
Abstract
The fungus Botrytis cinerea (Botryotinia fuckeliana) infects more than 500 plant species and causes a wide range of symptoms: soft rots, accompanied by collapse and water-soaking of tissues followed by the appearance of gray masses of conidia on leaves and soft fruits (gray mold), and spots that may turn brown to full-scale soft rotting on flower petals (Botrytis blight). In general, B. cinerea is responsible for severe economic losses that are either due to the damage of growing plants in the field or the rot of harvested fruits, flowers, and vegetables during storage under cold and humid conditions. B. cinerea has adapted to the plant host and its environment by evolving strategies to use plant tissues for proliferation in terms of a necrotrophic lifestyle and to survive biotic (host responses) as well as abiotic stresses such as sunlight. B. cinerea maintains a complex regulatory network of light-sensitive proteins and signal transduction pathways to use light for coordinating stress responses, virulence, and reproduction. Different light-controlled reproduction cycles enable B. cinerea to live in moderate climate zones by infecting and propagating in summer and remaining dormant in winter when green host tissues are unavailable as a nutrient source.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Adaptations to the Plant Host
The interaction between B. cinerea and its host plants cover a spectrum of outcomes, ranging from full immunity (no infection) to full susceptibility (fast infection) and all the intermediate stages in between. Another lifestyle option, although not yet that well understood, is the asymptomatic colonization of the host (Williamson et al. 2007; Shaw et al. 2016; Veloso and van Kan 2018). The outcome of the interaction with the host depends on the host species and its capacity to prevent fungal growth, the B. cinerea strain and its growth characteristics, and the climate conditions which can be more favorable for the host or the pathogen. Notably, B. cinerea field populations are characterized by high levels of variability in morphology and preferred mode of reproduction, virulence, spectra of secondary metabolites produced, and fungicide resistance. The genetic variability can be achieved through sexual reproduction, hyphal anastomosis, transposable elements, heterokaryosis, heteroploidy, or viruses (Coley-Smith et al. 1980; Elad et al. 2004; Fillinger and Elad 2016). In this regard, it is important to use reference host plants and B. cinerea strains in the laboratory for elucidating the mechanisms and regulation of pathogen attack, host immune response, and fungal reproduction. Phaseolus vulgaris (French bean) and Arabidopsis thaliana are valuable model experimental hosts that exhibit high and low susceptibility to aggressive B. cinerea strains such as B05.10 (Fig. 14.1). Thus, the use of the highly sensitive host, P. vulgaris, enables the study of the interaction from the fungal perspective by revealing differences between the capacity of B. cinerea mutants to enter and colonize plant tissues (Giesbert et al. 2012; de Vallee et al. 2019). In contrast, the role of plant immune responses can be studied in (genetically modified) A. thaliana, which represents a “poor host” because of its potent phytoalexin camalexin (Denby et al. 2004; Rowe and Kliebenstein 2008; Mengiste et al. 2009).

Infection cycle of B. cinerea B05.10 under laboratory conditions. (a) On Phaseolus vulgaris (French bean) as a highly susceptible host. Primary leaves of living plants are inoculated with plugs of vegetative mycelia (two-day-old complete medium (CM) cultures) or conidial suspensions (2 × 105 conidia/ml Gamborg B5 + 2% glucose) and are incubated under humid conditions and natural light conditions at room temperature. The fungus establishes primary (restricted) infections that spread quickly. After one week the whole leaf is infected: the plant tissue collapses and becomes watery (soft rot), which is accompanied by the formation of gray conidiophores and conidia. The penetration process is monitored on isolated onion epidermis. Hyphae deriving from vegetative mycelia produce infection cushions; conidia form short germ tubes that directly penetrate the epidermal cells (hyphae growing inside the onion epidermis are colorless while fungal structures outside the host are stained with lactophenol blue). Dpi—days post-inoculation. (b) On the model plant Arabidopsis thaliana as a lowly susceptible host. Leaves of living plants can be infected by conidia as described above, resulting in fast spreading and soft rot formation. In comparison to French bean, no defined stages (i.e., primary and secondary infections) can be distinguished. Due to the high level of basal resistance, the outcomes of different assays may vary significantly. A. thaliana is likely not a (natural) host for B. cinerea outside the laboratory
The life cycle of B. cinerea includes vegetative mycelia for substrate colonization, (macro)conidia for dispersal, sclerotia for survival and as a prerequisite for sexual development, microconidia as male gametes, and fruiting bodies bearing the sexual spores (Fig. 14.2). The relationship with the host further includes the development of penetration structures and the active secretion of factors for killing host cells and for obtaining nutrition from the dead tissue.

Reproduction cycles of light-responsive B. cinerea strains. Young vegetative mycelia may develop either macro-conidia for dispersal (summer cycle, green) or sclerotia for survival (winter cycle, blue). Both conidia and sclerotia germinate asexually under favorable conditions. Fertilization of the sclerotia by microconidia of the opposite mating type is part of the sexual cycle that results in the formation of apothecia as fruiting bodies in spring. The apothecia contain asci with eight ascospores in ordered tetrads. Light promotes conidiation and represses sclerotial development. Sclerotia and microconidia are formed under the same conditions that is the absence of light. Left bottom corner: conidiophores with conidia and sclerotia (blue arrows) formed by B. cinerea on Hydrangea plants from the greenhouse (left) and a dark cold storage (right)
1.1 Penetration Structures
B. cinerea enters the host by using two different pre-penetration structures (Backhouse and Willets 1987; Choquer et al. 2007). The conidia that are ubiquitously distributed in the air germinate after landing on the plant surface under favorable conditions and form short germ tubes with apical swellings (appressorium-like structures), which directly penetrate the plant surface. Alternatively, hyphae derived from already established mycelia or older elongated germ tubes form multicellular penetration structures, the so-called infection cushions. The penetration process by germ tubes and infection cushions can be easily monitored on onion epidermis by using lactophenol blue staining for distinguishing structures inside and outside of the onion cells (Fig. 14.1).
Several signaling pathways are implicated in the early stages of plant infection. Deletion of the genes encoding the mitogen-activated protein (MAP) kinases, BMP1 and BcSAK1, results in avirulent mutants (Zheng et al. 2000; Segmüller et al. 2007; Schamber et al. 2010). In contrast, deletion of the genes encoding the MAP kinase BMP1 and components of the cAMP cascade leads to mutants with retarded infection due to reduced penetration efficiency (Doehlemann et al. 2006b; Rui and Hahn 2007; Schumacher et al. 2008b). Deletion of genes encoding members of the NoxB (NADPH oxidase) complex results in another interesting phenotype: conidia germinate, form germ tubes and appressorium-like structures, which initiate penetration but without success. New outgrowths appear that are followed by further attempts to penetrate. Components of the NoxA complex are specifically involved in the formation of infection cushions. However, all NOX mutants eventually colonize the host tissue (Segmüller et al. 2008; Marschall et al. 2016). An intact fungal wall cell is critical for the differentiation of functional infections cushions as highlighted by phenotype analysis of Δbcchs3a, Δbccrz1, and Δbcskn7 mutants that are more strongly impaired in mycelia-derived than in conidia-derived infection processes (Schumacher et al. 2008a; Arbelet et al. 2010; Viefhues et al. 2015).
Taken together, conidia and mycelia of the same strain may have different penetration capabilities due to the two different kinds of infection structures whose formation is regulated by independent pathways. A recent study identified genes that are upregulated in mature infection cushions including two that encode secreted fasciclin-like proteins that are required for full virulence on P. vulgaris (Choquer et al. 2021).
1.2 Virulence Determinants
After penetration, the epidermal and underlying cells die and B. cinerea establishes a primary infection that is characterized by collapsed brownish tissue and defined margins. At this stage, hyphae are probably restricted to this region. The subcuticular mycelia may remain inactive in this restricted lesion for an indefinite duration (quiescence) and become active again under specific circumstances, such as during the ripening processes in which fungitoxic compounds decline in concentration, levels of sugars increase and the chemical composition of the cell wall changes (Prusky et al. 2013). Finally, B. cinerea overcomes the plant defense barriers and initiates vigorous secondary growth; hyphae grow invasively and decompose the plant tissue very rapidly resulting in soft rot and formation of conidiophores and conidia (Fig. 14.1).
B. cinerea secretes several non-host-specific cell death-inducing factors, which are low molecular weight metabolites or proteins (van Kan 2006; Amselem et al. 2011). The expression of these factors may be induced during the fungus-host interaction but synthesis is not restricted to this state. Two families of phytotoxic secondary metabolites, the botryanes (BOT) and the botcinins (BOA) contribute to full virulence (Dalmais et al. 2011; Viaud et al. 2016). Reactive oxygen species (ROS) play a dual role in the relationship as both interacting partners produce ROS. H2O2 generation is observed in the penetration structures, in and around the penetrated host cell wall in the early stages and in colonized tissues (Choquer et al. 2007; Heller and Tudzynski 2011). Several secreted necrosis-inducing proteins have been identified of but only a few contribute significantly to virulence. Cell wall-degrading enzymes (CWDEs) may be involved in different stages of plant infection: in penetration by lysing the cuticle or epidermal cell walls, in the manifestation of infection by degrading components of the plant defense, or in nutrition by effective decomposition of dead tissues. Some CWDEs also exhibit necrotizing activity (González et al. 2016). The redundancy of these groups of virulence determinates has been demonstrated by Hahn & co-workers who generated mutants using CRISPR/Cas9 technology where multiple genes encoding cell death-inducing proteins and key enzymes for the synthesis of BOT and BOA were deleted. On all tested tissues, infection efficiency of the mutants decreased with increasing number of deleted genes. However, even mutants with twelve genes deleted were eventually able to colonize the host tissue and to form conidia (Leisen et al. 2021).
A relatively new class of secreted effectors produced by B. cinerea are small RNAs (sRNAs) that derive from retrotransposons. They are taken up and recognized by the host resulting in the suppression of defense-related gene expression due to RNA interference (pathogen-induced gene silencing). Deletion of the genes encoding fungal Dicer-like proteins, BcDCL1 and BcDCL2, abolishes the generation of sRNAs and consequently results in reduced virulence. On the other hand, the mutation of AGO1 in A. thaliana that is required for active gene silencing leads to decreased susceptibility, indicating that cellular components of both interaction partners are involved in the process (Weiberg et al. 2013; Wang et al. 2016; Porquier et al. 2021).
Like other necrotrophs, B. cinerea induces plant defense responses by pathogen-associated molecular pattern (PAMP) recognition, such as cell wall fortification and production of antifungal compounds that activate JA/ET (jasmonate/ethylene)-dependent signaling pathways. Mutations that abolish JA and ET signaling result in increased susceptibility to B. cinerea (Mengiste et al. 2009). In addition, B. cinerea triggers SA (salicylic acid) signaling to induce the hypersensitive response (HR), a localized cell death reaction that is effective against biotrophic but supports necrotrophic pathogens (El Oirdi et al. 2011; Rossi et al. 2011). As JA/ET and SA pathways are antagonistic, the induction of SA signaling concurrently results in the suppression of JA/ET-associated defense responses (Robert-Seilaniantz et al. 2007; Mengiste 2012). Furthermore, B. cinerea may affect the hormone balance by fungal production of ethylene, auxins, and abscisic acid (ABA) (Sharon et al. 2004). However, the impact of fungal-derived phytohormones in the interaction is still unclear. JA-dependent defense responses against necrotrophs are downregulated in plants in which the shade avoidance response has been initiated. This program is induced by a low red/far-red light ratio which indicates the presence of competitors and the concomitant decrease of photosynthetic yields. The shade avoidance response consequently redirects more carbon resources to promote growth for optimizing photosynthetic yields and allows the plants to finish the life cycle at the expense of defense (Ballaré 2014; Ballaré and Pierik 2017; Liu et al. 2021). Because of its short repetitive infection cycles that yield high numbers of airborne conidia for consecutive infections of “immunocompromised” high-yielding host plant varieties planted in high densities, B. cinerea is a highly destructive pathogen in industrial monocultures.
2 Adaptations to Light
Sunlight is an important environmental factor in almost all ecosystems as it represents a source of energy, information, and stress. Cyanobacteria, algae and plants all use light for energy generation (photosynthesis). Less apparent is the utilization of light for repair of photodamaged DNA or for establishing proton gradients by microbial opsins. All organisms must protect themselves from the harmful effects of light such as UV radiation, ROS accumulation, heat, and desiccation. Finally, light quality and quantity can be used for signaling (photoinduction, photomorphogenesis), as a guide for directed growth (phototropism) and timing (photoentrainment), processes that include sensing and transduction into intracellular signals.
B. cinerea and other plant pathogens infecting the sun-exposed parts of the plant must cope with the high (UV, blue) light conditions the host plant seeks. Furthermore, the fungus experiences an altered light spectrum (‘green gap’) when it colonizes shaded parts of the plant. It is depleted for blue and red light that is absorbed by the plant chlorophyll and enriched for green and far-red light that is reflected or transmitted by the plant tissue. As these ambient light conditions trigger the shade avoidance response in the plant, the pathogens may trigger their own “shade response” such as the upregulation of virulence determinants and inoculum production assuming that new host tissues are close and highly susceptible.
2.1 Genetic Make-up: Photoperception
Light is perceived by chromophore-containing proteins called photoreceptors (PRs). FAD (flavin adenine dinucleotide), MTHF (5,10-methyltetrahydrofolate), retinal and bilin are bound by conserved input domains allowing for the distinction of four major PR classes: (i) near-UV/blue-sensing cryptochrome/photolyase-family proteins (CPFs), (ii) blue-sensing light-oxygen-voltage domain-containing proteins (LOVs), green-sensing membrane-associated opsins (OPSs) and red/far-red-sensing phytochromes (PHYs) (Heintzen 2012). Great numbers of PR-encoding genes are found in phototroph-associated microbes—including plant-pathogenic and rock-inhabiting fungi—pointing to the relevance of light in their life cycles (Schumacher and Gorbushina 2020; Losi and Gärtner 2021). B. cinerea possesses substantially more PRs than the long-established models in fungal photobiology, including Aspergillus nidulans which has three and Neurospora crassa which has eight PRs (Dasgupta et al. 2016). Striking is the expansion of PHYs (at least two) in the Leotiomycetes. The presence of two CPFs (BcCRY1, 2), four LOVs (BcWCL1, VVD1, LOV3, 4), two OPSs (BOP1, 2), and three PHYs (BcPHY1, 2, 3) enables B. cinerea to sense near-UV, blue, green, red and far-red light. Yet the mechanism and proteins involved for sensing UV (280–315 nm) are unknown and thus the total number of PRs of B. cinerea may be even larger. The eleven PRs of B. cinerea possess the conserved chromophore binding sites, and all are expressed in response to light. While deletion and overexpression mutants have been generated, characterization of these mutants is just beginning (Schumacher 2017).
2.2 Gene Expression: Photoregulation
Changes at the transcriptional level are prerequisites for all photoresponses and indicate that the organisms perceive and respond to light. But not all changes in gene expression will result in obvious phenotypes. Thus, PRs and light-responsive transcription factors (LTFs), which may control the expression of several genes have been studied to elucidate the regulatory networks and genes involved in the photoresponses.
The exposure of vegetative mycelia of B. cinerea B05.10 to full-spectrum light significantly alters gene expression (Fig. 14.3). The group of light-induced genes (LIGs) comprises genes involved in photoperception, stress responses, and transcriptional regulation. The group of light-repressed genes (LRGs) is smaller and enriched for genes encoding amino acid transporters (Schumacher et al. 2014). LIG expression occurs with different kinetics: genes are either subjected to photoadaptation or not. LIGs of the first group (early, late) mediate short-term (adaptive) responses. LIGs of the second group exhibit maximal expression after longer exposure to light and may be critical for development (dev LIGs). The expression levels of all PR-encoding genes are influenced by light: bccry1, bccry2, bcvvd1, and bop1 are early LIGs, bclov3, bcphy2 and bop2 are late LIGs, and bcphy1, bcphy3, bclov4, and bcwcl1 belong to the group of dev LIGs. Importantly, the blue light-sensing BcWCL1 as part of the White Collar complex (WCC) acts as the primary PR by modulating the expression levels of the remaining PR-encoding genes within a short period of time (Schumacher 2017). To date, eight LTFs out of the identified 30 LTFs (Schumacher et al. 2019) have been functionally characterized including the early LIGs that encode BcLTF3 and BcREG1, the late LIGs that encode BcLTF1, BcATF1 and BcLTF4, 5, 6, and the only studied dev LIG that encodes BcLTF2 (Michielse et al. 2011; Temme et al. 2012; Schumacher et al. 2014; Cohrs et al. 2016; Brandhoff et al. 2017).

Prominent light responses of B. cinerea WT:B05.10—morphogenesis and gene expression. (a) Light induces conidiation and represses sclerotium formation via transcriptional activation of BcLTF2. Low light dosages received in the competent stage of vegetative mycelia prevent the formation of sclerotia. Conidia and sclerotia as observed after two weeks of cultivation on solidified CM in 12 h light/12 h dark (LD) or constant darkness (DD) at 20 °C; vegetative mycelium as formed from a mycelial plug after two days of incubation in darkness. (b) Light-dark cycles are more effective than constant light (LL) and cause rhythmic conidiation (“rings”). Conidiophores with conidia are formed overnight on aerial mycelia that have been grown during the day (yellow brackets). (c) Transcriptional light responses are studied in surface-grown mycelia obtained from cellophane-covered solidified CM. Dark-grown mycelia are exposed to light pulses (LPs) before harvest. (d) Gene expression profiles determined by RT-qPCR of three light-responsive transcription factors (LTFs) exhibiting different kinetics. Bcltf3, ltf1, and ltf2 represent early, late, and developmental light-induced genes (LIGs) and affect conidiation
2.3 Reproduction: Photomorphogenesis and Phototropism
B. cinerea can reproduce asexually via macroconidia and sclerotia, and sexually via ascospores. The reproduction cycles depend on the availability of a host and the environmental conditions –mainly light and its absence, temperature, substrate—and are therefore linked to the seasons. Macroconidia are predominantly formed in the summer as inoculum for new infections (host tissue is available) and sclerotia in autumn for surviving the winter in the absence of a host. Sexual reproduction—fertilization of sclerotia with microconidia and subsequent apothecial development—is promoted by nutrient starvation and lower temperatures and happens in winter, resulting in the appearance of the fruiting bodies (apothecia) the following spring. Non-fertilized sclerotia and ascospores germinate with mycelia and conidia to initiate the summer cycle (Williamson et al. 2007; Amselem et al. 2011) (Fig. 14.2). Field populations of B. cinerea are highly diverse regarding the formation of these reproduction structures. Strains may or may not differentiate macroconidia, microconidia, sclerotia, and apothecia with ascospores during their life cycles. In general, B. cinerea strains can be classified into light-responsive strains that undergo photomorphogenesis, and blind strains that exhibit the same phenotypes in light and constant darkness (always mycelia, always sclerotia or always conidia). Light responsiveness allows for the formation of all structures—as found in strain B05.10 that is used in most laboratories as the host for genetic modification (Fig. 14.3). Photomorphogenesis is considered a characteristic feature of B. cinerea and its absence as the result of mutated regulatory genes.
Vegetative mycelia of B. cinerea B05.10 quickly colonize the host tissue (invasive growth) or the agar medium (saprophytic growth). For example, the growth rates on solidified complete medium are 10–15 mm per day so that the area of a standard Petri dish is colonized after three days of incubation under optimal temperature conditions (~20 °C). Little aerial hyphae and biomass accumulates as mycelia consist of thin layers of hyphae. Young vegetative mycelia are fully competent and can develop conidiophores with macroconidia or sclerotia. Light responsiveness depends on the physiological age and is less effective in aged colonies that have completely colonized the substrate or have already initiated one of the reproduction programs.
The (macro)conidia are short-lived asexual dispersal units. They are multinucleate, contain a two-layered cell wall—the outer wall is melanized—and are formed at denticles that arise from the spherical ampulla located at the tip of a conidiophore (synchronous holoblastic conidiogenesis) (Cole 1981). The development of conidia (conidiation) starting from initial conidiophores through to the presence of mature conidia may be completed within eight hours and requires light. Alternation of black light (near-UV) and white light, or alternation of full-spectrum light and darkness (Fig. 14.3) are most effective in promoting conidiation, while blue light alone inhibits conidiation and causes morphological changes in the various stages of conidiogenesis as mature conidiophores, ampullae at the tips of conidiophores, denticles, and conidial initials de-differentiate into sterile hyphae. Inhibition by blue light can be overcome by subsequent exposure to near-UV or far-red light. Red light does not promote conidiation after blue light inhibition and reduces conidiation in far-red light-treated colonies (Tan 1975; Suzuki et al. 1977).
Conidiophores positively respond to blue light after induction of conidiation by black light (positive phototropism)—most likely mediated through the blue light-sensing WCC—while green, yellow, and red light are ineffective (Jarvis 1972). Three TFs are known to be required for conidiogenesis: Bcreg1 mutants form conidiophores with ampullae and denticles but fail to form mature conidia, the mutation of bcltf3 phenocopies the blue light effect as the conidial initials grow out to sterile hyphae, and BcHOX8 is crucial for both proper conidiophore development and conidiogenesis (Michielse et al. 2011; Antal et al. 2012; Brandhoff et al. 2017). Initiation of conidiophore development requires BcLTF2 (sufficient and essential for conidiation), the stress-activated MAPK module, the response regulator BcSKN7, and the cell wall integrity-related MAPK module (Rui and Hahn 2007; Segmüller et al. 2007; Yan et al. 2010; Liu et al. 2011; Yang et al. 2012, 2015; Viefhues et al. 2015; Cohrs et al. 2016; Wang et al. 2018). Conidiation is repressed by the transcription factors BcLTF1, BcWCL1, BcMADS1, BcSTE12, the hybrid-histidine kinases BOS1 and BcSLN1 acting upstream of the BcSAK1 module, and the VELVET complex (Liu et al. 2008; Schamber et al. 2010; Schumacher et al. 2012, 2014, 2015; Canessa et al. 2013; Yang et al. 2013; Zhang et al. 2016; Müller et al. 2018; Ren et al. 2019).
The conidia may stay dormant for long periods, held in check by lack of moisture and nutrients. Germination starts with the swelling of the conidium, followed by the emergence of the germ tube. This is accompanied by nuclear divisions, the induction of genes encoding secreted proteins and lytic enzymes, and the production of an extracellular polysaccharide (EPS) matrix (Gull and Trinci 1971; Doss 1999; Leroch et al. 2013). Differences between isolates were observed regarding the induction of this process; some B. cinerea isolates germinate readily in distilled water, while others require sugars and amino acids (Blakeman 1975). Conidial germination on hydrophilic surfaces is stimulated by carbon sources and requires the MAP kinase BMP1, the Gα subunit BCG3 and the adenylate cyclase BAC, but not by the cAMP-dependent protein kinase (PKA), and is accompanied by the rapid degradation of trehalose, an important carbon storage compound and stress protectant (Doehlemann et al. 2006a, 2006b; Schumacher et al. 2008b). Germination on hydrophobic surfaces in the absence of nutrients is controlled by the MAP kinase BMP1 in a cAMP-independent manner (Doehlemann et al. 2006b). Other mutations result in multiple or elongated germ tubes with abnormal shapes, possibly due to defects in surface sensing (Rui and Hahn 2007; Kokkelink et al. 2011; Leroch et al. 2015). Experimental evidence for the secretion of self-inhibitors of germination by B. cinerea exist (Carlile and Sellin 1963; Kritzman et al. 1980); yet the compounds and regulatory networks remain elusive.
A recent study reported on the presence of 1-phenylethanol and 3-phenylpropanol and the absence of farnesol and tyrosol—four commonly involved compounds in quorum sensing in bacteria and fungi—in culture extracts of B. cinerea (Rosero-Hernandez and Echeverri 2020). Three forms of conidial fusions can be distinguished: fusions between conidia via a short connective hypha (conidial anastomosis tubes, CATs) (dominant type), fusions between germ tubes and conidia, and fusions between germ tubes directly or indirectly via CATs (Akutsu et al. 1981). Conidial/germling fusions are induced by a lack of nutrients and require the NoxA complex (Roca et al. 2012; Siegmund et al. 2015). Another factor influencing the germination process is light. Conidia germinate less well under green light conditions (Zhu et al. 2013), and germ tubes develop more frequently at the un-illuminated site of the conidium and grow away from the white light source (negative phototropism) (Robinson 1914; Gettkandt 1954; Islam et al. 1998). These observations suggest that light guides the germ tubes to the host surface and prevents conidial germination during the day.
Sclerotia are dark pigmented structures of limited growth, which allow the fungus to survive many years in the absence of suitable hosts or conditions favoring active growth. Sclerotial development consists of the three stages (i) initiation: formation of dichotomous branches, (ii) development: growth to the full size, and (iii) maturation: surface delimitation, internal changes, pigmentation of the peripheral hyphae, and exudation of water. The mature sclerotium has three distinct layers consisting of a melanized outer rind, a cortex of thin-walled pseudoparenchymatous cells, and a large medulla of loosely arranged filamentous hyphae. The formation of sclerotia is influenced by different factors, such as temperature, nutrition, pH, physical damage and growth against mechanical barriers, and is generally promoted by lower temperatures, high humidity, and the absence of light (Coley-Smith et al. 1980). The NoxA complex is essential for sclerotial development, which might be due to the inability of the mutants to form anastomoses (hyphal fusions) as common features of infection cushions and sclerotia (Segmüller et al. 2008; Siegmund et al. 2015). Several other mutants do not produce sclerotia, but this is due to the deregulation of morphogenetic programs rather than due to defects in sclerotial development. For example, inappropriate induction of vegetative growth resulting in always mycelia is caused by the overexpression of WCC components or deletion of the Gβγ dimer of heterotrimeric G proteins (Canessa et al. 2013; Tang et al. 2021). Conidiation in the dark by always conidia mutants has been described above. Only a few mutations are known to cause the always sclerotia phenotype; for example the deletion of bckdm1 encoding a putative histone 3 lysine 36 (H3K36) demethylase (Schumacher et al. 2019). The sclerotia may remain quiescent for long periods and can germinate in the presence of nutrients and after low temperature activation in three different ways: myceliogenically (by hyphae), sporogenically (by conidiophores), or carpogenically (by apothecia) (Coley-Smith and Cooke 1971) (Fig. 14.2).
Microconidia are the male gametes (spermatia). They are uninucleate, contain a lightly melanized cell wall, and are formed by phialides on conidial germ tubes, more mature hyphae, infection cushions or sclerotia under nutrient limitation conditions. They germinate exclusively in the presence of a female parent of the opposite mating type (Faretra and Antonacci 1987; Fukumori et al. 2004). The process of microconidiation is rather poorly studied. Nevertheless, microconidia and sclerotia are formed simultaneously in the laboratory—that is after two weeks in the absence of light under nutrient-rich conditions—suggesting that both production structures are formed at the same time in the field so that mating can occur to initiate the sexual cycle.
Apothecia (fruiting bodies) contain sexual spores. They arise from sclerotia after the sexual process (spermatization by microconidia) and are 2–20 mm in size. The upper part (disc) consists of a hymenial layer composed of paraphyses and asci containing the eight uninucleate ascospores in ordered tetrads. B. cinerea is self-sterile/heterothallic with the two mating types MAT1–1 and MAT1–2. Strains of the opposite mating types may be reciprocally crossed by using either sclerotia or microconidia. Derivatives of strain B05.10 and SAS56 (MAT1–1) can be crossed with the wild strain SAS405 (MAT1–2) (Faretra et al. 1988b; Amselem et al. 2011). There is little information about the physiology of the apothecia. Carpogenic germination of sclerotia requires pre-conditioning in cold, moist conditions, moderate temperatures, sufficient air, and light. Thereby, apothecial initials grow toward the light source (positive phototropism). Besides, the origin of the sclerotia plays a role as smaller sclerotia from minimal medium are unable to form apothecia likely due to the shortage of storage substances (Faretra and Antonacci 1987; Faretra et al. 1988a). Mutants may be female-sterile because they are unable to form sclerotia or the sclerotia are non-functional, but this does not imply male sterility (Segmüller et al. 2008; Jonkers et al. 2011; Siegmund et al. 2013). Genes expressed in different stages of apothecial development have been identified. Hydrophobins in maternal tissues and the mating type genes are essential for the regular development of apothecia (Terhem and van Kan 2014; Rodenburg et al. 2018).
The response of vegetative mycelia of B. cinerea to light is exceptional, and the switch to conidiation or sclerotial development can be easily monitored to identify the regulatory networks. Sclerotial development is the morphogenetic program that the fungus usually pursues (in the dark). Light prevents further sclerotial development in a competent stage by inducing the formation of aerial hyphae that develop into conidiophores and conidia. As a result, the upregulation of bcltf2 is sufficient for conidiation even in the dark (always conidia). The accumulation of vegetative mycelia (always mycelia)—optionally with aerial hyphae—suggests that the conidiation program is turned on and prevents sclerotial development but cannot be completed because of defects in conidiophore differentiation. Importantly, these regulators can be essential for full virulence. Thus, mutations of the BcVEL1-encoding gene were found to cause hyperconidiation (always conidia) and reduced virulence in wild strains T4 and 1750 (Schumacher et al. 2012, 2013). Recently, mutations in Bcin04g03490 encoding a putative transcriptional regulator with Zn2Cys6-type DNA binding and acetyltransferase domains were demonstrated to cause a mycelial morphotype and avirulence in B. cinerea field isolates (Acosta Morel et al. 2021). In these studies, mutated genes were identified in blind wild isolates. Their deletion in light-responsive B05.10, phenocopied blindness and reduced virulence, while addition of the intact genes from B05.10 rescued the phenotypes in the blind wild strains.
2.4 Metabolism: Photoprotection
Pigments are versatile secondary metabolites that protect cells from the detrimental effects of light—especially from UV-induced damage. Often, they have further beneficial functions such as the strengthening of reproduction structures or antimicrobial activities. Common in Ascomycota are the dark 1,8-dihydroxynaphthalene (DHN) melanin and the orange to yellow carotenoids. However, certain species evolved other pigment synthesis pathways leading to their characteristic pigmentations. While the genes for DHN melanogenesis and carotenogenesis are highly conserved in several species, their differential expression profiles reflect the adaptation to environmental light conditions.
Thus, fungi such as B. cinerea produce DHN melanin and carotenoids in a spatial and temporal manner while fungi in more extreme sun-exposed habitats usually exhibit constitutive melanogenesis and carotenogenesis leading to the term “black fungi” (Schumacher and Gorbushina 2020). Likewise, the designation “gray mold fungus” for B. cinerea is based on the production of DHN melanin during the latter stages of host infection resulting in plant debris covered with a layer of grayish conidiophores and conidia. The sclerotia of B. cinerea appear blackish and resemble the dense microcolonies of black fungi. DHN melanogenesis can be divided into three phases: (i) the de novo synthesis of the pentaketide T4HN (1,3,6,8-tetrahydroxynaphthalene), (ii) its modification to DHN, and (iii) the polymerization of DHN resulting in the highly complex DHN melanin (Butler and Day 1998). The genes involved in the first two phases are highly conserved in all fungi and are tightly, partially, or not clustered in the genome. In contrast, different multicopper oxidases (laccases) may carry out the cross-linking of DHN in distantly related Ascomycetes (Jia et al. 2021). Remarkably, B. cinerea possesses two different types of melanin polyketide synthases (phase I) mediating DHN melanogenesis together with a shared set of downstream-acting enzymes (phase II), and most likely again with different multicopper oxidases (phase III) in different developmental stages. BcPKS12 directly and BcPKS13 via BcYGH1 provide the precursor T4HN in either sclerotia or conidia and light-stressed mycelia; then T4HN is further converted by the same enzymes in both structures (Schumacher 2016). Melanogenesis is coupled with developmental programs because the genes involved are under the control of the respective master regulators. BcLTF2 as an activator of conidiation positively affects the expression of bcpks13 (dev LIG) and the core genes (conidial DHN melanogenesis), and bcltf2 expression is repressed by the WCC, BcLTF1, and the Velvet complex. Expression of bcpks12 and the core genes, and in turn sclerotial melanogenesis, depends on BcSMR1 (sclerotial melanin regulator) whose expression is predicted to be repressed by a yet unknown WCC-dependent LTF (Cohrs et al. 2016; Zhou et al. 2017). The upregulated expression of bcltf2, bcpks13 and the other melanogenic genes in the multicellular infection cushions suggests that the melanin is of the “conidial/BcPKS13 type” (Choquer et al. 2021). A recent study reported on the compartmentalization of melanogenic enzymes to avoid poisoning by pathway intermediates (Chen et al. 2021). Melanin is dispensable for virulence and the differentiation of the reproduction structures but contributes to their stability and longevity (Schumacher 2016; Zhu et al. 2017; Zhou et al. 2018).
B. cinerea possesses the highly conserved cluster of four genes required for carotenogenesis (bcphs1, bcphd1), retinal formation (bccao1), and green light sensing (bop2). The bifunctional BcPHS1 (phytoene synthase and lycopene cyclase) and the phytoene dehydrogenase (BcPHD1) may catalyze consecutive reactions leading to β-carotene and torulene. Retinal—the chromophore for green light absorption by opsins—can be formed by cleavage of β-carotene by the carotenoid oxygenase. The genes are induced by light and in a blue light/WCC- and BcLTF1-dependent manner (Schumacher 2017). In summary, carotenoids may help B. cinerea to resist light-induced stresses, for example, by scavenging of singlet molecular oxygen and free radicals and by protecting the integrity of the membranes. Furthermore, they represent the basis for the formation of retinal for the green light-absorbing opsins. In B. cinerea, the carotenoids are almost invisible as the faint orange pigmentation is masked by the dark melanin.
A few B. cinerea strains exhibit a remarkable red pigmentation due to the synthesis of bikaverin, a polyketide with antimicrobial properties that is typically produced by different Fusarium species (Dos Santos and Bicas 2021). The reddish B. cinerea strains possess the entire bikaverin gene cluster consisting of six genes, while grayish B. cinerea strains such as B05.10 lack the key enzyme-coding gene (Campbell et al. 2012; Schumacher et al. 2013). Bikaverin production by B. cinerea is relevant for interactions with bacteria and certain host species rather than for photoprotection (Spraker et al. 2018; Plesken et al. 2021). However, bikaverin synthesis is regulated by the VELVET complex (Schumacher et al. 2013).
B. cinerea also produces mycosporine-like amino acids (MAAs) for UV absorption and as compatibles solutes and antioxidants (Volkmann and Gorbushina 2006). Mycosporine-glutamicol—called mycosporine-2—was first isolated as a UV-absorbing compound (P310) from near-UV-treated cultures that had initiated conidiation. This compound was not detected in dark-grown vegetative cultures leading to the hypothesis that it may have a regulatory effect on conidiation (Tan and Epton 1974; Arpin et al. 1977).
The extracellular polysaccharide (EPS) matrix of B. cinerea consists mostly of cinerean [β-(1,3) (1,6)-d-glucan]. The polymer forms a capsule strongly adhering to the germinating conidia and hyphae and contributes to the adhesion to (host) surfaces, protection from light and concomitant heat and desiccation stresses, creation of a micro-environment for extracellular enzymes and as a nutrient reservoir. Thus, B. cinerea synthesizes cinerean under carbon surplus conditions and may degrade it by extracellular β-1,3 glucanases under carbon starvation conditions (Dubourdieu et al. 1981; Stahmann et al. 1992; Doss 1999).
2.5 Enzyme Systems: Photodamage Repair
The EPS matrix represents the first layer for protecting the cellular contents from environmental stresses, with the two-layered cell walls fortified with DHN melanin, and membranes with incorporated carotenoids providing second and third layers. The cellular lumen contains several primary metabolites functioning as UV screens (e.g., mycosporines), osmoprotectants (e.g., polyols, sugars, amino acids), and/or antioxidants (e.g., tocopherols, ascorbic acid, and carotenoids). Nonetheless, excessive sunlight (UV) may pass these lines of defense and cause mutagenic lesions in the DNA, disadvantageous conformational changes of proteins and the accumulation of harmful ROS. Therefore, enzymatic mechanisms must accomplish the repair of damaged DNA, misfolded proteins, and the detoxification of ROS. In fact, these genes are often upregulated upon different stresses via the stress-activated MAPK module and/or by light in a WCC-dependent fashion.
Photo-damaged DNA is repaired by the CPF protein BcCRY1 using visible light as a source of energy. The second related protein (BcCRY2) exerts a regulatory role in photomorphogenesis and does not exhibit photoreactivation activity (Cohrs and Schumacher 2017). With BcCRY1, a cyclobutane pyrimidine dimer (CPD) photolyase, B. cinerea possesses a single photolyase only. Other Leotiomycetes contain an additional (6–4)-pyrimidine-pyrimidone photoproduct (6-4PP) photolyase, and several Chaetothyriales (Eurotiomycetes) and Dothideomycetes even possess a second CPD photolyase (Schumacher and Gorbushina 2020). Expression of bccry1, bccry2, and bcuve1 encoding a conserved UV endonuclease for UVDE-dependent excision repair is induced by light in a WCC-dependent manner. Consequently, the Δbcwcl1 mutant exhibits a decreased UV tolerance (Cohrs and Schumacher 2017).
Refolding of partially denatured proteins in B. cinerea can be mediated by molecular co-chaperones including small heat shock proteins (HSPs). The expression of several genes encoding HSPs, dehydration-induced protein-like (dehydrins), and other stress-related proteins is induced upon exposure to light, as well as to temperature, osmotic and oxidative stress (Heller et al., 2012).
The oxidative stress response (OSR) system of B. cinerea comprises detoxifying enzymes, such as superoxide dismutases, catalases, peroxidases, peroxiredoxins, glutaredoxin, and thioredoxins and enzymes reducing glutathione and the thioredoxins (Viefhues et al. 2014). The corresponding genes are upregulated in response to oxidative stress by the transcriptional regulators BAP1 and BcSKN7 (Temme and Tudzynski 2009; Viefhues et al. 2015; Yang et al. 2015). The OSR system has a moderate basal activity that ensures that ROS produced as by-products of various metabolite pathways do not accumulate to toxic amounts. Simultaneous exposure to light and oxidative stress exceeds the buffer capacity, explaining the stronger effect of oxidants on B. cinerea in light compared to constant darkness. Both Δbcltf1 and Δbcwcl1 mutants affected in photoinduction of gene expression are more sensitive to oxidative stress in light (Canessa et al. 2013; Schumacher et al. 2014).
2.6 Circadian Clock: Photoentrainment
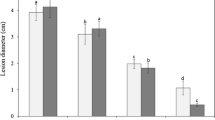
Several genes involved in coping with light-associated stress in B. cinerea are subject to photoregulation by the transcriptional WCC complex and BcLTF1 as a co-activator, as well as by the stress-activated MAP kinase BcSAK1 (Canessa et al. 2013; Schumacher et al. 2014) (unpublished). By this mechanism, the cell activates the needed stress protection measures in a timely fashion and saves resources under non-stressed conditions. Almost all organisms on earth are exposed to a 24 h day/night cycle of changing light and temperature conditions, leading to independent evolution of circadian clocks in the different kingdoms. As molecular machinery entrained by light and temperature cues, they can keep time and enable the organism to anticipate daily changing conditions and to adapt their metabolism in advance. The molecular basis of the fungal circadian clock has been intensively studied in N. crassa (Baker et al. 2012), but little is known about the presence and biological functions of the circadian clock in other fungi (Hevia et al. 2016). However, the maintenance of a clock (gain vs. loss) may reflect the relevance of light in an ecological niche. A functional circadian clock exists in B. cinerea that shares similarity with the N. crassa system, comprising the blue light-sensing WCC as the input pathway, and BcFRQ1 (frequency) as the oscillator. Bcfrq1 transcript and protein levels oscillate in constant darkness, which requires BcWCL1 as part of the WCC. Likewise, the entrainment of the clock by light requires the WCC. The output of the clock is visible on the host A. thaliana: larger lesions are formed when leaves are inoculated at dusk than at dawn which is in accordance with increased susceptibility of the host during the night. Δbcfrq1 mutants exhibit decreased conidiation accompanied by sclerotial development in the light (always sclerotia), suggesting that BcFRQ1 modulates photomorphogenesis in a nutrition-dependent manner (Canessa et al. 2013; Hevia et al. 2015).
3 Conclusion: Botrytis Exploits the Host for Nutrition and Sun Protection
B. cinerea maintains a highly sophisticated light signaling machinery that senses different light qualities to trigger a variety of responses, that are photoprotection, photomorphogenesis, positive and negative phototropism and photoentrainment. These processes render B. cinerea a valuable model to enlighten the role of light in parasitic fungus-plant interactions.
The vegetative mycelium of B. cinerea is the core of all infection and developmental programs in that it mediates the rapid colonization of dead plant tissues in terms of nectrotrophic (active killing and colonization) and saprophytic growth (colonization of already dead tissues). The vegetative mycelium is not visibly pigmented and thus considered to be sensitive to different biotic and abiotic stresses. However, the vegetative hyphae have a very limited half-life and are usually restricted to the invasive growth phase in which the hyphae are protected from light by the plant tissue. Fast colonization of fresh tissues and proper nutrient acquisition enables the rapid formation of well-protected and long-lasting reproduction structures (conidiophores with conidia and sclerotia) on the surfaces of rotted plant tissues. Depending on the light and temperature conditions, conidiation or sclerotial development is initiated for spreading of disease (summer cycle) or fungal survival (winter cycle). Taken together, B. cinerea uses the light regulatory networks to avoid light whenever possible; for example, by minimizing the half-life of sensitive cells that are hiding in plant tissues and by scheduling critical steps for the night including conidiogenesis, conidial germination, and penetration of plant tissues. Not least, the multinucleate nature of hyphae and conidia helps B. cinerea to survive in light-flooded habitats by compensating for detrimental mutations in single nuclei.
References
Acosta Morel W, Anta Fernandez F, Baroncelli R, Becerra S, Thon MR, van Kan JAL et al (2021) A major effect gene controlling development and pathogenicity in Botrytis cinerea identified through genetic analysis of natural mycelial non-pathogenic isolates. Front Plant Sci 12:663870
Akutsu K, Ko K, Misato T (1981) Role of conidial fusion in infection by Botrytis cinerea on cucumber leaves. Ann Phytopath Soc Jpn 47:15–23
Amselem J, Cuomo CA, van Kan JA, Viaud M, Benito EP, Couloux A et al (2011) Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet 7:e1002230
Antal Z, Rascle C, Cimerman A, Viaud M, Billon-Grand G, Choquer M, Bruel C (2012) The homeobox BcHOX8 gene in Botrytis cinerea regulates vegetative growth and morphology. PLoS One 7:e48134
Arbelet D, Malfatti P, Simond-Cote E, Fontaine T, Desquilbet L, Expert D et al (2010) Disruption of the Bcchs3a chitin synthase gene in Botrytis cinerea is responsible for altered adhesion and overstimulation of host plant immunity. Mol Plant-Microbe Interact 23:1324–1334
Arpin N, Favre-Bonvin J, Thivend S (1977) Structure de la mycosporine 2, nouvelle molecule, isolee de Botrytis cinerea. Tetrahedron Lett 18:819–820
Backhouse D, Willets HJ (1987) Development and structure of infection cushions of Botrytis cinerea. Trans Br Mycol Soc 89:89–95
Baker CL, Loros JJ, Dunlap JC (2012) The circadian clock of Neurospora crassa. FEMS Microbiol Rev 36:95–110
Ballaré CL (2014) Light regulation of plant defense. Annu Rev Plant Biol 65:335–363
Ballaré CL, Pierik R (2017) The shade-avoidance syndrome: multiple signals and ecological consequences. Plant Cell Environ 40:2530–2543
Blakeman JP (1975) Germination of Botrytis cinerea conidia in vitro in relation to nutrient conditions on leaf surfaces. Trans Br Mycol Soc 65:239–247
Brandhoff B, Simon A, Dornieden A, Schumacher J (2017) Regulation of conidiation in Botrytis cinerea involves the light-responsive transcriptional regulators BcLTF3 and BcREG1. Curr Genet 63:931–949
Butler MJ, Day AW (1998) Fungal melanins: a review. Can J Microbiol 44:1115–1136
Campbell MA, Rokas A, Slot JC (2012) Horizontal transfer and death of a fungal secondary metabolic gene cluster. Genome Biol Evol 4:289–293
Canessa P, Schumacher J, Hevia MA, Tudzynski P, Larrondo LF (2013) Assessing the effects of light on differentiation and virulence of the plant pathogen Botrytis cinerea: characterization of the White Collar Complex. PLoS One 8:e84223
Carlile M, Sellin MA (1963) An endogenous inhibition of spore germination in fungi. Trans Br Mycol Soc 46:15–18
Chen X, Zhu C, Na Y, Ren D, Zhang C, He Y et al (2021) Compartmentalization of melanin biosynthetic enzymes contributes to self-defense against intermediate compound scytalone in Botrytis cinerea. mBio 12
Choquer M, Fournier E, Kunz C, Levis C, Pradier JM, Simon A, Viaud M (2007) Botrytis cinerea virulence factors: new insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol Lett 277:1–10
Choquer M, Rascle C, Goncalves IR, de Vallee A, Ribot C, Loisel E et al (2021) The infection cushion of Botrytis cinerea: a fungal ‘weapon’ of plant-biomass destruction. Environ Microbiol 23:2293–2314
Cohrs KC, Schumacher J (2017) The two cryptochrome/photolyase family proteins fulfill distinct roles in DNA photorepair and regulation of conidiation in the gray mold fungus Botrytis cinerea. Appl Environ Microbiol 83:e00812–e00817
Cohrs KC, Simon A, Viaud M, Schumacher J (2016) Light governs asexual differentiation in the grey mould fungus Botrytis cinerea via the putative transcription factor BcLTF2. Environ Microbiol 18:4068–4086
Cole GT (1981) Conidiogenesis and conidiomatal ontogeny. Biol Conidial Fungi 2:271–327
Coley-Smith JR, Cooke RC (1971) Survival and germination of fungal sclerotia. Annu Rev Phytopathol 9:65–92
Coley-Smith JR, Verhoeff K, Jarvis WR (1980) The biology of botrytis. Academic Press, London
Dalmais B, Schumacher J, Moraga J, Pêcheur PLE, Tudzynski B, Collado IG, Viaud M (2011) The Botrytis cinerea phytotoxin botcinic acid requires two polyketide synthases for production and has a redundant role in virulence with botrydial. Mol Plant Pathol 12:564–579
Dasgupta A, Fuller KK, Dunlap JC, Loros JJ (2016) Seeing the world differently: variability in the photosensory mechanisms of two model fungi. Environ Microbiol 18:5–20
de Vallee A, Bally P, Bruel C, Chandat L, Choquer M, Dieryckx C et al (2019) A similar secretome disturbance as a hallmark of non-pathogenic Botrytis cinerea ATMT-mutants? Front Microbiol 10:2829
Denby KJ, Kumar P, Kliebenstein DJ (2004) Identification of Botrytis cinerea susceptibility loci in Arabidopsis thaliana. Plant J 38:473–486
Doehlemann G, Berndt P, Hahn M (2006a) Trehalose metabolism is important for heat stress tolerance and spore germination of Botrytis cinerea. Microbiology 152:2625–2634
Doehlemann G, Berndt P, Hahn M (2006b) Different signalling pathways involving a Galpha protein, cAMP and a MAP kinase control germination of Botrytis cinerea conidia. Mol Microbiol 59:821–835
Dos Santos MC, Bicas JL (2021) Natural blue pigments and bikaverin. Microbiol Res 244:126653
Doss RP (1999) Composition and enzymatic activity of the extracellular matrix secreted by germalings of Botrytis cinerea. Appl Environ Microbiol 65:404–408
Dubourdieu D, Ribereau-Gayon P, Fournet B (1981) Structure of the extracellular β-D-glucan from Botrytis cinerea. Carbohydr Res 93:294–299
El Oirdi M, El Rahman TA, Rigano L, El Hadrami A, Rodriguez MC, Daayf F et al (2011) Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant Cell 23:2405–2421
Elad Y, Williamson B, Tudzynski P, Delen N (2004) Botrytis: biology, pathology and control. Kluwer Academic Publishers, Dordrecht
Faretra F, Antonacci E (1987) Production of apothecia of Botryotinia fuckeliana (de Bary) Whetz. under controlled environmental conditions. Phytopathol Mediterr 26:29–35
Faretra F, Antonacci E, Pollastro S (1988a) Improvement of the technique used for obtaining apothecia of Botryotinia fuckeliana (Botrytis cinerea) under controlled conditions. Ann Microbiol Enzimol 38:29–40
Faretra F, Antonacci E, Pollastro S (1988b) Sexual behaviour and mating system of Botryotinia fuckeliana, teleomorph of Botrytis cinerea. J Gen Microbiol 134:2543–2550
Fillinger S, Elad Y (2016) Botrytis – the fungus, the pathogen and its management in agricultural systems. Springer
Fukumori Y, Nakajima M, Akutsu K (2004) Microconidia act the role as spermatia in the sexual reproduction of Botrytis cinerea. J Gen Plant Pathol 70:256–260
Gettkandt G (1954) Phototropism of germinating mycelia of some parasitic fungi. Scientific Journal of the Martin Luther University Halle-Wittenberg III:691–710
Giesbert S, Schumacher J, Kupas V, Espino J, Segmuller N, Haeuser-Hahn I et al (2012) Identification of pathogenesis-associated genes by T-DNA-mediated insertional mutagenesis in Botrytis cinerea: a type 2A phosphoprotein phosphatase and an SPT3 transcription factor have significant impact on virulence. Mol Plant-Microbe Interact 25:481–495
González C, Brito N, Sharon A (2016) Infection process and fungal virulence factors. In: Botrytis–the fungus, the pathogen and its management in agricultural systems. Springer, pp 229–246
Gull K, Trinci APJ (1971) Fine structure of spore germination in Botrytis cinerea. J Gen Microbiol 68:207–220
Heintzen C (2012) Plant and fungal photopigments. WIREs Membr Transp Signal 1:411–432
Heller J, Tudzynski P (2011) Reactive oxygen species in phytopathogenic fungi: signaling, development, and disease. Annu Rev Phytopathol 49:369–390
Hevia MA, Canessa P, Muller-Esparza H, Larrondo LF (2015) A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana. Proc Natl Acad Sci U S A 112:8744–8749
Hevia MA, Canessa P, Larrondo LF (2016) Circadian clocks and the regulation of virulence in fungi: getting up to speed. Semin Cell Dev Biol 57:147–155
Islam SZ, Honda Y, Sonhaji M (1998) Phototropism of conidial germ tubes of Botrytis cinerea and its implication in plant infection processes. Plant Dis 82:850–856
Jarvis W (1972) Phototropism in Botrytis cinerea. Trans Br Mycol Soc 58:526–527
Jia S-L, Chi Z, Chen L, Liu G-L, Hu Z, Chi Z-M (2021) Molecular evolution and regulation of DHN melanin-related gene clusters are closely related to adaptation of different melanin-producing fungi. Genomics 113:1962–1975
Jonkers W, Kan JAVAN, Tijm P, Lee YW, Tudzynski P, Rep M, Michielse CB (2011) The FRP1 F-box gene has different functions in sexuality, pathogenicity and metabolism in three fungal pathogens. Mol Plant Pathol 12:548–563
Kokkelink L, Minz A, Al-Masri M, Giesbert S, Barakat R, Sharon A, Tudzynski P (2011) The small GTPase BcCdc42 affects nuclear division, germination and virulence of the gray mold fungus Botrytis cinerea. Fungal Genet Biol 48:1012–1019
Kritzman G, Gilan D, Chet I (1980) Germination inhibitor in Botrytis allii spores. Phytoparasitica 8:73–76
Leisen T, Werner J, Pattar P, Ymeri E, Sommer F, Schroda M et al (2021) Multiple knockout mutants reveal a high redundancy of phytotoxic compounds that determine necrotrophic pathogenesis of Botrytis cinerea. bioRxiv
Leroch M, Kleber A, Silva E, Coenen T, Koppenhofer D, Shmaryahu A et al (2013) Transcriptome profiling of Botrytis cinerea conidial germination reveals upregulation of infection-related genes during the prepenetration stage. Eukaryot Cell 12:614–626
Leroch M, Mueller N, Hinsenkamp I, Hahn M (2015) The signalling mucin Msb2 regulates surface sensing and host penetration via BMP1 MAP kinase signalling in Botrytis cinerea. Mol Plant Pathol 16:787–798
Liu W, Leroux P, Fillinger S (2008) The HOG1-like MAP kinase Sak1 of Botrytis cinerea is negatively regulated by the upstream histidine kinase Bos1 and is not involved in dicarboximide- and phenylpyrrole-resistance. Fungal Genet Biol 45:1062–1074
Liu W, Soulie MC, Perrino C, Fillinger S (2011) The osmosensing signal transduction pathway from Botrytis cinerea regulates cell wall integrity and MAP kinase pathways control melanin biosynthesis with influence of light. Fungal Genet Biol 48:377–387
Liu Y, Jafari F, Wang H (2021) Integration of light and hormone signaling pathways in the regulation of plant shade avoidance syndrome. aBIOTECH 2:131–145
Losi A, Gärtner W (2021) A light life together: photosensing in the plant microbiota. Photochem Photobiol Sci 20:451–473
Marschall R, Siegmund U, Burbank J, Tudzynski P (2016) Update on Nox function, site of action and regulation in Botrytis cinerea. Fungal Biol Biotechnol 3:8
Mengiste T (2012) Plant immunity to necrotrophs. Annu Rev Phytopathol 50:267–294
Mengiste T, Laluk K, AbuQamar S (2009) Mechanisms of induced resistance against Botrytis cinerea. In: Postharvest pathology. Springer, pp 13–30
Michielse CB, Becker M, Heller J, Moraga J, Collado IG, Tudzynski P (2011) The Botrytis cinerea Reg1 protein, a putative transcriptional regulator, is required for pathogenicity, conidiogenesis, and the production of secondary metabolites. Mol Plant-Microbe Interact 24:1074–1085
Müller N, Leroch M, Schumacher J, Zimmer D, Könnel A, Klug K et al (2018) Investigations on VELVET regulatory mutants confirm the role of host tissue acidification and secretion of proteins in the pathogenesis of Botrytis cinerea. New Phytol 219:1062–1074
Plesken C, Pattar P, Reiss B, Noor ZN, Zhang L, Klug K et al (2021) Genetic diversity of Botrytis cinerea revealed by multilocus sequencing, and identification of B. cinerea populations showing genetic isolation and distinct host adaptation. Front Plant Sci 12:663027
Porquier A, Tisserant C, Salinas F, Glassl C, Wange L, Enard W et al (2021) Retrotransposons as pathogenicity factors of the plant pathogenic fungus Botrytis cinerea. Genome Biol 22:225
Prusky D, Alkan N, Mengiste T, Fluhr R (2013) Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu Rev Phytopathol 51:155–176
Ren W, Liu N, Yang Y, Yang Q, Chen C, Gao Q (2019) The sensor proteins BcSho1 and BcSln1 are involved in, though not essential to, vegetative differentiation, pathogenicity and osmotic stress tolerance in Botrytis cinerea. Front Microbiol 10:328
Robert-Seilaniantz A, Navarro L, Bari R, Jones JD (2007) Pathological hormone imbalances. Curr Opin Plant Biol 10:372–379
Robinson W (1914) Some experiments on the effect of external stimuli on the sporidia of Puccinia malvacearum (Mont.). Ann Bot XXVIII:331–340
Roca MG, Weichert M, Siegmund U, Tudzynski P, Fleissner A (2012) Germling fusion via conidial anastomosis tubes in the grey mould Botrytis cinerea requires NADPH oxidase activity. Fungal Biol 116:379–387
Rodenburg SY, Terhem RB, Veloso J, Stassen JH, van Kan JA (2018) Functional analysis of mating type genes and transcriptome analysis during fruiting body development of Botrytis cinerea. mBio 9:01939–01917
Rosero-Hernandez ED, Echeverri FL (2020) The search for quorum sensing in Botrytis cinerea: regulatory activity of its extracts on its development. Plants (Basel) 9
Rossi FR, Gárriz A, Marina M, Romero FM, Gonzalez ME, Collado IG, Pieckenstain FL (2011) The sesquiterpene botrydial produced by Botrytis cinerea induces the hypersensitive response on plant tissues and its action is modulated by salicylic acid and jasmonic acid signaling. Mol Plant-Microbe Interact 24:888–896
Rowe HC, Kliebenstein DJ (2008) Complex genetics control natural variation in Arabidopsis thaliana resistance to Botrytis cinerea. Genetics 180:2237–2250
Rui O, Hahn M (2007) The Slt2-type MAP kinase Bmp3 of Botrytis cinerea is required for normal saprotrophic growth, conidiation, plant surface sensing and host tissue colonization. Mol Plant Pathol 8:173–184
Schamber A, Leroch M, Diwo J, Mendgen K, Hahn M (2010) The role of mitogen-activated protein (MAP) kinase signalling components and the Ste12 transcription factor in germination and pathogenicity of Botrytis cinerea. Mol Plant Pathol 11:105–119
Schumacher J (2016) DHN melanin biosynthesis in the plant pathogenic fungus Botrytis cinerea is based on two developmentally regulated key enzyme (PKS)-encoding genes. Mol Microbiol 99:729–748
Schumacher J (2017) How light affects the life of Botrytis. Fungal Genet Biol 106:26–41
Schumacher J, Gorbushina AA (2020) Light sensing in plant- and rock-associated black fungi. Fungal Biol 124:407–417
Schumacher J, de Larrinoa IF, Tudzynski B (2008a) Calcineurin-responsive zinc finger transcription factor CRZ1 of Botrytis cinerea is required for growth, development, and full virulence on bean plants. Eukaryot Cell 7:584–601
Schumacher J, Kokkelink L, Huesmann C, Jimenez-Teja D, Collado IG, Barakat R et al (2008b) The cAMP-dependent signaling pathway and its role in conidial germination, growth, and virulence of the gray mold Botrytis cinerea. Mol Plant-Microbe Interact 21:1443–1459
Schumacher J, Pradier JM, Simon A, Traeger S, Moraga J, Collado IG et al (2012) Natural variation in the VELVET gene bcvel1 affects virulence and light-dependent differentiation in Botrytis cinerea. PLoS One 7:e47840
Schumacher J, Gautier A, Morgant G, Studt L, Ducrot PH, Le Pecheur P et al (2013) A functional bikaverin biosynthesis gene cluster in rare strains of Botrytis cinerea is positively controlled by VELVET. PLoS One 8:e53729
Schumacher J, Simon A, Cohrs KC, Viaud M, Tudzynski P (2014) The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLoS Genet 10:e1004040
Schumacher J, Simon A, Cohrs KC, Traeger S, Porquier A, Dalmais B et al (2015) The VELVET complex in the gray mold fungus Botrytis cinerea: impact of BcLAE1 on differentiation, secondary metabolism, and virulence. Mol Plant-Microbe Interact 28:659–674
Schumacher J, Studt L, Tudzynski P (2019) The putative H3K36 demethylase BcKDM1 affects virulence, stress responses and photomorphogenesis in Botrytis cinerea. Fungal Genet Biol 123:14–24
Segmüller N, Ellendorf U, Tudzynski B, Tudzynski P (2007) BcSAK1, a stress-activated mitogen-activated protein kinase, is involved in vegetative differentiation and pathogenicity in Botrytis cinerea. Eukaryot Cell 6:211–221
Segmüller N, Kokkelink L, Giesbert S, Odinius D, van Kan J, Tudzynski P (2008) NADPH oxidases are involved in differentiation and pathogenicity in Botrytis cinerea. Mol Plant-Microbe Interact 21:808–819
Sharon A, Elad Y, Barakat R, Tudzynski P (2004) Phytohormones in Botrytis-plant interactions. In: Botrytis: biology, pathology and control. Springer, pp 163–179
Shaw MW, Emmanuel CJ, Emilda D, Terhem RB, Shafia A, Tsamaidi D et al (2016) Analysis of cryptic, systemic Botrytis infections in symptomless hosts. Front Plant Sci 7:625
Siegmund U, Heller J, van Kan JA, Tudzynski P (2013) The NADPH oxidase complexes in Botrytis cinerea: evidence for a close association with the ER and the tetraspanin Pls1. PLoS One 8:e55879
Siegmund U, Marschall R, Tudzynski P (2015) BcNoxD, a putative ER protein, is a new component of the NADPH oxidase complex in Botrytis cinerea. Mol Microbiol 95:988–1005
Spraker JE, Wiemann P, Baccile JA, Venkatesh N, Schumacher J, Schroeder FC et al (2018) Conserved responses in a war of small molecules between a plant-pathogenic bacterium and fungi. mBio 9:e00820–00818
Stahmann KP, Pielken P, Schimz KL, Sahm H (1992) Degradation of extracellular beta-(1,3)(1,6)-D-Glucan by Botrytis cinerea. Appl Environ Microbiol 58:3347–3354
Suzuki Y, Kumagai T, Oda Y (1977) Locus of blue and near ultraviolet reversible photoreaction in the stages of conidial development in Botrytis cinerea. J Gen Microbiol 98:199–204
Tan KK (1975) Interaction of near-ultraviolet, blue, red, and far-red light in sporulation of Botrytis cinerea. Trans Br Mycol Soc 64:215–222
Tan K, Epton H (1974) Ultraviolet-absorbing compounds associated with sporulation in Botrytis cinerea. Trans Br Mycol Soc 63:157–167
Tang J, Wu M, Zhang J, Li G, Yang L (2021) Botrytis cinerea G protein beta subunit Bcgb1 controls growth, development and virulence by regulating cAMP signaling and MAPK sgnaling. J Fungi (Basel) 7
Temme N, Tudzynski P (2009) Does Botrytis cinerea ignore H(2)O(2)-induced oxidative stress during infection? Characterization of Botrytis activator protein 1. Mol Plant-Microbe Interact 22:987–998
Temme N, Oeser B, Massaroli M, Heller J, Simon A, Collado IG et al (2012) BcAtf1, a global regulator, controls various differentiation processes and phytotoxin production in Botrytis cinerea. Mol Plant Pathol 13:704–718
Terhem RB, van Kan JA (2014) Functional analysis of hydrophobin genes in sexual development of Botrytis cinerea. Fungal Genet Biol 71:42–51
van Kan JA (2006) Licensed to kill: the lifestyle of a necrotrophic plant pathogen. Trends Plant Sci 11:247–253
Veloso J, van Kan JAL (2018) Many shades of grey in Botrytis—host plant interactions. Trends Plant Sci
Viaud M, Schumacher J, Porquier A, Simon A (2016) Regulation of secondary metabolism in the gray mold fungus Botrytis cinerea. Host-Pathogen Interaction: Microbial Metabolism, Pathogenicity and Antiinfectives 201–216
Viefhues A, Heller J, Temme N, Tudzynski P (2014) Redox systems in Botrytis cinerea: impact on development and virulence. Mol Plant-Microbe Interact 27:858–874
Viefhues A, Schlathoelter I, Simon A, Viaud M, Tudzynski P (2015) Unraveling the function of the response regulator BcSkn7 in the stress sgnaling network of Botrytis cinerea. Eukaryot Cell 14:636–651
Volkmann M, Gorbushina AA (2006) A broadly applicable method for extraction and characterization of mycosporines and mycosporine-like amino acids of terrestrial, marine and freshwater origin. FEMS Microbiol Lett 255:286–295
Wang M, Weiberg A, Lin F-M, Thomma BP, Huang H-D, Jin H (2016) Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat Plants 2:16151
Wang Y, Yin Y, Wu S, Chui C, Ma T, Jiang H et al (2018) The MAPK kinase BcMkk1 suppresses oxalic acid biosynthesis via impeding phosphorylation of BcRim15 by BcSch9 in Botrytis cinerea. PLoS Pathog 14:e1007285
Weiberg A, Wang M, Lin F-M, Zhao H, Zhang Z, Kaloshian I et al (2013) Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 342:118–123
Williamson B, Tudzynski B, Tudzynski P, van Kan JA (2007) Botrytis cinerea: the cause of grey mould disease. Mol Plant Pathol 8:561–580
Yan L, Yang Q, Sundin GW, Li H, Ma Z (2010) The mitogen-activated protein kinase kinase BOS5 is involved in regulating vegetative differentiation and virulence in Botrytis cinerea. Fungal Genet Biol 47:753–760
Yang Q, Yan L, Gu Q, Ma Z (2012) The mitogen-activated protein kinase kinase kinase BcOs4 is required for vegetative differentiation and pathogenicity in Botrytis cinerea. Appl Microbiol Biotechnol 96:481–492
Yang Q, Chen Y, Ma Z (2013) Involvement of BcVeA and BcVelB in regulating conidiation, pigmentation and virulence in Botrytis cinerea. Fungal Genet Biol 50:63–71
Yang Q, Yin D, Yin Y, Cao Y, Ma Z (2015) The response regulator BcSkn7 is required for vegetative differentiation and adaptation to oxidative and osmotic stresses in Botrytis cinerea. Mol Plant Pathol 16:276–287
Zhang Z, Li H, Qin G, He C, Li B, Tian S (2016) The MADS-Box transcription factor Bcmads1 is required for growth, sclerotia production and pathogenicity of Botrytis cinerea. Sci Rep 6:33901
Zheng L, Campbell M, Murphy J, Lam S, Xu JR (2000) The BMP1 gene is essential for pathogenicity in the gray mold fungus Botrytis cinerea. Mol Plant-Microbe Interact 13:724–732
Zhou Y, Yang L, Wu M, Chen W, Li G, Zhang J (2017) A single-nucleotide deletion in the transcription factor gene bcsmr1 causes sclerotial-melanogenesis deficiency in Botrytis cinerea. Front Microbiol 8
Zhou Y, Li N, Yang J, Yang L, Wu M, Chen W et al (2018) Contrast between orange- and black-colored sclerotial isolates of Botrytis cinerea: melanogenesis and ecological fitness. Plant Dis 102:428–436
Zhu P, Zhang C, Xiao H, Wang Y, Toyoda H, Xu L (2013) Exploitable regulatory effects of light on growth and development of Botrytis cinerea. J Plant Pathol 95:509–517
Zhu P, Li Q, Zhang C, Na Y, Xu L (2017) Bcpks12 gene inactivation substantiates biological functions of sclerotium melanization in Botrytis cinerea. Physiol Mol Plant Pathol 98:80–84
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Schumacher, J. (2023). Role of Light in the Life Cycle of Botrytis cinerea. In: Scott, B., Mesarich, C. (eds) Plant Relationships. The Mycota, vol 5. Springer, Cham. https://doi.org/10.1007/978-3-031-16503-0_14
Download citation
DOI: https://doi.org/10.1007/978-3-031-16503-0_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-16502-3
Online ISBN: 978-3-031-16503-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)