed that fish scales are among the toughest biological materials (work of fracture \({\sim }~40 \text {J}/\text {m}^2\), Fig. 1). Fracture models suggest that inelastic deformations of the collagen fibrils, which operate over regions on the order of 1–2 mm around the crack tip is the main contributor to toughness (Dastjerdi and Barthelat 2015), a process similar to nacre (Barthelat and Rabiei 2011) and to advanced engineering polymers (Evans et al. 1986). The fibers also rotate towards the direction of pulling (Yang et al. 2014), providing a local “tendon” like structure to resist crack propagation. Delamination of the collagen fibers also produces a small bridging stress across the crack faces, but this mechanism can operate over large crack openings so that its contribution to toughness is not negligible (Dastjerdi and Barthelat 2015). For these mechanisms the interfaces between the fibers and the plies must be much weaker than the fibers. Delamination experiments indicated that these interface are indeed 400 times weaker than fish scale as a whole (Dastjerdi and Barthelat 2015).

Mechanical tests on individual scales: a tensile tests on intact and notched samples with b resulting nominal stress-displacement curves; c Fracture involves extensive delamination and bridging by collagen fibers (adapted from Dastjerdi and Barthelat 2015)

Overview of skin structure and mechanics: a Schematic of the network of collagen fibrils in the dermis; b tensile stress strain curve of human skin (Silver et al. 2003); c Notch performance of skin in tension. Rounding of the crack tip suppresses stress concentrations (Yang et al. 2015); d at the crack tip the collagen fibrils align along the direction of loading, forming a “tendon-like” barrier to crack propagation (Yang et al. 2015)
5 A Rubber-Like Material: Skin
Human skin is the largest organ in the body. A thin, deformable and tough layer, skin fulfills a wide range of critical functions that include protection against mechanical threats, pathogens or water loss, but also temperature regulation, sensing and excretion (sweat) (Fung 2004). Skin is composed of several layers: stratum corneum (outermost layer), epidermis, dermis, subcutaneous tissue (innermost skin layer) (Silver et al. 2003). The dermis layer is the thickest layer, and it is also the layer that governs the mechanical response of skin (Oxlund et al. 1988). The dermis is a three-dimensional network of cross-linked collagen and elastin fibers embedded in proteoglycans (Silver et al. 2003). The network of collagen fibrils largely dominates the mechanical response of the dermis, elastin providing recoil and “elasticity” to the skin once it is unloaded (Oxlund et al. 1988). Individual collagen fibers within the network are primarily parallel to the surface of skin (Fung 2004) (Fig. 6a). Within the plane of the dermis, the fibers follow a mostly random orientation, although preferred orientations are observed locally along lines which were first mapped by Langer (Ridge and Wright 1966a). Skin tends to be stiffer and stronger along these Langer lines (Ridge and Wright 1966a, b), and wounds or cuts that disrupt these lines can take longer to heal (surgeons favor incision parallel to these lines). The dermis shares many attributes with rubbers and other engineering elastomers in terms of structure, mechanics and properties (including fracture properties). Both are made of a random network of cross-linked fibers or molecules, and the elasticity of both can be captured with entropy-based elastic models (Fung 2004; Bischoff et al. 2000). Both materials are very soft and can undergo large deformations in tension. The high level of hydration of skin maintains the high mobility of the elastin and collagen components. Figure 6b shows a typical stress-strain curve of skin in tension. Up to about 30% strain, the collagen network offers only a small resistance to deformation as entropic elastic dominates. At larger strains skin enter a much stiffer, linear region where the collagen network progressively aligns and stiffens along stretch direction. Eventually the cross links break, fibers slip, and the overall collagenous network weakens which generate a “yield point” on the curve. Complete defibrillation follows with progressive tear of the skin. Skin provides a mechanical barrier to bites, scratches and small tears, so that fracture toughness is critical. Tear fracture tests reveal toughness in the order of \(10~\text {kJ/m}^2\) (Purslow 1983), which is towards the high end for biological materials (Fig. 1). Figure 6c shows a strip of skin containing a through cut in tension. Skin is easily deformed at low stresses, so that the initial slit immediately turns into a rounded ellipse as a tensile force is applied, which suppresses the stress singularity at the tip of the notch. In addition, the alignment of the collagen fibers in these deformed regions form a region of aligned collagen fibers which are transverse to the direction of crack propagation (Fig. 6d). Skin therefore “recruits” random collagen fibers with high deformations, to toughen the material locally with fiber reinforcements in a “tendon-like” material.
6 A Densely Mineralized Brick and Mortar Composite: Nacre
Minerals are widely used by living organisms for structural purposes, mainly for increasing the stiffness and hardness of otherwise relatively soft proteins. The tensile modulus of collagen, the most common structural protein, in a unidirectional material such as tendon is 1–2 GPa. This is relatively stiff for a protein, but too soft to fulfill skeletal functions or heavier protection. Structural biological materials therefore often incorporate stiff minerals to increase stiffness and hardness. Calcium carbonate (mollusk shells) or hydroxyapatite (human bone, teeth) are the most common minerals found in natural materials, although there are many others (Weiner and Addadi 1997). The general strategy for incorporating these minerals is for the organism to grow a scaffold of proteinaceous materials first, which is then mineralized over time. This process is highly regulated by complex biochemical processes combined with physical confinement to control the deposition sites, shape, size, and growth rate of mineral crystals (Weiner and Addadi 1997; Mann 2000). As a general rule, the minerals are in the form of nanograins (Rousseau et al. 2005), or in the form of mesocrystals (Colfen and Antonietti 2005): large crystals made of nanograins with uniform texture and bonded by organic materials. Therefore all mineralized tissues in nature are composite materials, even seemingly pure biominerals such as urchin spines (Seto et al. 2012). In general, stiff materials are also hard (Ashby 2010), and therefore incorporating minerals into soft matrices increases both stiffness and hardness. However, the deformability of the material also decreases and their fracture toughness also decreases in general, so that this “conflict” between strength and toughness is prominent in engineering materials (Ritchie 2011). Interestingly, natural materials alleviate this limitation by remarkable architectures and mechanisms which ensure high stiffness, hardness and fracture toughness.

Overview of the structure and mechanics of nacre: a Nacre has a three dimensional brick and mortar structure and is part of a two layer hard shell in mollusks (Barthelat et al. 2007); b schematic with dimensions. 20–30 nm thick layers of organic materials bond the microscopic tablets together; c when tension is applied along the direction of the tablets they slide on one another over large volumes. This behavior generates large strain, turning a brittle biomineral into a tough composite (Barthelat et al. 2007)

The fracture mechanics of nacre: a Experimental crack resistance curves for four types of nacre, together with in-situ optical images. The whitening regions are an indication of tablet sliding at the microscale (Rabiei et al. 2010); b Schematic showing the main toughening mechanisms (Barthelat and Rabiei 2011)
Mollusk shells provide remarkable examples of highly mineralized natural materials. The shells are mostly made of minerals (at least 95% volume) and contain only a small fraction (at most 5% volume) of organic materials (Currey and Taylor 1974). Among the different microstructures found in mollusk shells, nacre is the strongest and toughest (Currey and Taylor 1974). Nacre displays complex micro-mechanisms of deformation and fracture which generate high stiffness (70–80 GPa), high tensile strength (70–100 MPa) and a remarkably high fracture toughness (4–10 MPa.m1/2) (Currey 1977; Jackson et al. 1988; Wang et al. 2001). Nacre has a relatively simple, brick-wall-like architecture composed of mesocrystals of minerals in the shape of polygonal tablets (0.2–1 \(\upmu \) thick, 5–10 \(\upmu \) in diameter, Fig. 7a, b). In tension, the tablets can slide on one another, which generates relatively large deformations (up to almost 1% strain) accompanied with energy dissipation (Barthelat et al. 2007; Jackson et al. 1988; Wang et al. 2001), Fig. 7c. The sliding and pullout of the tablets are mediated by the thin (20–40 nm) interfaces between the tablets, which are rich in organic materials (Levi-Kalisman et al. 2001). The relatively large inelastic deformations and the energy dissipated at the interface translate into a material that can absorb deformations, deform to reduce the effects of stress concentrations, tolerate damage and absorb mechanical energy from impact. Figure 8a shows crack resistance curves obtained from four-point bending tests on single edge notched bend samples of nacre, prepared so a crack propagates across the tablets. In this configuration crack propagation in nacre is stable, and the curves display a very strong “R-curve” behavior, with a resistance to crack propagation (measured in kJ/m\(^2\)) that is initially low (low initiation toughness) but which increases significantly as the crack advances. This behavior, a characteristic of advanced structural materials, imparts the materials with tolerance to defects and stable crack propagation. This behavior also indicates that powerful toughening mechanisms are triggered by the stresses ahead of the advancing crack. The main two toughening mechanisms are both associated with the sliding of the micro tablets. If the cracks do not penetrate into the tablets, the intrinsic cohesion of nacre is provided by the interfaces. These interfaces are very weak, with a measured mode I fracture toughness of only about 10 J/m\(^{-2}\) (Currey 1977; Mayer 2005; Dastjerdi et al. 2013), which is about 100 times lower than the toughness of nacre across the direction of the tablets (Rabiei et al. 2010). As noted previously for tendon and fish scales, weak interfaces are a requirement for the interfaces to deflect and guide incoming cracks. Mechanical tests on demineralized nacre confirm that the organic materials have low strength but high deformability (Dastjerdi et al. 2013; Lopez et al. 2014). The low toughness of the interfaces is amplified by two micro-mechanisms, both associated with tablet sliding (Fig. 8b). A first amplification of the toughness of the interfaces is provided by crack bridging, with tablets interacting behind the main crack tip. A second amplification of toughness is more powerful and provided by the inelastic process zone that develops in front of the advancing crack. The high tensile stress in this region triggers tablet sliding in a relatively large region, millimeters in size. As the crack propagates into that inelastic region, the material on either side of the newly created crack faces unloads, leaving a wake of permanently deformed material with residual strains. The process of propagating a crack in nacre therefore involves the loading and unloading of a large volume of material, which dissipates a large amount of energy. Process zone toughening (Barthelat and Rabiei 2011) make nacre several orders of magnitude tougher than aragonite (Rabiei et al. 2010; Barthelat 2007; Wang et al. 2001). This critical mechanism is supported by experiments: nacres with small process zones have a low toughness (Fig. 8a). It is also supported by models, that show that transient process toughening can generate extremely large or even unbounded toughening (Barthelat and Rabiei 2011). High extensibility is critical to develop inelastic mechanisms over large volumes and generate toughness at the macroscale (Barthelat et al. 2007). Among other properties for the interfaces in nacre, it has been suggested that extensibility is the most important for the overall toughness of nacre (Nabavi et al. 2014). The properties of the interfaces appear to be fine-tuned to achieve high performance for the material (Nabavi et al. 2014).
7 A Mineralized Cross-Ply: Tooth Enamel
Tooth enamel is a very hard tissue that covers the surface of teeth. Its extremely high level of mineralization (\(\sim \)96% vol.) makes it the hardest tissue in the human body (Nanci and Ten Cate 2013). Surface hardness is a critical property for the functionality of teeth, to ensure efficient crushing and cutting food particles. Biting forces can be relatively large (100–1000 N), which involves very high and localized contact stresses involving teeth and food particles (Yahyazadehfar et al. 2014). Specific architectures and mechanisms are therefore required to resist surface cracking. The microstructure of enamel is made of mineral rods about 5 \(\upmu \) in diameter, which start at the surface of the tooth and end at the junction between enamel and the underlying dentin (the dentine-enamel junction, Fig. 9a). Individual rods are made of nano-crystallites of the mineral hydroxyapatite (Habelitz et al. 2001). The thin interface between the rods is rich in proteins. While the rods are perpendicular to the surface of the tooth near the surface (in the outer enamel region), in deeper regions (inner enamel region) they crisscross complex three-dimensional decussation patterns (Macho et al. 2003). Cracks emanating from the surface of the tooth are channeled away from the surface and along the parallel rods, preventing chipping of the enamel surface. Deeper within enamel, the decussation pattern impedes further crack growth (Yahyazadehfar et al. 2013) by a series of crack deflection and crack bridging (Bajaj et al. 2010) (Fig. 9a, b). The powerful toughening mechanisms at work in the inner region can be quantified by propagating a crack from the outer layer to the inner layer (Bajaj and Arola 2009). In this “forward” direction (Fig. 9c), the toughness remains about constant in the outer layer, and the crack is mostly channeled by the proteinaceous interfaces. When the crack enters the decussated region in inner enamel, the toughness rapidly increases as crack bridging takes place, up to four times the initial fracture toughness. Through-cracks in enamel are therefore very stable and are actually considered a normal aging process for teeth (Lin and Douglas 1994; Bechtle et al. 2010; Espinosa et al. 2011). More severe stresses may propagate the crack through the enamel layer where they meet another line of defense: the dentine-enamel junction (Imbeni et al. 2005) and underlying dentin (Kruzic et al. 2003). In contrast, cracks propagated in the “reverse” direction (from inner to outer enamel) are unstable and propagate more easily (Bajaj and Arola 2009) (Fig. 10c). Similar mechanisms are found in the shell of conch, another example of a highly mineralized biological material (>99% vol.). In the outer layers of the shell, mineral plies are perpendicular to the surface, and proteinaceous interfaces between the plies channel multiple cracks in periodic arrays (Kamat et al. 2000, 2004). In the middle layer, the layers form a \(\pm 45^{\circ }\) crossply, which creates large scale crack bridging and amplifies the toughness by two to three orders of magnitude (Kamat et al. 2000, 2004). In tooth enamel and conch shell the weaker organic interfaces only represent 1–5 wt%, but they are crucial to the toughness of these biological ceramics (Yahyazadehfar and Arola 2015).

Tooth enamel: a microstructure and interaction with a surface crack (Mirkhalaf et al. 2014); b Scanning electron micrograph of a crack propagated form the outer region (straight rods) to the inner region (decussated rods) (Bajaj et al. 2010); c Crack resistance curve for tooth enamel in the “forward” direction shows a pronounced rise in local toughness when the crack enters the decussation region. In the “reverse” direction the toughness is lower and the crack is unstable (Bajaj and Arola 2009)
8 A Complex Hierarchical Composite: Bone
Bone is a high-performance material which fulfills a variety of functions, the primary of which is mechanical support (Currey 2002). Bone must therefore be stiff and hard, but it is also surprisingly tough (Wegst and Ashby 2004) considering its contents of brittle minerals and soft proteins. Bone density and mineral content have traditionally served as the only predictors of bone strength, but these measures have limitations (Hui et al. 1988). More recent research has considered the material bone as a composite material in which minerals, collagen and extracollagenous proteins contribute to its mechanical performance (Burr 2002; Ritchie et al. 2009). Figure 10 shows the structure of cortical bone, which is the dense, outer layer of long bones (femur, tibia). The material bone is composed of approximately 60% weight of mineral (calcium and phosphate), 10–20% water and 20–30% of proteins. About 90% of the protein content is collagen type I, the remaining 10% non-collagenous proteins including fibronectin, osteonectin, sialoprotein, osteocalcin and osteopontin (Young 2003).

The hierarchical building blocks that form the structure of cortical (dense bone). The building blocks are joined by interfaces which are critical to deformation and fracture mechanisms (Barthelat et al. 2016)
The collagen fibrils in bone they are reinforced by nanocrystals of hydroxyapatite (Weiner and Wagner 1998; Hassenkam et al. 2004; Buehler 2007). The fibrils bundle into fibers, which form the building block of bone at the next hierarchical level. Fibers arrange into cross plies and lamellae at the microscopic scale (Fig. 10), and the lamellae wrap around the Haversian canals concentrically to form the osteons, which are the microscopic building blocks of mature cortical bone. The deformation and fracture of bone is complex and involves mechanisms at each of these length scales (Ritchie et al. 2009; Launey et al. 2022; Ural and Vashishth 2014; Ritchie et al. 2005). This description of the structure and mechanics of bone, based on hierarchical building blocks (Weiner and Wagner 1998; Rho et al. 1998), has dominated our conception of this material over the last 20 years. More recently however, the interfaces between these building blocks have been examined in more depth (Fig. 10), especially in the context of deformation and fracture (Barthelat et al. 2016; Buehler 2007; Thurner and Katsamenis 2014; Zimmermann et al. 2011; Taylor et al. 2007; Gupta et al. 2006; Fantner et al. 2005; Dunlop et al. 2011). For example, fibrils are held together by a 1–2 nm thick layer of non-collagenous interfibrillar matrix which contains a variety of proteins including osteocalcin and osteopontin (Ural and Vashishth 2014). This mixture of proteins is more compliant and weaker than the stiff mineralized and aligned collagen fibrils as demonstrated by cleavage and fracture surfaces of lamellar bone at the microscale (Fantner et al. 2005). The proteins at the interfaces are highly deformable, however, and separating the collagen fibrils in bone forms ligaments in the interfaces (Fantner et al. 2005) which are similar to observations on nacre. Figure 11a shows the stress-strain curve of bone. The modulus is in the range of 10–20 GPa and the tensile strength is about 100 MPa, which is about 10 times stiffer and two times stronger than tendon (respectively), a direct effect of the mineralization of bone. Beyond a “yield point” bone displays inelastic deformations and it can fail at strains in the order of 1–2%. The inelastic deformation of bone is critical to its ability to absorb impact energy without fracturing. The multiscale micromechanics governing this deformation includes the inelastic deformation of individual collagen fibrils (Tang et al. 2010), the sliding of minerals relative to collagen (Mercer et al. 2006), the sliding of fibrils on one another (Gupta et al. 2006), the accumulation of diffuse damage (Zioupos 1998), the development of dilatational bands at the nanoscale (Poundarik et al. 2012; Schwiedrzik et al. 2014), and shearing of the cement lines (Ascenzi and Bonucci 1972). While these inelastic deformations develop, bone must resist the propagation of large cracks emanating from bone’s heterogeneities and microcracks (Zioupos 1998). This competition between deformation and fracture is what makes the ductility of bone possible. It requires not only high toughness for the material bone, but also a rising crack resistance curve that would promote crack stability.

Mechanics of deformation and fracture in cortical bone: a Stress strain curves in tension and compression along the axial direction of a bovine femur (Mercer et al. 2006); b crack resistance curve obtained from mode I fracture tests across the longitudinal direction of human humeri (Nalla et al. 2006); c Multiscale toughening mechanisms in cortical bone: Nanoscale “ductility” (Gupta et al. 2006), bridging by collagen fibers, (Fantner et al. 2006) deflection on osteon lamellae (Peterlik et al. 2006), microcracks (Vashishth 2007), deflection on cement lines (Zioupos et al. 2008)
Figure 11b shows a set of experimental crack resistance curves for cortical bone, for a mode I crack propagating across the longitudinal direction of long bones (human humeri Nalla et al. 2006). The overall toughness (\(K_{IC}\)) of bone is on par with fish scale and nacre, and among the toughness natural materials (Fig. 1). Bone is also about 10 times tougher than skin and the pure hydroxyapatite mineral (in \(K_{IC}\) terms). Fracture toughness also increases significantly as the crack advances, an indication of powerful toughening mechanisms taking place. The ductility of the material bone at the tip of cracks generates a process zone toughening mechanism similar to nacre. Microcrack toughening was also suggested as a similar mechanism (Vashishth et al. 2003). Propagating a crack in bone also involves the pullout of individual fibers and fibrils out of the crack faces (Nalla et al. 2003; Fantner et al. 2004), crack bridging and added toughness for bone. Upon opening or shearing of the interface, these electrostatic sacrificial bonds can break and release ‘hidden lengths’ along the molecule, generating large deformations and energy dissipation at the molecular scale (Fantner et al. 2005). Tensile experiments on bovine cortical bone using stepwise changes in strain rates confirmed that the activation enthalpy associated to nonlinear deformation in bone corresponds to the disruption of electrostatic bonds (Gupta et al. 2007). A larger scales, mature cortical bone can therefore be interpreted as a unidirectional fiber reinforced composite, where the osteons are the fibers and the interstitial bone is the matrix (Burr et al. 1988). In a similar way that an interface of carbon or glass fibres in synthetic composites can deflect cracks and generate toughness by pullout, cracks can deflect or twist along the weaker cement lines (Launey et al. 2022; Koester et al. 2008). These powerful mechanisms make cortical bone five times tougher in the transverse direction compared to the longitudinal ‘splitting’ direction (Ritchie et al. 2005). To deflect incoming cracks, the cement line must be significantly weaker than both osteons and interstitial bone. Push-out tests on individual osteons have verified the low shear strength of the cement lines (8 MPa) (Ascenzi and Bonucci 1972; Bigley et al. 2006), about 10 times smaller than the surrounding interlamellar interfaces (73 MPa) (Dong et al. 2005). Once the cement line has broken, frictional pullout ensues (Bigley et al. 2006), a mechanism which is also observed and exploited in synthetic fibers used in engineering composites. The fracture toughness of cement lines can be estimated from the toughness of cortical bone in the splitting direction, because the crack mostly propagates along the cement lines in that orientation. By this measure, the toughness of the cement line is 1–2 MPa\(^{1/2}\), which is 10 times lower than the toughness of bone in the transverse direction (Koester et al. 2008). These experiments confirm the strong contrast of strength between cement lines and the surrounding bone material, which can be explained by differences in composition and structure. The main toughening mechanisms associated with the cement line are crack deflection and twisting (Ritchie et al. 2009; Koester et al. 2008; Ager et al. 2006), although debonding followed by frictional pullout has also been suggested as an important toughening mechanism associated with osteons (Piekarsk 1970; Hiller et al. 2003). Disrupting the finely tuned structure and mechanisms of these interfaces in bone can have a profound impact on overall performance. For example, suppressing key interface proteins such as osteopontin has immediate and dramatic consequences on the overall toughness (Poundarik et al. 2012; Thurner et al. 2010), and recent studies have shown that the decrease of the mechanical properties of bone with age can be explained by excess of stiff and brittle covalent cross links at the nano-interfaces (Zimmermann et al. 2011), with direct effects on the toughening mechanisms (Fig. 11b). These results make it clear that bone must be understood as an integration of structural building blocks connected by interfaces.
9 Summary and Overview of Toughening Mechanisms in Natural Materials
The examples discussed in this chapter show how natural materials can control deformation and prevent crack propagation by specific mechanisms and architectures. Tendon, fish scales, skin, nacre, tooth enamel, and bone display different types of architectures with a broad variety of toughening mechanisms. The loose organization of the collagen fibrils in skin allows for large deformations, crack tip rounding, and local re-orientation of the collagen fibrils. In more organized architectures like fish scales, bone, tooth enamel, and bone, crack deflection is prominent, and it is possible only if the interface is significantly weaker than the rest of the material (He and Hutchinson 1989). The inter-fibrillar interfaces in tendon must be much weaker than the collagen fibrils, the thin proteinaceous interfaces in nacre must be weaker than the mineral tablets, the thin cement lines in cortical bone must be weaker than the osteons. Weak interfaces are therefore a requirement for high toughness, a somewhat counterintuitive rule prevalent in fiber-reinforced composites (Evans 1989) and multilayered ceramics (Clegg et al. 1990). Weak interfaces are an universal theme in biological materials is the result of the adaptation and specialization of biological materials for specific functions. More isotropic structures are less specialized, and show less extreme mechanical properties (Nikolov et al. 2010). Crack deflection can trigger further toughening mechanisms such as crack bridging as seen in fish scales, tooth enamel, nacre or bone. The most powerful toughening mechanisms are associated with process zone toughening and inelastic deformations. This type of mechanism relies on volumetric dilation of the inelastic region and residual strains to generate toughness as the crack advances. In addition to spending energy to generate new surfaces, the material dissipates energy by the loading of the material ahead of the crack and unloading it behind the crack. Process zone toughening is the main toughening mechanisms in fish scales (Dastjerdi and Barthelat 2015) and nacre (Barthelat and Rabiei 2011). In these hard yet tough materials, inelastic deformations over large volumes near cracks cannot be dissociated from, and are critical to toughening mechanisms in ways similar to metals (Anderson 2005), phase transforming ceramics (Evans 1990) and reinforced epoxies (Evans et al. 1986). The amount of precision and effectiveness of toughening mechanisms in natural materials, as well as its controlled occurrence over multiple length scales, will continue to fascinate materials scientists and inspire new engineering materials in years to come. Other remarkable features such as self-healing to repair fatigue microcrack to remodeling to adapt to local stresses are unique to biological materials, but will perhaps also be used in the engineering materials of tomorrow.
References
Ager JW, Balooch G, Ritchie RO (2006) Fracture, aging, and disease in bone. J Mater Res 21:1878–92
Anderson TL (2005) Fracture mechanics: fundamentals and applications. Taylor & Francis, Boca Raton, FL
Ascenzi A, Bonucci E (1972) Shearing properties of single osteons. Anat Rec 172:499–510
Ashby M (2010) Materials selection in mechanical design. 4th Revised edition ed. Butterworth-Heinemann Ltd., Oxford
Bajaj D, Arola D (2009) Role of prism decussation on fatigue crack growth and fracture of human enamel. Acta Biomater 5:3045–56
Bajaj D, Park S, Quinn GD, Arola D (2010) Fracture processes and mechanisms of crack growth resistance in human enamel. Jom 62:76–82
Barthelat F (2007) Biomimetics for next generation materials. Philos Trans R Soc A-Math Phys Eng Sci 365:2907–19
Barthelat F (2015) Architectured materials in engineering and biology: fabrication, structure, mechanics and performance. Int Mater Rev 60:413–30
Barthelat F, Rabiei R (2011) Toughness amplification in natural composites. J Mech Phys Solids 59:829–40
Barthelat F, Tang H, Zavattieri PD, Li CM, Espinosa HD (2007) On the mechanics of mother-of-pearl: a key feature in the material hierarchical structure. J Mech Phys Solids 55:225–444
Barthelat F, Yin Z, Buehler MJ (2016) Structure and mechanics of interfaces in biological materials. Nat Rev Mater 1
Bass EC, Ashford FA, Segal MR, Lotz JC (2004) Biaxial testing of human annulus fibrosus and its implications for a constitutive formulation. Ann Biomed Eng 32:1231–42
Bechtle S, Fett T, Rizzi G, Habelitz S, Klocke A, Schneider GA (2010) Crack arrest within teeth at the dentinoenamel junction caused by elastic modulus mismatch. Biomaterials 31:4238–47
Bigi A, Burghammer M, Falconi R, Koch MHJ, Panzavolta S, Riekel C (2001) Twisted plywood pattern of collagen fibrils in teleost scales: an X-ray diffraction investigation. J Struct Biol 136:137–43
Bigley RF, Griffin LV, Christensen L, Vandenbosch R (2006) Osteon interfacial strength and histomorphometry of equine cortical bone. J Biomech 39:1629–40
Binici H, Aksogan O, Bodur MN, Akca E, Kapur S (2007) Thermal isolation and mechanical properties of fibre reinforced mud bricks as wall materials. Constr Build Mater 21:901–6
Bischoff JE, Arruda EM, Grosh K (2000) Finite element modeling of human skin using an isotropic, nonlinear elastic constitutive model. J Biomech 33:645–52
Browning A, Ortiz C, Boyce MC (2013) Mechanics of composite elasmoid fish scale assemblies and their bioinspired analogues. J Mech Behav Biomed Mater 19:75–86
Bruet BJF, Song JH, Boyce MC, Ortiz C (2008) Materials design principles of ancient fish armour. Nat Mater 7:748–56
Buehler MJ (2007) Molecular nanomechanics of nascent bone: fibrillar toughening by mineralization. Nanotechnology 18
Buehler MJ (2006) Nature designs tough collagen: explaining the nanostructure of collagen fibrils. Proc Natl Acad Sci USA 103:12285–90
Burr DB (2002) The contribution of the organic matrix to bone’s material properties. Bone 31:8–11
Burr DB, Schaffler MB, Frederickson RG (1988) Composition of the cement line and its possible mechanical role as a local interface inhuman compact-bone. J Biomech 21:939–945
Clegg WJ, Kendall K, Alford NM, Button TW, Birchall JD (1990) A simple way to make tough ceramics. Nature 347:455–7
Colfen H, Antonietti M (2005) Mesocrystals: inorganic superstructures made by highly parallel crystallization and controlled alignment. Angew Chem-Int Ed 44:5576–91
Cook J, Gordon JE, Evans CC, Marsh DM (1964) A mechanism for the control of crack propagation in all-brittle systems. Proc R Soc Lond A 282:508–20
Currey JD (1977) Mechanical properties of mother of pearl in tension. Proc R Soc Lond B 196:443–63
Currey JD (1999) The design of mineralised hard tissues for their mechanical functions. J Exp Biol 202:3285–94
Currey JD (2002) Bones: structure and mechanics. Princeton University Press, Princeton, NJ
Currey JD, Taylor JD (1974) The mechanical behavior of some molluscan hard tissues. J Zool (Lond) 173:395–406
Dastjerdi AK, Barthelat F (2015) Teleost fish scales amongst the toughest collagenous materials. J Mech Behav Biomed Mater 52:95–107
Dastjerdi AK, Rabiei R, Barthelat F (2013) The weak interfaces within tough natural composites: experiments on three types of nacre. J Mech Behav Biomed Mater 19:50–60
Dong XN, Zhang X, Guo XE (2005) Interfacial strength of cement lines in human cortical bone. Mech Chem Biosyst: MCB 2:63–8
Dunlop JWC, Weinkamer R, Fratzl P (2011) Artful interfaces within biological materials. Mater Today 14:70–8
Eppell SJ, Smith BN, Kahn H, Ballarini R (2006) Nano measurements with micro-devices: mechanical properties of hydrated collagen fibrils. J R Soc Interface 3:117–21
Espinosa HD, Juster AL, Latourte FJ, Loh OY, Gregoire D, Zavattieri PD (2011) Tablet-level origin of toughening in abalone shells and translation to synthetic composite materials. Nat Commun 2
Espinosa HD, Rim JE, Barthelat F, Buehler MJ (2009) Merger of structure and material in nacre and bone - perspectives on de novo biomimetic materials. Prog Mater Sci 54:1059–100
Evans AG (1989) The mechanical performance of fiber-reinforced ceramic matrix composites. Mater Sci Eng A 107:227–39
Evans AG (1990) Perspective on the development of high-toughness ceramics. J Am Ceram Soc 73:187–206
Evans AG, Ahmad ZB, Gilbert DG, Beaumont PWR (1986) Mechanisms of toughening in rubber toughened polymers. Acta Metall 34:79–87
Fantner GE, Birkedal H, Kindt JH, Hassenkam T, Weaver JC, Cutroni JA et al (2004) Influence of the degradation of the organic matrix on the microscopic fracture behavior of trabecular bone. Bone 35:1013–22
Fantner GE, Hassenkam T, Kindt JH, Weaver JC, Birkedal H, Pechenik L et al (2005) Sacrificial bonds and hidden length dissipate energy as mineralized fibrils separate during bone fracture. Nat Mater 4:612–6
Fantner GE, Rabinovych O, Schitter G, Thurner P, Kindt JH, Finch MM et al (2006) Hierarchical interconnections in the nano-composite material bone: fibrillar cross-links resist fracture on several length scales. Compos Sci Technol 66:1205–11
Forest Products Laboratory. Wood handbook: wood as an engineering material. [Madison]; Washington: The Laboratory; For sale by the Supt. of Docs., U.S. Govt. Print. Off.; 1974
Fratzl P (2003) Cellulose and collagen: from fibres to tissues. Curr Opin Colloid Interface Sci 8:32–9
Fratzl P, Weinkamer R (2007) Nature’s hierarchical materials. Prog Mater Sci 52:1263–334
Fratzl P, Misof K, Zizak I, Rapp G, Amenitsch H, Bernstorff S (1998) Fibrillar structure and mechanical properties of collagen. J Struct Biol 122:119–22
Fung YC (2004) Biomechanics: mechanical properties of living tissues. Springer, New York
Fung DT, Wang VM, Laudier DM, Shine JH, Basta-Pljakic J, Jepsen KJ et al (2009) Subrupture tendon fatigue damage. J Orthop Res 27:264–73
Garrano AMC, La Rosa G, Zhang D, Niu LN, Tay FR, Majd H et al (2012) On the mechanical behavior of scales from Cyprinus carpio. J Mech Behav Biomed Mater 7:17–29
Gautieri A, Uzel S, Vesentini S, Redaelli A, Buehler MJ (2009) Molecular and mesoscale mechanisms of osteogenesis imperfecta disease in collagen fibrils. Biophys J 97:857–65
Gupta HS, Wagermaier W, Zickler GA, Aroush DRB, Funari SS, Roschger P et al (2005) Nanoscale deformation mechanisms in bone. Nano Lett 5:2108–11
Gupta HS, Wagermaier W, Zickler GA, Hartmann J, Funari SS, Roschger P et al (2006) Fibrillar level fracture in bone beyond the yield point. Int J Fract 139:425–36
Gupta HS, Seto J, Wagermaier W, Zaslansky P, Boesecke P, Fratzl P (2006) Cooperative deformation of mineral and collagen in bone at the nanoscale. Proc Natl Acad Sci USA 103:17741–6
Gupta HS, Fratzl P, Kerschnitzki M, Benecke G, Wagermaier W, Kirchner HOK (2007) Evidence for an elementary process in bone plasticity with an activation enthalpy of 1 eV. J R Soc Interface 4:277–82
Habelitz S, Marshall SJ, Marshall GW, Balooch M (2001) Mechanical properties of human dental enamel on the nanometre scale. Arch Oral Biol 46:173–83
Haraldsson BT, Aagaard P, Krogsgaard M, Alkjaer T, Kjaer M, Magnusson SP (2005) Region-specific mechanical properties of the human patella tendon. J Appl Physiol 98:1006–12
Hassenkam T, Fantner GE, Cutroni JA, Weaver JC, Morse DE, Hansma PK (2004) High-resolution AFM imaging of intact and fractured trabecular bone. Bone 35:4–10
He MY, Hutchinson JW (1989) Crack deflection at an interface between dissimilar elastic materials. Int J Solids Struct 25:1053–67
Herod TW, Chambers NC, Veres SP (2016) Collagen fibrils in functionally distinct tendons have differing structural responses to tendon rupture and fatigue loading. Acta Biomater 42:296–307
Hiller LP, Stover SM, Gibson VA, Gibeling JC, Prater CS, Hazelwood SJ et al (2003) Osteon pullout in the equine third metacarpal bone: effects of ex vivo fatigue. J Orthop Res 21:481–488
Hui SL, Slemenda CW, Johnston CC (1988) Age and bone mass as predictors of fracture in a prospective-study. J Clin Investig 81:1804–9
Ikoma T, Kobayashi H, Tanaka J, Walsh D, Mann S (2003) Microstructure, mechanical, and biomimetic properties of fish scales from Pagrus major. J Struct Biol 142:327–33
Imbeni V, Kruzic JJ, Marshall GW, Marshall SJ, Ritchie RO (2005) The dentin-enamel junction and the fracture of human teeth. Nat Mater 4:229–32
Jackson AP, Vincent JFV, Turner RM (1988) The mechanical design of nacre. Proc R Soc Lond B 234:415–40
Kamat S, Su X, Ballarini R, Heuer AH (2000) Structural basis for the fracture toughness of the shell of the conch Strombus gigas. Nat Mater 405:1036–40
Kamat S, Kessler H, Ballarini R, Nassirou M, Heuer AH (2004) Fracture mechanisms of the Strombus gigas conch shell: II-micromechanics analyses of multiple cracking and large-scale crack bridging. Acta Mater 52:2395–406
Ker RF (2007) Mechanics of tendon, from an engineering perspective. Int J Fatigue 29:1001–9
Koester KJ, Ager JW III, Ritchie RO (2008) The true toughness of human cortical bone measured with realistically short cracks. Nat Mater 7:672–7
Kruzic J, Nalla RK, Kinney JH, Ritchie RO (2003) Crack blunting, crack bridging and resistance-curve fracture mechanics in dentin: effect of hydration. Biomaterials 24:5209–21
Launey ME, Buehler MJ, Ritchie RO (2022) On the mechanistic origins of toughness in bone. In: Clarke DR, Ruhle M, Zok F (eds) Annual review of materials research, vol 402010, pp 25–53
Lawn BR (1993) Fracture of brittle solids, 2nd edn. Cambridge University Press, New York
Lemmen C, Khan A (2012) A simulation of the neolithic transition in the indus valley. In: Giosan L, Fuller DQ, Nicoll K, Flad RK, Clift PD (eds) Climates, Landscapes, and Civilizations, pp 107-114
Levi-Kalisman Y, Falini G, Addadi L, Weiner S (2001) Structure of the nacreous organic matrix of a bivalve mollusk shell examined in the hydrated state using Cryo-TEM. J Struct Biol 135:8–17
Lichtenegger HC, Schoberl T, Ruokolainen JT, Cross JO, Heald SM, Birkedal H et al (2003) Zinc and mechanical prowess in the jaws of Nereis, a marine worm. Proc Natl Acad Sci USA 100:9144–9
Lin C, Douglas W (1994) Structure-property relations and crack resistance at the bovine dentin-enamel junction. J Dent Res 73:1072–8
Lin YS, Wei CT, Olevsky EA, Meyers MA (2011) Mechanical properties and the laminate structure of Arapaima gigas scales. J Mech Behav Biomed Mater 4:1145–56
Liu WT, Zhang Y, Li GY, Miao YQ, Wu XH (2008) Structure and composition of teleost scales from snakehead Channa argus (Cantor) (Perciformes: Channidae). J Fish Biol 72:1055–67
Lopez MI, Martinez PEM, Meyers MA (2014) Organic interlamellar layers, mesolayers and mineral nanobridges: contribution to strength in abalone (Haliotis rufescence) nacre. Acta Biomater 10:2056–64
Macho GA, Jiang Y, Spears IR (2003) Enamel microstructure-a truly three-dimensional structure. J Hum Evol 45:81–90
Mann S (2000) The chemistry of form. Angew Chem-Int Ed 39:3393–406
Mayer G (2005) Rigid biological systems as models for synthetic composites. Science 310:1144–7
Mercer C, He MY, Wang R, Evans AG (2006) Mechanisms governing the inelastic deformation of cortical bone and application to trabecular bone. Acta Biomater 2:59–68
Meunier J (1981) ‘Twisted plywood’ structure and mineralization in the scales of a primitive living fish Amia calva. Tissue Cell 13:165–71
Meunier FJ (1984) Spatial organization and mineralization of the basal plate of elasmoid scales in osteichthyans. Am Zool 24:953
Meunier FJ, Castanet J (1982) Organisation spatiale des fibres de collagène de la plaque basale des écailles des Téléostéens. Zool Scripta 11:141–53
Meyers MA, Chen PY, Lin AYM, Seki Y (2008) Biological materials: structure and mechanical properties. Prog Mater Sci 53:1–206
Meyers MA, Lin YS, Olevsky EA, Chen PY (2012) Battle in the Amazon: arapaima versus piranha. Adv Eng Mater 14:B279–B88
Mirkhalaf M, Dastjerdi AK, Barthelat F (2014) Overcoming the brittleness of glass through bio-inspiration and micro-architecture. Nat Commun 5
Nabavi A, Capozzi A, Goroshin S, Frost DL, Barthelat F (2014) A novel method for net-shape manufacturing of metal-metal sulfide cermets. J Mater Sci 49:8095–106
Nalla RK, Kinney JH, Ritchie RO (2003) Mechanistic fracture criteria for the failure of human cortical bone. Nat Mater 2:164–8
Nalla RK, Kruzic JJ, Kinney JH, Balooch M, Ager JW, Ritchie RO (2006) Role of microstructure in the aging-related deterioration of the toughness of human cortical bone. Mater Sci Eng: C 26:1251–60
Nanci A, Ten Cate AR (2013) Ten Cate’s oral histology: development, structure, and function. Elsevier, St. Louis, Mo
Nikolov S, Petrov M, Lymperakis L, Friák M, Sachs C, Fabritius H-O et al (2010) Revealing the design principles of high-performance biological composites using ab initio and multiscale simulations: the example of lobster cuticle. Adv Mater 22:519–26
Nikolov S, Petrov M, Lymperakis L, Friak M, Sachs C, Fabritius H-O, et al (2010) Revealing the design principles of high-performance biological composites using ab initio and multiscale simulations: the example of lobster cuticle. Adv Mater 22:519–526
Oxlund H, Manschot J, Viidik A (1988) The role of elastin in the mechanical-properties of skin. J Biomech 21:213–8
Peterlik H, Roschger P, Klaushofer K, Fratzl P (2006) From brittle to ductile fracture of bone. Nat Mater 5:52–5
Piekarsk K (1970) Fracture of bone. J Appl Phys 41:215–223
Poissant J, Barthelat F (2012) In situ mechanical testing of hydrated biological nanofibers using a nanoindenter transducer. Exp Mech 52:1287–95
Poundarik AA, Diab T, Sroga GE, Ural A, Boskey AL, Gundberg CM et al (2012) Dilatational band formation in bone. Proc Natl Acad Sci USA 109:19178–83
Provenzano PP, Vanderby R (2006) Collagen fibril morphology and organization: implications for force transmission in ligament and tendon. Matrix Biol 25:71–84
Purslow PP (1983) Measurement of the fracture-toughness of extensible connective tissues. J Mater Sci 18:3591–398
Puxkandl R, Zizak I, Paris O, Keckes J, Tesch W, Bernstorff S et al (2002) Viscoelastic properties of collagen: synchrotron radiation investigations and structural model. Philos Trans R Soc Lond Ser B-Biol Sci 357:191–7
Raabe D, Sachs C, Romano P (2005) The crustacean exoskeleton as an example of a structurally and mechanically graded biological nanocomposite material. Acta Mater 53:4281–92
Rabiei R, Bekah S, Barthelat F (2010) Failure mode transition in nacre and bone-like materials. Acta Biomater 6:4081–9
Rho JY, Kuhn-Spearing L, Zioupos P (1998) Mechanical properties and the hierarchical structure of bone. Med Eng Phys 20:92–102
Ridge MD, Wright V (1966) Directional effects of skin - a bio-engineering study of skin with particular reference to Langers lines. J Investig Dermatology 46:341–346
Ridge MD, Wright V (1966) The directional effects of skin: a bio-engineering study of skin with particular reference to Langer’s lines*. J Investig Dermatology 46:341–6
Ritchie RO (2011) The conflicts between strength and toughness. Nat Mater 10:817–22
Ritchie RO, Kinney JH, Kruzic JJ, Nalla RK (2005) A fracture mechanics and mechanistic approach to the failure of cortical bone. Fatigue Fract Eng Mater Struct 28:345–71
Ritchie RO, Buehler MJ, Hansma P (2009) Plasticity and toughness in bone. Phys Today 62:41–7
Rousseau M, Lopez E, Stempfle P, Brendle M, Franke L, Guette A et al (2005) Multiscale structure of sheet nacre. Biomaterials 26:6254–62
Scheyer TM, Sander PM, Joyce WG, Boehme W, Witzel U (2007) A plywood structure in the shell of fossil and living soft-shelled turtles (Trionychidae) and its evolutionary implications. Org Divers Evol 7:136–44
Schonborner AA, Boivin G, Baud CA (1979) The mineralization processes in teleost fish scales. Cell Tissue Res 202:203–12
Schwiedrzik J, Raghavan R, Buerki A, LeNader V, Wolfram U, Michler J et al (2014) In situ micropillar compression reveals superior strength and ductility but an absence of damage in lamellar bone. Nat Mater 13:740–7
Seshaiya RV, Ambujabay P, Kalyani M (1963) Amino acid composition of icthylepidin from fish scales. Asp Protein Struct 343–348
Seto J, Ma Y, Davis SA, Meldrum F, Gourrier A, Kim Y-Y et al (2012) Structure-property relationships of a biological mesocrystal in the adult sea urchin spine. Proc Natl Acad Sci 109:3699–704
Shen ZL, Dodge MR, Kahn H, Ballarini R, Eppell SJ (2008) Stress-strain experiments on individual collagen fibrils. Biophys J 95:3956–63
Silver FH, Siperko LM, Seehra GP (2003) Mechanobiology of force transduction in dermal tissue. Ski Res Technol 9:3–23
Svensson RB, Herchenhan A, Starborg T, Larsen M, Kadler KE, Qvortrup K et al (2017) Evidence of structurally continuous collagen fibrils in tendons. Acta Biomater 50:293–301
Szczesny SE, Caplan JL, Pedersen P, Elliott DM (2015) Quantification of interfibrillar shear stress in aligned soft collagenous tissues via notch tension testing. Scientific Reports, vol 5
Tang Y, Ballarini R, Buehler MJ, Eppell SJ (2010) Deformation micromechanisms of collagen fibrils under uniaxial tension. J R Soc Interface 7:839–50
Taylor D, Hazenberg JG, Lee TC (2007) Living with cracks: damage and repair in human bone. Nat Mater 6:263–8
Thurner PJ, Katsamenis OL (2014) The role of nanoscale toughening mechanisms in osteoporosis. Curr Osteoporos Rep 12:351–6
Thurner PJ, Chen CG, Ionova-Martin S, Sun L, Harman A, Porter A et al (2010) Osteopontin deficiency increases bone fragility but preserves bone mass. Bone 46:1564–1573
Torres FG, Troncoso OP, Nakamatsu J, Grande CJ, Gomez CM (2008) Characterization of the nanocomposite laminate structure occurring in fish scales from Arapaima Gigas. Mater Sci Eng C-Biomim Supramol Syst 28:1276–83
Ural A, Vashishth D (2014) Hierarchical perspective of bone toughness - from molecules to fracture. Int Mater Rev 59:245–63
Uzel SGM, Buehler MJ (2011) Molecular structure, mechanical behavior and failure mechanism of the C-terminal cross-link domain in type I collagen. J Mech Behav Biomed Mater 4:153–61
Vashishth D (2007) Hierarchy of bone microdamage at multiple length scales. Int J Fatigue 29:1024–33
Vashishth D, Tanner KE, Bonfield W (2003) Experimental validation of a microcracking-based toughening mechanism for cortical bone. J Biomech 36:121–4
Vernerey FJ, Barthelat F (2010) On the mechanics of fishscale structures. Int J Solids Struct 47:2268–75
Vincent JFV, Bogatyreva OA, Bogatyrev NR, Bowyer A, Pahl AK (2006) Biomimetics: its practice and theory. J R Soc Interface 3:471–82
Wang RZ, Suo Z, Evans AG, Yao N, Aksay IA (2001) Deformation mechanisms in nacre. J Mater Res 16:2485–93
Wegst UGK, Ashby MF (2004) The mechanical efficiency of natural materials. Philos Mag 84:2167–81
Wegst UGK, Bai H, Saiz E, Tomsia AP, Ritchie RO (2015) Bioinspired structural materials. Nat Mater 14:23–36
Weiner S, Addadi L (1997) Design strategies in mineralized biological materials. J Mater Chem 7:689–702
Weiner S, Wagner HD (1998) The material bone: structure mechanical function relations. Annu Rev Mater Sci 28:271–98
Yahyazadehfar M, Arola D (2015) The role of organic proteins on the crack growth resistance of human enamel. Acta Biomater 19:33–45
Yahyazadehfar M, Bajaj D, Arola DD (2013) Hidden contributions of the enamel rods on the fracture resistance of human teeth. Acta Biomater 9:4806–14
Yahyazadehfar M, Ivancik J, Majd H, An B, Zhang D, Arola D (2014) On the mechanics of fatigue and fracture in teeth. Appl Mech Rev 66:1–19
Yang W, Chen IH, Gludovatz B, Zimmermann EA, Ritchie RO, Meyers MA (2013a) Natural flexible dermal armor. Adv Mater 25:31–48
Yang W, Gludovatz B, Zimmermann EA, Bale HA, Ritchie RO, Meyers MA (2013b) Structure and fracture resistance of alligator gar (Atractosteus spatula) armored fish scales. Acta Biomater 9:5876–89
Yang W, Sherman VR, Gludovatz B, Mackey M, Zimmermann EA, Chang EH et al (2014) Protective role of Arapaima gigas fish scales: structure and mechanical behavior. Acta Biomater 10:3599–614
Yang W, Sherman VR, Gludovatz B, Schaible E, Stewart P, Ritchie RO et al (2015) On the tear resistance of skin. Nat Commun 6:6649
Young MF (2003) Bone matrix proteins: their function, regulation, and relationship to osteoporosis. Osteoporos Int 14:S35–S42
Zhu D, Ortega CF, Motamedi R, Szewciw L, Vernerey F, Barthelat F (2012) Structure and mechanical performance of a “modern’’ fish scale. Adv Eng Mater 14:B185–B94
Zhu D, Szewciw L, Vernerey F, Barthelat F (2013) Puncture resistance of the scaled skin from striped bass: collective mechanisms and inspiration for new flexible armor designs. J Mech Behav Biomed Mater 24:30–40
Zimmermann EA, Gludovatz B, Schaible E, Dave NKN, Yang W, Meyers MA et al (2013) Mechanical adaptability of the Bouligand-type structure in natural dermal armour. Nat Commun 4
Zimmermann EA, Schaible E, Bale H, Barth HD, Tang SY, Reichert P et al (2011) Age-related changes in the plasticity and toughness of human cortical bone at multiple length scales. Proc Natl Acad Sci USA 108:14416–21
Zioupos P (1998) Recent developments in the study of failure of solid biomaterials and bone: ‘fracture’ and ‘pre-fracture’ toughness. Mater Sci Eng: C 6:33–40
Zioupos P, Gresle M, Winwood K (2008) Fatigue strength of human cortical bone: age, physical, and material heterogeneity effects. J Biomed Mater Res Part A 86A:627–36
Zok FW (2006) Developments in oxide fiber composites. J Am Ceram Soc 89:3309–24
Zylberberg L, Bereiterhahn J, Sire JY (1988) Cytoskeletal organization and collagen orientation in the fish scales. Cell Tissue Res 253:597–607
Acknowledgements
This chapter is based on lecture notes from a CISM school on “Mechanics and Physics of Fracture: Multi-scale modeling of the failure behavior of solids” held September 26–30, 2016. FB would like to acknowledge the contribution of Will Pro to the final preparation steps of this chapter.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 CISM International Centre for Mechanical Sciences
About this chapter

Cite this chapter
Barthelat, F. (2023). The Fracture Mechanics of Biological Materials. In: Ponson, L. (eds) Mechanics and Physics of Fracture. CISM International Centre for Mechanical Sciences, vol 608. Springer, Cham. https://doi.org/10.1007/978-3-031-18340-9_4
Download citation
DOI: https://doi.org/10.1007/978-3-031-18340-9_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-18339-3
Online ISBN: 978-3-031-18340-9
eBook Packages: EngineeringEngineering (R0)