
Abstract
Increasingly, consumers are paying attention to healthier food diets, “healthy” food attributes (such as “freshness”, “naturalness” and “nutritional value”), and the overall sustainability of production and processing methods. Other significant trends include a growing demand for regional and locally produced/supplied and less processed food. To meet these demands, food production and processing need to evolve to preserve the raw material and natural food properties while ensuring such sustenance is healthy, tasty, and sustainable. In parallel, it is necessary to understand the influence of consumers’ practices in maintaining the beneficial food attributes from purchasing to consumption. The whole supply chain must be resilient, fair, diverse, transparent, and economically balanced to make different food systems sustainable. This chapter focuses on the role of dynamic value chains using biodiverse, underutilised crops to improve food system resilience and deliver foods with good nutritional and health properties while ensuring low environmental impacts, and resilient ecosystem functions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
There are about 50,000 edible plants on the Earth; however, current food systems are concentrated on only three: wheat, maize, and rice (Khoury et al., 2014). These species provide more than 50% of the plant-based calories consumed by the world’s population and occupy 40% of the world’s arable land. The lack of agricultural diversity has severe consequences on biodiversity and global environmental sustainability, namely soil degradation and higher global emissions (FAO, 2010a). Therefore, it is vital to stimulate the cultivation of less common species, known by “underutilised crops” (UCs) and enhance awareness of, plus improve where necessary, their nutritional and environmental proprieties. Here we describe how this can be achieved via integrating UCs in value chains while realising their benefits using a “system function approach”.
Agri-food systems comprise actors and activities involved in the production, processing, distribution, consumption, and disposal of food products (FAO, 2021). By 2050, food demand is projected to increase by 60% relative to 2005. However, this projection is highly sensitive to, among other things, consumption patterns (diets), distribution, and levels of food waste (FAO, 2018b; Hunter et al., 2017). By 2067 the population is expected to reach 10.4 billion, with Africa and Asia accounting for 81% of this growth (Britt et al., 2018). Meanwhile, there is increasing pressure on agriculture and the broader land sector to deliver food, feed, fibre, fuel, bio-based materials, and ecosystem services – including nature-based solutions to climate change (Huppmann et al., 2018). Rapid cuts in greenhouse gas (GHG) emissions, alongside adaptation to a changing climate, are essential to maintain food system viability, let alone sustainability. Food systems account for 21–37% of global anthropogenic GHG emissions (Masson-Delmotte et al., 2019). On current trajectories, these emissions alone (excluding other industry, transport, and building sources) could exceed Paris Agreement targets for climate stabilisation (Clark et al., 2020). Similarly, food systems are key drivers of “Planetary Boundaries” exceedances across land use, biodiversity loss, and nutrient cycling (Steffen et al., 2015). Livestock production dominates many of these impacts (Foley et al., 2011; Rogelj et al., 2018; Vermeulen et al., 2012; Eshel et al., 2014; FAO, 2018a; Steinfeld et al., 2006). Up to now, food system intensification has been highly successful at delivering more output per unit of land (Burney et al., 2010). This has helped reducing GHG emission intensity per unit of output and sustain increasing levels of consumption to the point where a larger share of global population is obese rather than under-nourished (Benton & Bailey, 2019). To meet increasing food demands, focus during since the 60’s was on intensification of agricultural systems, characterized by low crop diversity and large use of chemical inputs. Together with the implementation of low-input agronomic practices, crop diversification is highlighted today as a key issue for future sustainable development of agroecosystems valorising natural and cultivated biodiversity for agricultural purposes (Stagnari et al., 2017), resulting in greater ecosystem services and resilience (Springmann et al., 2018). Going beyond the recent focus on efficiency to deliver more food, fibre, and fuel at a dramatically lower aggregate environmental cost and with resilience is a massive challenge – necessitating transformative change beyond the incremental improvement of business-as-usual (Fanzo et al., 2021). The effective transformation will require integration of demand-side measures (e.g., reduced consumption of livestock products) alongside reconfiguration of value chains. This, in turn, will deliver both food and value-added more fairly, changing primary production to provide food and a plethora of ecosystem services, while preserving large areas of land for nature-based solutions (Fanzo et al., 2021; IPCC, 2019; Willett et al., 2019).
Sustainable diets can be defined as diets with a low environmental impact that contribute to food and nutrition security, and health in the present and future generations. They are “protective and respectful of biodiversity and ecosystems, culturally acceptable, accessible, economically fair and affordable; nutritionally adequate, safe and healthy; while optimising natural and human resources” (FAO, 2010a). Consumers are increasingly aware of the impact of their choices and behaviours on environmental sustainability. Accordingly, they are looking for demonstrations of sustainability commitment from food industries along the entire product life cycle. These include the interest in the origin of ingredients, higher demand for locally sourced products, and clean labels. For instance, food trends for 2022 are in line with those observed in the last years, looking ahead to an increase in the demand for plant-based and alternative proteins, better ingredients, and functional foods and beverages (Mintel, 2021).
This chapter explores the prospects of better integration of UCs in value chains and presents how their multiple benefits can be realised in a systems function approach. We show the current status of underutilised crops from cultivation to various uses and benefits. Then we turn to challenges in the value chain from farm to fork. Finally, we introduce policies that could improve investment in underutilised crops.
2 The Role of Underutilised Crops
2.1 What Are Underutilised Crops?
Staple crops currently dominate agricultural production and global food supplies. Diets around the globe are becoming more standardised, relying on very few crops or crop varieties– together with high intakes of sugar and oil. With staple crops’ technological and policy investments, these new dietary habits have led to the neglect of about 7000 plant species documented as human food (FAO, 2010a). These neglected, underutilised, minor, or orphan crops are locally adapted to challenging environments, domesticated by local communities, and require lower inputs than staple crops, but are characterised by having low agricultural production and no perceived economic importance in advanced economies (Ebert, 2014). There are many possible definitions for a UC, and taking into account the FAO’s work on this topic, these can be defined as ‘a neglected, but valuable species, landrace, variety, or cultivar that has limited current use in a given geographic, social, and economic context and that holds great promise to diversify agricultural systems, create resilient agroecosystems, diversify diets, and create economically viable dynamic value chains (for feed, food, and non-food uses)’.
UCs are therefore considered key to sustainable food security in the future, as scientists have been discussing their role in climate change mitigation and potential for exploitation for years (Padulosi et al., 2011). Given their more rustic nature, they contribute to agroecological resilience through system diversification and are an important component of the culture and diets of specific regions of the world (Massawe et al., 2016). However, they are underexploited and under-conserved resources, with minimal research and investment in their development (FAO, 2010b). For example, UC seed conservation is mainly made by gene banks, complemented by family or local seed networks (Padulosi et al., 2011). Current factors hindering the broader incorporation of UCs in food systems have been reviewed. They include their lower productivity and yield potential, lack of trait improvement and processing technology, limited market availability, lower cooking quality, and lack of knowledge both at the cultivation and at the consumption level (Bekkering & Tian, 2019; Hunter et al., 2019; Saini et al., 2021).
General examples of UCs include millets, roots and tubers, pulse crops, fruits and vegetables, and tree nuts (FAO, 2010b). These are traditionally grown for food, fibre, fodder, oil, or medicinal value (Ebert, 2014). Although these potential uses are acknowledged, further exploitation is yet to be developed due to these crops’ semi-domesticated and neglected nature (Murthy & Bapat, 2020).
Millets, such as pearl millet (Pennisetum glaucum), foxtail millet (Setaria italica), barnyard millet (Echinochloa spp.), little millet (Panicum miliar), kodo millet (Paspalum scrobiculatum), finger millet (Eleusine coracana), are small-grained C4 cereals with a rich nutritional profile and many of them resilient to drought and high temperatures (Saini et al., 2021).
There are also many C3 underutilized cereals emmer (Triticum dicoccum), einkorn (T. monococcum), spelt (T. spelta), or rye (Secale cereale). Other UCs such as amaranth (Amaranthus spp.), buckwheat (Fagopyrum esculentum), chia (Salvia hispanica), or quinoa (Chenopodium quinoa), are considered pseudocereals because despite having seeds resembling the cereal grains, do not belong to Poaceae. The other large family containing many UCs is the Fabaceae, including many species of interest for forage or pasture, but also many others whose seeds are valued for food and feed. These species are known as grain legumes or pulses, and include good examples of warm-season pulses like winged bean (Psophocarpus tetragonolobus), horse gram (Macrotyloma uniflorum), lima bean (Phaseolus lunatus), hyacinth bean (Lablab purpureus), cowpea (Vigna unguiculata), mung bean (Vigna radiata) or barbara groundnut (Vigna subterranea), and of cool-season ones like faba bean (Vicia faba), grass pea (Lathyrus sativus) or lupin (Lupinus albus), among many others (Rubiales et al., 2021).
Underutilised roots and tubers are represented mainly by taro (Colocasia esculenta), yam (Dioscorea sp.), ulluco (Ullucus tuberosus), yautia (Xanthosoma sagittifolium), arrowroot (Maranta arundinaceae), and giant swamp taro (Cyrtosperma paeonifolius), and sweet potato (Ipomoea batatas) (FAO, 2010b; Li et al., 2020b). These crops are essential food on small-holder farms in marginal rural areas where they can be the primary source of nutrients during periods of food scarcity (Siddique et al., 2021).
Fruits and vegetables, such as wild melon (Citrullus lanatus), wild mustard (Sinapis arvensis), jackfruit (Artocarpus heterophyllus), mangosteen (Garcinia mangostana), African eggplant (Solanum aethiopicum), or grape (Vitis spp.) (FAO, 2010b; Massawe et al., 2016) are essential foods to support small-scale farmers and serve as the primary source of nourishment in underdeveloped countries (Siddique et al., 2021). Hence, these underutilised horticultural crops are vital in supporting nutritional security and avoiding malnutrition and hidden hunger issues with respect to the lack of specific vitamins and micronutrients (Nandal & Bhardwaj, 2014). Unfortunately, most tree nuts are also considered underutilised, as little or no research has been dedicated to their development. Some examples include cashew nut (Anacardium occidentale), Brazil nut (Bertholletia excelsa), chestnut (Castanea sativa), acorns (Quercus nuts), and tiger nut (Cyperus esculentus) (Asare et al., 2020; Murthy & Bapat, 2020). Their importance has been put forward in the latest diet recommendations due to their high contents in bioactive compounds and their biological activity (Willett et al., 2019). The harvested area and production levels of some examples of UCs mentioned above are listed in Table 9.1, although is still challenging to trace UCs production numbers on a worldwide scale. However, for the crops where numbers are available, increasing UCs agricultural production needs further investment in technology and research. This could be focused on their beneficial impacts and resilience in semi-arid and arid areas and their adaptation to different climatic scenarios. Additionally, when looking at the countries with higher levels of UCs production (Table 9.1), food systems benefit from UCs incorporation and adaptation.
2.2 Preservation of Biodiversity
Biodiversity and ecosystems are the very foundations of human existence and contribute to human well-being in three fundamental ways: through the production of goods (food, fibres, water, air, medicines, and recreational spaces); provision of services (cultural, religious, aesthetic, and spiritual); and the processes that balance and regulate the above (pollination, prevention of soil erosion, microclimate control and nutrient cycling and transfer) (Buiatti et al., 2010). Cultural and natural biodiversity that include thousands of UCs are the basis of agrobiodiversity, which is preserved, alike nutrition and health, by traditional farming practices and cultural identities. These practices also make long-term sustainable use of natural resources and the environment, increasing productivity and ensuring food security and sovereignty (Buiatti et al., 2010). Unfortunately, however, biodiversity and agrobiodiversity are in a state of decline worldwide, and with it, the inclusion of UCs in local agri-food systems. The key factors contributing to the loss of biodiversity include unsustainable farming, fishing, and forest practices which lead, among other things, to natural resource consumption, habitat loss and fragmentation, soil deterioration, water and atmospheric pollution, and genetic pollution (MEA, 2005). Moreover, global climate change threatens biodiversity by altering habitats and modifying the equilibria of crucial species. At the same time, the narrow spectrum of products traded from agriculture, forestry, and fisheries make ecosystems increasingly vulnerable (FAO, 2019a).
When considering genetic resources for food and agriculture (GRFA), we refer to crop diversity created by man (FAO, 1999). It underpins agriculture’s productivity, resilience, and adaptive capacity and is an integral part of people’s cultural identity (IAAKSTD, 2009). Given that they supply most of the food for human consumption, they are fundamental for creating sustainable agriculture and food safety. Yet we are losing them at an alarming rate. Since agriculture began to develop about 15,000 years ago, it is estimated that around 10,000 species have been used for human food. Currently, no more than 120 cultivated species provide 90% of the human food supplied by plants. Only four plant species (potatoes, rice, maize, and wheat) and three animal species (cattle, pigs, and chickens) provide more than 50% of all human food. The degree of biodiversity in agroecosystems depends on: (i) variety of vegetation inside and around the agroecosystem; (ii) length of different crops; (iii) intensity of management; and (iv) degree of isolation from wild vegetation (Altieri, 1999).
The best way to conserve traditionally cultivated plants, including UCs, and raised animals, is to utilise them. Two distinctive conservation methods of UCs can be identified as in-situ (and on farms) and ex-situ practices. The first is carried out in conditions that allow a natural and continuous evolution and co-adaptation through cultivation or breeding. In contrast, the second entails protecting endangered species and genetic resources (plant varieties and animal breeds) outside their natural habitat, for example, by preserving seeds in a germplasm bank. All conservation measures should be planned and implemented on a scale determined by ecological and social criteria, focusing on densely populated areas, and protected natural areas. It is interesting to note that the leading cause of the loss of Genetic Resources for Food and Agriculture (GRFA) would appear to be underutilisation as opposed to overexploitation. Given the high interdependency of countries on GRFA, international cooperation in this area is not an option but a must. This cooperation has led to intergovernmental negotiations and the adoption of the legally binding International Treaty for Plant Genetic Resources for Food and Agriculture (ITPGRFA). The objectives of the ITPGRFA (FAO, 2001) are the conservation and sustainable use of all plant genetic resources for food and agriculture and the fair and equitable sharing of the benefits arising out of their use, in harmony with the Convention on Biological Diversity, for sustainable agriculture and food security.
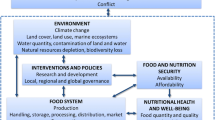
Wider cultivation of UCs would help to deliver on the pledge to diversify agricultural systems, create resilient agroecosystems, diversify diets, and develop economically viable dynamic value chains for feed, food, and non-food uses (Fig. 9.1) (Bavec et al., 2017; Gregory et al., 2019). Therefore, characterising their nutritional and health attributes is essential to promote their wider adoption amongst populations.

The role of underutilised crops (UCs) in the preservation of biodiversity. (Modified from Gregory et al., 2019)
2.3 Health and Nutritional Benefits of UCs
Recent data suggests that ending world hunger and malnutrition in all its forms is becoming increasingly more challenging, particularly exacerbated by the COVID-19 pandemic (Lowe, 2021). Indeed, the latest international reports highlight concerns regarding the world’s ability to meet the sustainable development agenda by 2030, including the Zero Hunger target (FAO, 2021). Around 118 million more people were facing hunger in 2020 compared to 2019, representing 768 million undernourished people worldwide (FAO, 2021). Limited access to a healthy, balanced, diverse, and nutritious diet, particularly in low- and middle-income countries, contributes to this scenario (Li et al., 2020a, b). In 2020, nearly 1 in 3 people, around 2.37 billion people in the world, did not have access to adequate food, 40% of which or almost 928 million, faced severe levels of food insecurity (FAO, 2021). Simultaneously, overweight and obesity rates continuously spread worldwide as technological developments push societies towards more sedentary lifestyles with easier access to highly caloric but nutritionally poor and highly processed foods and beverages (Popkin et al., 2020). Hence, it is estimated that multiple micronutrient deficiencies, frequently underlying a “hidden hunger” phenomenon, affect more than 2 billion people globally (von Grebmer et al., 2014). For instance, of the 7 billion world’s population, more than 1.6 billion and more than 200 million people suffer from iron deficiency and vitamin A insufficiency, respectively (Li et al., 2020a, b). Such nutrient deficits can not only impair human health but may ultimately hamper socioeconomic development (Ibeanu et al., 2020).
Although a shortage in food supply to adequately feed the growing world’s population has become a critical reality, it is advocated that current food systems should shift from the quantity-oriented mindset to a more quality-focused approach. This will allow exploration of new and more sustainable means to nourish populations (Hunter et al., 2019). The investment in UCs seems required to restore sustainable agriculture practices and address the global food challenges (Li & Siddique, 2020). These crops represent a local, affordable, sustainable, and culturally acceptable way to improve diversity in food supply systems and, therefore, access to nutrient-dense foods. Yet, many countries fail to recognise their rightful value (Hunter et al., 2019). Among various socio-economic and political reasons, the over-reliance on more profitable high-yielding monocultures has caused the marginalisation of minor crops, including primarily wild or semi-domesticated crops (Li et al., 2020a, b). However, data suggests that these crops often provide greater levels of essential nutrients in comparison to current major staple crops, including vitamin C, vitamin A, iron, calcium, and fibre (Hunter et al., 2019), which aligns with the most frequent limiting dietary micronutrients (Lowe, 2021). For example, an indigenous Brazilian fruit, camu-camu (Myrciaria dubia), has been found to contain 40 times more vitamin C than the typical orange (Citrus sinensis) (Hunter et al., 2019). Like pearl millet (P. glaucum), traditional crops in Pakistan and Nepal possess higher amounts of iron, zinc, riboflavin, and folic acid than rice, maize, and wheat (Adhikari et al., 2017). These three staple crops provide more than 50% of the world’s plant-derived calories (Dulloo et al., 2016). Noteworthy, many of these crops are native in the poorest world’s regions characterised by nutrient-deficient and health-impaired individuals (FAO, 2019a). In Kenya, locally grown leafy greens, such as amaranth (A. dubius), were introduced into school meals in an attempt to mitigate undernutrition since they have been shown to possess almost 3.5 times more vitamin A (beta-carotene equivalent) and 6 times more iron than the ordinary cabbage (Brassica oleracea) (Hunter et al., 2019). Inter and intra-species differences regarding the nutritional composition of UCs have been reported across the literature and justify the need for further research regarding the health potential of these foods (Hunter et al., 2019). Also, challenges are still present when considering the processing associated with including UCs in food product development.
2.4 Integration of UCs in Food production and Processing
It is vital to provide populations with diverse and nurturing foods to keep them healthy without damaging the environment (Willett et al., 2019). As the global population keeps growing, it presses for the intensification of the current food system, causing environmental impact to increase beyond sustainable levels (Poore & Nemecek, 2018). The highest impact of food production comes from raw materials sourcing (Poore & Nemecek, 2018; Willett et al., 2019). Therefore, many environmental problems can be easily traced back to this point: from deforestation to the desertification of arable land to lixiviation, loss in biodiversity and others (Mentis, 2020; Zhao et al., 2015).
Farming has a considerable environmental impact that could be lowered. Intensification leads to higher yields per given area and higher resource consumption (fuel, irrigation, fertilisers, and pesticides). Sustainable farming can help lower the impact from these inputs (Commission on Sustainable Agriculture and Climate Change, 2012) and growing quality plant proteins tackles many different sustainability points (e.g., agrobiodiversity and potentially avoiding high-impact animal-protein production). As mentioned above, presently, only a few crops are responsible for almost 50% of global food intake (FAO, 2018b). This becomes a problem when the repetitive growth of the same cultures reduces soil biodiversity and depletes nutrients beyond natural replenishment rates (Zhao et al., 2015). This then leads to an increasing need for synthetic fertilisers that cause additional damage to the environment, as explained previously. Increasing the consumption of different proteins is often suggested to diversify diets. Multiple studies show these alternative proteins and developed products have a lower impact (Smetana et al., 2015, 2021; Tello et al., 2021). Increasing the production and consumption of UCs can also decrease the environmental strain of the repetitive growth of crops.
Crop replacement is not easy since their cultivation is adapted to the different areas where they are grown. One possibility that can help with this is the revalorization of crops adapted to those areas, but that lost competitiveness. These crops, however, may need to be improved to increase their competitiveness and lead to agricultural diversification and reduction of risks (e.g., pest attacks that can destroy entire cultures; Popp et al., 2013). Another possibility could be the introduction on new UCs, their adaptation needing testing and probably further improvement. Still, the use of these crops can lead to novel product development, and this, in return, revalorize the crops.
The development of these novel products should consider consumer trends with more fresh-like attributes and long shelf-life (Palou et al., 2020). Due to this, implementation of novel technologies such as pulsed electric fields, high-pressure processing, and high-pressure homogenisation or ultrasound, can provide an interesting starting point. All these technologies can provide potential solutions to the pressing challenge of sustainability (Matthews et al., 2019). The plant protein industry shows many advantages and strengths over the animal protein industry, as shown by Petrusán et al. (2016). These advantages of plant proteins could underpin the success of integrating UC-derived products through marketing and certification strategies that support their broader commercialisation. As a result of the increasing demand of local vegetable protein food, both in traditional uses and in novel processed food business (Cusworth et al., 2021) legume cultivation is speedily recovering.
2.5 Consumers, Cultural Barriers, and Leverages
Most of the research to increase the consumption of UCs has been from the supply side (Cheng et al., 2017; Dawson et al., 2009; Mayes et al., 2012). These have focused on highlighting their nutritional and environmental properties to justify the additional effort in improving the characteristics of those crops (e.g., yields, agronomic properties, environmental impact). Understanding consumer knowledge, acceptance, and preferences for UCs are essential in enhancing their consumption levels to increase micronutrient intake.
A barrier to higher adoption rates of UCs as a staple food is the rise of convenience foods and modern consumption patterns. In particular, consumers in developing countries are increasingly abandoning the traditional diets that these crops are part of and are replacing them with western diets (Cordain et al., 2005). Likewise, in industrialized countries, many recipes and products have fallen into oblivion in the last century, partly due to the change in direction to a society where meat is the dominant food (Holm & Møhl, 2000). For example, in Germany, the consumption of legumes decreased from 20.7 kg in 1850 to 3.0 kg per capita and year in 2017 (BLE, n.d.; Teuteberg, 2006). In this context, many legumes varieties became extinct or have been forgotten, e.g., lentils from Swabian Alb (Reif et al., 2021). Moreover, consuming such traditional UCs often requires know-how, i.e., how to prepare and cook them (sometimes depending on the stage of maturity), and perhaps even knowing which cultivar (variety) is more desirable for a particular use. These knowledge gaps often render UCs non-competitive against well-known and globally consumed staples such as rice, wheat, maize, soybean, and potatoes.
There are several approaches to increase the consumption of underutilised crops. One way is to convey knowledge about such food products and their preparation. Activities like the Bavarian specialty database (https://www.spezialitaetenland-bayern.de/spezialitaeten) or the Slow Food ‘Ark of Taste’ (https://www.fondazioneslowfood.com/it/arca-del-gusto-slow-food) try to preserve and to promote the knowledge and to create consumer awareness. Through such measures, consumer preferences for traditional specialties (Profeta et al., 2007) and authentic foods (Wirsig et al., n.d.) can be addressed and triggered. Furthermore, consumers are becoming increasingly conscious of their food basket’s health and nutritional profile (Profeta, 2019). The tendency is to avoid chemicals and synthetic foods and preference for nutrition through foods that bring “natural” attributes. In this context, many UCs have advantages compared to staple foods, as outlined in the chapters before. In this situation, marketing communication to the final consumer should highlight UCs’ special health and environmental characteristics.
Looking at new ways of incorporating UCs into consumers’ diets requires creativity. UCs could easily fit into a modern lifestyle by adding value and creating ready-made convenience products. Finished convenience products do not need consumers to prepare or cook the corresponding UC. Thus, by finding novel and innovative methods to organise, sell and consume UCs, consumers can discover more diverse ways to enjoy this nutritional and culturally relevant food source. Also, due to the dominant role of taste in consumers’ purchase decisions, there is the need to bring UCs closer to consumers’ preferences. In this way, the value chain for UCs will get sustainable economic, environmental, food security, and nutritional benefits.
2.6 The Role of Markets, Labelling, and Certification
Many UCs are locally popular crops, are nutritionally superior, they generate income, are resistant to drought, they conserve natural resources, are tasty and delicious, are necessary for climate adaptation, and often have long culinary traditions. Still, they continue to be marginalised by research and undervalued by development (Eyzaguirre et al., 1999). Most importantly, market factors are responsible for rendering these crops underutilised; consequently, UCs become unable to meet the global market requirements, industrialised agri-techniques, and uniformity standards. Similarly, the policy is also often divisive, even “food discriminatory”, and this explains why UCs are undervalued and underinvested (Chishakwe, 2008). Furthermore, increasing their value for more comprehensive production and commerce depends on research-intensive activities. These crops are mostly not suitable for cultivation or cannot meet uniformity standards, often due to genetic erosion. UCs are niche-specific, versatile, and differ substantially from mainstream crop value chains. Breeding programs, seed multiplication, collective actions of value chain actors are indispensable for market development (Stamp et al., 2012). Increasing the consumption of UCs requires not only systemic demand-markets development, but more expansive capacity building in the value chain. Any attempt at commercialising UCs requires demand expansion, increased supply, marketing channels efficiency, and a supply control mechanism (Gruere et al., 2008). Scarano et al. (2021) identified several research-intensive factors that could raise awareness of and fully realise the benefits of UCs. These include research on the genetic traits linked to the climate adaptation; characterisation of main nutrient classes and their biosynthesis pathways; quantification and characterisation of the main antimetabolic factors/antinutrients; and understanding biological activities in the prevention of human diseases. Finally, any research on UCs needs to benefit from the full participation in exploration and action learning of value chain actors in a participatory setting (Vernooy, 2021). In sum, value addition would be a high potential for UCs in a diverse and sustainable food system only if more significant investment in research and development becomes more available.
Consumers are increasingly interested in local, traditional, or sustainably produced fruits, vegetables, or arable crops. This provides an excellent premise to label such products to make consumers aware of unique product qualities, taste, shape, and colour (Wirsig et al., 2011). However, there is no label for UCs in the food market. Nonetheless, at least in the European Union, there are different food quality labels as, e.g., PDO (Protected Designation of Origin), PGI (Protected Geographical Indication), and TSG (Traditional Specialties Guaranteed), or Protected Mountain Products. These allow covering aspects of crop diversity or seed origin (Benner et al., 2008; Profeta et al., 2006). According to this scheme, many underutilised food products, e.g., Bamberger Hörnla or Alho da Graciosa (Berbereia, 2015), are protected and profit from marketing campaigns promoting the EU quality system. A recent case study from Germany shows the positive effects of this official labelling scheme for such products (Chilla et al., 2020). Since the EU regulation even allows applications for PDO, PGI, and TSG from third countries, there is a legal labelling framework that nearly all countries can use. Despite this, the existing regulation was not specially developed for underutilised groups. Such a long and complicated application process is too great a task for small-producer groups. In the next revision of the EU regulation, the unique requirements and needs of producers of UCs should be considered to improve their access to the existing scheme.
2.7 The Non-food Uses for Underutilised Crops
Crops are most likely underutilised when their potential is unknown, or their availability is not sufficient to establish an economically feasible utilisation. This is particularly challenging in rural areas where long distances need to be bridged to harvest, treat, and utilise biomass. Nevertheless, there are economic opportunities, and most UCs possess a high potential to serve as a source of food and non-food products. A combination of both uses may foster the cultivation of UCs. As with UCs considered for food use, the prospect of UCs for non-food use can be assessed based on the biochemical components such as lipid, carbohydrate, and protein contents. Depending on the composition, tailor-made harvesting and utilisation approaches can be developed, allowing the implementation of a biorefinery and the generation of products and services even in rural areas.
An example of a successful new crop is late-harvested grass, usually cut in autumn at natural conservation areas. While fresh grass has been considered as feedstock in green biorefineries or as feed, late-harvested grass utilisation is still at the early stage. The biochemical composition is the reason for the different utilisation intensities of fresh and late-harvested grasses. On a dry matter basis (w/w), grass can contain 20–30% cellulose, 15–25% hemicellulose, 3–10% lignin, 6–25% protein, 1–2.5% fat, 1–2.5% starch, and 5–20% ash (Grass, 2004). Fresh grass is rich in proteins and is easier to digest. Contrarily, matured grass contains less metabolisable energy, for instance, due to a reduced degradable protein content (Bovolenta et al., 2008; Waramit et al., 2012; Boob et al., 2019; Koidou et al., 2019) and reduced nutrient contents such as P, N, and K (Bokdam & Wallis de Vries, 1992; Mládek et al., 2011; Schlegel et al., 2016; Boob et al., 2019; Koidou et al., 2019).
Even though the protein content is comparably low, protein extraction can be worthwhile. About 30–60% (w/w) of the original protein can be recovered by mechanical pressing or alkaline extraction (Bals et al., 2012; Hermansen et al., 2017). The highest value arises from the fibres present in the grass. After mechanical pressing, up to 95% (w/w) of the fibres remain in the press cake (O’Keeffe et al., 2011) and can be used as a feedstock in pulp and paper production (Finell, 2003), for biocomposites (Biowert, 2021), or building materials (King et al., 2013).
Although late-harvested grass has been investigated as a substrate for combustion (Tonn et al., 2010; Lewandowski et al., 2003), pyrolysis (Wilson et al., 2013; Mos et al., 2013) or as lignocellulosic feedstock in fermentation (Dien et al., 2018; Jungers et al., 2013) a biorefinery that operates purely on late-harvested grass is currently not working. As mentioned above, the challenges are the availability of biomass as late-harvested grass appears once, maximum twice per year. However, the availability of biomass and the services that the biomass delivers during its cultivation stage should be considered. Late-harvested grass is vital to conserving biodiversity and storing carbon in the soil. Thus, the use of late-harvested grass can be an example where ecosystem services are preserved, and the potential of the biomass is simultaneously utilised.
2.8 Environmental Benefits of UCs
Modern crop varieties deliver reliable and high yields, but the widespread adoption of monocultures in intensive agriculture often leads to environmental depletion and higher chemical inputs. Most of the cereal crops that dominate global production, such as wheat, rice, and maize, require an increased water supply and have low adaptive resilience to water shortage, raising concerns about their suitability to under the forecasted scenarios of more frequent and severe droughts (Mueller et al., 2012). Pesticides and herbicides target harmful organisms that can harm or compete with crops. Still, they can also reach animals and plants beyond the seemingly restricted area of their application. For example, several pesticides are harmful to bees and other insects, limiting their ability to pollinate crops and other plants (Uhl & Brühl, 2019). Phytosanitary products also impair soil microorganisms involved in carbon and nitrogen cycling, contributing to climate change. Highly disturbed soils with low microbial biodiversity quickly lose carbon to waterways and the atmosphere, propelling the accumulation of greenhouse gases in the atmosphere (Lazcano et al., 2021). This will ultimately lead to warming temperatures and extreme weather events, further impairing plant and soil communities and favouring the adaptation of invasive species that disrupt native ecosystems (Diffenbaugh et al., 2008). Intensive agriculture is also largely reliant on nitrogen fertilisers, which can run off into waterways, decrease the available oxygen in the water and cause eutrophication of both fresh and saltwater ecosystems, making them uninhabitable for aquatic organisms (Huang et al., 2017).
In the forthcoming decades, food systems are estimated to have an increasing environmental impact by intensifying global ecological pressures and destabilising key ecosystem processes, fostering climate change (Springmann et al., 2018). On the other hand, climate change will also pose challenges to ecosystems worldwide, as plants will have to endure in drier, saltier soils (Onyekachi et al., 2019). UCs are typically native to the environments in which they are grown, thus requiring fewer external and economic inputs than conventional crops. They can show adaptation to dryland cropping systems, high water use efficiency, and short growing seasons while delivering similar yields to major cereal crops (Karunaratne et al., 2015). They can also prosper in harsh environments and poor soils by fixing carbon from the atmosphere and nitrogen in the ground, offering opportunities for nutrient use efficiency and lowering global GHG emissions (Mabhaudhi et al., 2019). Developing powerful sustainable and bio-based agronomic strategies for crop nutrition, irrigation, soil fertility, and stress tolerance could allow a significant reduction in the use of chemical fertilisers and water for agriculture (Karkanis et al., 2018; Karavidas et al., 2022). They can also improve environmental resilience and quality of crops (Rivero et al., 2022; Dubey et al., 2020). Moreover, integrating neglected landraces, ecotypes, and varieties with increased nutrient use efficiency, water use efficiency, and stress tolerance into such farming systems could help in this direction (Dwivedi et al., 2016; Rivero et al., 2022). Specifically, UCs can contribute to environmental resilience and in mitigating climate change by the following means:
-
(a)
Delivering tolerance to drought, salt, and toxic metals stress, as is the case of several cultivars of tomato, chickpea, barley, rice, wheat, and sunflower that possess specific genes involved in abiotic-stress tolerance (Mammadov et al., 2018 and references therein; Kumar Rai et al., 2021);
-
(b)
Improving water use efficiency, as they can grow as a dryland crop without supplemental irrigation (e.g., millets), as well as by improving water quality (e.g., winged bean); (Kamel et al., 2018 and references therein);
-
(c)
Fostering biodiversity and beneficial wild animals, thus promoting resilience against pests and diseases as part of integrated pest management, as detailed above (Villegas-Fernández et al., 2011; Sardana et al., 2017; Mammadov et al., 2018 and references therein);
-
(d)
Decreasing the need for inputs and supporting natural carbon and nitrogen cycles, particularly concerning legumes that promote the accumulation of nitrogen in the soil while capturing carbon from the atmosphere (Mabhaudhi et al., 2019 and references therein);
-
(e)
Reducing the high environmental impact of large-scale food and feed production and consumption worldwide by creating shorter value chains and decreasing transportation burdens (Weinberger & Swai, 2006; Will, 2008; Imathiu, 2021).
The exploitation of UCs as part of a holistic transformation of food systems plays a pivotal role in environmental sustainability (Haddad et al., 2016). Table 9.2 showcases the environmental and ecosystem services provided by distinct UCs that can lever the security of the global food supply while ensuring the sustainable use of environmental resources. Figure 9.2 illustrates the multiple benefits of UCs that go beyond the farm level.

Illustration of the diversity of benefits delivered by underutilised crops (UCs), from the farm to the final consumer
2.9 Genetics and Breeding of UCs
Being minor crops, there has been a lag in the overall genetic improvement of UCs due to limited investment compared to major crops. Applicable breeding methods are the same that could be used for any crop, from classical selection, to genomic assisted-breeding, being the availability of resources and the targets what makes the difference. Breeding more adapted and productive cultivars, thus meeting producer and consumer needs, enables a wider adoption in the value chain. However, when the surfaces are limited, the return of the breeding activity is not sufficient to support strong breeding programs. The agroecological transition requires not only greater UCs cultivation but also different cultivation approches, such as intercropping, organic, etc., each one requiring specific breeding strategies. Greater adaptation to low input conditions will be a leading priority in UC breeding, particularly for organic systems. The global change and increasing instability of the climate pose additional challenges to breeders, emphasizing a need for greater nutrient use efficieny and greater tolerance to major abiotic stresses (Rubiales et al., 2021). The need to improve pest and disease resistance will be increasingly critical with the mandatory decrease in pesticide uses and with the expected effects of climatic change on the geographic distribution and frequency of epidemic outbreaks (Skendžić et al., 2021). Also, consumer preferences are affecting breeding priorities in terms of quality, that used to focus mainly on improving protein yield and reducing “undesirable” compounds contents, currently demanding increasing attention to important sensory or processing traits (Vaz Patto et al., 2015; Mecha et al., 2021).
Breeding relies on genetic diversity, and, for this, collection and conservation of genetic resourses is crucial. The breeding of elite cultivars of any crop tends to focus on selected germplams, progressively reducing the genetic diversity in the given species used in agriculture. This would be easily remedied by pre-breeding, with infusion of genetic diversity coming from landraces, ecotypes, or wild relative. However, UCs breeders have to cope with ever-increasing quantitative target traits with modest budgets, being often forced to focus on short-term breeding goals, preventing the needed exploitation of valuable germplasm that would require lengthy pre-breeding (Dwivedi et al., 2016; Rubiales et al., 2015). There are already excellent global collections in which wild and cultivated (e.g., landraces, old varieties) accessions of most crop species, including most UCs, are effectively stored, multiplied, and shared (EURISCO, 2022; GENESYS, 2022). However, a real limitation for effective use in breeding is the insufficient characterization (phenotypic and genotypic data) of these stored accessions.
Despite the modest investment made on UCs, significant advances were made in biotechnology and genomics over the last two decades, with funded initiatives and web resources available (Gregory et al., 2019; Jamnadass et al., 2020; Rubiales et al., 2021). This offers great opportunities to adapt to UCs advanced tools already used form major crops, such as whole-genome and transcriptome sequencing, genomic selection, genome editing and speed breeding (Kamenya et al., 2021). In fact, the list of UCs with their genomes sequenced is rapidly growing (see tables in Kamenya et al., 2021; Rubiales et al., 2021), and any case, with the dropping of sequencing costs, most of UCs will likely have their genomes sequenced in the next decade. Still, when no whole-genome secuence is yet available, comparative genomics could be exploited alongside other tools enabling single nucleotide polymorphisms (SNP) calling. For instance, Diversity Arrays Technology (DArT) has been successfully used for genetic characterization and mapping in many UCs. More recently DArT-sequencing (DArT-seq) or other restriction-associated DNA sequencing (RADseq) genotyping methods, including genotyping-by-sequencing, are being used for rapid marker discovery in many UCs (as reviewed by Kamenya et al., 2021; Rubiales et al., 2021). Mapping studies in biparental populations and genome-wide association studies (GWAS) are being used to identify markers that explain trait variation in a chosen population. Also, monogenic traits can be exploited by marker assisted selection. However, most agronomically important traits are polygenic, thus genomic selection could help to incorporate small-effect loci into prediction equations. Genomic selection has potential for UC breeding, enhancing selection efficiency once prediction equations are available (Annicchiarico et al., 2020). To develop these prediction equations not only Next Generation Sequencing (NGS) genotyping data are needed, but, most importantly, good phenotypic data. Field phenotyping remains a bottleneck for crop genetic improvement. Therefore, affordable low-cost phenotyping tools are needed to decrease the cost of field evaluations (Araus et al., 2018).
2.10 Agronomic Challenges of UC Cultivation
Growing UCs sometimes comes with agronomic challenges. As for any crop, UCs’ demand for nutrients is not constant during the growth period, as nutrient availability is affected by environmental factors such as soil type and climate (Havlin, 2020). Therefore, to better utilise UCs, the supply and demand of fertiliser can be synchronised by fine-tuning its application to the needs of such crops, and thus the input be significantly reduced without compromising yield (Shah & Wu, 2019; Gatsios et al., 2021a, b). The loss of nutrients from the soil can also be appreciably reduced by the use of new intelligent fertilisers, such as nano-fertilisers, slow-release fertilisers, fertilisers enriched with nitrification inhibitors, compost, and microbial biostimulants such as arbuscular mycorrhiza fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) (Mejias et al., 2021; Rouphael & Colla, 2020a, b; Ghafoor et al., 2021; Alonso-Ayuso et al., 2016; Cristofano et al., 2021; Sabatino et al., 2020). Applying such integrated nutrient management (INM) strategies in UC cultivation could enhance nutrient use efficiency (Shah & Wu, 2019.). Similarly, grafting onto nitrogen-efficient rootstocks can also lead to reduced nitrogen application (Liang et al., 2021). Some UCs could also be used as rootstocks, the wild relatives of cultivated crops (Razi et al., 2021). Introducing these crops to innovative farming practices spanning from the agroecological (integrated, organic, conservation) to high controlled technology (soilless culture, vertical farming) could improve their performance. Importantly, it could also lead to a measurable increase in farm income (Savvas & Gruda, 2018; Gatsios et al., 2021a, b; van Delden et al., 2021).
Organic crop production is facing the challenge of the yield gap due to nitrogen shortage availability at critical growth stages (Ponisio et al., 2015; Birkhofer et al., 2016). Identifying elite and UC genotypes suitable for low-input farming systems may also reduce the yield gap (Ntatsi et al., 2018a, b; Anastasi et al., 2019; Ronga et al., 2021). Taking also into consideration that organic farming relies on the inclusion of legumes as green manure, or in the rotation, due to the contribution of significant quantities of atmospheric nitrogen (N2) (Gatsios et al., 2019, 2021a, c), the need to use legumes with high biological nitrogen fixation (BNF) ability are imperative for enhancing nitrogen inputs to the soil, thereby improving crop yield (Ntatsi et al., 2018a, b).
In addition, due to climate change, choosing the appropriate tillage system is extremely important. The adoption of conservation tillage systems (e.g., reduced tillage or no-tillage) can make a significant contribution to the reduction of greenhouse gas emissions due to a decrease in fuel consumption and lowered soil mineralisation rates (Stošić et al., 2021). Also, several studies show that conservation tillage systems improve soil properties (e.g., soil organic matter and water storage) and increases crop yields (Li et al., 2020a, b; Dong et al., 2021). Thus, for all the reasons mentioned above, it is essential to evaluate the effects of tillage systems on the growth and yield of UCs.
Another important limiting factor in UC cultivation is their competition with weeds and the lack of registered herbicides integrated into weed management programs. Thus, weed control is mainly based on hand hoeing and mechanical equipment. As in other “minor crops,” these species should be planted in rows at distances to allow natural weed control (Karkanis et al., 2022). An appropriate design of the crop rotation system can also make a significant contribution to weed management (Kanatas, 2020; Shahzad et al., 2021). Ideally, this should be done using a ‘Theory of Change’ approach where the system’s long-term and robust (stable) functional capacities determine the degree to which a system is resilient. All these obstacles and opportunities pave the way for developing new agri-food systems, including UCs. There is a need to implement a ‘Theory of Change’ approach where food system actors are included in the process of problem identification and solving, using true multi-actor approaches. The views and knowledge from breeders, farmers, chefs, consumers, food retailers, scientists, food/non-food industry and civil society in general need to be integrated to strengthen the evidence base of UCs multiple dimensions of value. This ‘Theory of Change’ approach for UCs will help also to identify the governance and policy frameworks needed for effective implementation of UCs in food and non-food value chains and ensure that agrobiodiversity is used sustainably.
3 Integrating UCs Using a Theory of Change Approach
Resilience can be defined as the maintenance of system functions in the face of stress from biotic or abiotic perturbations, whether gradual or sudden. The functional capacities of an ecological system are determined by interactions between biotic and abiotic components and the influence of specific pedoclimate, biogeography, land-use or -management approaches, socio-economic- and -technical aspects on the resulting ecosystem processes. Systems comprising a balance of functions tend to be more stable due to internal regulation of specific essential processes, or “system-function indicators”, such as primary production, nutrient-, carbon- and water cycling, etc. These system-function indicators should be selected and monitored at the relevant spatial scale, such as field, farm, catchment, or bioregion. These system function indicators can also serve as a measure of system resilience where acceptable upper- and lower-thresholds can be defined.
3.1 Defining Better Farming System Functions with UCs
What constitutes a well-functioning and resilient farming system depends upon the ecological interactions at field- farm- and catchment-scales. Better farming operations can be defined as those which maintain a balance of all the essential farmed habitat functions such that they maintain stable levels over time, in response to shocks, and with minimal inputs from outside the system – since external inputs present dependencies, and therefore a risk.
-
(a)
System functional indicators can be divided into biotic, abiotic, and socioeconomic categories. Biotic indicators include crop productivity and yield qualities, non-crop vegetational diversity, and the diversity and functional composition of trophic groups of microbes and invertebrates. These are organisms responsible for ecological processes needed to maintain system functions of soil and water quality, nutrient cycling, primary productivity, pollination, and the trophic and competitive interactions driving population regulation. Abiotic indicators relate to soil physical structure and environmental pollution (greenhouse gas emissions, leaching, and erosion). Socioeconomic indicators include cost-benefit analysis at the farm business scale and social aspects (employment, countryside access, etc.) beyond the farm gate, depending on the system boundaries (Hawes et al., 2009, 2016)
-
(b)
Stability is defined here as a fluctuation within the upper- and lower limits or thresholds, which will vary depending on the environment and desired system states. Resilience is then determined by the system’s capacity (farm) to keep within these thresholds over time and is the speed at which the system returns to a stable state following a disturbance. Resilience is strongly influenced by diversity and by a system feature called “functional redundancy” or “compensating complementarity”. The similarity in functional role between species allows those functions to be maintained in the face of species extinctions (Ehrlich & Ehrlich, 1981), i.e., where numerous species possess a specific ecosystem function, the loss of one or few can be compensated by the others present in the system. In this way, system function is not compromised by such loss(es). Sufficient diversity accommodates functional redundancy and is an insurance measure for protection against shocks, as may occur due to management or climate (Yachi & Loreau, 1999).
-
(c)
Minimal inputs should be the defining feature of well-functioning production systems that are semi “closed” (Hadavi & Ghazijahani, 2018), i.e., reduce reliance on external inputs by enhancing resource use efficiency and introducing nature-based solutions, minimising pollution and diversity loss, and so maintain stable functioning. However, fully closed systems are not entirely possible at the field-farm scale since harvested material must be removed for consumption. Therefore, offtake or loss from the system must be replaced to maintain stable states. Consequently, maintaining productivity (offtake) demands renewable and sustainably (and preferably locally) sourced inputs. Suppose the offtake is consumed locally and sourced from the same region. In that case, the system could be considered “closed” within a more comprehensive spatial boundary (i.e., bioregion) than the literal confines of the farm-scale management unit. Furthermore, reducing reliance on external inputs requires that resource use efficiency is optimised. This can be accomplished through agronomy (e.g., precision fertiliser placement in time and space precise targeting of crop protection chemicals through forecasting and mapping technology), plant diversity (e.g., niche complementarity giving rise to complete utilisation of inputs and selection of varieties to optimise resource capture and pest and disease resistance), soil biophysical function (e.g., microbial and invertebrate communities for nutrient turnover and optimal rooting for uptake efficiency), and non-crop biodiversity (e.g., alternative resources to support pollinator and natural enemy populations). Finally, interventions such as minimum tillage, cover cropping, riparian buffers, field margins, and fertiliser injection can be used to help “close the loop” by minimising inputs losses through erosion, leaching, and GHG emissions.
3.2 Implementing the System-Functions Approach
In the contexts defined above, management of production systems for the needs, or “health” of the environment, society, and economics requires optimisation across system functions. There will be inevitable trade-offs, at least in the short term, e.g., productivity/profit versus diversity/ecological functions, until the long-term benefits of more sustainable approaches can be achieved. As such, decisions need to be agreed on what system functions (health states) are desired or/and are to be prioritised. It is then necessary that: key indicators are identified for the desired functions; that upper- and lower-thresholds of acceptability in these functions are determined; and that they can be practically applied at different scales (e.g., field-farm-catchment-region-national). Finally, questions are raised at each step, and decisions need to be made for successful design and implementation, as illustrated in the conceptual model (Fig. 9.3).

A schematic flow-diagram illustrating the main steps involved in implementing the Ecosystem Function Approach. The approach is socioecological and demands the involvement of cooperative communities of stakeholders from across the value chain at appropriate spatial scales – from “system baselining” to “scaling-up and –out” of the approach. The pivotal importance of the interactive and cooperative socio-ecological approach is highlighted by the facilitative communities and capacities necessary to underpin the success of the process
There are various national and global environmental impact accounting tools, especially concerning GHG emissions. However, such inventories present data at national levels. Moreover, they do not dissect the detail of landscape structures and land-uses at levels related with confidence to ecosystem functions. So, the Ecosystem Function Approach has not yet been achieved for conventional farming systems, let alone those using less common agronomic strategies or underutilised crops.
Even with an agreed indicator set and using accredited, open, and transparent monitoring- and accounting- strategies, we still face the challenge of how monitoring approaches can be effectively taken up? Successful implementation of the system function approach requires an objective assessment of impact through accurate baselining and subsequent monitoring of the effect of any change in management intervention. Monitoring needs to be sensitive enough to detect trends over time. Land managers can ensure (and prove) that their interventions result in a move in the right direction towards the set target. Traditionally, agri-environment schemes have been incentivised through payments based on implementing a specified management intervention (length of hedgerow planted, area of cover crop sown, etc.). Still, these schemes suffer from a lack of evidence for any subsequent ecosystem function benefit. They frequently fall short of their original goal (biodiversity gain, species conservation, etc.) (Hawes et al., 2016). An alternative in the form of outcomes-based monitoring allows a proper assessment of impact, the opportunity for iterative development of improved management, and incentive payment is based on the extent to which the goals have been met. However, this approach requires indicator monitoring protocols that are quick, inexpensive, and easy to carry out by the land managers themselves while providing sufficiently accurate data that can detect trends in the right (or wrong) direction. Some examples are currently being piloted by the James Hutton Institute and NatureScot with farmers across Scotland and Ireland, focusing on biodiversity conservation for specific habitats and species. There is a need to test the quality and objectivity of data collected and then extend these specific protocols to more generally applicable assessments of farmland system functioning.
Research and innovation to realise validated approaches of system function accounting are rare and generally restricted to field-farm scales, which are also experimental sites, rather than in commercial farm settings. Across Europe, there are only a minimal number of long-term experimental platforms, mainly in arable stages, and livestock and perennial orchard systems, offering well-developed frameworks of indicators to the main system elements. However, such long-term farm-monitoring platforms appear absent for even major cropped systems and critical commercial species, such as tomatoes (Quesada et al., 2019; Tran et al., 2021). Also, there are no known life cycle analysis (LCA) studies of such platforms’ ex-farmgate impacts (or functions). Additionally, it may be that a typical synthesis of system function indicators could be achieved even from the existing platforms, however limited. It may be possible to identify that sub-set of system-function indicators that are relatively easy for farmers to carry out themselves. Automated, high throughput technological solutions for monitoring system function – such as satellite imagery, other remote methods, and molecular diagnostics – also have the potential to bolster farmer-led data collection. Nonetheless, these will require a significant increase in research and development support before ground-truth testing and subsequent roll out.
3.3 Novel AI Methods for Integrating UCs in Sustainable Food production Systems
The transition to sustainable agriculture with UCs requires simultaneously considering the questions “How much food needs to be produced?” and “How will this food be produced?”. So far, the agri-food sector has failed to address this challenge comprehensively and successfully. A promising approach to this challenge is the introduction of sustainable agriculture (Piñeiro et al., 2020; Rocchi et al., 2020). It is becoming increasingly clear that the transition to sustainable agriculture is impossible without using modern information technologies and artificial intelligence (AI) methods. With their help, the discovery of synergistic links between environmental conditions, biodiversity, and food production has been dramatically accelerated, enabling the adoption of sustainable agriculture. Cropland is no longer considered a basic input for food production, but a complex dynamic agri-ecosystem managed based on cognitive approaches. This means constantly monitoring its condition and maintaining a stable balance between “how much” and “how” by flexible management decisions. Artificial intelligence has become a new tool with which agriculture successfully introduces new principles and criteria for sustainable food production (Liakos et al., 2018).
Artificial intelligence is the computer science of complex dynamic systems that help extract information from large amounts of data, research already carried out, and experts’ experience and knowledge (e.g., agronomists, pedologists, entomologists) (Russel & Norvig, 2021). The information gathered in this way is integrated into knowledge structures that help us understand, predict, and manage complex dynamic systems such as sustainable food production. This type of research approach allows us to acquire new knowledge very quickly and design scenarios for an efficient transition to sustainable multifunctional agriculture. Artificial intelligence, therefore, plays a critical role in the development of modern decision support systems for sustainable food production (Zhai et al., 2020).
One such system that illustrates the use of artificial intelligence for assessment of the sustainability of agri-food chains, including legumes as the target UC is the PATHFINDER (http://pathfinder.ijs.si/) Decision Support System (DSS) (Fig. 9.4). The system assesses the sustainability and its pillars (environmental, economic, social) of both the individual links and the chain as a whole. If the user wants to improve sustainability, the DSS finds and suggests changes to enhance sustainability or its unique sustainability pillars of the whole agri-food chain. With the help of artificial intelligence methods, a system like this can be further developed and upgraded to consider dynamic agri-food chains that would introduce, promote and strengthen the role of UCs in the agri-food chain.

Landing page of the PATHFINDER web-based DSS (http://pathfinder.ijs.si/)
Artificial intelligence is a very effective new tool to build advanced decision support systems that enable qualitative and quantitative breakthroughs in agriculture. With its help, we can make responsible decisions about measures to achieve the Sustainable Development Goals in general and sustainable food production in particular.
3.4 Social and Economic Considerations
In this section, the various opportunities and challenges of UCs are discussed from a socio-economic perspective. Figure 9.5 presents an overview of findings from the literature at the farm level, the potential impacts for the local community, and value chain aspects. We conceptualise farmers as both community and value chain actors; the value chain is embedded within the community but goes beyond (e.g., remote consumers). The community includes both actors directly involved in the value chain and indirectly impacted members.

Challenges (+) and opportunities (−) of UCs found in the literature: aspects of underutilized crops with empirical evidence described in the paper itself are above the dashed line, while aspects without empirical evidence are below. References referring to potential effects of UC in a hypothetical manner are marked with (?). a Guida et al., 2017; b Siracusa et al., 2013; c Galmes et al. 2011; d Padulosi et al., 2002; e Padulosi et al. 2013; f FAO, 2010b; g Padulosi et al., 1999; h Altieri & Merrick, 1987; i (Baker & Russell, 2017; j Baldermann et al., 2016; k Burgess, 1994; l Camacho-Henriquez et al., 2016; m Karunaratne et al., 2015; n Mabhaudhi et al., 2016; o Murevanhema & Jideani, 2013; p Nandal and Bhardwaj, 2014; q Nganga, 2014; r Traoré et al., 2020; s Van Oosterom et al., 2002; t Vijayalakshmi et al., 2010; u Will, 2008
The literature used is composed of scientific articles with experiments (Guida et al., 2017; Siracusa et al., 2013; Galmes et al., 2011; Karunaratne et al., 2015; Nandal & Bhardwaj, 2014; Van Oosterom et al., 2002), review articles (Mabhaudhi et al., 2016; Murevanhema & Jideani, 2013; Nandal & Bhardwaj, 2014), case studies (Baker & Russell, 2017), book chapters (Padulosi et al., 2002; Camacho-Henriquez et al., 2016) and reports (FAO, 2010b). The cases studies reported are from all continents (e.g., potato landrace in Peru (Camacho-Henriquez et al., 2016), pearl millet or sorghum in Burkina Faso (Camacho-Henriquez et al., 2016), tomato landraces in Italy (Guida et al., 2017; Siracusa et al., 2013), wheat landraces in the United States (Baker & Russell, 2017), and finger millet in India (Vijayalakshmi et al., 2010), etc.). Yet, a lot of unknowns remain regarding the European context. The studies compare situations before and after UC introduction initiatives (e.g., Vijayalakshmi et al., 2010) or reach the characteristics of UC towards their mainstream equivalents through quantitative analysis, for example, in terms of yields (e.g., Traoré et al., 2020; Van Oosterom et al., 2002). Studies also discuss the advantages and issues of using UCs (e.g., Baldermann et al., 2016; Burgess, 1994) or present detailed case studies of UCs (e.g., (Baker & Russell, 2017; Camacho-Henriquez et al., 2016; Nandal & Bhardwaj, 2014).
Aspects that are the most recurrent in the literature are the low level of external inputs needed in the farming systems due to UCs good adaptation to their local context (Guida et al., 2017; Galmes et al., 2011; FAO, 2010b; Padulosi et al., 1999; Altieri & Merrick, 1987; (Baker & Russell, 2017; Baldermann et al., 2016; Burgess et al., 1994; Camacho-Henriquez et al., 2016; Karunaratne et al., 2015; Mabhaudhi et al., 2016; Traoré et al., 2020; Vijayalakshmi et al., 2010). There are, however, difficulties in implementing and managing local value chains that can be competitive with mainstream crops, such as the need to access plant materials (Padulosi et al., 2002; Altieri & Merrick, 1987), to connect with other VC actors (e.g., consumers) (Padulosi et al., 2002; FAO, 2010b; Baker & Russell, 2017; Baldermann et al., 2016; Mabhaudhi et al., 2016), to create partnerships (Camacho-Henriquez et al., 2016; Murevanhema et al., 2013), and to find necessary structures, infrastructures, funds, knowledge, etc. (Padulosi et al., 2002; Baker & Russell, 2017; Baldermann et al., 2016).
3.4.1 Scotland as a Socio-economic Case Study
The Scottish socioeconomic paradigm and its impact on the Scottish arable system are particular, with barley occupying around two-thirds of cultivated arable land annually. Moreover, this barley is apportioned 2:1 for animal feed and malting markets, respectively (Scottish Government, 2021). In particular, the high demand for Scottish whisky, a heritage product, facilitates economic security mainly via a global trade which accounts for 75% of the total value of Scottish drink and food exports (21% of all of the UK), and high tax revenue for the government which is are currently estimated at £5.5bn in Gross Value Added (GVA) (Scottish Whisky Association, 2021), and this is doubled when beer and other spirits are taken into consideration. Nevertheless: how can crop systems be diversified using UCs while maintaining the commercial success of the whisky sector?
An exemplary Scottish farm (www.arbikie.com) took the approach of developing a short value chain which they termed their ‘field to bottle’ approach, and which encompasses five key elements (attractive location, traditional ethos, master craftsmen, small scale, and very close proximity of crop-production and distilling-elements), and the offer of products whose provenance and environmental credentials are fully traceable. Arbikie developed rye as a forgotten Scottish crop to diversify their cropped system and reintroduce Scottish Whisky made from rye after a 150-year absence. Arbikie’s approach is now allied to the use of intercropping, the use of heritage barley types, and adoption of under-cultivated crops, including pea (Pisum sativum), used to produce the world’s first climate-positive gin and vodka (Lienhardt et al., 2019a, b) known as the Nàdar Collection (Arbikie Distillery, 2021). Other smaller production units have advertised their products along with similar principles, advertising their products based on their whole value chain (e.g., Nc’nean Distillery, 2021).
A defining feature here is that sustainability matters have emerged as the language of modern marketing. Additionally, the increasing awareness among consumers of “greenwashing” (Chen & Chang, 2013) has meant that the importance of any sustainability-related marketing claim should be evidenced. This requires open access to and transparent data about the claims made and value chain operations and processes (Beulens et al., 2005). Additionally, independent agencies should validate evidence using methods and procedures that are also approved, accredited, or certified.
Given the complex nature of environmental sustainability and resilience assessments and the need to adopt the Ecosystem Function Approach (described in this chapter – Sect. 3.3), producers and processors forming business partnerships with specialists, including research and technology organisations, are becoming commonplace. It is these research organisations that develop and offer state-of-the-art methodologies, which can be exploited to evidence new unique selling points. These are critical in a competitive marketplace, and in one where consumers have become very well informed. One solution is to account for ecosystem service functions alongside environmental impact assessments using LCA – since evidence of reduced impact does not necessarily inform on improved ecosystem functions (Koellner et al., 2013).
4 Programmes, Policies, and Research to Promote the Inclusion of UCs in Agro Food Systems
Public and private policies can directly influence the adoption of UCs (Table 9.3). Several International Fund for Agricultural Development (IFAD)-funded projects of Bioversity International confirmed UCs’ livelihood benefits to poor people in numerous countries, including Bolivia, Peru, Guatemala, Mali, Nepal, and India (Padulosi et al., 2013). Their Holistic Value Chain Approach created participatory interventions at different value chain stages to overcome barriers. Public food procurement (e.g., school, hospital food programs) could shape diets by offering healthy and nutritious food for students purchased from local producers. Agrobiodiversity conservation programs that link UC farmer groups with public food procurement proved effective. For example, the Bioversity International program (‘Linking agrobiodiversity value chains, climate adaptation, and nutrition’, and ‘Empowering the poor to manage risk’) targeted the promotion of UCs in African countries funded by the IFAD and the European Commission. Finally, thousands of followers can reach a broader public via food champions and influencers. For example, when Crops for the Future launched the Forgotten Foods Network, they partnered with Prince Charles of Wales.
In Europe, perhaps the most common means by which UC could be supported is via the Common Agricultural Policy (CAP). The CAP has several functions, including increasing productivity while stabilising markets by avoiding the over-production of dominant crops and crop products, protecting income for farmers, ensuring food availability, and the affordability of food for consumers (EC, 2021). Ensuring sufficient levels of crop diversity is not the main aim of the CAP. Nevertheless, it is possible that the production of specific UCs could be encouraged via direct financial support under either CAP Pillar 1- or Pillar 2-payments. While provided by the EU, this money is administered by national or federal governments. While Pillar 1 payments relate to the area of land owned, payment is made on the basis that additional ‘cross compliant’ criteria are met. Such cross-compliance criteria can include attaining specific standards, often referred to as “Greening,” as these encompass protection measures for the environment and biodiversity – through this ambition also aims to ensure production levels are maintained and even increased (Erjavec et al., 2015). Among interventions intended to aid the environment, there is: maintaining permanent grassland; maintaining a not (necessarily) cropped “ecological focus area” (EFA), of at least 7% of the total farm area; and crop diversification, which is defined as cultivating more than 2 crops when the area which can be cropped is >10 ha, or 3 crops if >30 ha. So, a farmer need not cultivate many crop species to qualify for payment, and the current diversification standard (3 crop minimum), highlights that holistic crop rotations are largely an ideal. There is, in fact, a “crop sequence” whose composition is determined by the demand of dominant markets and less by the protection of either crop from disease or the production environment from degradation. There is no substantial accommodation of high crop diversity or UCs. Though favourable markets exist, neither is there a restriction to using underutilised species under the 2 or 3 crop-minimum rules. Also, it is possible that where a crop also fulfils environmental or biodiversity protection goals, a crop can qualify payment as an EFA too. This service has been (controversially) acknowledged for grain legumes like fava beans. No synthetic fertiliser has been applied and based on its ability to provide a resource to pollinating- and beneficial insects. However, a different reality is that while the upscaling of the cultivation and consumption of legumes is required, these are common crops which domestic EU market has elected to import. While grain legumes are under-cultivated (in Europe), they qualify as UC species.
The fact is that there are no specific means by which government schemes support crop diversification via the use of UCs and that the use of UCs is mainly realised in short-value chains (Will, 2008). These, as mentioned before, are often cultivated by small-holder farmers, and utilised by relatively small processing units operated by artisans. As such, UCs are a bastion for maintaining and developing regional food cultures and ensuring food security among the neediest in many parts of the less-industrialised world (Massawe et al., 2016). In Europe, the farmers who most commonly grow underutilised crops own a land area that is too small (<5 ha) for the production unit to qualify for income protection via the CAP. Nevertheless, several Non-Governmental Organisations and community-led groups support underutilised crops, including Crops for the Future, La Via Campesina (the International Peasants’ Movement), and the Permaculture Association, as well as community seed banks (Let’s Liberate Diversity) and Slow Food. The EU agricultural policy did not identify and define UCs per se but considers them context dependent. Therefore, UCs are not the target of any CAP policies specifically. UCs can still be embedded into the current CAP, but the current governance system marginalises them. Only recently, the Farm-to-Fork Strategy and the Biodiversity Strategy, as part of the EU Green Deal Roadmap, started to focus on the sustainability of cropping systems, which creates room for valuing UCs.
Therefore, the commercial success of UCs is often achieved by the entrepreneurship of individuals who recognise the potential of underutilised crops in strong existing markets for products already accepted by consumers. The markets of UC-based products also usually involve highlighting historic food cultures, forgotten heritage, and any additional attractive environmental, nutritional, and organoleptic attributes.
5 Concluding Remarks: The Critical Importance of a ‘Theory of Change’ Approach for the Promotion of UCs
As showcased above, UCs provide multiple nutritional, health, environmental, social, and economic benefits that go well beyond the farm level and contribute to agri-food system resilience. However, a ‘Theory of Change’ Approach is needed for UCs to be successfully included in sustainable agri-food systems. This approach recognises the need for an unprecedented degree of multi-actor strategies, whole-of-society engagement, and transformative actions. It is being developed and implemented, e.g., in the European H2020 project RADIANT (Realising Dynamic Value Chains for Underutilised Crops). It recognises the need to: (i) support multilateral learning among farmers, breeders, chefs, food retailers, scientists, representatives of food/non-food industry, and civil society; (ii) strengthen the evidence base in multiple dimensions of UC value (agronomic, environmental, economic and resilience), that also go beyond farm level (nutritional, techno-functional, health) and devise tools that integrate and showcase the potential benefits for adoption into new marketing schemes; (iii) identify the governance and policy frameworks needed for effective implementation of UCs in food and non-food value chains; and (iv) ensure that agrobiodiversity is used sustainably to meet people’s needs and that agrobiodiversity promoting actions are supported by enabling conditions (educational, financial, technological, and capacity) that effectively get UCs to farmers fields and consumers’ tables.
Implementation of the ‘Theory of Change’ Approach needs to be allied to equally facilitative socio-ecological frameworks or communities to ensure that farmers, other land managers, and citisens more generally co-develop and co-deliver interventions. These social networks and partnerships will need to seek agreed system function targets in a manner that avoids polarisation, adopting a flexible and non-prescriptive approach to land management – based on the rate or degree of change from the baseline, rather than absolute values. This will also help ensure that data-gathers and -users are receptive to the approaches and accept downstream data management, − analyses, and syntheses. Whatever procedures are undertaken, the transition from the dominant paradigm of conventional, high-input, intensive farming to more integrated, regenerative approaches supported by healthy ecosystem function and UC inclusion exposes farmers to risk. Direct and indirect costs are likely to be incurred before longer-term efficiency gains and system function improvements materialise. Although financial remuneration for undertaking the transition is required, it can be expected that such an incentive would not be necessary for the long term.
References
Adhikari, L., Hussain, A., & Rasul, G. (2017). Tapping the potential of neglected and underutilized food crops for sustainable nutrition security in the mountains of Pakistan and Nepal. Sustainability, 9(2), 291. https://doi.org/10.3390/su9020291
Alonso-Ayuso, M., Gabriel, J. L., & Quemada, M. (2016). Nitrogen use efficiency and residual effect of fertilizers with nitrification inhibitors. European Journal of Agronomy, 80, 1–8. https://doi.org/10.1016/j.eja.2016.06.008
Altieri, M. A. (1999). The ecological role of biodiversity in agroecosystems. Agriculture, Ecosystems and Environment, 74(1), 19–31.
Altieri, M. A., Merrick, L., (1987). In Situ Conservation of Crop Genetic Resources through Maintenance of Traditional Farming Systems, Economic Botany 41(1): 86–96.
Anastasi, U., Corinzia, S. A., Cosentino, S. L., et al. (2019). Performances of durum wheat varieties under conventional and no-chemical input management systems in a semiarid Mediterranean environment. Agronomy, 9, 788. https://doi.org/10.3390/agronomy9120788
Annicchiarico, P., Nazzicari, N., Laouar, M., et al. (2020). Development and proof-of-concept application of genome-enabled selection for pea grain yield under severe terminal drought. International Journal of Molecular Sciences, 21, 2414. https://doi.org/10.3390/ijms21072414
Araus, J. L., Kefauver, S. C., Zaman-Allah, M., et al. (2018). Translating high-throughput phenotyping into genetic gain. Trends in Plant Science, 23, 451–466. https://doi.org/10.1016/j.tplants.2018.02.001
Arbikie Distillery. (2021). The Nadar collection. https://arbikie.com/pages/nadar-collection. Accessed 31 Jan 2022.
Asare, P. A., Kpankpari, R., Adu, M. O., et al. (2020). Phenotypic charcaterization of tiger nuts (Cyperus esculentus L.) from major growing areas in Ghana. Scientific World Journal, 2020, 7232591. https://doi.org/10.1155/2020/7232591
Azam, M. M., Waris, A., & Nahar, N. M. (2005). Properties and potential of fatty acid methyl esters of some non-traditional seed oils for use as biodiesel in India. Biomass & Bioenergy, 29, 293–302.
Azeke, M. A., Elsanhoty, R. M., Egielewa, S. J., et al. (2011). The effect of germination on the phytase activity, phytate and total phosphorus contents of some Nigerian-grown grain legumes. Journal of the Science of Food and Agriculture, 91(1), 75–79.
Baker, B. P., & Russell, J. A. (2017). Capturing a value-added niche market: Articulation of local organic grain. American Journal of Agricultural Economics, 99, 532–545.
Baldermann, S., Blagojević, L., Frede, K., et al. (2016). Are neglected plants the food for the future? Critical Reviews in Plant Sciences, 35(2), 106–119.
Ballhorn, D. J., Kautz, S., Heil, M., et al. (2009). Cyanogenesis of wild lima bean (Phaseolus lunatus L.) is an efficient direct defence in nature. PLoS One, 4(5), e5450.
Bals, B. D., Dale, B. E., & Balan, V. (2012). Recovery of leaf protein for animal feed and high-value uses. In C. Bergeron, D. J. Carrier, & S. Ramaswamy (Eds.), Biorefinery co-products (pp. 179–197). Wiley.
Bationo, A., Ntare, B., Tarawali, S., et al. (2002). Soil fertility management and cowpea production in the semiarid tropics. In Challenges and opportunities for enhancing sustainable cowpea production (pp. 301–318). IITA.
Bavec, F., Lisec, U., & Bavec, M. (2017). Importance of underutilized field crops for increasing functional biodiversity. In B. Şen & O. Grillo (Eds.), Selected studies in biodiversity. IntechOpen. https://doi.org/10.5772/intechopen.70472
Bekkering, C. S., & Tian, L. (2019). Thinking outside of the cereal box: Breeding underutilized (pseudo) cereals for improved human nutrition. Frontiers in Genetics, 10, 1289. https://doi.org/10.3389/fgene.2019.01289
Benner, E., Profeta, A., Wirsig, A., et al. (2008). Übergangsregelung Zum Herkunftsschutz Bei Agrarprodukten Und Lebensmitteln Aus Dem Blickwinkel Der Transaktions- Und Der Informationsökonomie. https://doi.org/10.22004/AG.ECON.52644
Benton, T. G., & Bailey, R. (2019). The paradox of productivity: Agricultural productivity promotes food system inefficiency. Global Sustainability, 2, e6. https://doi.org/10.1017/SUS.2019.3
Berbereia, V. L. D. (2015). Promoção do uso de alimentos promotores de saúde na dieta de grupos socioeconómicos desfavorecidos. https://repositorio.uac.pt/handle/10400.3/3551
Beulens, A. J., Broens, D. F., Folstar, P., et al. (2005). Food safety and transparency in food chains and networks relationships and challenges. Food Control, 16, 481–486.
Bioversity International and IFAD. (2021). How to do. Mainstreaming NUS in national policy for nutritionoutcomes. IFAD. https://www.ifad.org/en/web/knowledge/-/how-to-do-note-mainstreaming-nus-in-national-policy-for-nutrition-outcomes
Biowert (2021) Products. https://biowert.com/products. Accessed 8 Nov 2021.
Birkhofer, K., Smith, H. G., & Rundlöf, M. (2016). Environmental impacts of organic farming. In eLS. Wiley. https://doi.org/10.1002/9780470015902.a0026341
Bishop, K. R., Betzelberger, A. M., Long, S. P., et al. (2015). Is there potential to adapt soybean (Glycine max Merr.) to future [CO2]? An analysis of the yield response of 18 genotypes in free- air CO2 enrichment. Plant, Cell & Environment, 38, 1765–1774.
BLE. (n.d.). Pro-Kopf-Konsum von Hülsenfrüchten in Deutschland bis 2016/2017 | Statista. https://de.statista.com/statistik/daten/studie/175416/umfrage/pro-kopf-verbrauch-von-huelsenfruechten-in-deutschland-seit-1935/. Accessed 21 Jan 2022.
Bokdam, J., & Wallis de Vries, M. F. (1992). Forage quality as a limiting factor for cattle grazing in isolated Dutch nature reserves. Conservation Biology, 6, 399–408.
Boob, M., Elsaesser, M., Thumm, U., et al. (2019). Harvest time determines quality and usability of biomass from lowland hay meadows. Agriculture, 9(9), 198.
Bourgault, M., Brand, J., Tausz-Posch, S., et al. (2017). Yield, growth and grain nitrogen response to elevated CO2 in six lentil (Lens culinaris) cultivars grown under Free Air CO2 Enrichment (FACE) in a semi-arid environment. European Journal of Agronomy, 87, 50–58. https://doi.org/10.1016/j.eja.2017.05.003
Bovolenta, S., Spanghero, M., Dovier, S., et al. (2008). Chemical composition and net energy content of alpine pasture species during the grazing season. Animal Feed Science and Technology, 140, 164–177.
Britt, J. H., Cushman, R. A., Dechow, C. D., et al. (2018). Invited review: Learning from the future – A vision for dairy farms and cows in 2067. Journal of Dairy Science, 101(5), 3722–3741. https://doi.org/10.3168/jds.2017-14025
Budak, H., Kantar, M., & Kurtoglu, K. Y. (2013). Drought tolerance in modern and wild wheat. The Scientific World Journal, 2013, 548246. https://doi.org/10.1155/2013/548246
Buiatti, M., Esquinas-Alcázar, J., Lazzerini, G., et al. (2010). Biodiversity and ecosystems. In AAVV. Food policy and sustainability. SlowFood.
Burgess, M. A. (1994). Cultural responsibility in the preservation of local economic plant resources. Biodiversity and Conservation, 3, 126–136.
Burney, J. A., Davis, S. J., & Lobell, D. B. (2010). Greenhouse gas mitigation by agricultural intensification. https://doi.org/10.1073/pnas.0914216107
Camacho-Henriquez, A., Kraemer, F., Galluzzi, G., et al. (2016). Decentralized collaborative plant breeding for utilization and conservation of neglected and underutilized crop genetic resources. In Advances in plant breeding strategies: Breeding, biotechnology and molecular tools (Vol. 1, pp. 25–61).
Chen, Y. S., & Chang, C. H. (2013). Greenwash and green trust: The mediation effects of green consumer confusion and green perceived risk. Journal of Business Ethics, 114, 489–500.
Cheng, A., Mayes, S., Dalle, G., et al. (2017). Diversifying crops for food and nutrition security – A case of teff. Biological Reviews, 92(1), 188–198. https://doi.org/10.1111/BRV.12225
Chilla, T., Fink, B., Balling, R., et al. (2020). The EU food label ‘protected geographical indication’: Economic implications and their spatial dimension. Sustainability, 12(14), 5503. https://doi.org/10.3390/SU12145503
Chishakwe, N. E. (2008). The role of policy in the conservation and extended use of underutilised plant species: A cross-national policy analysis. N. 31 p. ISBN: 978-92-9043-782-6.
Clark, M. A., Domingo, N. G. G., Colgan, K., et al. (2020). Global food system emissions could preclude achieving the 1.5° and 2°C climate change targets. Science, 370(6517), 705–708. https://doi.org/10.1126/SCIENCE.ABA7357/SUPPL_FILE/ABA7357_DATAS3.ZIP
Collinson, S. T., Azam-Ali, S. N., Chavula, K. M., et al. (1996). Growth, development and yield of bambara groundnut (Vigna subterranea) in response to soil moisture. Journal of Agricultural Science, 126, 307–318.
Collinson, S. T., Clawson, E. J., Azam-Ali, S. N., et al. (1997). Effect of soil moisture deficits on the water relations of bambara groundnut (Vigna subterranea L. Verdc.). Journal of Experimental Botany, 48, 877–884.
Commission on Sustainable Agriculture and Climate Change (2012). Achieving food security in the face of climate change. Final report from the Commission on Sustainable Agriculture and Climate Change Commission Secretariat. www.ccafs.cgiar.org/commission
Cordain, L., Eaton, S. B., Sebastian, A., et al. (2005). Origins and evolution of the Western diet: Health implications for the 21st century. The American Journal of Clinical Nutrition, 81(2), 341–354. https://doi.org/10.1093/AJCN.81.2.341
Cristofano, F., El-Nakhel, C., & Rouphael, Y. (2021). Biostimulant substances for sustainable agriculture: Origin, operating mechanisms and effects on cucurbits, leafy greens, and nightshade vegetables species. Biomolecules, 11, 1103. https://doi.org/10.3390/biom11081103
Cusworth, G., Garnett, T., & Lorimer, J. (2021). Legume dreams: The contested futures of sustainable plant-based food systems in Europe. Global Environmental Change, 69, 102321. https://doi.org/10.1016/j.gloenvcha.2021.102321
Dantas, E. F., Freitas, A. D., Lyra, M. D., et al. (2019). Biological fixation, transfer and balance of nitrogen in passion fruit (Passiflora edulis Sims) orchard intercropped with different green manure crops. Australian Journal of Crop Science, 13(03), 465–471.
Das, S., Khound, R., Santra, M., et al. (2019). Beyond bird feed: Proso millet for human health and environment. Agriculture, 9(3), 64. https://doi.org/10.3390/agriculture9030064
Dawson, I. K., Hedley, P. E., Guarino, L., et al. (2009). Does biotechnology have a role in the promotion of underutilised crops? Food Policy, 34(4), 319–328. https://doi.org/10.1016/J.FOODPOL.2009.02.003
Dien, B. S., Mitchell, R. B., Bowman, M. J., et al. (2018). Bioconversion of pelletized big bluestem, switchgrass, and low-diversity grass mixtures into sugars and bioethanol. Frontiers in Energy Research, 6, 129. https://doi.org/10.3389/fenrg.2018.00129
Diffenbaugh, N., Krupke, C. H., White, M. A., et al. (2008). Global warming presents new challenges for maize pest management. Environmental Research Letters, 3(4). https://doi.org/10.1088/1748-9326/3/4/044007
Dong, L., Si, T., Li, Y.-E., et al. (2021). The effect of conservation tillage in managing climate change in arid and semiarid areas—A case study in Northwest China. Mitigation and Adaptation Strategies for Global Change, 26(4), 17. https://doi.org/10.1007/s11027-021-09956-3
Dubey, P. K., Singh, G. S., & Abhilash, P. C. (2020). Adaptive agronomic practices for sustaining food production. In Adaptive Agricultural Practices (Springer briefs in environmental science). Springer. https://doi.org/10.1007/978-3-030-15519-3_2
Dulloo, M., Drucker, A., Gaisberger, H., et al. (2016). Mainstreaming agrobiodiversity in sustainable food systems: Scientific foundations for an agrobiodiversity index – Summary (pp. 22–25). Bioversity International.
Dwivedi, S. L., Ceccarelli, S., Blair, M. W., et al. (2016). Landrace germplasm for improving yield and abiotic stress adaptation. Trends in Plant Science, 21(1), 31–42. https://doi.org/10.1016/j.tplants.2015.10.012
Ebert, A. W. (2014). Potential of underutilized traditional vegetables and legume crops to contribute to food and nutritional security, income and more sustainable production systems. Sustainability, 6, 319–335. https://doi.org/10.3390/su6010319
Ehrlich, P. R., & Ehrlich, A. H. (1981). Extinction: The causes and consequences of the disappearance of species (pp. 72–98). Random House.
Ekanayake, S., Nair, B., Jansz, E. R., et al. (2003). Effect of processing on the protein nutritional value of Canavalia gladiate seeds. Die Nahrung, 47(4), 256–260.
Erjavec, E., Lovec, M., & Erjavec, K. (2015). From “Greening” to “Greenwash”: the drivers and discourses of CAP 2020 reform. In: J.F. Swinnen (Ed.), The Political Economy of the 2014-2020 Common Agricultural Policy: An Imperfect Storm. Rowman & Littlefield, London and CEPS, Brussels, ISBN: 978-1-78348-484-3, p. 215–244.
Eshel, G., Shepon, A., Makov, T., et al. (2014). Land, irrigation water, greenhouse gas, and reactive nitrogen burdens of meat, eggs, and dairy production in the United States. Proceedings of the National Academy of Sciences of the United States of America, 111(33), 11996–12001. https://doi.org/10.1073/pnas.1402183111
EURISCO. (2022). European search catalogue for plant genetic resources. Available at: https://eurisco.ipk-gatersleben.de/. Accessed 5 Feb 2022.
European Commission. (2021). The common agricultural policy at a glance: The common agricultural policy supports farmers and ensures Europe’s food security. https://ec.europa.eu/info/food-farming-fisheries/key-policies/common-agricultural-policy/cap-glance_en. Accessed 31 Jan 2022.
Eyzaguirre, P., Padulosi, S., & Hodgkin, T. (1999). IPGRI’s strategy for neglected and underutilised species and the human dimension of agrobiodiversity. In S. Padulosi (Ed.), Priority setting for underutilised and neglected plant species of the Mediterranean region. Report of the IPGRI conference, 9–11. February 1998, ICARDA, Aleppo. Syria. International Plant Genetic Resources Institute.
Fahey, J. W. (2005). Moringa oleifera: A review of the medical evidence for its nutritional, therapeutic, and prophylactic properties. Part 1. Phytochemistry, 47, 123–157.
Fanzo, J., Haddad, L., Schneider, K. R., et al. (2021). Viewpoint: Rigorous monitoring is necessary to guide food system transformation in the countdown to the 2030 global goals. Food Policy, 104, 102163. https://doi.org/10.1016/J.FOODPOL.2021.102163
FAO. (1999). Agricultural biodiversity. In Multifunctional Character of Agriculture and Land Conference, Maastricht, 12–17 Sept 1999.
FAO. (2001). International treaty on plant genetic resource for food and agriculture, Rome.
FAO. (2010a). Proceedings of the International Scientific Symposium Biodiversity and Sustainable diets united against hunger. 3–5 Nov 2010. Available in: https://www.fao.org/3/i3004e/i3004e00.pdf
FAO (2010b). The second report on the state of the world’s plant genetic resources for food and agriculture (pp. 183–201). Food and Agriculture Organization of the United Nations, Rome.
FAO. (2018a). Global livestock environmental assessment model. http://www.fao.org/fileadmin/user_upload/gleam/docs/GLEAM_2.0_Model_description.pdf
FAO. (2018b). The future of Food and Agriculture: Alternative pathways to 2050 | Knowledge for policy. https://knowledge4policy.ec.europa.eu/publication/future-food-agriculture-alternative-pathways-2050_en
FAO. (2019a). The state of the world’s biodiversity for food and agriculture. In J. Bélanger & D. Pilling (Eds.), FAO commission on genetic resources for food and agriculture assessments. 572 pp.
FAO (2019b). Mountain agriculture: Opportunities for harnessing zero hunger in Asia. Food and Agriculture Organization of the United Nations, Bangkok.
FAO. (2021). The state of food and agriculture 2021. https://doi.org/10.4060/CB4476EN.
FAOSTAT, Food and Agriculture Organization of the United Nations. Available at: https://www.fao.org/faostat/en/#data/QCL. Accessed 25 Jan 2022.
Ferchichi, S., Hessini, K., Dell Aversana, E., et al. (2018). Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Functional Plant Biology, 45(11), 1096–1109.
Finell, M. (2003). The use of reed canary grass (Phalaris arundinacea) as a short fibre raw material for the pulp and paper industry (Dissertation). Swedish University of Agricultural Sciences.
Foley, J. A., Ramankutty, N., Brauman, K. A., et al. (2011). Solutions for a cultivated planet. Nature, 478(7369), 337–342.
Foyer, C. H., Lam, H. M., Nguyen, H. T., et al. (2016). Neglecting legumes has compromised human health and sustainable food production. Nature Plants, 2, 16112.
Fraj, M. B., Al-Dakheel, A. J., McCann, I. R., et al. (2013). Selection of high yielding and stable safflower (Carthamus tinctorius L.) genotypes under salinity stress. Agricultural Science Research Journal, 3, 273–283.
Fraj, M. B., Al-Dakheel, A. J., McCann, I. R., et al. (2014). Selection of salt-tolerant triticale (x Triticum secale Wittmack) genotypes for grain and forage end-uses. American-Eurasian Journal of Agricultural and Environmental, 14, 445–454.
Freitas, R. M., Dombroski, J. L., Freitas, F. C., et al. (2019). Water use of cowpea under deficit irrigation and cultivation systems in semi-arid region. Revista Brasileira de Engenharia Agrícola e Ambiental, 23(4), 271–276.
Galmés, J., Conesa, M. À., Ochogavía, J. M., Perdomo, J. A., Francis, D. M., Ribas-Carbó, M., Savé, R., Flexas, J., Medrano, H., & Cifre, J. (2011). Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant, Cell & Environment, 34, 245–260. https://doi.org/10.1111/j.1365-3040.2010.02239.x
Gatsios, A., Ntatsi, G., Celi, L., et al. (2019). Nitrogen nutrition optimization in organic greenhouse tomato through the use of legume plants as green manure or intercrops. Agronomy, 9, 766. https://doi.org/10.3390/agronomy9110766
Gatsios, A., Ntatsi, G., Yfantopoulos, D., et al. (2021). Effects of different organic soil amendments on nitrogen nutrition and yield of organic greenhouse tomato crop. Nitrogen, 2, 347–358. https://doi.org/10.3390/nitrogen2030024
Gatsios, A., Ntatsi, G., Celi, L., et al. (2021a). Impact of legumes as a pre-crop on nitrogen nutrition and yield in organic greenhouse tomato. Plants, 10, 468. https://doi.org/10.3390/plants10030468
Gatsios, A., Ntatsi, G., Celi, L., et al. (2021b). Legume-based mobile green manure can increase soil nitrogen availability and yield of organic greenhouse tomatoes. Plants, 10, 2419. https://doi.org/10.3390/plants10112419
GENESYS. (2022). The global gateway to genetic resources. Available online at: https://www.genesys-pgr.org. Accessed 5 Feb 2022.
Ghafoor, I., Habib-ur-Rahman, M., Ali, M., et al. (2021). Slow-release nitrogen fertilizers enhance growth, yield, NUE in wheat crop and reduce nitrogen losses under an arid environment. Environmental Science and Pollution Research, 28, 43528–43543. https://doi.org/10.1007/s11356-021-13700-4
Grass, S. (2004). Utilisation of grass for production of fibres, protein, and energy. In Biomass and agriculture. Sustainability, markets and policies (pp. 169–177). OECD.
Gregory, P. J., Mayes, S., Hui, C. H., et al. (2019). Crops For the Future (CFF): An overview of research efforts in the adoption of underutilised species. Planta, 250, 979–988. https://doi.org/10.1007/s00425-019-03179-2
Gruère, G. P., Giuliani, A., & Smale, M. (2008). Marketing underutilized plant species for the poor: A conceptual framework. In Agrobiodiversity Conservation and Economic Development (pp. 86–105). Routledge, London, UK.
Guida, G., Sellami, M. H., Mistretta, C., et al. (2017). Agronomical, physiological and fruit quality responses of two italian long-storage tomato landraces under rain-fed and full irrigation conditions. Agricultural Water Management, 180, 126–135.
Habiyaremye, C., Barth, V., Highet, K., et al. (2017). Phenotypic responses of twenty diverse proso millet (Panicum miliaceum L.) accessions to irrigation. Sustainability, 9, 389.
Hadavi, E., & Ghazijahani, N. (2018). Closed and semi-closed systems in agriculture. In E. Lichtfouse (Ed.), Sustainable agriculture reviews 33. Springer. https://doi.org/10.1007/978-3-319-99076-7_10
Haddad, L., Hawkes, C., Webb, P. et al. (2016). A new global research agenda for food. Nature 540, 30–32. https://doi.org/10.1038/540030a
Hadebe, S., Modi, A. T., & Mabhaudhi, T. (2017). Drought tolerance and water use of cereal crops: A focus on sorghum as a food security crop in Sub-Saharan Africa. Journal of Agronomy and Crop Science, 203, 177–191.
Havlin, J. L. (2020). Soil: Fertility and nutrient management. In Landscape and land capacity (pp. 251–265). CRC Press, Boca Raton, Florida, USA
Hawes, C., Haughton, A. J., Bohan, D. A., et al. (2009). Functional approaches for assessing plant and invertebrate abundance patterns in arable systems. Basic and Applied Ecology, 10, 34–47.
Hawes, C., Begg, G. S., Iannetta, P., et al. (2016). A whole-systems approach for assessing measures to improve arable ecosystem sustainability. Ecosystem Health and Sustainability, 2, e01252.
Hermansen, J., Jørgensen, U., Lærke, P. E., et al. (2017). Green biomass - protein production through bio-refining. DCA Report.
Holm, L., & Møhl, M. (2000). The role of meat in everyday food culture: An analysis of an interview study in Copenhagen. Appetite, 34(3), 277–283. https://doi.org/10.1006/APPE.2000.0324
Huang, J., Xu, C., Ridoutt, B. G., et al. (2017). Nitrogen and phosphorus losses and eutrophication potential associated with fertilizer application to cropland in China. Journal of Cleaner Production, 159, 171–179.
Hunter, M. C., Smith, R. G., Schipanski, M. E., et al. (2017). Agriculture in 2050: Recalibrating targets for sustainable intensification. Bioscience, 67(4), 386–391. https://doi.org/10.1093/BIOSCI/BIX010
Hunter, D., Borelli, T., Beltrame, D. M. O., et al. (2019). The potential of neglected and underutilized species for improving diets and nutrition. Planta, 250(3), 709–729. https://doi.org/10.1007/s00425-019-03169-4
Huppmann, D., Rogelj, J., Kriegler, E., Krey, V., & Riahi, K. (2018). A new scenario resource for integrated 1.5 °C research. In Nature Climate Change (Vol. 8, Issue 12, pp. 1027–1030). Nature Publishing Group. https://doi.org/10.1038/s41558-018-0317-4
Ibeanu, V. N., Edeh, C. G., & Ani, P. N. (2020). Evidence-Based Strategy for Prevention of Hidden Hunger among Adolescents in a Suburb of Nigeria. BMC Public Health, 20(1), 1–10. https://doi.org/10.1186/s12889-020-09729-8
Imathiu, S. (2021). Neglected and underutilized cultivated crops with respect to indigenous African leafy vegetables for food and nutrition security. Journal of Food Security, 9(3), 115–125.
International assessment of agricultural knowledge, science and technology for development. (2009). Agriculture at a crossroads: A global report. Island Press.
IPCC. (2019). Climate change and land. An IPCC Special Report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems. www.ipcc.ch
Jahn, S. A. A., Musnad, H. A., & Burgstaller, H. (1986). The tree that purifies water: Cultivating multipurpose Moringaceae in the Sudan. Unasylva, 38, 23–28.
Jamnadass, R., Mumm, R. H., Hale, I., et al. (2020). Enhancing African orphan crops with genomics. Nature Genetics, 52, 356–360. https://doi.org/10.1038/s41588-020-0601-x
Jansen, M., Guariguata, M. R., Raneri, J. E., et al. (2020). Food for thought: The underutilized potential of tropical tree-sourced foods for 21st century sustainable food systems. People and Nature, 2(4), 1006–1020.
Jungers, J. M., Fargione, J. E., Sheaffer, C. C., et al. (2013). Energy potential of biomass from conservation grasslands in Minnesota, USA. PLoS One, 8(4), e61209. https://doi.org/10.1371/journal.pone.0061209
Kamel, S. H., Zaki, Z. M., & Kassim, J. (2018). The effectiveness of Psophocarpus tetragonolobus’s seed as turbidity removal. International Journal of Engineering and Technology, 7, 144–146.
Kamenya, S. N., Mikwa, E. O., Song, B., & Odeny, D. A. (2021). Genetics and breeding for climate change in orphan crops. Theoretical and Applied Genetics, 134, 1787–1815. https://doi.org/10.1007/s00122-020-03755-1
Kanatas, P. (2020). Mini-review: The role of crop rotation, intercropping, sowing dates and increased crop density towards a sustainable crop and weed management in arable crops. Agraarteadus, 31(1), 22–27. https://doi.org/10.15159/jas.20.11
Karavidas, I., Ntatsi, G., Vougeleka, V., et al. (2022). Agronomic practices to increase the yield and quality of common bean (Phaseolus vulgaris L.): A systematic review. Agronomy, 12, 271. https://doi.org/10.3390/agronomy12020271
Karkanis, A., Ntatsi, G., Lepse, L., et al. (2018). Faba bean cultivation – Revealing novel managing practices for more sustainable and competitive European cropping systems. Frontiers in Plant Science, 9, 1115. https://doi.org/10.3389/fpls.2018.01115
Karkanis, A., Polyzos, N., Kompocholi, M., et al. (2022). Rock samphire, a candidate crop for saline agriculture: Cropping practices, chemical composition and health effects. Applied Sciences, 12, 737. https://doi.org/10.3390/app12020737
Karunaratne, A. S., Walker, S., & Azam-Ali, S. N. (2015). Assessing the productivity and resource-use efficiency of underutilised crops: Towards an integrative system. Agricultural Water Management, 147, 129–134.
Katschnig, D., Broekman, R. A., & Rozema, J. (2013). Salt tolerance in the halophyte Salicornia dolichostachya Moss: Growth, morphology and physiology. Environmental and Experimental Botany, 92, 32–42.
Keatinge, J. D. H., Waliyar, F., Jamnadass, R. H., et al. (2010). Re-learning old lessons for the future of food-by bread alone no longer: Diversifying diets with fruit and vegetables. Crop Science, 50, 51–62.
Khoury, C. K., Bjorkman, A. D., Dempewolf, H., et al. (2014). Increasing homogeneity in global food supplies and the implications for food security. Proceedings of the National Academy of Sciences of the United States of America, 111, 4001–4006. https://doi.org/10.1073/pnas.1313490111
King, C., Richardson, M., McEniry, J., et al. (2013). Potential use of fibrous grass silage press-cake to minimise shrinkage cracking in low-strength building materials. Biosystems Engineering, 115, 203–210.
Koellner, T., De Baan, L., Beck, T., et al. (2013). UNEP-SETAC guideline on global land use impact assessment on biodiversity and ecosystem services in LCA. The International Journal of Life Cycle Assessment, 18, 1188–1202.
Koidou, M., Mountousis, I., Dotas, V., et al. (2019). Temporal variations of herbage production and nutritive value of three grasslands at different elevation zones regarding grazing needs and welfare of ruminants. Archives Animal Breeding, 62, 215–226.
Kuo, C. G., Chen, H. M., & Sun, H. C. (1992). Membrane thermostability and heat tolerance of vegetable leaves. In Adaptation of food crops to temperature and water stress (pp. 160–168). AVRDC - The World Vegetable Center.
Karunaratne, A. S., Walker, S., & Azam-Ali, S. N. (2015). Assessing the productivity and resource-use efficiency of underutilised crops: Towards an integrative system. Agricultural Water Management, 147, 129–134.
Lakew, B., Eglinton, J., Henry, R. J., Baum, M., Grando, S., & Ceccarelli, S. (2011). The potential contribution of wild barley (Hordeum vulgare ssp. spontaneum) germplasm to drought tolerance of cultivated barley (H. vulgare ssp. vulgare). Field Crops Research, 120(1), 161–168.
Lazcano, C., Zhu-Barker, X., & Decock, C. (2021). Effects of organic fertilizers on the soil microorganisms responsible for N2O emissions: A review. Microorganisms, 9(5), 983. https://doi.org/10.3390/microorganisms9050983
Lewandowski, I., Clifton-Brown, J. C., Andersson, B., et al. (2003). Environment and harvest time affects the combustion qualities of Miscanthus genotypes. Agronomy Journal, 95, 1274–1280.
Li, X., & Siddique, K. H. M. (2020). Future smart food: Harnessing the potential of neglected and underutilized species for zero hunger. Maternal and Child Nutrition, 16(S3), 1–22. https://doi.org/10.1111/mcn.13008
Li, J., Wang, Y. K., Guo, Z., et al. (2020a). Effects of conservation tillage on soil physicochemical properties and crop yield in an arid loess plateau, China. Scientific Reports, 10(1), 4716. https://doi.org/10.1038/s41598-020-61650-7
Li, X., Yadav, R., & Siddique, K. H. M. (2020b). Neglected and underutilized crop species: The key to improving dietary diversity and fighting hunger and malnutrition in Asia and the Pacific. Frontiers in Nutrition, 7, 593711. https://doi.org/10.3389/fnut.2020.593711
Liakos, K. G., Busato, P., Moshou, D., et al. (2018). Machine learning in agriculture: A review. Sensors (Switzerland), 18(8). https://doi.org/10.3390/s18082674
Liang, J., Chen, X., Guo, P., et al. (2021). Grafting improves nitrogen-use efficiency by regulating the nitrogen uptake and metabolism under low-nitrate conditions in cucumber. Scientia Horticulturae, 289, 110454. https://doi.org/10.1016/j.scienta.2021.110454
Lienhardt, T., Black, K., Saget, S., et al. (2019a). Just the tonic! Legume biorefining for alcohol has the potential to reduce Europe’s protein deficit and mitigate climate change. Environment International, 130, 104870. https://doi.org/10.1016/j.envint.2019.05.064
Lienhardt, T., Black, K., Saget, S., et al. (2019b). Data for life cycle assessment of legume biorefining for alcohol. Data in Brief, 25. https://doi.org/10.1016/j.dib.2019.104242
Linnemann, A. R., & Azam-Ali, S. (1993). Bambara groundnut (Vigna subterranea). In J. T. William (Ed.), Pulses and vegetables (pp. 13–57). Chapman and Hall – CRC Press, Boca Raton, Florida, USA.
Lowe, N. M. (2021). The global challenge of hidden hunger: Perspectives from the field. Proceedings of the Nutrition Society, 80(3), 283–289. https://doi.org/10.1017/S0029665121000902
Mabhaudhi, T., O’Reilly, P., Walker, S., et al. (2016). Opportunities for underutilised crops in Southern Africa’s post–2015 development agenda. Sustainability, 8(4), 302.
Mabhaudhi, T., Chimonyo, V. G. P., Hlahla, S., et al. (2019). Prospects of orphan crops in climate change. Planta, 250(3). https://doi.org/10.1007/s00425-019-03129-y
Mafakheri, M., & Kordrostami, M. (2020). Newly revealed promising gene pools of neglected brassica species to improve stress-tolerant crops. In M. Hasanuzzaman (Ed.), The plant family Brassicaceae. Springer. https://doi.org/10.1007/978-981-15-6345-4_4
Mammadov, J., Buyyarapu, R., Guttikonda, S. K., et al. (2018). Wild relatives of maize, rice, cotton, and soybean: Treasure troves for tolerance to biotic and abiotic stresses. Frontiers in Plant Science, 9. https://doi.org/10.3389/fpls.2018.00886
Mandizvo, T., Odindo, A. O., & Mashilo, J. (2021). Citron watermelon potential to improve crop diversification and reduce negative impacts of climate change. Sustainability, 13(4), 2269. https://doi.org/10.3390/su13042269
Massawe, F., Mayes, S., & Cheng, A. (2016). Crop diversity: An unexploited treasure trove for food security. Trends in Plant Science, 21, 365–368. https://doi.org/10.1016/j.tplants.2016.02.006
Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., et al. (2019). Global warming of 1.5°C An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty Summary for Policymakers Edited by Science Officer Science Assistant Graphics Officer Working Group I Technical Support Unit. https://report.ipcc.ch/sr15/pdf/sr15_spm_final.pdf
Matthews, N. E., Stamford, L., & Shapira, P. (2019). Aligning sustainability assessment with responsible research and innovation: Towards a framework for Constructive Sustainability Assessment. Sustainable Production and Consumption, 20, 58–73. https://doi.org/10.1016/j.spc.2019.05.002
Mayes, S., Massawe, F. J., Alderson, P. G., et al. (2012). The potential for underutilized crops to improve security of food production. Journal of Experimental Botany, 63(3), 1075–1079. https://doi.org/10.1093/JXB/ERR396
Mecha, E., Natalello, S., Carbas, B., et al. (2021). Disclosing the nutritional quality diversity of Portuguese common beans—The missing link for their effective use on protein quality breeding programmes. Agronomy, 11, 221. https://doi.org/10.3390/agronomy11020221
Mejias, J. H., Salazar, F., Pérez-Amaro, L., et al. (2021). Nanofertilizers: A cutting-edge approach to increase nitrogen use efficiency in grasslands. Frontiers in Environmental Science, 9, 635114. https://doi.org/10.3389/fenvs.2021.635114
Meldrum G., Sthapit S., Rojas W., et al. (2016) Methodology mapping for resilient production systems: Approaches and results from surveys in Bolivia, India, and Nepal. Proceedings of the 3rd International Conference on Neglected and Underutilized Species: For a Food-Secure Africa, Accra, Ghana, 25–27 September 2013, pp. 108–112.
Mentis, M. (2020). Environmental rehabilitation of damaged land. Forest Ecosystems, 7(1). https://doi.org/10.1186/s40663-020-00233-4
Millennium Ecosystem Assessment (MEA). (2005). Ecosystems and human well-being: Synthesis. Island Press, Washington, DC, 137p. https://www.millenniumassessment.org/documents/document.356.aspx.pdf
Mintel. (2021). Global consumer trends 2022. Available in: https://downloads.mintel.com/private/r3Mts/files/889468/
Mládek, J., Hejcman, M., Hejduk, S., et al. (2011). Community seasonal development enables late defoliation without loss of forage quality in semi-natural grasslands. Folia Geobotanica, 46, 17–34.
Mndzebele, B., Ncube, B., Fessehazion, M. K., et al. (2020). Effects of cowpea-amaranth intercropping and fertiliser application on soil phosphatase activities, available soil phosphorus, and crop growth response. Agronomy, 10(1), 79. https://doi.org/10.3390/agronomy10010079
Morris, J. B. (2009). Morphological and reproductive characterization in hyacinth bean, Lablab purpureus (L.) sweet germplasm with clinically proven nutraceutical and pharmaceutical traits for use as a medicinal food. Journal of Dietary Supplements, 6(3), 263–279.
Mos, M., Banks, S. W., Nowakowski, D. J., et al. (2013). Impact of Miscanthus x giganteus senescence times on fast pyrolysis bio-oil quality. Bioresource Technology, 129, 335–342.
Mueller, N., Gerber, J., Johnston, M., et al. (2012). Closing yield gaps through nutrient and water management. Nature, 490, 254–257. https://doi.org/10.1038/nature11420
Murevanhema, Y. Y., & Jideani, V. A. (2013). Potential of bambara groundnut (Vigna subterranea (L.) verdc) milk as a probiotic beverage-a review. Critical Reviews in Food Science and Nutrition, 53(9), 954–967.
Murthy, H. N., & Bapat, V. A. (2020). Importance of underutilized fruits and nuts. In H. N. Murthy & V. A. Bapat (Eds.), Bioactive compounds in underutilized fruits and nuts (pp. 3–19). Springer Nature. https://doi.org/10.1007/978-3-030-30182-8
Naeem, M., Khan, M. M., & Morris, J. (2009). Agrobotanical attributes, nitrogen-fixation, enzyme activities and nutraceuticals of hyacinth bean (Lablab purpureus L.): A bio-functional medicinal legume. American Journal of Plant Physiology, 4, 58–69.
Nandal, U., & Bhardwaj, R. L. (2014). The role of underutilized fruits in nutritional and economic security of tribals: A review. Critical Reviews in Food Science and Nutrition, 54, 880–890. https://doi.org/10.1080/10408398.2011.616638
Nc’Nean Distillery. (2021). Sustainability report. https://ncnean.com/pages/sustainability. Accessed 31 Jan 2022.
Ndjiondjop, M. N., Manneh, B., Cissoko, M., et al. (2010). Drought resistance in an interspecific backcross population of rice (Oryza spp.) derived from the cross WAB56-104 (O. sativa) x CG14 (O. glaberrima). Plant Science, 179, 364–373.
Nganga, S. (2014). African leafy vegetables and household wellbeing in Kenya: A disaggregation by gender. Current Research Journal of Social Sciences, 6(4), 82–94.
Nielsen, D. C., & Vigil, M. F. (2017). Water use and environmental parameters influence proso millet yield. Field Crops Research, 212, 34–44.
Ntatsi, G., Karkanis, A., Yfantopoulos, D., et al. (2018a). Evaluation of the field performance, nitrogen fixation efficiency and competitive ability of pea landraces grown under organic and conventional farming systems. Archives of Agronomy and Soil Science, 65, 294–307. https://doi.org/10.1080/03650340.2018.1501155
Ntatsi, G., Karkanis, A., Yfantopoulos, D., et al. (2018b). Impact of variety and farming practices on growth, yield, weed flora and symbiotic nitrogen fixation in faba bean cultivated for fresh seed production. Acta Agriculturae Scandinavica, Section B — Soil & Plant Science, 68, 619–630. https://doi.org/10.1080/09064710.2018.1452286
O’Keeffe, S., Schulte, R., Sanders, J., et al. (2011). I. Technical assessment for first generation green biorefinery (GBR) using mass and energy balances: Scenarios for an Irish GBR blueprint. Biomass and Bioenergy, 35, 4712–4723.
Obasi, M., & Ezedinma, F. (1991). Floral biology of Kerstings groundnut (Kerstingiella geocarpa harms). Ghana Journal of Agricultural Science, 24–27, 145–149.
Oelke, E. A., Oplinger, E. S., Teynor, T. M., et al. (1992). Safflower. Alternative crops field manual. Universities of Wisconsin Cooperative Extension Service, University of Minnesota Center for Alternative Plant and Animal Products & Minnesota Extension Service. http://www.hort.purdue.edu/newcrop/afcm/safflower.html
Onyekachi, O. G., Boniface, O. O., Gemlack, N. F., et al. (2019). The effect of climate change on abiotic plant stress: A review. In A. De Oliveira (Ed.), Abiotic and biotic stress in plants. 13pp. Available at: https://www.intechopen.com/chapters/64791. Accessed 27 Jan 2022.
Oyelakin, O. S., & Olaniyi, O. W. (2019). Effects of intercropping maize (Zea mays L.) with Jack beans (Canavalia ensiformis L.) at different spacing and weeding regimes on crops productivity. International Journal of Agricultural and Biosystems Engineering, 13(3), 61–65.
Padulosi, S., Eyzaquirre, P., & Hodgkin, T. (1999). Challenges and strategies in promoting conservation and use of neglected and underutilized crop species. In J. Janick (Ed.), Perspectives on new crops and new uses (p. 140). ASHS Press.
Padulosi, S., Hodgkin, T., Williams, J., & Haq, N. (2002). 30 underutilized crops: trends, challenges and opportunities in the 21st century. In: Managing plant genetic diversity (eds J.M.M. Engels, V. Ramanatha Rao, A.H.D. Brown and M.T. Jackson) 323–487
Padulosi, S., Heywood, V., Hunter, D., et al. (2011). Underutilized species and climate change: Current status and outlook. In S. S. Yadav, R. J. Redden, J. L. Hatfield, H. Lotze-Campen, & A. E. Hall (Eds.), Crop adaptation to climate change. John Wiley & Sons, Ltd.: Blackwell Publishing Ltd.
Padulosi, S., Thompson, J., & Rudebjer, P. (2013). Fighting poverty, hunger and malnutrition with neglected and underutilised species (NUS): Needs, challenges and the way forward. Bioversity International 60 p. https://cgspace.cgiar.org/handle/10568/68927
Palou, E., López-Malo, A., Barbosa-Canovas, G. V., et al. (2020). High-pressure preservation of foods. In Handbook of food preservation (pp. 843–872). CRC Press. https://doi.org/10.1201/9780429091483-54
Parihar, A. K., Basandrai, A. K., Kushwaha, K. P., et al. (2018). Targeting test environments and rust-resistant genotypes in lentils (Lens culinaris) by using heritability-adjusted biplot analysis. Crop & Pasture Science, 69, 1113–1125.
Parvin, S., Uddin, S., Bourgault, M., et al. (2019a). Effect of heat wave on N2 fixation and N remobilization of lentil (Lens culinaris MEDIK) grown under Free Air CO2 Enrichment in a Mediterranean-type environment. Plant Biology, 22(S1), 123–132.
Parvin, S., Uddin, S., Tausz-Posch, S., et al. (2019b). Elevated CO2 improves yield and N2 fixation but not grain N concentration of faba bean (Vicia faba L.) subjected to terminal drought. Environmental and Experimental Botany, 165, 161–173. https://doi.org/10.1016/j.envexpbot.2019.06.003
Petrusán, J. I., Rawel, H., & Huschek, G. (2016). Protein-rich vegetal sources and trends in human nutrition: A review. Current Topics in Peptide and Protein Research, 17, 1–19.
Piñeiro, V., Arias, J., Dürr, J., et al. (2020). A scoping review on incentives for adoption of sustainable agricultural practices and their outcomes. Nature Sustainability, 3(10), 809–820. https://doi.org/10.1038/s41893-020-00617-y
Ponisio, L. C., Gonigle, L. K. M., Mace, K. C., et al. (2015). Diversification practices reduce organic to conventional yield gap. Proceedings of the Royal Society B Biological Sciences, 282, 20141396. https://doi.org/10.1098/rspb.2014.1396
Poore, J., & Nemecek, T. (2018). Reducing food’s environmental impacts through producers and consumers. Science, 360(6392), 987–992. https://doi.org/10.1126/science.aaq0216
Popkin, B. M., Corvalan, C., & Grummer-Strawn, L. M. (2020). Dynamics of the double burden of malnutrition and the changing nutrition reality. The Lancet, 395(10217), 65–74. https://doi.org/10.1016/S0140-6736(19)32497-3
Popoola, J. O., Ojuederie, O. B., Omonhinmin, C. A., et al. (2019). Neglected and underutilized legume crops: Improvement and future prospects. In F. Shah, Z. Khan, & A. Iqbal (Eds.), Recent advances in grain crops research. IntechOpen. https://doi.org/10.5772/intechopen.87069
Popp, J., Pető, K., & Nagy, J. (2013). Pesticide productivity and food security. A review. Agronomy for Sustainable Development, 33(1), 243–255. https://doi.org/10.1007/s13593-012-0105-x
Profeta, A. (2019). The impact of health claims in different product categories. Journal of International Food & Agribusiness Marketing, 32(2), 123–140. https://doi.org/10.1080/08974438.2019.1599753
Profeta, A., Balling, R., & Enneking, U. (2006). Geschützte HerkunftsangabenStatus Quo und Entwicklung der Nutzung der Verordnung (EG) 510/06 › GJAE - German Journal of Agricultural Economics - DE. German Journal of Agricultural Economics, 55(8), 353–358. https://www.gjae-online.de/de/articles/geschutzte-herkunftsangabenstatus-quo-und-entwicklung-der-nutzung-der-verordnung-eg-510-06/
Profeta, A., Enneking, U., & Balling, R. (2007). Wahrnehmung von regionalen Lebensmittelspezialitäten in Deutschland – Eine deutschlandweite, repräsentative Konsumentenbefragung. Berichte Über Landwirtschaft: Zeitschrift Für Agrarpolitik Und Landwirtschaft, 85(2), 238–251.
Quesada, N., Iannetta, P. P. M., White, P. J., et al. (2019). What evidence exists on the effectiveness of the techniques and management approaches used to improve the productivity of field grown tomatoes under conditions of water-, nitrogen- and/or phosphorus-deficit? A Systematic Map Protocol. Environmental Evidence, 8, 26. https://doi.org/10.1186/s13750-019-0172-4
Rai, K. K., Pandey, N., Meena, R. P., et al. (2021). Biotechnological strategies for enhancing heavy metal tolerance in neglected and underutilized legume crops: A comprehensive review. Ecotoxicology and Environmental Safety, 208, 111750. https://doi.org/10.1016/j.ecoenv.2020.111750
Rao, N. K., & Shahid, M. (2016). Neglected and underutilized crops for sustainable agricultural production in marginal areas. In R. A. Hall & P. Rudebjer (Eds.), Proceeding of the 3rd international conference on neglected and under-utilized crops for food secure Africa, 25–27 September, Accra, Ghana (pp. 130–139). Bioversity International.
Rao, N. K., Shahid, M., Al-Shankiti, A., & Elouafi, I. (2014). Neglected and underutilized species for food and income security in marginal environments. Acta Horticulturae, 1051, 91–104.
Ravelombola, W., Shi, A., & Weng, Y. (2018). Association analysis of salt tolerance in cowpea (Vigna unguiculata (L.) Walp) at germination and seedling stages. Theoretical and Applied Genetics, 131, 79. https://doi.org/10.1007/s00122-017-2987-0
Razi, K., Bae, D.-W., & Muneer, S. (2021). Target-based physiological modulations and chloroplast proteome reveals a drought resilient rootstock in okra (Abelmoschus esculentus) genotypes. International Journal of Molecular Sciences, 22(23), 12996. https://doi.org/10.3390/ijms222312996
Reif, T. M., Zikeli, S., Rieps, A. M., et al. (2021). Reviving a neglected crop: A case study on lentil (Lens culinaris Medikus subsp. culinaris) cultivation in Germany. Sustainability, 13(1), 133. https://doi.org/10.3390/SU13010133
Rivero, R. M., Mittler, R., Blumwald, E., et al. (2022). Developing climate-resilient crops: Improving plant tolerance to stress combination. The Plant Journal, 109, 373–389. https://doi.org/10.1111/tpj.15483
Rocchi, L., Boggia, A., & Paolotti, L. (2020). Sustainable agricultural systems: A bibliometrics analysis of ecological modernization approach. Sustainability (Switzerland), 12(22), 1–16. https://doi.org/10.3390/su12229635
Rogelj, J. D., Shindell, K., Jiang, S., et al. (2018). Mitigation pathways compatible with 1.5°C in the context of sustainable development. In V. Masson-Delmotte, P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, & T. Waterfield (Eds.), Global warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. In Press. https://www.ipcc.ch/site/assets/uploads/sites/2/2019/02/SR15_Chapter2_Low_Res.pdf
Ronga, D., Caradonia, F., Vitti, A., et al. (2021). Agronomic comparisons of heirloom and modern processing tomato genotypes cultivated in organic and conventional farming systems. Agronomy, 11, 349. https://doi.org/10.3390/agronomy11020349
Rouphael, Y., & Colla, G. (2020a). Editorial: Biostimulants in agriculture. Frontiers in Plant Science, 1, 40. https://doi.org/10.3389/fpls.2020.00040
Rouphael, Y., & Colla, G. (2020b). Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy, 10, 1461. https://doi.org/10.3390/agronomy10101461
Rubiales, D., & Flores, F. (2020). Adaptation of one-flowered vetch (Vicia articulata Hornem.) to Mediterranean rain fed conditions. Agronomy, 10(3), 383. https://doi.org/10.3390/agronomy10030383
Rubiales, D., Fondevilla, S., Chen, W., et al. (2015). Achievements and challenges in legume breeding for pest and disease resistance. Critical Reviews in Plant Science, 34, 195–236. https://doi.org/10.1080/07352689.2014.898445
Rubiales, D., Emeran, A. A., & Flores, F. (2020). Adaptation of grass pea (Lathyrus sativus) to Mediterranean environments. Agronomy, 10(9), 1295; doi:10.3390/agronomy10091295.
Rubiales, D., Annicchiarico, P., Vaz Patto, M. C., & Julier, B. (2021). Legume breeding for the agroecological transition of global agri-food systems: A European perspective. Frontiers in Plant Science, 12, 782574. https://doi.org/10.3389/fpls.2021.782574
Russel, S. J., & Norvig, P. (2021). Artificial intelligence: A modern approach (4th ed.). Hoboken Person. ISBN 9780134610993.
Sabatino, L., Iapichino, G., Consentino, B. B., et al. (2020). Rootstock and arbuscular mycorrhiza combinatorial effects on eggplant crop performance and fruit quality under greenhouse conditions. Agronomy, 10, 693. https://doi.org/10.3390/agronomy10050693
Saini, S., Saxena, S., Samtiya, M., et al. (2021). Potential of underutilized millets as nutri-cereal: An overview. Journal of Food Science and Technology, 58, 4465–4477. https://doi.org/10.1007/s13197-021-04985-x
Sardana, V., Mahajan, G., Jabran, K., et al. (2017). Role of competition in managing weeds: An introduction to the special issue. Crop Protection, 95, 1–7.
Savvas, D., & Gruda, N. (2018). Application of soilless culture technologies in the modern greenhouse industry - A review. European Journal of Horticultural Science, 83, 280–293. https://doi.org/10.17660/eJHS.2018/83.5.5
Scarano, A., Semeraro, T., Chieppa, M., et al. (2021). Neglected and underutilised plant species (NUS) from the Apulia Region worthy of being rescued and re-included in daily diet. Horticulturae, 7(7), 177. https://doi.org/10.3390/horticulturae7070177
Schlegel, P., Wyss, U., Arrigo, Y., et al. (2016). Mineral concentrations of fresh herbage from mixed grassland as influenced by botanical composition, harvest time and growth stage. Animal Feed Science and Technology, 219, 226–233.
Scottish Government. (2021). Cereal and oilseed rape harvest: Final. https://www.gov.scot/publications/cereal-oilseed-rape-harvest-2021-final-estimates/. Accessed 31 Jan 2022.
Scottish Whisky Association. (2021). Facts and figures. https://www.scotch-whisky.org.uk/insights/facts-figures/. Accessed 31 Jan 2022.
Shah, F., & Wu, W. (2019). Soil and crop management strategies to ensure higher crop productivity within sustainable environments. Sustainability, 11, 1485. https://doi.org/10.3390/su11051485
Shahzad, M., Hussain, M., Jabran, K., et al. (2021). The impact of different crop rotations by weed management strategies’ interactions on weed infestation and productivity of wheat (Triticum aestivum L.). Agronomy, 11, 2088. https://doi.org/10.3390/agronomy11102088
Shannon, M. C., & Grieve, C. M. (1999). Tolerance of vegetable crops to salinity. Scientia Horticulturae, 78, 5–36.
Shayanowako, A., Morrissey, O., Tanzi, A. S., et al. (2021). African leafy vegetables for improved human nutrition and food system resilience in Southern Africa: A scoping review. Sustainability, 13, 2896.
Siddique, K. H. M., Li, X., & Gruber, K. (2021). Rediscovering Asia’s forgotten crops to fight chronic and hidden hunger. Nature Plants, 7, 116–122. https://doi.org/10.1038/s41477-021-00850-z
Siracusa, L., Avola, G., Patanè, C., et al. (2013). Re-evaluation of traditional Mediterranean foods. The local landraces of ‘Cipolla di Giarratana’ (Allium cepa L.) and long-storage tomato (Lycopersicon esculentum L.): Quality traits and polyphenol content. Journal of the Science of Food and Agriculture, 93(14), 3512–3519.
Skendžić, S., Zovko, M., Živković, I. P., et al. (2021). The impact of climate change on agricultural insect pests. Insects, 12, 440. https://doi.org/10.3390/insects12050440
Slabbert, R., & van den Heever, E. (2006). Selection of traditional crops for improved drought tolerance in leafy amaranth: Moving towards sustainable food supply. In International conference on indigenous vegetables and legumes prospectus for fighting poverty, hunger malnutrition (Vol. 752, pp. 281–286). ISHS.
Smetana, S., Mathys, A., Knoch, A., et al. (2015). Meat alternatives: Life cycle assessment of most known meat substitutes. The International Journal of Life Cycle Assessment, 20(9), 1254–1267. https://doi.org/10.1007/s11367-015-0931-6
Smetana, S., Spykman, R., & Heinz, V. (2021). Environmental aspects of insect mass production. Journal of Insects as Food and Feed, 7(5), 553–571. https://doi.org/10.3920/JIFF2020.0116
Soares, J., Deuchande, T., Valente, L., et al. (2019). Growth and nutritional responses of bean and soybean genotypes to elevated CO2 in a controlled environment. Plants, 8, 465. https://doi.org/10.3390/plants8110465
Souza, L. A., López Andrade, S. A., Ribeiro Souza, S. C., et al. (2013). Evaluation of mycorrhizal influence on the development and phytoremediation potential of Canavalia Gladiata in Pb-contaminated soils. International Journal of Phytoremediation, 15, 465–476.
Springmann, M., Clark, M. A., Mason-D’Croz, D., et al. (2018). Options for keeping the food system within environmental limits. Nature, 562, 519–525. https://doi.org/10.1038/s41586-018-0594-0
Stagnari, F., Maggio, A., Galieni, A., & Pisante, M. (2017). Multiple benefits of legumes for agriculture sustainability: an overview. Chemical and Biological Technologies in Agriculture, 4(1), 1–13.
Stamp, P., Messmer, R., & Walter, A. (2012). Competitive underutilized crops will depend on the state funding of breeding programmes: An opinion on the example of Europe. Plant Breeding, 131(4), 461–464. https://doi.org/10.1111/j.1439-0523.2012.01990.x
Steffen, W., Richardson, K., Rockström, J., et al. (2015). Planetary boundaries: Guiding human development on a changing planet. Science, 347(6223), 1259855.
Steinfeld, H., Gerber, P., Wassenaar, T., et al. (2006). Livestock’s long shadow. FAO of the UN.
Stošić, M., Ivezić, V., & Tadić, V. (2021). Tillage systems as a function of greenhouse gas (GHG) emission and fuel consumption mitigation. Environmental Science and Pollution Research, 28(13), 16492–16503. https://doi.org/10.1007/s11356-020-12211-y
Tello, A., Aganovic, K., Parniakov, O., et al. (2021). Product development and environmental impact of an insect-based milk alternative. Future Foods, 4. https://doi.org/10.1016/J.FUFO.2021.100080
Teuteberg, H. J. (2006). Der Verzehr von Nahrungsmitteln in Deutschland pro Kopf und Jahr seit Beginn der Industrialisierung (1850–1975). https://doi.org/10.4232/1.8252.
Thanh, P. T., Sripichitt, P., Chanprame, S., et al. (2006). Transfer of drought resistant character from wild rice (Oryza meridionalis and Oryza nivara) to cultivated rice (Oryza sativa L.) by backcrossing and immature embryo culture. Kasetsart Journal of Social Sciences, 40, 582–594.
Tonn, B., Thumm, U., & Claupein, W. (2010). Semi-natural grassland biomass for combustion: Influence of botanical composition, harvest date and site conditions on fuel composition. Grass and Forage Science, 65, 383–397.
Tran, F., Holland, J. E., Quesada, N., et al. (2021). What evidence exists on the effectiveness of the techniques and management approaches used to improve the productivity of field grown tomatoes under conditions of water-, nitrogen- and/or phosphorus-deficit? A systematic map. Environmental Evidence, 10, 14. https://doi.org/10.1186/s13750-021-00229-9
Traoré, H., Barro, A., Yonli, D., et al. (2020). Water conservation methods and cropping systems for increased productivity and economic resilience in Burkina Faso. Water (Switzerland), 12(4), 976.
Uhl, P., & Brühl, C. A. (2019). The impact of pesticides on flower-visiting insects: A review with regard to European risk assessment. Environmental Toxicology and Chemistry, 38(11), 2355–2370.
von Grebmer, K., Saltzman, A., Birol, E., et al. (2014). Global hunger index: The challenge of hidden hunger. Welthungerhilfe, International Food Policy Research Institute, and Concern Worldwide.
Van Delden, S. H., SharathKumar, M., Butturini, M., et al. (2021). Current status and future challenges in implementing and upscaling vertical farming systems. Nature Food, 2, 944–956. https://doi.org/10.1038/s43016-021-00402-w
Van Oosterom, E. J., O’Leary, G. J., Carberry, P. S., et al. (2002). Simulating growth, development, and yield of tillering pearl millet. III. Biomass accumulation and partitioning. Field Crops Research, 79(2–3), 85–106.
Vaz Patto, M. C., & Rubiales, D. (2014). Lathyrus diversity: Available resources with relevance to crop improvement. Annals of Botany, 113(6), 895–908. https://doi.org/10.1093/aob/mcu024
Vaz Patto, M. C., Amarowicz, R., Aryee, A., et al. (2015). Achievements and challenges in improving the nutritional quality of food legumes. Critical Reviews in Plant Sciences, 34, 105–143. https://doi.org/10.1080/07352689.2014.897907
Vermeulen, S., Campbell, B. M., & Ingram, J. S. I. (2012). Climate change and food systems. Annual Review of Environment and Resources, 37, 195–222. https://doi.org/10.1146/annurev-environ-020411-130608
Vernooy, R. (2021). Action research on neglected and underutilised species. A methodological guide. The Alliance of Bioversity International and CIAT.
Vidigal, P., Duarte, B., Cavaco, A. R., et al. (2018). Preliminary diversity assessment of an undervalued tropical bean (Lablab purpureus (L.) Sweet) through fatty acid profiling. Plant Physiology and Biochemistry, 132, 508–514.
Vijayalakshmi, D., Geetha, K., Gowda, J., et al. (2010). Empowerment of women farmers through value addition on minor millets genetic resources in Karnataka. Indian Journal of Plant Genetic Resources, 23(1), 132–135.
Villegas-Fernández, Á. M., Sillero, J. C., Emeran, A. A., et al. (2011). Multiple-disease resistance in Vicia faba: Multi-environment field testing for identification of combined resistance to rust and chocolate spot. Field Crops Research, 124, 59–65.
Wang, S. T., & Ebert, A. W. (2013). Breeding of leafy amaranth for adaptation to climate change. In R. Holmer, G. Linwattana, P. Nath, & J. D. H. Keatinge (Eds.), High value vegetables in Southeast Asia: Production, supply and demand; proceedings of the SEAVEG 2012 regional symposium (pp. 36–43). AVRDC - The World Vegetable Center.
Waramit, N., Moore, K. J., & Fales, S. L. (2012). Forage quality of native warm-season grasses in response to nitrogen fertilization and harvest date. Animal Feed Science and Technology, 174, 46–59.
Weinberger, K. (2003). Impact analysis of Mungbean research in South and Southeast Asia. Final report of GTZ Project. Shanhua, Taiwan. https://assets.publishing.service.gov.uk/media/57a08b1ced915d3cfd000b38/DFID_impact_case_study_Mungbean_FINAL_1_.pdf
Weinberger, K., & Swai, I. (2006). Consumption of traditional vegetables in Central and North-Eastern Tanzania. Ecology of Food and Nutrition, 45, 87–103.
Wilker, J., Navabi, A., Rajcan, I., et al. (2019). Agronomic performance and nitrogen fixation of heirloom and conventional dry bean varieties under low-nitrogen field conditions. Frontiers in Plant Science, 10, 952. https://doi.org/10.3389/fpls.2019.00952
Will, M. (2008). Promoting value chains of neglected and underutilized species for pro-poor growth and biodiversity conservation: Guidelines and good practices. Bioversity International; Global Facilitation Unit for Underutilized Species. isbn:978-92-9043-775-8. Available on-line: https://cgspace.cgiar.org/handle/10568/104850
Willett, W., Rockström, J., Loken, B., et al. (2019). Food in the Anthropocene: The EAT-lancet commission on healthy diets from sustainable food systems. Lancet (London, England), 393(10170), 447–492. https://doi.org/10.1016/S0140-6736(18)31788-4
Wilson, D. M., Dalluge, D. L., Rover, M., et al. (2013). Crop management impacts biofuel quality: Influence of switchgrass harvest time on yield, nitrogen and ash of fast pyrolysis products. Bioenergy Research, 6, 103–113.
Wirsig, A., Profeta, A., Häring, A., et al. (n.d.). Branding of rural regions and autochthon agricultural products linked to their terroir Section 2. Agricultural Economics and Rural Sociology Branding of rural regions and autochthon agricultural products linked to their terroir.
Wirsig, A., Profeta, A., Häring, A., & Cerjak, M. (2011). Branding of rural regions and autochthon agricultural products linked to their terroir. Proceedings. 43rd Proceedings. 46th Croatian and 6th Croatian and 3rd International Symposium on Agriculture. Opatija. Croatia (301-305) Agriculture. Opatija. Croatia (301- XXX), January, 301–305.
Wright, D., Neupane, S., Heidecker, T., et al. (2021). Understanding photothermal interactions will help expand production range and increase genetic diversity of lentil (Lens culinaris Medik.). Plants, People and Planet, 3, 171–181. https://doi.org/10.1002/ppp3.10158
Yachi, S., & Loreau, M. (1999). Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America, 96, 1463–1468. https://doi.org/10.1073/pnas.96.4.1463
Zhai, Z., Martínez, J. F., Beltran, V., et al. (2020). Decision support systems for agriculture 4.0: Survey and challenges. Computers and Electronics in Agriculture, 170. https://doi.org/10.1016/j.compag.2020.105256
Zhao, J., Zeng, Z., He, X., et al. (2015). Effects of monoculture and mixed culture of grass and legume forage species on soil microbial community structure under different levels of nitrogen fertilization. European Journal of Soil Biology, 68, 61–68. https://doi.org/10.1016/j.ejsobi.2015.03.008
Acknowledgements
This research was supported by the European Union’s Horizon 2020 Research and Innovation Programme through the project “Realising Dynamic Value Chains for Underutilised Crops” (RADIANT), Grant Agreement number 101000622. The authors would also like to thank the scientific collaboration under the FCT project UIDB/50016/2020. in. The James Hutton Institute (CH and PPMI) are supported by the “Rural and Environmental Science and Analytical Services” (RESAS), a Division of the Scottish Government.
Conflicts of Interest
The authors declare no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Pinto, E. et al. (2023). Healthier and Sustainable Food Systems: Integrating Underutilised Crops in a ‘Theory of Change Approach’. In: Galanakis, C.M. (eds) Biodiversity, Functional Ecosystems and Sustainable Food Production. Springer, Cham. https://doi.org/10.1007/978-3-031-07434-9_9
Download citation
DOI: https://doi.org/10.1007/978-3-031-07434-9_9
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-07433-2
Online ISBN: 978-3-031-07434-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)