
Abstract
Angiogenesis is an essential process for tumor growth, progression, and metastasis, and it is one of the hallmarks of cancer. In addition to being structurally atypical, tumor blood vessels exhibit distinctly abnormal molecular phenotypes compared to their normal counterparts. As noticed by Aird: “The phenotypes of endothelial cells vary in structure and function, in space and time, and health and disease.” The palette of tumor endothelial phenotypes results from the specific conditions that guide the formation of these vessels, including activation of specific signaling pathways and a range of environmental pressures.
The focus of this chapter is to outline the roles endothelial cells play in basic physiological and pathological processes, to provide an overview of the mechanisms of dysregulated tumor angiogenesis, and to point out well-established and some novel molecular markers and phenotypes of tumor endothelial cells. We list key experimental and clinical studies that discuss the clinical relevance of specific molecular markers in predicting prognosis and therapy response, supporting the importance of tumor-associated angiogenesis in pathological processes such as metastasis. In addition to this, we discuss novel therapeutic approaches based on exploiting the molecular specificity of tumor endothelial cells to provide selective and efficient therapies.

Pathological stimulation of endothelial cells in tumors induces abnormal vascular phenotypes with altered function. Molecular markers of tumor endothelial cells can aid in predicting prognosis and response to therapy in cancer
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
FormalPara Take-Home Lessons-
Endothelial cells actively control the function of specific organs, acting as “gatekeepers” of their microenvironment
-
Blood vessel formation in physiological processes results in the formation of mature and fully functional vessels
-
Initiation of angiogenesis is an important requirement for tumor growth and progression
-
Tumor vessels are morphologically and molecularly abnormal with significant heterogeneity
Introduction
Tumor blood vessels display markedly abnormal phenotypes and are molecularly distinct from normal blood vessels. Under healthy conditions, endothelial cells act as gatekeepers of tissue homeostasis. They line the inner surface of all blood and lymphatic vessels, where they create a semipermeable barrier between blood or lymph and its surrounding tissues. This single cell layer is a highly metabolically active organ, and dynamic endothelial cell phenotypes constantly shift in response to cues within the extracellular environment, governing the needs of the tissues they serve. Thus, signals generated during homeostasis, hypoxia, inflammation, and repair are pivotal for the endothelial cell capacity to maintain blood fluidity, regulate blood flow and control trans-endothelial extravasation of solutes, macromolecules, hormones, and circulating immune cells. Accordingly, a growing body of evidence suggests that tumor cells and their microenvironment induce characteristic phenotypic changes in the surrounding tumor vasculature, reflected in unique gene expression profiles when compared to normal endothelial cells. However, different tumor types and stages of progression, show diverse patterns of altered gene expression, and no unique tumor endothelial marker has so far been proven to be suitable as a durable target for anticancer therapy. In recent years, novel tools have made it possible to analyze the tumor microenvironment at a much higher resolution, providing hope that the near future may bring improved prediction of therapy response and perhaps better patient outcomes.
Healthy Endothelial Cells Are Gatekeepers of Tissue Homeostasis
Endothelial Cell Phenotype Vary Across Vascular Beds, Branches, and Activation Status
The vascular network, often referred to as the vascular tree, forms a highly branched closed circulation that extends into all organs, nourishes every tissue, and provides a gateway for extravasation of fluids, solutes, macromolecules, hormones, and circulating immune cells. Microvascular beds, composed of arterioles, capillaries and postcapillary venules, make up the greatest surface of the vascular circulation. This is also where most physiological processes brought on by the endothelium occur [1], and, accordingly, where tumor-induced vascular pathology most commonly manifests.
Endothelial phenotypes change along the vascular tree, in line with the functionality of the vascular segments. Briefly, the arterial segment of the circulation is responsible for delivering nutrients and oxygen, the venous segment predominantly takes part in waste management, and the thin-walled capillary network represents the main site for exchange of gases and molecules between blood and tissues. To accommodate the different requirements imposed by these functional differences, arterial, venous, and capillary endothelial cells differ both on the structural and molecular levels [2].
The tissue microenvironment drives endothelial heterogeneity by two distinct mechanisms. Firstly, cell-derived, biochemical and biomechanical signals contribute to activation of specific genes and signaling pathways, in turn producing tissue-specific phenotypes. For example, blood pressure, blood flow, shear stress, blood-gas concentration, pH, and plasma components of arterial and venous blood are very different, all providing input that contributes to the heterogeneity of endothelial cells (EC) lining the different vascular segments. Secondly, specific traits of the microenvironment may induce epigenetic modifications in endothelial cells that cause site-specific heterogeneity [3]. Such epigenetic signatures, like those contributing to arterial or venous endothelial profiles, are generally more robust than those imposed by dynamic signals.
Structural heterogeneity can be examined by light and electron microscopy and includes variations in cellular morphology, thickness, nuclear orientation, size, surface properties, and the types of cell junctions present [2, 4]. The thickness of endothelial cells varies between 0.1 μm in capillaries and 1 μm in the aorta. Moreover, while commonly flat, endothelial cells in the high endothelial venules of secondary lymphoid organs take on a cuboidal morphology [1]. Endothelial cells tend to be larger in big veins compared to big arteries. Nuclei of endothelial cells lining big arterial vessels, like the aorta, are positioned downstream related to the blood flow, opposite from big vein vessels where endothelial nuclei are positioned upstream relative to the middle cell axis [1].
It has been already discussed that the phenotype of the endothelial cells can vary significantly between different tissue types, allowing tissue-specific endothelial cells to exert their specific functions [5, 6]. Depending on the tissue type they are isolated from, EC has been shown to respond differently to different signaling molecules. Endothelial cells isolated from different segments of the vascular tree display specific metabolic signatures and vary in their response to stimulation. In one study, endothelial cells isolated from coronary arteries, coronary veins, and the capillary network showed different capabilities to produce prostaglandins [2, 7]. Another study from Johnson et al. [2, 8] found significantly higher angiotensin converting enzyme activity in endothelial cells isolated from the arterial wall than in venous endothelial cells. Conversely, endothelial cells isolated from large veins produced significantly more tissue plasminogen activator (t-PA) compared to endothelial cells from arteries. Differences have been noted in the endothelial response to vasoactive molecules and inflammatory cytokines [2], with a more intense response being observed in postcapillary venules, in line with their prominent function in tissue leukocyte recruitment.
In addition to the variation found along the vascular tree, endothelial phenotypes also vary between different tissues to accommodate tissue-specific vascular functions [5, 6]. Tissue-based heterogeneity is most pronounced in the capillary segment [9], and microvascular endothelial cells isolated from different anatomical sites demonstrate distinct gene expression profiles [10]. While endothelial cells in some organs, like brain, liver, and kidney, express unique sets of genes, endothelial cells in other organs, like heart and aorta, display a more generalized endothelial signature [11].
The interaction between endothelial cells and their tissue-specific microenvironment starts during development and continues throughout life in a two-way process. First, endothelial cells actively contribute to specific organ function, acting as “gatekeepers of their microenvironment” [5, 12]. Endothelial cells take active part in guiding and controlling tissue development, during the embryonic stage and postnatally, before proceeding to control tissue homeostasis and regeneration in adulthood. For example, during brain development, endothelial cells contribute to regulation of neuronal differentiation [13, 14], while in adulthood they take part in maintaining the blood–brain barrier. On the other hand, the microenvironment also provides input that shapes the colorful range of EC phenotypes.
Hence, it is not surprising that endothelial cells also differ in their response to signaling molecules, depending on their tissue of origin. For example, lung endothelial cells are highly specialized in gas exchange and at the same time assist in providing a prompt immune response, in line with their proximity to the external environment [15]. Moreover, endothelial cells in the heart muscle specialize in providing a prompt and regular supply of fatty acids required for proper cardiomyocyte metabolism [6]. In order to investigate and further understand the nature and molecular origin of tissue-specific endothelial heterogeneity, Jambusaria et al. performed a study using the RiboTag transgenic mouse model that enables direct isolation of tissue-specific ribosome-associated mRNAs from complex tissues without cell dissociation [5]. This study provided surprising insight into the molecular milieu responsible for shaping tissue-specific EC phenotypes. The analyses revealed that genes responsible for physiological processes typical for neuronal cells, such as neurotransmitter transport and axon development, were highly expressed by endothelial cells of the brain. Similarly, endothelial-enriched genes in lung tissue were related to immune function and endothelial-enriched genes in the myocardium to myofiber assembly, muscle tissue development, and myocardial contraction [5]. These findings are intriguing, because they suggest that a certain level of plasticity exists between endothelial cells and surrounding tissue-specific cell types. Another study, based on analysis of 100,000 single cells from the Tabula Muris study, also detected transcripts of hepatocyte and cardiomyocyte genes in endothelial cells of the respective organs, but found no evidence for endothelial expression of neuronal, kidney, or lung transcripts [11].
Endothelial Cells Are Essential Regulators of Vascular Function
Under physiological conditions, one of the main functions of endothelium is to form a tightly regulated barrier at the interface between the blood and the surrounding tissue. The position of endothelial cells is optimal for allowing them to respond to physical and chemical signals, either carried in the blood from distant anatomical sites or produced locally by tissue-resident cells. Activated endothelial cells produce a wide range of factors that in turn regulate vascular tone, cellular adhesion, blood fluidity, smooth muscle cell proliferation, and vessel wall inflammation and remodeling [16]. Moreover, in inflammation, endothelial cells take an active part in regulating vascular permeability and blood flow, the development of tissue edema and hyperemia, as well as tissue leukocyte recruitment [17] (Fig. 3.1).

Normal endothelial cells maintain a healthy tissue microenvironment by regulating blood flow, preventing coagulation and exerting strict control over the movement of substances and cells between blood and tissues
Endothelial cells regulate blood flow in close crosstalk with vascular smooth muscle cells [18]. Importantly, they produce and secrete the vasodilators nitric oxide (NO) and prostacyclin (PGI2) [19,20,21]. NO is released after stimulation of EC with a number of molecules, including bradykinin, angiotensin II, histamine, acetylcholine, adenine nucleotides, and arachidonic acid [22]. Apart from NO, EC also produce and secrete PGI2 that also causes relaxation of underlying smooth muscle cells [23]. In addition to mediating vasodilation, EC also play an active role in the opposing process of vasoconstriction through the synthesis of molecules with vasoconstrictor activity, including angiotensin II, endothelins, prostaglandin H2, thromboxane A2, and reactive oxygen species [24,25,26].
Endothelial cells also secrete a number of molecules, including NO and PGI2, that contribute to maintaining the balance between coagulation and anticoagulation. Both NO and PGI2 are major anticoagulatory signaling factors that exert their function by increasing cAMP levels in platelets to prevent their aggregation [27, 28]. Another anticoagulatory pathway supported by endothelial cells is the protein C/protein S pathway, initiated by the interaction between thrombomodulin (receptor on the surface of EC) and thrombin with the consequent activation of protein C. Protein C consequently inactivates coagulation factors VIIIa and Va [29].
Healthy endothelial cells also contribute to innate immune responses that protect the body from invading microorganisms and tissue damage. Inflammatory activated endothelial cells mount an efficient response that facilitates elimination of the underlying cause of inflammation. For example, stimulation by hypoxia activated thrombocytes or histamine induces the release of endothelial Weibel Palade bodies. These storage granules contain preformed von Willebrand factor (VWF) that induces a pro-coagulant state, as well as membrane-bound P-selectin and platelet activating factor that allows neutrophils recruitment within minutes [30, 31]. Furthermore, inflammatory signaling molecules and pathogen-associated molecules, such as inflammatory cytokines and endotoxin, induce and upregulate the expression of inflammatory molecules that further drives the endothelial phenotype toward efficient leukocyte recruitment, vascular leakiness and thrombus formation. Additionally, with production and secretion of the chemokines, endothelial cells play an important roles in trafficking dendritic cells and T cells. Namely, endothelial cells are capable of producing and secreting CCL5, also known by the acronym RANTES (Regulated on Activation, Normal T Expressed and Secreted), which is a very powerful T cell attractant [32, 33]. Additionally, activated endothelial cells are capable of synthetizing and secreting CCL21 which directly stimulates chemotactic migration of dendritic cells [33, 34]. Heparin sulfate is another molecule produced by endothelial cells which contributes to trafficking of both dendritic and T cells [33]. Activated endothelial cells express on their surface adhesion molecules, such as E-selectin, vascular cell-adhesion molecule 1 (VCAM1), and intercellular adhesion molecule 1 (ICAM1), well-known for their importance in binding and redirecting of immune cells [35]. This pro-inflammatory state facilitates the efficient elimination of invading microbes and damaged cells and tissues. However, if excessive or prolonged, it may detrimentally affect tissue function. Hence, endothelial cells have also developed mechanisms to limit inflammation and promote resolution. For example, in viral infections, endothelial sphingosine-1-phosphate-receptor signaling protects against cytokine storm and related mortality [36]. Endothelial cells also take part in crosstalk with other cell types and influence their response to stimuli. For example, in ischemia, endothelial cells secrete factors that stimulate macrophage polarization toward a phenotype that promotes tissue repair rather than inflammation [37].
Tumor Vasculature Is Dysfunctional and Contributes to Pathological Conditions
Tumor Blood Vessels Display Abnormal Phenotypes and Are Structurally Different from Normal Vessels
As a distorted reflection of their functions in maintaining tissue homeostasis in health and during infections, endothelial cells also play an important role in malignant processes. The vasculature in solid tumors is morphologically and physiologically abnormal, and this contributes to cancer cell extravasation and metastatic processes [38] (Fig. 3.2).

Tumor endothelial cells are dysfunctional and contribute to microenvironment that affects tumor growth
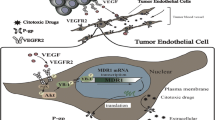
Blood vessel formation in physiological processes, such as embryonic development, growth, and wound healing, is a strictly controlled physiological process, which results in the formation of mature and fully functional vessels. This fine-tuned regulatory machinery is disrupted in tumors, due to the persisting presence of proangiogenic factors. One of these is VEGFA, a key driver of the abnormal tumor endothelial phenotype [39]. In this context, excessive VEGF signaling loosens the tight junctions between endothelial cells, inducing dysfunctional activation and vascular leakiness. Over time, the strong proangiogenic profile of the tumor microenvironment leads to the formation of aberrant and immature vasculature. Newly formed tumor vessels may fail to mature, and the clear distinction between arterioles, capillaries, and venules might be lost. Because of this dysregulation, tumor vessels are typically tortuous and branch irregularly in a dysfunctional network, often with the consequence of disordered blood flow [40,41,42]. This again results in poor perfusion and hypoxic areas within the tumor mass [43,44,45,46]. Although most tumors are densely vascularized, the malfunctional vasculature means that they are also usually hypoxic and nutrient-deprived [47], resulting in a vicious cycle that reinforces the proangiogenic microenvironment.
Tumor vasculature is morphologically and physiologically abnormal which contributes to cancer cell extravasation and metastatic process [38]. What is being observed in the tumor vasculature is that VEGF signaling causes loosening of the tight junctions between EC and makes such blood vessel prone to leakage. With addition of high interstitial fluid pressure and lack of pericyte lining, cancer cells are easily shad in the vasculature [48]. In addition to these mechanical shading mechanisms, it has been shown that EC can actively promote metastasis by secreting specific molecules and activating specific molecular pathways [38]. For example, it has been demonstrated that activation of Notch signaling pathway in tumor endothelial cells (TEC) contributes to inducing stemness in cancer cells together with inducing higher tumorogenicity, chemoresistance, and higher metastatic potential [49,50,51,52,53,54,55]. TEC secrete chemokines that has been known to be involved in tumor progression and metastasis [38]. In the study of Yadav et al. [56], it has been shown that TEC play a significant role in protecting CTC from anoikis. Namely, circulating TEC present in the cancer patients’ circulation have been reported to express high levels of adhesion molecules which bind to CTC and protect them from undergoing anoikis [56,57,58]. Maishi et al. have reported the role of TEC in the initiation of tumor metastasis [59]. They isolated two types of TEC, highly metastatic TEC (HM-TEC) and low metastatic TEC (LM-TEC) [60]. Their work demonstrated that high metastatic TEC have higher expression of angiogenesis related genes, higher genetic instability and they exert stem-like properties. They further demonstrated that HM-TEC have higher ability to attract and adhere to CTC when compared to LM-TEC and normal EC. HM-TEC had upregulated biglycan which has been suggested as a key molecule on TEC, that is enabling tumor cells to get through the vessel barrier into the circulation. In addition, biglicans are proved to be capable of binding to the toll-like receptor 2 (TLR2) and toll-like receptor 4 (TLR4) on tumor cells causing activation of NF-κB and ERK signaling and consequently unleashing their migratory potential [38]. In another study of Branco-Price et al. from 2012, it has been discussed the importance of hypoxia inducible transcription factors in regulation of metastatic processes. They concluded that the loss of HIF-1α in endothelial cells leads to reduction of NO synthesis, which consequently suppresses the migration of tumor cells through the endothelium and prevents metastasis. In the contrary of the loss of HIF-2α which has an opposite effect thus supporting the metastatic process [61].
Molecular Signatures of Tumor Endothelial Cells
Molecular Signatures and Markers of Tumor Endothelial Cells Are Heterogeneous
The structural differences between normal and tumor endothelial cells are reflected by distinct molecular characteristics [62,63,64,65,66,67]. As discussed in the previous section, both the tumor microenvironment and the range of mechanisms used by tumors to satisfy their needs for nutrients and oxygen exert pressure on endothelial cells. This influences endothelial gene expression and induces a range of variations from the normal endothelial profile. Furthermore, the transcriptional and molecular signatures of tumor endothelial cells differ in relation to tumor type, anatomical localization, different pathohistological stages, and even between different blood vessels in the same tumor [68, 69]. As stated by Aird: “the phenotypes of endothelial cells vary in structure and function, in space and time, and in health and disease” [3]. The specific molecular traits that make up this heterogeneity may either be triggered by the pathological process itself or arise from its underlying cause [70]. In this section, we will discuss specific molecular markers arising from different tumor endothelial phenotypes. Many of these molecular markers are also expressed by normal endothelial cells, where they take part in physiological processes, including development, angiogenesis, survival, and extracellular matrix (ECM) remodeling. Other markers suggest the presence of specific subpopulations, like endothelial progenitors or tumor cells involved in vascular mimicry. Finally, the expression level of pan-endothelial marker genes can be measured to indicate the fraction of endothelial cells and therefore the vascular density of a tumor [71].
Over the past few decades, many different methods have been used to discover and validate novel tumor endothelial markers. The range of tools includes histological studies, targeted techniques like immunostaining, flow cytometry and cell sorting by flow-assisted and magnetic techniques, experiments in cell culture and unbiased transcriptomic and proteomic analyses, including serial analysis of gene expression and, more recently, single cell transcriptomics. In the case of antibody-based imaging, the introduction of digital image analysis and digital scoring methods have improved quantitation and eliminated the subjectivity of manual analyses [72]. In this section, we will discuss a range of studies undertaken to reveal specific tumor endothelial signatures that could be used as prognostic and predictive markers.
Markers of Endothelial Progenitor Cells
Tumors are capable of recruiting the endothelial progenitor cells (EPC) from the bone marrow and guide their differentiation into mature endothelial cells [69]. The first identification of putative EPC was in 1997 by Asahara et al. [73]. Using two antigens (CD34 and the VEGF receptor Flk-1) shared between angioblasts and hematopoietic stem cells, the authors isolated putative angioblasts from peripheral blood. Both C34 and Flk-1 are expressed by all hematopoietic stem cells but are lost during differentiation of the hematopoietic lineages while remaining present in most adult EC. CD34+/Flk-1+ cells were isolated from the human peripheral blood using antibody-coated magnetic beads and then cultured in vitro using collagen and fibronectin-coated dishes [73]. Following this, further characterization of EPC was done by other authors. Yin et al. in 1997 in their study were using peripheral and umbilical cord blood, fetal liver and fetal and adult bone marrow from which they isolated CD34 positive cells and performed in vitro colony assays and in vivo engraftment assays. They recognized AC133 as a novel hematopoietic stem cell marker present on EPC but not on mature EC [74]. This was in concordance with Peichev et al., who demonstrated that a small subpopulation of CD34 cells isolated from umbilical cord blood and fetal liver using MACS immunomagnetic technique, express both AC133 and VEGFR-2. When they cultured these cells in the presence of VEGF, FGF-2, and collagen, EPC cells differentiated into AC133-/VEGFR-2+ mature EC [75]. In more recent years, the study by Romagnani et al. used CD14+ cells isolated from peripheral blood, and by using highly sensitive antibody-conjugated magnetofluorescent liposomes (ACMFL) technique, the authors concluded that almost all CD14+ cell from the bone marrow were also CD34low cells. These double positive CD14+/CD34low cells express other embryonic stemness markers such as Nanog and Oct-4 [76]. The idea of EPC generated in the bone marrow, being capable of entering circulation and taking part in neovascularization, was confirmed in human, dog, and mouse transplantation model studies of Shi et al; Asahara et al.; Lin et al.; and Kalka et al. [77,78,79,80,81]. Therefore, EPCs are usually characterized by the expression of vascular markers such as CD14, CD34, CD133, VEGFR1, Tie-2 (endothelial tyrosine kinase receptor), but also Oct-4 and Nanog. Additionally, they are characterized by the ability to uptake low-density lipoproteins and to bind ulex-lectin [69, 76, 82]. EPC’s role in tumor vascularization has been confirmed in a study by Asahara et al. from 1999, where the authors used mice that were subjected to bone marrow transplantation from transgenic mice constitutively expressing β- galactosidase gene regulated by an EC specific promoter. These mice were then inoculated with murine colon cancer cells and 3 weeks later, histological examination of the developed tumors has shown Lac-Z positive cells both in the tumor mass and integrated into the endothelium layer of tumor vessels [77, 78]. In years to come, there were multiple efforts to use circulating endothelial cells (CEC) as markers [83]. This idea was supported by numerous studies discussed in Bertolini et al. [84] where a number of approaches have been suggested (flow cytometry, positive enrichment using immunobeads, etc.) in order to follow the kinetics, number, and viability of CEC and CEP. These parameters have been recognized to correlate with the clinical outcome and prognosis in cancer patients receiving anti-angiogenic therapy [84]. Because there is still no perfect single marker identified as being suitable for this purpose, multiparametric single cell analysis is needed at this point. In 2010, the predictive value of CEC in colorectal cancer patients, undergoing bevacizumab-based combination therapy, has been confirmed in a study by Ronzoni et al. [85], where CEC levels in circulation correlated with the therapy response. In a study by Mehran et al. from 2014, an attempt has been made to identify the tumor specific CEC by observing tumor specific endothelial markers on their surface (TEM7/8), first in preclinical cancer models and afterwards in patients with esophageal and non-small cell lung cancer. They observed that CEC levels decreased after tumor resection [86]. Following this, there was another study from 2016 by Cima et al., where they reported identification of tumor-derived circulating endothelial cell clusters in early stage colorectal cancer patients, suggesting their use as noninvasive screening for colorectal cancer [87]. A study from 2017 by Rahbari et al. suggested that CEC are of higher prognostic value when detected in blood of patients suffering from metastatic colorectal cancer than detection of CTC [88] (Fig. 3.3).

Tumor endothelial cells display a range of molecular markers that can be used to characterize the tumor and predict prognosis and therapy response (endothelial cells in green, tumor cells in grey)
Markers of Vascular Mimicry
Vascular mimicry (VM) is a strategy mostly used by highly aggressive tumors [69, 89, 90]. In this case, tumor cells driven by hypoxia create vascular channels made of endothelial-like cancer stem cells (CSC) [89] which are capable of delivering oxygen and nutrients to the tumor tissue. There is a number of potential mechanisms that support the formation and occurrence of VM. For example, the role of EMT in VM forming cancer cells has been discussed [91]. It has been known that cells undergoing EMT lose some of the epithelial markers including α-catenin, E-cadherin, and zonula occludins-1, and at the same time they upregulated some of the mesenchymal markers, such as fibronectin, cadherin-2, vimentin, and VE-cadherin. VE-cadherin is proven to be crucial for VM formation [91]. The first identification of the vascular mimicry process goes back to 1999 by Maniotis et al. where they used tissue sections of uveal and melanoma origin to identify functional vascular-like channels, which were not giving staining for known vascular markers. They examined tissue samples histologically, using various microscopy techniques, including light and TEM microscopy for visualization of these, matrix embedded channels; and IHC panel for endothelial cell markers to confirm their phenotype as CD31, CD34, Flk-1, factor VIII, and ulex eurapeus I lectin negative cells [92]. Further molecular characterization revealed that these specific cells express CD271 (Low-affinity Nerve Growth Factor Receptor) also known as mesenchymal stem cell marker and they often lack expression of CD31 and VWf [89, 93]. Using the Crispr/CAS9 method, the importance of VE-Cadherin in the occurrence of VM has been supported [94]. Additionally, it has been revealed that in breast cancer, these endothelial-like cells overexpress HIF1α, EphA2, and VE-cadherin [89]. Aside from this, CSC characterized by the CD44+/CD24-profile and high expression of ALDH1 has been kept accountable for regulation of vascular mimicry in triple negative breast cancer [89]. In a meta-analysis from 2013, including 22 clinical studies, Cao et al. evaluated VM as a prognostic marker in 3062 patients and across 15 malignancies, including metastatic conditions of sarcoma, liver, lung, melanoma, and colon cancers. They concluded that VM+ cancers give a poor 5-year overall survival rate when compared to VM cancers [95]. Subsequently, there were other studies where VM has been investigated from the ground of cancer differentiation and metastatic potential [96] and as an unfavorable prognostic indicator in breast cancer [97]. In 2019, Zhang et al. performed a meta-analysis study to define a prognostic value of vascular mimicry in advanced melanoma patients. The authors found a significant association between VM and poor prognosis in melanoma patients [98]. They concluded that VM status can be relied on as an accurate prognostic biomarker when diagnosed using CD31-/Pas+ IHC staining, which gives a relatively accurate VM diagnostic value of 75% sensitivity and 70% specificity, compared to the less accurate CD34-/PAS+ staining or PAS+ staining alone [98].
Markers of Endothelial Trans-differentiation
Trans-differentiation of CSC to endothelial and smooth muscle-like cells has been confirmed in many different tumor types [99,100,101,102,103]. These cells often carry similar somatic mutations as the accompanying tumor epithelia confirming their malignant origin [103, 104]. In an example, this process has been confirmed in glioma CD133+ stem cells cultured in endothelial promoting media. As a result glioma cancer stem cells were forming tubular structures expressing VWF, CD34, CD144, CD31, Tie-2, endothelial NOS, and VEGFR2 [104, 105]. In the study by Bussolati et al., it has been demonstrated that CD105+ cells isolated from renal carcinomas have the possibility to differentiate into VWF/KDR/VEGFR3/CD31 positive endothelial cells in vitro and in murine models [106]. CD105 showed high correlation with other prognostic factors in breast cancer when used as a neovascularization marker [107]. In the study from 2014, CD105 has been evaluated and confirmed as a potential independent predictive marker in patients with clear-cell renal cell carcinoma after resection [108]. Aomatsu et al., in the study from 2012, demonstrated the predictive value of CD133 for neoadjuvant chemotherapy in breast cancer patients [109]. In addition, Kim et al. in 2015 published a study where they evaluated the potential predictive value of CD133 and ALDH1 in breast cancer patients [110].
General Tumor Endothelial Markers
The first observation of molecular heterogeneity in TEC was in 1992 when endosialin, also known as tumor endothelial marker (TEM) 1 or CD248, has been identified by Rettig et al. [111]. This discovery has been followed by the identification of prostate-specific membrane antigen on the membrane of TEC (normally not present on the surface of normal endothelial cells) [112]. Similar studies have revealed other TEC markers such as endothelial-specific molecule 1 (ESM1), endoglin (CD105), αvβ3 and α5β1 integrin [113,114,115,116]. Some other reports concluded that a number of genes appear to be downregulated in TEC, like intercellular adhesion molecule 1 (ICAM-1) and CD34 [117, 118].
Using the SAGE method (serial analysis of gene expression) on endothelial cells isolated from colorectal cancer and compared with normal colon tissue as a control, St. Croix et al. in 2000 have identified a total number of 46 transcripts that are more than tenfold upregulated in TECs compared to normal colon endothelial cells. This study yielded eight new TEMs [63]. TEMs have been extensively exploited as vascular prognostic markers in various tumor tissues [119,120,121]. For example, high expression of TEM7 has been related to metastasis and poor survival of osteogenic sarcoma patients [122]. Even though TEM markers have been defined as specific and reliable, additional studies showed that some of these newly identified markers, even in the beginning believed to be TEC specific [111], are not specific for TEC. TEM1 (endosialin) has been proven to be expressed in some fibroblasts, perivascular cells, and developing tissues [123, 124]. Additionally, TEM7 has been identified in the brain and sarcoma cell lines [125, 126] and TEM8 has been known to be present in the vessels of developing corpus luteum [63].
Nevertheless, the importance of TEM markers and their clinical relevance has been confirmed in the following years. In a study from 2015, Zhang et al. reported TEM7 as a key prognostic marker in resectable gastric cancer [127] and in the study from 2018, Czekierdowski et al. reported a prognostic significance of TEM7 and nestin in high grade serous ovarian cancer [128]. Further, TEM8 has been reported as a relevant prognostic marker in a number of studies. Gutwein et al. discussed the relevance of TEM8 as a marker in triple negative breast cancer being consistently expressed in significantly higher amounts in tumor tissue compared to healthy controls as well as in metastatic sites. Notably, TEM8 was not heavily expressed in the tumor-associated endothelium as confirmed by dual IHC, where the TEM8-positive cells did not show co-expression with SMA, calponin, CD31, or CD34 [129]. An interesting diagnostic approach using anti-TEM8 antibody labeled with 89Zr and immune-PET imaging was described by Kuo et al. in 2014 in a murine model [130]. The authors suggested that TEM8 targeted PET imaging could improve the diagnostic of angiogenic tumors, which might be susceptible to anti-TEM8 therapy. This type of PET imaging can also help in defining an appropriate therapeutic dose needed for optimal tumor uptake [130]. In the more recent study from 2020, Pietrzyk et al. concluded that TEM8 is a superior prognostic marker than the routinely used Ca 19–9 marker, exerting higher sensitivity and specificity. They were measuring serum levels of TEM8 pointing to its potential as an early diagnostic marker as well as a clinical predictor of progression and prognosis in patients suffering from colorectal cancers [131]. In addition to these studies, endoglin (CD105), another TEM marker, has been confirmed as a prognostic factor related to poor prognosis, metastasis and tumor recurrence in various cancers [132,133,134,135].
Microvascular mural cells (MMC) take part in the building of the microvasculature three together with endothelial cells. They consist of rather heterogeneous cell populations of pericytes, vascular smooth muscle cells, and intermediate cell types which are still not fully characterized [136]. These cells take part in the regulation of many biological functions but also play a major role in tumor pathology. Namely, it has been known that specific mural cells can have a particular effect on cancer therapy response and overall prognosis, the metastatic potential of the tumor and immune surveillance. Better characterization of these cells in the last few years conducted on murine metastasis models gave insights into the specific molecules and signaling pathways that can be related to “bad” versus “good” mural cell phenotypes. In an example, it has been known that mural microvascular cells expressing endosialin/CD248, KLF4, or having CD45-/VLA-1 bright phenotype are related to a higher risk of metastasis [136]. In addition to this, some studies pointed to the importance of MMC in premetastatic niches where metastasis supportive perivascular cells expressing KLF4 gene promoted metastasis by inducing fibronectin rich microenvironment [137, 138]. Expression of PDGFRb and NG2 are suggested as markers of “good” MMC phenotypes as they have been related to increased chemosensitivity and drug efficiency, better lymphocyte infiltration and better response to immunotherapy, and being antimetastatic [139,140,141,142,143,144,145]. On the other side MMC expressing desmin, Rgs5 and a-SMA are correlated to higher resistance to chemotherapy, loss of immunosurveillance and resistance to angiogenic therapy. In addition to this, a-SMA together with PDGFRb has been correlated with worse prognosis in a number of different studies [72, 137, 146,147,148,149,150].
The SAGE method has been used in studies on many other tumors such as breast cancer, ovarian cancer, and gliomas [62, 151,152,153,154]. These studies concluded that most of the overexpressed genes were tumor specific, or they were specific to high invasiveness and higher tumor grade, and a few of them were shared between different tumor types. For example, MMP9 has been connected to both ovary cancers and breast cancers. SPARC was shared between breast, colon, and brain tumors and HEYL between breast and colon cancers [3, 155]. Nevertheless, HEYL was particularly distinct to invasive breast cancer TEC, indicating stage and tumor type specificity of TEC molecular signature [3, 155]. In one glioma study, 14 endothelial markers have been identified in the malignant brain compared to a healthy brain, further confirming the existence of a TEC-specific transcriptional profile, different from the physiological signature. This has been supplemented by demonstrating that plasmalemmal vesicle associated protein 1 (PV-1), which is usually suppressed in the blood–brain barrier (BBB), is highly expressed in high grade gliomas and that this expression pattern is governed by glioma cells implying the high specificity and importance as a novel brain TEC marker [152, 156]. In a study conducted by Seaman et al., on healthy and tumor affected liver tissues, 13 genes were identified which were at least tenfold upregulated in malignant tissues compared to controls. Furthermore, additional studies on colorectal carcinoma, breast and ovarian cancers, confirmed the position of TEC molecular signatures being highly specific and distinct from the normal endothelial cells signature [153, 157,158,159].
All previous studies were generally facing the same limitations by being performed on the cell bulk which enabled discoveries of only a limited number of endothelial cell phenotypes [160,161,162,163]. Introducing robust single cell transcriptomic methods in endothelial cell studies allowed more detailed understanding of EC phenotypes and changed the perspective on tumor endothelial cells. In a study from 2020, Goveia et al. [68] described 16 previously unknown endothelial cell phenotypes in a lung tumor. The authors identified 2 novel capillary phenotypes that could be induced by cancer-derived cytokines and which were named scavenging capillaries, being characterized by upregulation of scavenging receptors (CD52, CD68) and genes related to macrophages and antigen presenting. They identified two distinct alveolar capillary EC phenotypes they named type I and type II. Type I was characterized by overexpression of endomucin (EMCN) and lower expression of von Willebrand factor (VWF) compared to type II, together with high expression of EDNRB and IL1RL1. Compared to all other clusters they identified, capillary EC expressed a signature of genes characteristic to MHC II antigen presenting cells, although they were lacking costimulatory molecules CD80 and CD86 [68]. Further, in the tumor endothelial cells, they identified tip EC (expressing high levels of tip cell marker CXCR4 and low expression of CD36, CA4, and HLA-II—capillary markers) that were expressing genes associated with migration, matrix remodeling, and VEGF signaling. Along with this, immature TEC phenotypes have been identified, similar to tip cells but characterized by overexpression of genes involved in vessel barrier integrity, vessel maturation, and Notch signaling, resembling stalk-like cells. TECs exert a specific phenotype of postcapillary veins EC (ACKR1high/VWF high) which upregulated immunomodulatory factors and ribosomal proteins corresponding to high endothelial venules in inflamed tissues including CCL14. By using CyTOF, the authors generally confirmed increased levels of capillary EC markers HLA-II and CD36 in normal EC but being downregulated in TEC [68]. The study gave some novel prognostic implications, suggesting that patients expressing high levels of gene signatures associated with angiogenic tip TEC, immature, activated postcapillary TEC or lymphatic TEC indicated worse prognosis and shorter overall survival.
Recently, in a study from 2021, Kahn et al. [71] designed a classifier for estimation of tumor vascularity that was named endothelial index (EI). In their approach, the authors used computer algorithms to correlate histological VD with mRNA expression data. The list of seven genes appearing as endothelial classifiers can quite precisely predict the VD, as confirmed in 31 different human malignancies. Genes used for this endothelial index (EI) consisted of angiopoietin 2 (ANGPT2), cadherin 5 (CDH5), ETS-related gene (ERG), endothelial cell selective adhesion molecule (ESAM), endothelial cell specific molecule 1 (ESM1), intracellular adhesion molecule 2 (ICAM2), and tyrosine kinase with immunoglobulin like domains 1 (TIE1). Although the EI score was very precise in estimating VD, it failed to deliver accurate prognostic information. As the authors suggest, one of the possible reasons for this is that in their study the observed 95% confidence interval (CI) was very wide when calculating the hazard ratio (HR) in relation to EI and overall survival. This observation was still in concordance with a study that pointed out the limitations of using VD alone in predicting response to anti-angiogenesis therapy [164]. Following this, the authors attempted in identifying specific signaling pathways related to variations in tumor vascularity and they focused on genes overexpressed in high EI tumors (EI score > 0.9—hypervascular tumors) and compared them with low EI tumors (EI score < 0.1—hypovascular tumors). Using EI score to stratify tumors as hyper/hypo vascularized, they concluded that VEGFA/VEGFR2 signaling is correlated to higher EI score but was not the only pathway that was strongly correlated with higher vascularization. Stratification on high versus low EI enabled them to the identification of multiple alternative signaling pathways which correlated strongly with high EI. This led to the identification of 24 hub genes in total, representing in the best manner the signaling networks of hypervascularized tumors. Using this information, the authors were able to group all tumor types they examined into six distinct vascular microenvironment signatures with specific signaling patterns. This vascular microenvironment approach was able to provide additional prognostic value beyond the use of VD and EI alone as it was based on a more robust metric of tumor–vessel signaling and mirrors comprehensive molecular and cellular interactions which are responsible for driving tumor vascularization. In the long run, this novel approach could enable more patient-oriented anti-vascular cancer therapy, with better therapy response prediction, promising the best possible results and therapy outcomes.
Tumor Endothelial-Specific Molecular Signatures and Signaling Pathways as Potential Therapeutic Targets
The Emergence of the Anti-angiogenic Therapy Concept
Cancer cells, being highly metabolically active, demand a high input of oxygen and nutrients from the blood and need to be in close proximity to the vasculature [69]. Led by this reasoning, Judah Folkman proposed the theory that the initiation of angiogenesis constitutes an important requirement for tumor growth and progression. Initial findings showed that more aggressive and quickly growing tumors are highly vascularized in comparison with less aggressive and slowly growing tumors [165]. This led Folkman to identify and isolate a tumor-derived angiogenic factor. His findings suggested that modulation of angiogenic signaling pathways to block tumor vessel formation could be a strategy to suppress tumor growth by starvation. In the following years, his work inspired many studies committed to identify and isolate tumor-derived angiogenic factors and describe their signaling pathways [166, 167]. The approval of this new family of anticancer drugs is undoubtedly one of the most important advances in the clinical oncology therapeutic approaches. Although, the high expectations has been given, these therapies often endure significant drawbacks. The first to prove the existence of VEGF independent tumors in 2003 was a study by Viloria-Petit et al. [168]. There is a number of molecules that are known to be secreted by the tumor mass and are able to promote angiogenesis. These molecules are VEGF, FGF2, angiopoietins, apelin, PDGF, and a variety of chemokines [69]. Many of these have important roles in physiological conditions. As in the beginning targeting VEGF gave promise and hope to the patients suffering from malignant diseases, very often the anti-angiogenic therapy failed to give expecting therapeutic results due to the number of mechanism tumor tissue avoid the therapeutic effect of the drugs, either by recruiting alternative ways of angiogenesis or by developing mechanism of resistance to the therapy.
As discussed above, a tumor can recruit additional tactics to fulfill its hunger for nutrients, which is even more evident when one of the angiogenesis pathways has been targeted. In addition to this, many times it is not perfectly clear if a patient expresses the specific drug target, or whether drug-target interaction will yield the desired therapeutic effect. Due to the high cost, of both drugs in use and the valuable reaction time patients and physicians have, it is of crucial importance to select and classify the patients according to their molecular signatures and identify the most fruitful molecular pathways which could be targeted. Although recent clinical trials data clearly point the value of anti-angiogenic drugs in prolonging the survival of patients, there are still challenges to meet when development of alternative therapeutic targets and their preclinical and clinical validations comes into focus [84].
VEGFR-Directed Therapies
As previously mentioned, VEGF is a well-known angiogenesis and vasculogenesis promotor both in cancer and physiological conditions. In a tumor, VEGF is often related to increased vessel density (VD), higher invasiveness and tumor progression, metastasis, and recurrence [69, 169]. VEGF is upregulated in hypoxic tumor conditions and regulates vessel formation by activation of VEGFR2 on endothelial cells [69, 170]. Activation of VEGFR2 causes the cascade of responses by various signaling pathways such as ERK and PI3K/Akt that as a consequence have endothelial cell proliferation, migration, invasion, and survival [69, 171, 172]. Endothelial invasion is regulated by expression of MMP-2, MMP-9, and urokinase plasmin activator which are helping in dissolving the basal membrane and ECM facilitating the formation of capillary sprouts [171, 173]. Dysregulated VEGF might as well play a significant role in metastasis by taking part in junctional remodeling, vascular hyperpermeability, and increased interstitial pressure which all together can allow the escape of tumor cells into the bloodstream [69, 174]. As previously mentioned, VEGF pathway has been very early recognized as an important regulator of tumor angiogenesis, and therefore efforts has been made to exploit it as antiangiogenic therapeutic target. Targeting this pathway has been looked upon as an effective approach, especially in solid tumors, as confirmed by numbers of preclinical and clinical studies [175]. This led to the development of multiple strategies to target this pathway which consists of molecules preventing VEGF binding to the receptors (bevacizumab, pegaptanib, VEGF trap) [176,177,178,179,180], antibodies against VEGFR-2 (ramucirumab) [181], and molecules which can inhibit the kinase activity of VEGFR-2 consequentially blocking the VEGF signaling pathway (sunitinib, pazapanib and sorafenib) [182,183,184,185,186,187]. There are 11 different drugs approved for targeting VEGF [188]. The whole complexity of the tumor angiogenic process is still not completely understood and there are alternative pathways involved; therefore, the main difficulty in anti VEGF-VEGFR therapy is that it is not the only angiogenic pathway active in tumors (Fig. 3.4).

Tumors can adapt alternative strategies to overcome insufficient perfusion and oxygenation. These include recruitment of endothelial progenitor cells from the bone marrow, vascular mimicry, and endothelial trans-differentation of cancer stem cells. (endothelial cells in green, tumor cells in grey)
Other Candidate Pathways for Anti-angiogenic Drugs
FGF2, also known as basic FGF (bFGF), exerts its function by binding to FGFR and has an important role in the regulation of many physiological functions. In tumors, it can increase cell proliferation and resistance to apoptosis. By increasing invasiveness and motility of cancer cells, regulating CSC as well as governing angiogenesis, it promotes metastasis [69, 189, 190]. FGF signals in a paracrine manner. After its release into the ECM, it induces secretion of matrix metalloproteinases (MMPs) and collagenases that cause ECM degradation. FGF also regulates endothelial metabolism through Myc-dependent glycolysis, which is proven to be essential for endothelial proliferation, sprouting, and migration [190, 191]. FGF2 can act as an alternative to VEGF orchestrated angiogenesis and a possible reason why VEGF therapies fail to give expected results, as suggested by a study where downregulation of FGF receptor restored tumor sensitivity to anti-VEGF therapies in a murine model [192] making FGF2-FGFR2 a potential therapeutic target targeting both angiogenesis and CSC [189, 193, 194]. Studies in mice were showing promising results in using combined ANGPT-2/VEGFR2 blockage, which in return slowed down the tumor growth, induced vessel normalization, and blocked macrophage recruitment consequently prolonging survival in glioma, breast cancer colorectal cancer, and renal cancer mouse models [69, 195, 196].
Ephrin signaling contributes to embryogenesis, tissue organization, cell migration, and vasculogenesis [69, 197,198,199]. In addition to its physiological roles, ephrin signaling has important roles in cancer angiogenesis, tumor progression, and metastasis. Dysregulation of ephrin signaling has been identified in a variety of cancers including breast, liver, brain, colon, melanoma, and prostate cancer [200,201,202]. The EphB2–ephB4 signaling axis was shown essential for tumor angiogenesis and progression in glioma, by altering vascular morphology, pericyte vessel coverage, and the resistance to antiangiogenic therapy [69, 203, 204]. Additionally, EphB2 signaling has been recognized as an important factor for proliferation and perivascular invasion of glioblastoma stem cells [205]. Ephrins A1/2 have been recognized as important markers of blood vessels in preclinical models of Kaposi sarcoma and breast carcinoma [201]. Their functional role and potential therapeutic target were proven by utilizing soluble EphA receptors that were able to induce a decrease of VD, reduce tumor volume, and cell proliferation [206,207,208].
Apelin—apelin receptor signaling increases in both endothelial cells and tumors in many malignancies. Increased expression of apelin has been shown to be a good predictor of disease progression and overall poor outcome [209,210,211,212,213,214,215,216]. Apelin expression is driven by tissue hypoxia and is known to promote tumor growth by direct stimulation of the tumor cell migration and metastasis [69, 211, 217,218,219,220]. Apelin pathway is an attractive therapeutic target, and some studies have shown usefulness in targeting apelin pathway in breast, glioma, and lung cancer, where it showed reduced tumor growth, reducing metastatic potential and improved vessel structure and function [221,222,223]. The study undertaken on renal cell carcinoma APLNR receptor overexpression in a group of patients was correlated with lower PD-L1 expression indicating its role in cancer immunomodulation [215].
Angiopoietins, aside from their physiological role in vasculogenesis, angiogenesis, and blood vessel remodeling, also have functions in tumor angiogenesis and tumor growth. The angiopoietin signaling pathway consists of three ligands, ANGPT-1, ANGPT-2, and ANGPT-4, which bind endothelial receptor tyrosine kinases, including TIE-1 and TIE-2 [69]. TIE-2 has an important role in both physiological and tumor processes, and its function depends on the ligand. Levels of ANGPT-2 in tumor-associated vessels increase in response to hypoxia and VEGF. Excessive ANGPT-2 has been also shown to cause decreased efficiency of anti-VEGF therapy in glioblastoma [69, 224, 225].
Tumors also adopt strategies that are not commonly part of physiological angiogenic processes to afford their increased oxygen and nutrient demands. Many tumors are capable of recruiting endothelial progenitor cells (EPC) from the bone marrow and guide their differentiation into mature endothelial cells [69]. Vascular mimicry (VM) is another tactic where the tumor cells take on the role of endothelial cells and line vascular channels [89]. Progressing along this line, cancer stem cells (CSC) can also trans-differentiate into endothelial cells and thereby take part in new vessel formation and tumor vascularization [69, 102, 226]. In addition to providing mechanisms for tumor escape from anti-angiogenic therapy, these mechanisms also add to the high endothelial molecular heterogeneity in tumors and the complexity of the molecular processes involved in tumor angiogenesis.
Vascular mimicry, aside from being an indicator of bad prognosis and bad survival, can also be exploited as a therapeutic target. Many attempts have been made to target the VM process specifically in malignant tumors. Specific therapy targeting CSC, using oncolytic measles viruses toward CD133, showed useful effects in mouse models, having potential both against vascular mimicry and endothelial trans-differentiation [227]. It has been discussed that the use of vincristine in combination with a specific inhibitor of the sarcoma family kinases could have a positive effect on inhibition of vascular mimicry. In addition to this, some other compounds have been suggested to have a positive effect on inhibition of VM, such as brucine (exerting its effect by modifying the structure of actin and tubulin), hinokitiol (governs EGFR proteasome degradation), and 6′-bis (2.3-dimethoxybenzoyl)- α,α-d-trehalose (DMBT) a derivative of brartemicin [89]. Brucin has been also reported to have a negative effect on expression of some key mediators of metastasis and VM formation such as MMP-2, MMP-9, and EpHhA2 in triple negative breast cancer cell line MDA-MB-231 [228]. In the study of Serwe et al. from 2012 has been reported the effect of flavone isoxanthohumol which was capable of decreasing the formation of VM in triple negative breast cancer cell line MDA-MB-231 cultured in Matrigel by blocking IL-4, IL-6, and IFN-γ-dependent Jak/Stat pathway [229]. Furthermore, in 2019 a study conducted by Kumar et al. on potential effect of tivantinib on VM in melanoma cell lines was presented [230]. It has been known that tivantinib targets MET and microtubule construction by affecting proteins such as vinculin and RhoC. In their study they suggest that tivantinib could target VM in melanoma, aside its already known effects on MET [230]. Specific therapeutic approach targeting VM has been undergoing clinical trials (NCT03582618). In this trial, a specific molecular compound named CVM-1118 has been used. This chemical is classified as a phenyl-quinoline derivate with reported anti-mutagenic and anti-neoplastic properties [231]. Another approach is to target the activity of FAK/y658 VE-cadherin with PF-271 which can consequently repress genes involved in VM promotion [232]. In addition to this, inhibition of VE-PTP/Tie-2 with AKB-9778 has been discussed as a way to control the VM forming capacity of pseudo endothelial cells [231, 233]. An approach for targeting pericytes has been proposed as well, where PDGFb axis is being targeted using STI-571 which has been proven efficient in vitro and in vivo in murine models [234,235,236].
Targeting chemokine pathways might have the potential as an anti-angiogenic treatment. It has been known that certain chemokines, such as CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7 and CXCL8, have proangiogenic effects when bound to CXCR2 receptor on TEC [237, 238]. It has been known that this effect of the CXCR2 receptor comes from the increased expression of VEGF in ovarian cancer [239]. Inhibition of this receptor has been shown to cause inhibition of angiogenesis and tumor growth in a murine model of pancreatic ductal adenocarcinoma [240]. CXCR4 is shown to be highly expressed in tumor vasculature and to promote endothelial cell sprouting [241]. IL8 or CXCL8 has been proven as a very important regulator of tumor angiogenesis, chemoresistance, and cancer stemness [242,243,244,245]. It is able to promote angiogenesis by inducing proangiogenic factors, namely VEGF, MMP-2, and MMP-9, and by governing endothelial survival [69, 246, 247]. CCL2 has been described to govern endothelial permeability and higher metastatic potential when bound to CCR2 receptor on endothelial cells [248]. Furthermore, endothelial cells expressing CCR2 can be recruited by tumor secretion of CCL2. Blocking the function of CCR2 by using its antagonist CCX872 has shown to increase the survival in Her2/neu mice models [249].
It has been demonstrated that annexin A1 and aminopeptidase—P are highly expressed in the lung cancer vasculature and therefore might be potential therapeutic targets. Furthermore, it has been demonstrated that 125I labeled anti-annexin A1 antibodies accumulate in the lung tumor tissue in rats, destroying the tumor and prolonging the survival [250].
In recent years, knowledge accumulated on molecular characteristics of tumor vasculature was exploited for designing vaccines targeting specific proangiogenic antigens. This anti-angiogenic vaccination program is suggested to be undertaken along with the immunotherapy and chemotherapy. A number of antigens has been proposed to be used for the development of vaccines. VEGF being an important factor in vasculogenesis has tried to been exploited first in 2001 as reported by Wei et al. [251]. In their study they used a Xenopus levis VEGF DNA vaccine that showed promising results in several murine models. The first clinical trials came in 2014 as reported by Gavilondo et al. [252] where they reported a phase I clinical study (CENTAURO) on 30 patients with advanced solid tumors, using CIGB-247 vaccine which was consisting of human VEGF variant molecule together with bacterial adjuvant. This study revealed some very promising data. Namely, after 8 weeks after subcutaneous immunization and after revaccination on week 12, this vaccine showed immunogenicity in three sequential analyses of the patients’ serum and neither physiological parameters have been altered by vaccination nor it has triggered the autoimmune response, giving high promises as a novel therapeutic strategy. Along with VEGF, VEGFR2 (FLK-1) has been as well exploited as a potential vaccine antigen giving promising results as reported in a number of studies [253,254,255,256,257,258,259,260]. There has been a number of attempts to exploit bFGF (FGF-2) and FGFR (CD331) in anti-angiogenic vaccine development. Recombinant protein vaccines, xenogeneic plasmid DNA vaccines, peptide vaccines with adjuvants, and liposome-based peptide vaccines have been tested in preclinical murine models [261,262,263,264]. In the study from 2004 by Plum et al. using liposome based peptide vaccine against (FGF-2), anti-angiogenic response has been reported without any adverse physiological effects [265]. In a study from 2005, McNeel et al. reported a successfully completed phase I clinical trial of monoclonal anti- αvβ3 antibody for the treatment of advanced solid tumors named MEDI-522 [266]. Endoglin, being highly expressed on the surface of TEC, has been used as a potential vaccine as an antigen on bacterial surface or oral DNA vaccine [267, 268]. There was a number of monoclonal antibodies and vaccines developed to target EGFR. Many of these are already in clinical use including monoclonal antibodies cetuximab, panitumumab, and nimotuzumab, and the small tyrosine kinase molecules erlotinib and gefitinib. A vaccine based on EGF bounded to a carrier protein from Neisseria meningitidis, CIMAvax® has been proven beneficial in patients with non-small-cell lung cancer and there were no adverse effects noted in postoperative wound healing [269, 270].
Concluding Remarks/Summary
With this chapter, we tried to bring closer to the reader a complex topic of molecular diversity of tumor endothelium, and a possibility to exploit specific molecular markers and signatures as prognostic and predictive indicators and therapeutic targets. It has been known that one of the hallmarks of cancer is angiogenesis. Tumors, having high metabolic demands, use angiogenesis to meet their increasing needs for oxygen and nutrients. Tumor angiogenesis leads to aberrant vessel formation and aberrant endothelial phenotypes and functions. Here, various studies have been discussed, undertaken in order to reveal specific tumor endothelial signatures, with a possibility for prognostic and predictive use. Because there is no perfect single marker that would be suitable for providing robust information on prognosis and prediction, multiparametric single cell analysis might be necessary, and some single cell studies have been discussed in this chapter. Focus has also been given to experimental and clinical studies, which all clearly point to the fact that tumor related angiogenesis is different from physiological vasculogenesis and that through all of its challenging complexity there is a possibility to exploit its diagnostic and therapeutic value. In the era of precision therapies, there is an increased importance for specific companion biomarkers in order to sort out the patients into subgroups with an aim to define responders, better select the therapeutic strategy, and increase the efficiency of treatment. Biomarkers reflecting the anti-angiogenic therapy response are of high importance. Based on specific molecular pathways, many novel therapeutic approaches have been suggested. In addition, novel vaccines have been designed targeting specific TEM proteins and have been shown to be successful in reducing tumor angiogenesis and preventing the occurrence of metastasis, while being successful in avoiding autoimmune response against normal vessels. With the development of more powerful single cell and imaging methods, we can expect to map the tumor microenvironment at a higher resolution, possibly yielding better biomarkers and better therapeutic options in the near future.
References
Aird WC. Phenotypic heterogeneity of the endothelium: I. Structure, function, and mechanisms. Circ Res. 2007;100(2):158–73. https://doi.org/10.1161/01.RES.0000255691.76142.4a.
Krüger-Genge A, et al. Vascular endothelial cell biology: an update. Int J Mol Sci. 2019;20(18):4411. https://doi.org/10.3390/ijms20184411.
Aird WC. Molecular heterogeneity of tumor endothelium. Cell Tissue Res. 2009;335:271–81. https://doi.org/10.1007/s00441-008-0672-y.
Aird WC. Endothelial cell heterogeneity. Crit Care Med. 2003;31(4 Suppl):S221–30. https://doi.org/10.1097/01.CCM.0000057847.32590.C1.
Jambusaria A, et al. Endothelial heterogeneity across distinct vascular beds during homeostasis and inflammation. eLife. 2020;9:e51413. https://doi.org/10.7554/eLife.51413.
Potente M, Mäkinen T. Vascular heterogeneity and specialization in development and disease. Nat Rev Mol Cell Biol. 2017;18(8):477–94. https://doi.org/10.1038/nrm.2017.36.
Gerritsen ME, Printz MP. Sites of prostaglandin synthesis in the bovine heart and isolated bovine coronary microvessels. Circ Res. 1981;49(5):1152–63. https://doi.org/10.1161/01.res.49.5.1152.
Johnson AR. Human pulmonary endothelial cells in culture. Activities of cells from arteries and cells from veins. J Clin Invest. 1980;65(4):841–50. https://doi.org/10.1172/JCI109736.
Kalucka J, et al. Single-cell transcriptome atlas of murine endothelial cells. Cell. 2020;180(4):764–779.e20. https://doi.org/10.1016/j.cell.2020.01.015.
Chi JT, et al. Endothelial cell diversity revealed by global expression profiling. Proc Natl Acad Sci U S A. 2003;100(19):10623–8. https://doi.org/10.1073/pnas.1434429100.
Paik DT, et al. Single-cell RNA sequencing unveils unique transcriptomic signatures of organ-specific endothelial cells. Circulation. 2020;142(19):1848–62. https://doi.org/10.1161/CIRCULATIONAHA.119.041433.
Augustin HG, Koh GY. Organotypic vasculature: from descriptive heterogeneity to functional pathophysiology. Science (New York, NY). 2017;357(6353):eaal2379. https://doi.org/10.1126/science.aal2379.
Bussmann J, Wolfe SA, Siekmann AF. Arterial-venous network formation during brain vascularization involves hemodynamic regulation of chemokine signaling. Development. 2011;138(9):1717–26. https://doi.org/10.1242/dev.059881.
Matsuoka H, et al. The retinoic acid receptor-related orphan receptor α positively regulates tight junction protein claudin domain-containing 1 mRNA expression in human brain endothelial cells. J Biochem. 2017;161(5):441–50. https://doi.org/10.1093/jb/mvw092.
Gillich A, et al. Capillary cell-type specialization in the alveolus. Nature. 2020;586(7831):785–9. https://doi.org/10.1038/s41586-020-2822-7.
Deanfield JE, Halcox JP, Rabelink TJ. Endothelial function and dysfunction: testing and clinical relevance. Circulation. 2007;115(10):1285–95. https://doi.org/10.1161/CIRCULATIONAHA.106.652859.
Pober JS, Sessa WC. Inflammation and the blood microvascular system. Cold Spring Harb Perspect Biol. 2014;7(1):a016345. https://doi.org/10.1101/cshperspect.a016345.
Michiels C. Endothelial cell functions. J Cell Physiol. 2003;196(3):430–43. https://doi.org/10.1002/jcp.10333.
Gori T. Endothelial function: a short guide for the interventional cardiologist. Int J Mol Sci. 2018;19(12):3838. https://doi.org/10.3390/ijms19123838.
Miura H, Gutterman DD. Human coronary arteriolar dilation to arachidonic acid depends on cytochrome P-450 monooxygenase and Ca2+-activated K+ channels. Circ Res. 1998;83(5):501–7. https://doi.org/10.1161/01.res.83.5.501.
Palmer RM, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–6. https://doi.org/10.1038/327524a0.
Loh YC, et al. Overview of the microenvironment of vasculature in vascular tone regulation. Int J Mol Sci. 2018;19(1):120. https://doi.org/10.3390/ijms19010120.
Moncada S, Higgs EA, Vane JR. Human arterial and venous tissues generate prostacyclin (prostaglandin x), a potent inhibitor of platelet aggregation. Lancet. 1977;1(8001):18–20. https://doi.org/10.1016/s0140-6736(77)91655-5.
Chien S. Effects of disturbed flow on endothelial cells. Ann Biomed Eng. 2008;36(4):554–62. https://doi.org/10.1007/s10439-007-9426-3.
Garland CJ, Dora KA. EDH: endothelium-dependent hyperpolarization and microvascular signalling. Acta Physiol (Oxf). 2017;219(1):152–61. https://doi.org/10.1111/apha.12649.
Yanagisawa M, et al. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature. 1988;332(6163):411–5. https://doi.org/10.1038/332411a0.
Cines DB, et al. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood. 1998;91(10):3527–61.
de Graaf JC, et al. Nitric oxide functions as an inhibitor of platelet adhesion under flow conditions. Circulation. 1992;85(6):2284–90. https://doi.org/10.1161/01.cir.85.6.2284.
Sadler JE. Thrombomodulin structure and function. Thromb Haemost. 1997;78(1):392–5.
Denis CV, et al. Defect in regulated secretion of P-selectin affects leukocyte recruitment in von Willebrand factor-deficient mice. Proc Natl Acad Sci U S A. 2001;98(7):4072–7. https://doi.org/10.1073/pnas.061307098.
Eppihimer MJ, et al. Heterogeneity of expression of E- and P-selectins in vivo. Circ Res. 1996;79(3):560–9. https://doi.org/10.1161/01.res.79.3.560.
Salsman VS, et al. Crosstalk between medulloblastoma cells and endothelium triggers a strong chemotactic signal recruiting T lymphocytes to the tumor microenvironment. PLoS One. 2011;6(5):e20267. https://doi.org/10.1371/journal.pone.0020267.
Young MR. Endothelial cells in the eyes of an immunologist. Cancer Immunol Immunother. 2012;61(10):1609–16. https://doi.org/10.1007/s00262-012-1335-0.
Johnson LA, Jackson DG. Inflammation-induced secretion of CCL21 in lymphatic endothelium is a key regulator of integrin-mediated dendritic cell transmigration. Int Immunol. 2010;22(10):839–49. https://doi.org/10.1093/intimm/dxq435.
Pober JS, Sessa WC. Evolving functions of endothelial cells in inflammation. Nat Rev Immunol. 2007;7(10):803–15. https://doi.org/10.1038/nri2171.
Teijaro JR, et al. Endothelial cells are central orchestrators of cytokine amplification during influenza virus infection. Cell. 2011;146(6):980–91. https://doi.org/10.1016/j.cell.2011.08.015.
Zhang J, et al. Endothelial lactate controls muscle regeneration from ischemia by inducing M2-like macrophage polarization. Cell Metab. 2020;31(6):1136–1153.e7. https://doi.org/10.1016/j.cmet.2020.05.004.
Maishi N, Hida K. Tumor endothelial cells accelerate tumor metastasis. Cancer Sci. 2017;108(10):1921–6. https://doi.org/10.1111/cas.13336.
Geindreau M, Ghiringhelli F, Bruchard M. Vascular endothelial growth factor, a key modulator of the anti-tumor immune response. Int J Mol Sci. 2021;22(9):4871. https://doi.org/10.3390/ijms22094871.
Brat DJ, Van Meir EG. Glomeruloid microvascular proliferation orchestrated by VPF/VEGF: a new world of angiogenesis research. Am J Pathol. 2001;158(3):789–96. https://doi.org/10.1016/S0002-9440(10)64025-4.
Nagy JA, et al. Heterogeneity of the tumor vasculature. Semin Thromb Hemost. 2010;36(3):321–31. https://doi.org/10.1055/s-0030-1253454.
Pettersson A, et al. Heterogeneity of the angiogenic response induced in different normal adult tissues by vascular permeability factor/vascular endothelial growth factor. Lab Invest. 2000;80(1):99–115. https://doi.org/10.1038/labinvest.3780013.
Baluk P, Hashizume H, McDonald DM. Cellular abnormalities of blood vessels as targets in cancer. Curr Opin Genet Dev. 2005;15(1):102–11. https://doi.org/10.1016/j.gde.2004.12.005.
Bennewith KL, Durand RE. Quantifying transient hypoxia in human tumor xenografts by flow cytometry. Cancer Res. 2004;64(17):6183–9. https://doi.org/10.1158/0008-5472.CAN-04-0289.
Kimura H, et al. Fluctuations in red cell flux in tumor microvessels can lead to transient hypoxia and reoxygenation in tumor parenchyma. Cancer Res. 1996;56(23):5522–8.
McDonald DM, Baluk P. Imaging of angiogenesis in inflamed airways and tumors: newly formed blood vessels are not alike and may be wildly abnormal: Parker B. Francis lecture. Chest. 2005;128(6 Suppl):602S–8S. https://doi.org/10.1378/chest.128.6_suppl.602S-a.
Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis. Nature. 2011;473(7347):298–307. https://doi.org/10.1038/nature10144.
Chang YS, et al. Mosaic blood vessels in tumors: frequency of cancer cells in contact with flowing blood. Proc Natl Acad Sci U S A. 2000;97(26):14608–13. https://doi.org/10.1073/pnas.97.26.14608.
Cao Z, et al. Angiocrine factors deployed by tumor vascular niche induce B cell lymphoma invasiveness and chemoresistance. Cancer Cell. 2014;25(3):350–65. https://doi.org/10.1016/j.ccr.2014.02.005.
Cao Z, et al. Molecular checkpoint decisions made by subverted vascular niche transform indolent tumor cells into chemoresistant cancer stem cells. Cancer Cell. 2017;31(1):110–26. https://doi.org/10.1016/j.ccell.2016.11.010.
Ghiabi P, et al. Endothelial cells provide a notch-dependent pro-tumoral niche for enhancing breast cancer survival, stemness and pro-metastatic properties. PLoS One. 2014;9(11):e112424. https://doi.org/10.1371/journal.pone.0112424.
Lu J, et al. Endothelial cells promote the colorectal cancer stem cell phenotype through a soluble form of Jagged-1. Cancer Cell. 2013;23(2):171–85. https://doi.org/10.1016/j.ccr.2012.12.021.
Pedrosa AR, et al. Endothelial Jagged1 promotes solid tumor growth through both pro-angiogenic and angiocrine functions. Oncotarget. 2015;6(27):24404–23. https://doi.org/10.18632/oncotarget.4380.
Wieland E, et al. Endothelial Notch1 Activity facilitates metastasis. Cancer Cell. 2017;31(3):355–67. https://doi.org/10.1016/j.ccell.2017.01.007.
Zhu TS, et al. Endothelial cells create a stem cell niche in glioblastoma by providing NOTCH ligands that nurture self-renewal of cancer stem-like cells. Cancer Res. 2011;71(18):6061–72. https://doi.org/10.1158/0008-5472.CAN-10-4269.
Yadav A, et al. Tumor-associated endothelial cells promote tumor metastasis by chaperoning circulating tumor cells and protecting them from anoikis. PLoS One. 2015;10(10):e0141602. https://doi.org/10.1371/journal.pone.0141602.
Beerepoot LV, et al. Increased levels of viable circulating endothelial cells are an indicator of progressive disease in cancer patients. Ann Oncol. 2004;15(1):139–45. https://doi.org/10.1093/annonc/mdh017.
Mancuso P, et al. Resting and activated endothelial cells are increased in the peripheral blood of cancer patients. Blood. 2001;97(11):3658–61. https://doi.org/10.1182/blood.v97.11.3658.
Maishi N, et al. Tumour endothelial cells in high metastatic tumours promote metastasis via epigenetic dysregulation of biglycan. Sci Rep. 2016;6:28039. https://doi.org/10.1038/srep28039.
Ohga N, et al. Heterogeneity of tumor endothelial cells: comparison between tumor endothelial cells isolated from high- and low-metastatic tumors. Am J Pathol. 2012;180(3):1294–307. https://doi.org/10.1016/j.ajpath.2011.11.035.
Branco-Price C, et al. Endothelial cell HIF-1α and HIF-2α differentially regulate metastatic success. Cancer Cell. 2012;21(1):52–65. https://doi.org/10.1016/j.ccr.2011.11.017.
Buckanovich RJ, et al. Tumor vascular proteins as biomarkers in ovarian cancer. J Clin Oncol. 2007;25(7):852–61. https://doi.org/10.1200/JCO.2006.08.8583.
St. Croix B, et al. Genes expressed in human tumor endothelium. Science. 2000;289(5482):1197–202. https://doi.org/10.1126/science.289.5482.1197.
Dieterich LC, et al. Transcriptional profiling of human glioblastoma vessels indicates a key role of VEGF-A and TGFβ2 in vascular abnormalization. J Pathol. 2012;228(3):378–90. https://doi.org/10.1002/path.4072.
Roudnicky F, et al. Endocan is upregulated on tumor vessels in invasive bladder cancer where it mediates VEGF-A-induced angiogenesis. Cancer Res. 2013;73(3):1097–106. https://doi.org/10.1158/0008-5472.CAN-12-1855.
Zhang L, et al. Tumor-derived vascular endothelial growth factor up-regulates angiopoietin-2 in host endothelium and destabilizes host vasculature, supporting angiogenesis in ovarian cancer. Cancer Res. 2003;63(12):3403–12.
Zhao Q, et al. Single-cell transcriptome analyses reveal endothelial cell heterogeneity in tumors and changes following antiangiogenic treatment. Cancer Res. 2018;78(9):2370–82. https://doi.org/10.1158/0008-5472.CAN-17-2728.
Goveia J, et al. An integrated gene expression landscape profiling approach to identify lung tumor endothelial cell heterogeneity and angiogenic candidates. Cancer Cell. 2020;37(1):21–36.e13. https://doi.org/10.1016/j.ccell.2019.12.001.
Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci. 2020;77(9):1745–70. https://doi.org/10.1007/s00018-019-03351-7.
Hwa C, Sebastian A, Aird WC. Endothelial biomedicine: its status as an interdisciplinary field, its progress as a basic science, and its translational bench-to-bedside gap. Endothelium. 2005;12(3):139–51. https://doi.org/10.1080/10623320500192016.
Kahn BM, et al. The vascular landscape of human cancer. J Clin Invest. 2021;131(2):e136655. https://doi.org/10.1172/JCI136655.
Mezheyeuski A, et al. Survival-associated heterogeneity of marker-defined perivascular cells in colorectal cancer. Oncotarget. 2016;7(27):41948–58. https://doi.org/10.18632/oncotarget.9632.
Asahara T, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275(5302):964–7. https://doi.org/10.1126/science.275.5302.964.
Yin AH, et al. AC133, a novel marker for human hematopoietic stem and progenitor cells. Blood. 1997;90(12):5002–12. https://doi.org/10.1182/blood.v90.12.5002.
Peichev M, et al. Expression of VEGFR-2 and AC133 by circulating human CD34+ cells identifies a population of functional endothelial precursors. Blood. 2000;95(3):952–8. https://doi.org/10.1182/blood.v95.3.952.003k27_952_958.
Romagnani P, et al. CD14+CD34low cells with stem cell phenotypic and functional features are the major source of circulating endothelial progenitors. Circ Res. 2005;97(4):314–22. https://doi.org/10.1161/01.RES.0000177670.72216.9b.
Asahara T, et al. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ Res. 1999a;85(3):221–8. https://doi.org/10.1161/01.RES.85.3.221.
Asahara T, et al. VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 1999b;18(14):3964–72. https://doi.org/10.1093/emboj/18.14.3964.
Kalka C, et al. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc Natl Acad Sci. 2000;97(7):3422–7. https://doi.org/10.1073/pnas.97.7.3422.
Lin Y, et al. Origins of circulating endothelial cells and endothelial outgrowth from blood. J Clin Investig. 2000;105(1):71–7. https://doi.org/10.1172/JCI8071.
Shi Q, et al. Evidence for circulating bone marrow-derived endothelial cells. Blood. 1998;92(2):362–7. https://doi.org/10.1182/blood.v92.2.362.
Shin JW, et al. Isolation of endothelial progenitor cells from cord blood and induction of differentiation by Ex Vivo expansion. Yonsei Med J. 2005;46(2):260–7. https://doi.org/10.3349/ymj.2005.46.2.260.
Blann AD, et al. Circulating endothelial cells. Biomarker of vascular disease. Thromb Haemost. 2005;93(2):228–35. https://doi.org/10.1160/TH04-09-0578.
Bertolini F, et al. The multifaceted circulating endothelial cell in cancer: towards marker and target identification. Nat Rev Cancer. 2006;6(11):835–45. https://doi.org/10.1038/nrc1971.
Ronzoni M, et al. Circulating endothelial cells and endothelial progenitors as predictive markers of clinical response to bevacizumab-based first-line treatment in advanced colorectal cancer patients. Ann Oncol. 2010;21(12):2382–9. https://doi.org/10.1093/annonc/mdq261.
Mehran R, et al. Tumor endothelial markers define novel subsets of cancer-specific circulating endothelial cells associated with antitumor efficacy. Cancer Res. 2014;74(10):2731–41. https://doi.org/10.1158/0008-5472.CAN-13-2044.
Cima I, et al. Tumor-derived circulating endothelial cell clusters in colorectal cancer. Sci Transl Med. 2016;8(345):345ra89. https://doi.org/10.1126/scitranslmed.aad7369.
Rahbari NN, et al. Prognostic value of circulating endothelial cells in metastatic colorectal cancer. Oncotarget. 2017;8(23):37491–501. https://doi.org/10.18632/oncotarget.16397.
Andonegui-Elguera MA, et al. An overview of vasculogenic mimicry in breast cancer. Front Oncol. 2020;10(February):1–8. https://doi.org/10.3389/fonc.2020.00220.
Lizárraga-Verdugo E, et al. Cancer stem cells and its role in angiogenesis and vasculogenic mimicry in gastrointestinal cancers. Front Oncol. 2020;10(March):1–8. https://doi.org/10.3389/fonc.2020.00413.
Luo Q, et al. Vasculogenic mimicry in carcinogenesis and clinical applications. J Hematol Oncol. 2020;13(1):1–15. https://doi.org/10.1186/s13045-020-00858-6.
Maniotis AJ, et al. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol. 1999;155(3):739–52. https://doi.org/10.1016/S0002-9440(10)65173-5.
Álvarez-Viejo M, et al. CD271 as a marker to identify mesenchymal stem cells from diverse sources before culture. World J Stem Cells. 2015;7(2):470–6. https://doi.org/10.4252/wjsc.v7.i2.470.
Delgado-Bellido D, et al. VE-cadherin promotes vasculogenic mimicry by modulating kaiso-dependent gene expression. Cell Death Differ. 2019;26(2):348–61. https://doi.org/10.1038/s41418-018-0125-4.
Cao Z, et al. Tumour vasculogenic mimicry is associated with poor prognosis of human cancer patients: a systemic review and meta-analysis. Eur J Cancer. 2013b;49(18):3914–23. https://doi.org/10.1016/j.ejca.2013.07.148.
Yang JP, et al. Tumor vasculogenic mimicry predicts poor prognosis in cancer patients: a meta-analysis. Angiogenesis. 2016;19(2):191–200. https://doi.org/10.1007/s10456-016-9500-2.
Shen Y, et al. Tumor vasculogenic mimicry formation as an unfavorable prognostic indicator in patients with breast cancer. Oncotarget. 2017;8(34):56408–16. https://doi.org/10.18632/oncotarget.16919.
Zhang Z, et al. The role of vascular mimicry as a biomarker in malignant melanoma: a systematic review and meta-analysis. BMC Cancer. 2019;19(1):1–12. https://doi.org/10.1186/s12885-019-6350-5.
Alvero AB, et al. Stem-like ovarian cancer cells can serve as tumor vascular progenitors. Stem Cells. 2009;27(10):2405–13. https://doi.org/10.5949/liverpool/9780853236788.003.0003.
Bussolati B, et al. Endothelial cell differentiation of human breast tumour stem/progenitor cells. J Cell Mol Med. 2009;13(2):309–19. https://doi.org/10.1111/j.1582-4934.2008.00338.x.
Li F, Xu J, Liu S. Cancer stem cells and neovascularization. Cells. 2021;10(5):1070. https://doi.org/10.3390/cells10051070.
Mei X, et al. Glioblastoma stem cell differentiation into endothelial cells evidenced through live-cell imaging. Neuro-Oncology. 2017;19(8):1109–18. https://doi.org/10.1093/neuonc/nox016.
Wang R, et al. Glioblastoma stem-like cells give rise to tumour endothelium. Nature. 2010;468(7325):829–33. https://doi.org/10.1038/nature09624.
Ricci-Vitiani L, et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature. 2010;468(7325):824–30. https://doi.org/10.1038/nature09557.
Zhao Y, et al. Endothelial cell transdifferentiation of human glioma stem progenitor cells in vitro. Brain Res Bull. 2010;82(5–6):308–12. https://doi.org/10.1016/j.brainresbull.2010.06.006.
Bussolati B, et al. Identification of a tumor-initiating stem cell population in human renal carcinomas. FASEB J. 2008;22(10):3696–705. https://doi.org/10.1096/fj.08-102590.
Rau K-M, et al. Neovascularization evaluated by CD105 correlates well with prognostic factors in breast cancers. Exp Ther Med. 2012;4(2):231–6. https://doi.org/10.3892/etm.2012.594.
Saroufim A, et al. Tumoral CD105 is a novel independent prognostic marker for prognosis in clear-cell renal cell carcinoma. Br J Cancer. 2014;110(7):1778–84. https://doi.org/10.1038/bjc.2014.71.
Aomatsu N, et al. CD133 is a useful surrogate marker for predicting chemosensitivity to neoadjuvant chemotherapy in breast cancer. PLoS One. 2012;7(9):e45865. https://doi.org/10.1371/journal.pone.0045865.
Kim SJ, et al. Prognostic impact and clinicopathological correlation of CD133 and ALDH1 expression in invasive breast cancer. J Breast Cancer. 2015;18(4):347–55. https://doi.org/10.4048/jbc.2015.18.4.347.
Rettig WJ, et al. Identification of endosialin, a cell surface glycoprotein of vascular endothelial cells in human cancer. Proc Natl Acad Sci U S A. 1992;89(22):10832–6. https://doi.org/10.1073/pnas.89.22.10832.
Chang SS, et al. Prostate-specific membrane antigen is produced in tumor-associated neovasculature. Clinical Cancer Res. 1999;5(10):2674–81.
Abid MR, et al. Vascular endocan is preferentially expressed in tumor endothelium. Microvasc Res. 2006;72(3):136–45. https://doi.org/10.1016/j.mvr.2006.05.010.
Burrows FJ, et al. Up-regulation of endoglin on vascular endothelial cells in human solid tumors: implications for diagnosis and therapy. Clin Cancer Res. 1995;1(12):1623–34.
Gasparini G, et al. Vascular integrin alpha(v)beta3: a new prognostic indicator in breast cancer. Clin Res. 1998;4(11):2625–34.
Kim S, et al. Regulation of angiogenesis in vivo by ligation of integrin alpha5beta1 with the central cell-binding domain of fibronectin. Am J Pathol. 2000;156(4):1345–62. https://doi.org/10.1016/s0002-9440(10)65005-5.
Griffioen AW, et al. Endothelial intercellular adhesion molecule-1 expression is suppressed in human malignancies: the role of angiogenic factors. Cancer Res. 1996;56(5):1111–7.
Hellwig SMM, et al. Endothelial CD34 is suppressed in human malignancies: role of angiogenic factors. Cancer Lett. 1997;120(2):203–11. https://doi.org/10.1016/S0304-3835(97)00310-8.
Davies G, et al. Levels of expression of endothelial markers specific to tumour-associated endothelial cells and their correlation with prognosis in patients with breast cancer. Clin Exp Metastasis. 2004;21(1):31–7. https://doi.org/10.1023/b:clin.0000017168.83616.d0.
Rmali KA, et al. Tumour endothelial marker 8 (TEM-8) in human colon cancer and its association with tumour progression. Eur J Surg Oncol. 2004;30(9):948–53. https://doi.org/10.1016/j.ejso.2004.07.023.
Rmali KA, Puntis MCA, Jiang WG. Prognostic values of tumor endothelial markers in patients with colorectal cancer. World J Gastroenterol. 2005;11(9):1283–6. https://doi.org/10.3748/wjg.v11.i9.1283.
Fuchs B, et al. High expression of tumor endothelial marker 7 is associated with metastasis and poor survival of patients with osteogenic sarcoma. Gene. 2007;399(2):137–43. https://doi.org/10.1016/j.gene.2007.05.003.
MacFadyen J, et al. Endosialin is expressed on stromal fibroblasts and CNS pericytes in mouse embryos and is downregulated during development. Gene Expr Patterns. 2007;7(3):363–9. https://doi.org/10.1016/j.modgep.2006.07.006.
Opavsky R, et al. Molecular characterization of the mouse Tem1/endosialin gene regulated by cell density in vitro and expressed in normal tissues in vivo. J Biol Chem. 2001;276(42):38795–807. https://doi.org/10.1074/jbc.M105241200.
Halder C, et al. Preferential expression of the secreted and membrane forms of tumor endothelial marker 7 transcripts in osteosarcoma. Anticancer Res. 2009;29(11):4317–22.
Lee HK, et al. Cloning, characterization and neuronal expression profiles of tumor endothelial marker 7 in the rat brain. Brain Res Mol Brain Res. 2005a;136(1–2):189–98. https://doi.org/10.1016/j.molbrainres.2005.02.010.
Zhang ZZ, et al. TEM7 (PLXDC1), a key prognostic predictor for resectable gastric cancer, promotes cancer cell migration and invasion. Am J Cancer Res. 2015;5(2):772–81.
Czekierdowski A, et al. Prognostic significance of TEM7 and nestin expression in women with advanced high grade serous ovarian cancer. Ginekol Pol. 2018;89(3):135–41. https://doi.org/10.5603/GP.a2018.0023.
Gutwein LG, et al. Tumor endothelial marker 8 expression in triple-negative breast cancer. Anticancer Res. 2011;31(10):3417–22. Available at: https://ar.iiarjournals.org/content/31/10/3417
Kuo F, et al. Immuno-PET imaging of tumor endothelial Marker 8 (TEM8). Mol Pharm. 2014;11(11):3996–4006. https://doi.org/10.1021/mp500056d.
Pietrzyk Ł, et al. Clinical value of detecting tumor endothelial marker 8 (Antxr1) as a biomarker in the diagnosis and prognosis of colorectal cancer. Cancer Manag Res. 2021;13:3113–22. https://doi.org/10.2147/CMAR.S298165.
Basilio-de-Oliveira RP, Pannain VLN. Prognostic angiogenic markers (endoglin, VEGF, CD31) and tumor cell proliferation (Ki67) for gastrointestinal stromal tumors. World J Gastroenterol. 2015;21(22):6924–30. https://doi.org/10.3748/wjg.v21.i22.6924.
Salvesen HB, et al. Significance of CD 105 expression for tumour angiogenesis and prognosis in endometrial carcinomas. APMIS. 2003;111(11):1011–8. https://doi.org/10.1111/j.1600-0463.2003.apm1111103.x.
Straume O, Akslen LA. Expression of vascular endothelial growth factor, its receptors (FLT-1, KDR) and TSP-1 related to microvessel density and patient outcome in vertical growth phase melanomas. Am J Pathol. 2001;159(1):223–35. https://doi.org/10.1016/S0002-9440(10)61688-4.
Svatek RS, et al. Preoperative plasma endoglin levels predict biochemical progression after radical prostatectomy. Clin Cancer Res. 2008;14(11):3362–6. https://doi.org/10.1158/1078-0432.CCR-07-4707.
Östman A, Corvigno S. Microvascular mural cells in cancer. Trends Cancer. 2018;4(12):838–48. https://doi.org/10.1016/j.trecan.2018.10.004.
Sinha D, et al. Pericytes promote malignant ovarian cancer progression in mice and predict poor prognosis in serous ovarian cancer patients. Clin Cancer Res. 2016;22(7):1813–24. https://doi.org/10.1158/1078-0432.CCR-15-1931.
Viski C, et al. Endosialin-expressing pericytes promote metastatic dissemination. Cancer Res. 2016;76(18):5313–25. https://doi.org/10.1158/0008-5472.CAN-16-0932.
Cantelmo AR, et al. Inhibition of the glycolytic activator PFKFB3 in endothelium induces tumor vessel normalization, impairs metastasis, and improves chemotherapy. Cancer Cell. 2016;30(6):968–85. https://doi.org/10.1016/j.ccell.2016.10.006.
Cooke VG, et al. Pericyte depletion results in hypoxia-associated epithelial-to-mesenchymal transition and metastasis mediated by met signaling pathway. Cancer cell. 2012;21(1):66–81. https://doi.org/10.1016/j.ccr.2011.11.024.
Hamzah J, et al. Vascular normalization in Rgs5-deficient tumours promotes immune destruction. Nature. 2008;453(7193):410–4. https://doi.org/10.1038/nature06868.
Hong J, et al. Role of tumor pericytes in the recruitment of myeloid-derived suppressor cells. J Natl Cancer Inst. 2015;107(10):djv209. https://doi.org/10.1093/jnci/djv209.
Lyle LT, et al. Alterations in pericyte subpopulations are associated with elevated blood-tumor barrier permeability in experimental brain metastasis of breast cancer. Clin Cancer Res. 2016;22(21):5287–99. https://doi.org/10.1158/1078-0432.CCR-15-1836.
Nisancioglu MH, Betsholtz C, Genové G. The absence of pericytes does not increase the sensitivity of tumor vasculature to vascular endothelial growth factor-A blockade. Cancer Res. 2010;70(12):5109–15. https://doi.org/10.1158/0008-5472.CAN-09-4245.
Xian X, et al. Pericytes limit tumor cell metastasis. J Clin Invest. 2006;116(3):642–51. https://doi.org/10.1172/JCI25705.
Cao Y, et al. Pericyte coverage of differentiated vessels inside tumor vasculature is an independent unfavorable prognostic factor for patients with clear cell renal cell carcinoma. Cancer. 2013a;119(2):313–24. https://doi.org/10.1002/cncr.27746.
Corvigno S, et al. Markers of fibroblast-rich tumor stroma and perivascular cells in serous ovarian cancer: inter- and intra-patient heterogeneity and impact on survival. Oncotarget. 2016;7(14):18573–84. https://doi.org/10.18632/oncotarget.7613.
Frödin M, et al. Perivascular PDGFR-β is an independent marker for prognosis in renal cell carcinoma. Br J Cancer. 2017;116(2):195–201. https://doi.org/10.1038/bjc.2016.407.
Tolaney SM, et al. Role of vascular density and normalization in response to neoadjuvant bevacizumab and chemotherapy in breast cancer patients. Proc Natl Acad Sci U S A. 2015;112(46):14325–30. https://doi.org/10.1073/pnas.1518808112.
Zhou W, et al. Targeting glioma stem cell-derived pericytes disrupts the blood-tumor barrier and improves chemotherapeutic efficacy. Cell Stem Cell. 2017;21(5):591–603.e4. https://doi.org/10.1016/j.stem.2017.10.002.
Bhati R, et al. Molecular characterization of human breast tumor vascular cells. Am J Pathol. 2008;172(5):1381–90. https://doi.org/10.2353/ajpath.2008.070988.
Madden SL, et al. Vascular gene expression in nonneoplastic and malignant brain. Am J Pathol. 2004;165(2):601–8. https://doi.org/10.1016/s0002-9440(10)63324-x.
Parker BS, et al. Alterations in vascular gene expression in invasive breast carcinoma. Cancer Res. 2004;64(21):7857–66. https://doi.org/10.1158/0008-5472.CAN-04-1976.
Pen A, et al. Molecular markers of extracellular matrix remodeling in glioblastoma vessels: microarray study of laser-captured glioblastoma vessels. Glia. 2007;55(6):559–72. https://doi.org/10.1002/glia.20481.
Dudley AC. Tumor endothelial cells. Cold Spring Harb Perspect Med. 2012;2(3):1–18. https://doi.org/10.1101/cshperspect.a006536.
Carson-Walter EB, et al. Plasmalemmal vesicle associated protein-1 is a novel marker implicated in brain tumor angiogenesis. Clinical Cancer Res. 2005;11(21):7643–50. https://doi.org/10.1158/1078-0432.CCR-05-1099.
van Beijnum JR, et al. Gene expression of tumor angiogenesis dissected: specific targeting of colon cancer angiogenic vasculature. Blood. 2006;108(7):2339–48. https://doi.org/10.1182/blood-2006-02-004291.
Hoda MA, et al. Temsirolimus inhibits malignant pleural mesothelioma growth in vitro and in vivo: synergism with chemotherapy. J Thorac Oncol. 2011;6(5):852–63. https://doi.org/10.1097/JTO.0b013e31820e1a25.
Lu C, et al. Gene alterations identified by expression profiling in tumor-associated endothelial cells from invasive ovarian carcinoma. Cancer Res. 2007;67(4):1757–68. https://doi.org/10.1158/0008-5472.CAN-06-3700.
Coppiello G, et al. Meox2/Tcf15 heterodimers program the heart capillary endothelium for cardiac fatty acid uptake. Circulation. 2015;131(9):815–26. https://doi.org/10.1161/CIRCULATIONAHA.114.013721.
Marcu R, et al. Human organ-specific endothelial cell heterogeneity. iScience. 2018;4:20–35. https://doi.org/10.1016/j.isci.2018.05.003.
Nolan DJ, et al. Molecular signatures of tissue-specific microvascular endothelial cell heterogeneity in organ maintenance and regeneration. Dev Cell. 2013;26(2):204–19. https://doi.org/10.1016/j.devcel.2013.06.017.Molecular.
Sabbagh MF, et al. Transcriptional and epigenomic landscapes of CNS and non-CNS vascular endothelial cells. eLife. 2018;7:e36187. https://doi.org/10.7554/eLife.36187.
Tewari KS, et al. Final overall survival of a randomized trial of bevacizumab for primary treatment of ovarian cancer. J Clin Oncol. 2019;37(26):2317–28. https://doi.org/10.1200/JCO.19.01009.
Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285(21):1182–6. https://doi.org/10.1056/NEJM197111182852108.
Cao Y, et al. Forty-year journey of angiogenesis translational research. Sci Transl Med. 2011;3(114):114rv3. https://doi.org/10.1126/scitranslmed.3003149.
Folkman J, et al. Isolation of a tumor factor responsible for angiogenesis. J Exp Med. 1971;133(2):275–88. https://doi.org/10.1084/jem.133.2.275.
Viloria-Petit A, et al. Contrasting effects of VEGF gene disruption in embryonic stem cell-derived versus oncogene-induced tumors. EMBO J. 2003;22(16):4091–102. https://doi.org/10.1093/emboj/cdg408.
Apte RS, Chen DS, Ferrara N. VEGF in signaling and disease: beyond discovery and development. Cell. 2019;176(6):1248–64. https://doi.org/10.1016/j.cell.2019.01.021.
Ferrara N. Vascular endothelial growth factor: basic science and clinical progress. Endocr Rev. 2004;25(4):581–611. https://doi.org/10.1210/er.2003-0027.
Jiang BH, Liu LZ. Chapter 2 PI3K/PTEN signaling in angiogenesis and tumorigenesis. Adv Cancer Res. 2009;102(09):19–65. https://doi.org/10.1016/S0065-230X(09)02002-8.
Takahashi T, et al. A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-γ and DNA synthesis in vascular endothelial cells. EMBO J. 2001;20(11):2768–78. https://doi.org/10.1093/emboj/20.11.2768.
van Hinsbergh VWM, Koolwijk P. Endothelial sprouting and angiogenesis: matrix metalloproteinases in the lead. Cardiovasc Res. 2008;78(2):203–12. https://doi.org/10.1093/cvr/cvm102.
Weis S, et al. Endothelial barrier disruption by VEGF-mediated Src activity potentiates tumor cell extravasation and metastasis. J Cell Biol. 2004;167(2):223–9. https://doi.org/10.1083/jcb.200408130.
Niu G, Chen X. Vascular endothelial growth factor as an anti-angiogenic target for cancer therapy. Curr Drug Targets. 2010;11(8):1000–17. https://doi.org/10.2174/138945010791591395.
Bell C, et al. Oligonucleotide NX1838 inhibits VEGF165-mediated cellular responses in vitro. In Vitro cell Dev Biol Anim. 1999;35(9):533–42. https://doi.org/10.1007/s11626-999-0064-y.
Lee JH, et al. A therapeutic aptamer inhibits angiogenesis by specifically targeting the heparin binding domain of VEGF165. Proc Natl Acad Sci. 2005b;102(52):18902–7. https://doi.org/10.1073/pnas.0509069102.
Lin YS, et al. Preclinical pharmacokinetics, interspecies scaling, and tissue distribution of a humanized monoclonal antibody against vascular endothelial growth factor. J Pharmacol Exp Ther. 1999;288(1):371–8.
Lockhart AC, et al. Phase I study of intravenous vascular endothelial growth factor trap, aflibercept, in patients with advanced solid tumors. J Clin Oncol. 2010;28(2):207–14. https://doi.org/10.1200/JCO.2009.22.9237.
Zhou B, Wang B. Pegaptanib for the treatment of age-related macular degeneration. Exp Eye Res. 2006;83(3):615–9. https://doi.org/10.1016/j.exer.2006.02.010.
Spratlin JL, et al. Phase I pharmacologic and biologic study of ramucirumab (IMC-1121B), a fully human immunoglobulin G1 monoclonal antibody targeting the vascular endothelial growth factor receptor-2. J Clin Oncol. 2010;28(5):780–7. https://doi.org/10.1200/JCO.2009.23.7537.
Faivre S, et al. Safety, pharmacokinetic, and antitumor activity of SU11248, a novel oral multitarget tyrosine kinase inhibitor, in patients with cancer. J Clin Oncol. 2006;24(1):25–35. https://doi.org/10.1200/JCO.2005.02.2194.
Hutson TE, et al. Efficacy and safety of pazopanib in patients with metastatic renal cell carcinoma. J Clin Oncol. 2010;28(3):475–80. https://doi.org/10.1200/JCO.2008.21.6994.
Iwamoto FM, et al. Phase II trial of pazopanib (GW786034), an oral multi-targeted angiogenesis inhibitor, for adults with recurrent glioblastoma (North American Brain Tumor Consortium Study 06-02). Neuro-Oncology. 2010;12(8):855–61. https://doi.org/10.1093/neuonc/noq025.
Sloan B, Scheinfeld NS. Pazopanib, a VEGF receptor tyrosine kinase inhibitor for cancer therapy. Curr Opin Investig Drugs. 2008;9(12):1324–35.
Strumberg D, et al. Phase I clinical and pharmacokinetic study of the Novel Raf kinase and vascular endothelial growth factor receptor inhibitor BAY 43-9006 in patients with advanced refractory solid tumors. J Clin Oncol. 2005;23(5):965–72. https://doi.org/10.1200/JCO.2005.06.124.
Wilhelm SM, et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004;64(19):7099–109. https://doi.org/10.1158/0008-5472.CAN-04-1443.
Zirlik K, Duyster J. Anti-angiogenics: current situation and future perspectives. Oncol Res Treat. 2018;41(4):166–71. https://doi.org/10.1159/000488087.
Jimenez-Pascual A, et al. FGF2: a novel druggable target for glioblastoma? Expert Opin Ther Targets. 2020;24(4):311–8. https://doi.org/10.1080/14728222.2020.1736558.
Turner N, Grose R. Fibroblast growth factor signalling: from development to cancer. Nat Rev Cancer. 2010;10(2):116–29. https://doi.org/10.1038/nrc2780.
Yu P, et al. FGF-dependent metabolic control of vascular development. Nature. 2017;545(7653):224–8. https://doi.org/10.1038/nature22322.
Incio J, et al. Obesity promotes resistance to anti-VEGF therapy in breast cancer by up-regulating IL-6 and potentially FGF-2. Sci Transl Med. 2018;10(432):eaag0945. https://doi.org/10.1126/scitranslmed.aag0945.
Brady N, et al. The FGF/FGFR axis as a therapeutic target in breast cancer. Expert Rev Endocrinol Metab. 2013;8(4):391–402. https://doi.org/10.1586/17446651.2013.811910.
Jain VK, Turner NC. Challenges and opportunities in the targeting of fibroblast growth factor receptors in breast cancer. Breast Cancer Res. 2012;14(3):208. https://doi.org/10.1186/bcr3139.
Kloepper J, et al. Ang-2/VEGF bispecific antibody reprograms macrophages and resident microglia to anti-tumor phenotype and prolongs glioblastoma survival. Proc Natl Acad Sci U S A. 2016;113(16):4476–81. https://doi.org/10.1073/pnas.1525360113.
Peterson TE, et al. Dual inhibition of Ang-2 and VEGF receptors normalizes tumor vasculature and prolongs survival in glioblastoma by altering macrophages. Proc Natl Acad Sci U S A. 2016;113(16):4470–5. https://doi.org/10.1073/pnas.1525349113.
Adams RH, Klein R. Eph receptors and ephrin ligands. Essential mediators of vascular development. Trends Cardiovasc Med. 2000;10(5):183–8. https://doi.org/10.1016/s1050-1738(00)00046-3.
Holder N, Klein R. Eph receptors and ephrins: effectors of morphogenesis. Development. 1999;126(10):2033–44.
Kullander K, Klein R. Mechanisms and functions of Eph and ephrin signalling. Nat Rev Mol Cell Biol. 2002;3(7):475–86. https://doi.org/10.1038/nrm856.
Dodelet VC, Pasquale EB. Eph receptors and ephrin ligands: embryogenesis to tumorigenesis. Oncogene. 2000;19(49):5614–9. https://doi.org/10.1038/sj.onc.1203856.
Ogawa K, et al. The ephrin-A1 ligand and its receptor, EphA2, are expressed during tumor neovascularization. Oncogene. 2000;19(52):6043–52. https://doi.org/10.1038/sj.onc.1204004.
Surawska H, Ma PC, Salgia R. The role of ephrins and Eph receptors in cancer. Cytokine Growth Factor Rev. 2004;15(6):419–33. https://doi.org/10.1016/j.cytogfr.2004.09.002.
Noren NK, et al. Interplay between EphB4 on tumor cells and vascular ephrin-B2 regulates tumor growth. Proc Natl Acad Sci U S A. 2004;101(15):5583–8. https://doi.org/10.1073/pnas.0401381101.
Uhl C, et al. EphB4 mediates resistance to antiangiogenic therapy in experimental glioma. Angiogenesis. 2018;21(4):873–81. https://doi.org/10.1007/s10456-018-9633-6.
Krusche B, et al. EphrinB2 drives perivascular invasion and proliferation of glioblastoma stem-like cells. elife. 2016;5:1–32. https://doi.org/10.7554/elife.14845.
Brantley DM, et al. Soluble Eph A receptors inhibit tumor angiogenesis and progression in vivo. Oncogene. 2002;21(46):7011–26. https://doi.org/10.1038/sj.onc.1205679.
Cheng N, et al. Inhibition of VEGF-dependent multistage carcinogenesis by soluble EphA receptors. Neoplasia. 2003;5(5):445–56. https://doi.org/10.1016/s1476-5586(03)80047-7.
Dobrzanski P, et al. Antiangiogenic and antitumor efficacy of EphA2 receptor antagonist. Cancer Res. 2004;64(3):910–9. https://doi.org/10.1158/0008-5472.can-3430-2.
Berta J, et al. Apelin expression in human non-small cell lung cancer: role in angiogenesis and prognosis. J Thorac Oncol. 2010;5(8):1120–9. https://doi.org/10.1097/JTO.0b013e3181e2c1ff.
Feng M, et al. Tumor apelin, not serum apelin, is associated with the clinical features and prognosis of gastric cancer. BMC Cancer. 2016;16(1):1–8. https://doi.org/10.1186/s12885-016-2815-y.
Heo K, et al. Hypoxia-induced up-regulation of apelin is associated with a poor prognosis in oral squamous cell carcinoma patients. Oral Oncol. 2012;48(6):500–6. https://doi.org/10.1016/j.oraloncology.2011.12.015.
Kälin RE, et al. Paracrine and autocrine mechanisms of apelin signaling govern embryonic and tumor angiogenesis. Dev Biol. 2007;305(2):599–614. https://doi.org/10.1016/j.ydbio.2007.03.004.
Lacquaniti A, et al. Apelin beyond kidney failure and hyponatremia: a useful biomarker for cancer disease progression evaluation. Clin Exp Med. 2015;15(1):97–105. https://doi.org/10.1007/s10238-014-0272-y.
Seaman S, et al. Genes that distinguish physiological and pathological angiogenesis. Cancer Cell. 2007;11(6):539–54. https://doi.org/10.1016/j.ccr.2007.04.017.
Tolkach Y, et al. Apelin and apelin receptor expression in renal cell carcinoma. Br J Cancer. 2019;120(6):633–9. https://doi.org/10.1038/s41416-019-0396-7.
Wysocka MB, Pietraszek-Gremplewicz K, Nowak D. The role of apelin in cardiovascular diseases, obesity and cancer. Front Physiol. 2018;9:557. https://doi.org/10.3389/fphys.2018.00557.
Berta J, et al. Apelin promotes lymphangiogenesis and lymph node metastasis. Oncotarget. 2014;5(12):4426–37. https://doi.org/10.18632/oncotarget.2032.
Hall C, et al. Inhibition of the apelin/apelin receptor axis decreases cholangiocarcinoma growth. Cancer Lett. 2017;386:179–88. https://doi.org/10.1016/j.canlet.2016.11.025.
Lv D, et al. PAK1-cofilin phosphorylation mediates human lung adenocarcinoma cells migration induced by apelin-13. Clin Exp Pharmacol Physiol. 2016;43(5):569–79. https://doi.org/10.1111/1440-1681.12563.
Macaluso NJM, et al. Discovery of a competitive apelin receptor (APJ) antagonist. ChemMedChem. 2011;6(6):1017–23. https://doi.org/10.1002/cmdc.201100069.
Harford-Wright E, et al. Pharmacological targeting of apelin impairs glioblastoma growth. Brain. 2017;140(11):2939–54. https://doi.org/10.1093/brain/awx253.
Mastrella G, et al. Targeting APLN/APLNR improves antiangiogenic efficiency and blunts proinvasive side effects of VEGFA/VEGFR2 blockade in glioblastoma. Cancer Res. 2019;79(9):2298–313. https://doi.org/10.1158/0008-5472.CAN-18-0881.
Uribesalgo I, et al. Apelin inhibition prevents resistance and metastasis associated with anti-angiogenic therapy. EMBO Mol Med. 2019;11(8):e9266. https://doi.org/10.15252/emmm.201809266.
Chae S, et al. Angiopoietin-2 interferes with anti-VEGFR2- induced vessel normalization and survival benefit in mice bearing gliomas. Clin Cancer Res. 2010;16(14):3618–27. https://doi.org/10.1158/1078-0432.CCR-09-3073.Angiopoietin-2.
Shim WSN, Ho IAW, Wong PEH. Angiopoietin: a TIE(d) balance in tumor angiogenesis. Mol Cancer Res. 2007;5(7):655–65. https://doi.org/10.1158/1541-7786.MCR-07-0072.
Ping YF, Bian XW. Consice review: contribution of cancer stem cells to neovascularization. Stem Cells. 2011;29(6):888–94. https://doi.org/10.1002/stem.650.
Bach P, et al. Specific elimination of CD133+ tumor cells with targeted oncolytic measles virus. Cancer Res. 2013;73(2):865–74. https://doi.org/10.1158/0008-5472.CAN-12-2221.
Xu MR, et al. Brucine suppresses vasculogenic mimicry in human triple-negative breast cancer cell line MDA-MB-231. In: W.-L. Lu, Editor. Biomed Res Int. 2019;2019:6543230. https://doi.org/10.1155/2019/6543230.
Serwe A, et al. Inhibition of TGF-β signaling, vasculogenic mimicry and proinflammatory gene expression by isoxanthohumol. Investig New Drugs. 2012;30(3):898–915. https://doi.org/10.1007/s10637-011-9643-3.
Kumar SR, et al. Molecular targets for tivantinib (ARQ 197) and vasculogenic mimicry in human melanoma cells. Eur J Pharmacol. 2019;853:316–24. https://doi.org/10.1016/j.ejphar.2019.04.010.
Fernández-Cortés M, Delgado-Bellido D, Javier Oliver F. Vasculogenic mimicry: become an endothelial cell “But not so much”. Front Oncol. 2019;9(AUG):1–6. https://doi.org/10.3389/fonc.2019.00803.
Jean C, et al. Inhibition of endothelial FAK activity prevents tumor metastasis by enhancing barrier function. J Cell Biol. 2014;204(2):247–63. https://doi.org/10.1083/jcb.201307067.
Goel S, et al. Effects of vascular-endothelial protein tyrosine phosphatase inhibition on breast cancer vasculature and metastatic progression. J Natl Cancer Inst. 2013;105(16):1188–201. https://doi.org/10.1093/jnci/djt164.
George D. Platelet-derived growth factor receptors: a therapeutic target in solid tumors. Semin Oncol. 2001;28:27–33. https://doi.org/10.1016/S0093-7754(01)90100-9.
Thijssen VL, et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. J Pathol. 2018;246(4):447–58. https://doi.org/10.1002/path.5152.
Uehara H, et al. Effects of blocking platelet-derived growth factor-receptor signaling in a mouse model of experimental prostate cancer bone metastases. J Natl Cancer Inst. 2003;95(6):458–70. https://doi.org/10.1093/jnci/95.6.458.
Heidemann J, et al. Angiogenic effects of interleukin 8 (CXCL8) in human intestinal microvascular endothelial cells are mediated by CXCR2. J Biol Chem. 2003;278(10):8508–15. https://doi.org/10.1074/jbc.M208231200.
Kitadai Y, et al. Regulation of disease-progression genes in human gastric carcinoma cells by interleukin 8. Clin Cancer Res. 2000;6(7):2735–40.
Yang G, et al. CXCR2 promotes ovarian cancer growth through dysregulated cell cycle, diminished apoptosis, and enhanced angiogenesis. Clin Cancer Res. 2010;16(15):3875–86. https://doi.org/10.1158/1078-0432.CCR-10-0483.
Ijichi H, et al. Inhibiting Cxcr2 disrupts tumor-stromal interactions and improves survival in a mouse model of pancreatic ductal adenocarcinoma. J Clin Invest. 2011;121(10):4106–17. https://doi.org/10.1172/JCI42754.
Xu J, et al. Vascular CXCR4 expression promotes vessel sprouting and sensitivity to sorafenib treatment in hepatocellular carcinoma. Clin Cancer Res. 2017;23(15):4482–92. https://doi.org/10.1158/1078-0432.ccr-16-2131.
Chen L, et al. The IL-8/CXCR1 axis is associated with cancer stem cell-like properties and correlates with clinical prognosis in human pancreatic cancer cases. Sci Rep. 2014;4:1–7. https://doi.org/10.1038/srep05911.
Ha H, Debnath B, Neamati N. Role of the CXCL8-CXCR1/2 axis in cancer and inflammatory diseases. Theranostics. 2017;7(6):1543–88. https://doi.org/10.7150/thno.15625.
Milosevic V, et al. Wnt/IL- 1β/IL -8 autocrine circuitries control chemoresistance in mesothelioma initiating cells by inducing ABCB5. Int J Cancer. 2020;146(1):192–207. https://doi.org/10.1002/ijc.32419.
Smith DR, et al. Inhibition of interleukin 8 attenuates angiogenesis in bronchogenic carcinoma. J Exp Med. 1994;179(5):1409–15. https://doi.org/10.1084/jem.179.5.1409.
Martin D, Galisteo R, Gutkind JS. CXCL8/IL8 stimulates vascular endothelial growth factor (VEGF) expression and the autocrine activation of VEGFR2 in endothelial cells by activating NFkappaB through the CBM (Carma3/Bcl10/Malt1) complex. J Biol Chem. 2009;284(10):6038–42. https://doi.org/10.1074/jbc.C800207200.
Scapini P, et al. CXCL1/macrophage inflammatory protein-2-induced angiogenesis in vivo is mediated by neutrophil-derived vascular endothelial growth factor-A. J Immunol. 2004;172(8):5034–40. https://doi.org/10.4049/jimmunol.172.8.5034.
Wolf MJ, et al. Endothelial CCR2 signaling induced by colon carcinoma cells enables extravasation via the JAK2-Stat5 and p38MAPK pathway. Cancer Cell. 2012;22(1):91–105. https://doi.org/10.1016/j.ccr.2012.05.023.
Chen X, et al. CCL2/CCR2 regulates the tumor microenvironment in HER-2/neu-driven mammary carcinomas in mice. PLoS One. 2016;11(11):e0165595. https://doi.org/10.1371/journal.pone.0165595.
Oh P, et al. Subtractive proteomic mapping of the endothelial surface in lung and solid tumours for tissue-specific therapy. Nature. 2004;429(6992):629–35. https://doi.org/10.1038/nature02580.
Wei YQ, et al. Immunogene therapy of tumors with vaccine based on Xenopus homologous vascular endothelial growth factor as a model antigen. Proc Natl Acad Sci U S A. 2001;98(20):11545–50. https://doi.org/10.1073/pnas.191112198.
Gavilondo JV, et al. Specific active immunotherapy with a VEGF vaccine in patients with advanced solid tumors. results of the CENTAURO antigen dose escalation phase I clinical trial. Vaccine. 2014;32(19):2241–50. https://doi.org/10.1016/j.vaccine.2013.11.102.
Chen R, et al. Anti-metastatic effects of DNA vaccine encoding single-chain trimer composed of MHC I and vascular endothelial growth factor receptor 2 peptide. Oncol Rep. 2015;33(5):2269–76. https://doi.org/10.3892/or.2015.3820.
Ishizaki H, et al. Inhibition of tumor growth with antiangiogenic cancer vaccine using epitope peptides derived from human vascular endothelial growth factor receptor 1. Clin Cancer Res. 2006;12(19):5841–9. https://doi.org/10.1158/1078-0432.CCR-06-0750.
Liang P, et al. Construction of a DNA vaccine encoding Flk-1 extracellular domain and C3d fusion gene and investigation of its suppressing effect on tumor growth. Cancer Immunol Immunother. 2010;59(1):93–101. https://doi.org/10.1007/s00262-009-0727-2.
Liu JY, et al. Immunotherapy of tumors with vaccine based on quail homologous vascular endothelial growth factor receptor-2. Blood. 2003;102(5):1815–23. https://doi.org/10.1182/blood-2002-12-3772.
McKinney KA, et al. Effect of a novel DNA vaccine on angiogenesis and tumor growth in vivo. Arch Otolaryngol. 2010;136(9):859–64. https://doi.org/10.1001/archoto.2010.139.
Wada S, et al. Rationale for antiangiogenic cancer therapy with vaccination using epitope peptides derived from human vascular endothelial growth factor receptor 2. Cancer Res. 2005;65(11):4939–46. https://doi.org/10.1158/0008-5472.CAN-04-3759.
Xie K, et al. Anti-tumor effects of a human VEGFR-2-based DNA vaccine in mouse models. Genet Vaccines Ther. 2009;7:10. https://doi.org/10.1186/1479-0556-7-10.
Zuo SG, et al. Orally administered DNA vaccine delivery by attenuated Salmonella typhimurium targeting fetal liver kinase 1 inhibits murine Lewis lung carcinoma growth and metastasis. Biol Pharm Bull. 2010;33(2):174–82. https://doi.org/10.1248/bpb.33.174.
He Q, et al. Inhibition of tumor growth with a vaccine based on xenogeneic homologous fibroblast growth factor receptor-1 in mice. J Biol Chem. 2003;278(24):21831–6. https://doi.org/10.1074/jbc.M300880200.
Li M, et al. bFGF peptide combined with the pVAX-8CpG plasmid as adjuvant is a novel anticancer vaccine inducing effective immune responses against Lewis lung carcinoma. Mol Med Rep. 2012;5(3):625–30. https://doi.org/10.3892/mmr.2011.725.
Plum SM, et al. Administration of a liposomal FGF-2 peptide vaccine leads to abrogation of FGF-2-mediated angiogenesis and tumor development. Vaccine. 2000;19(9–10):1294–303. https://doi.org/10.1016/s0264-410x(00)00210-3.
Zheng SJ, et al. Synergistic anti-tumor effect of recombinant chicken fibroblast growth factor receptor-1-mediated anti-angiogenesis and low-dose gemcitabine in a mouse colon adenocarcinoma model. World J Gastroenterol. 2007;13(17):2484–9. https://doi.org/10.3748/wjg.v13.i17.2484.
Plum SM, et al. Generation of a specific immunological response to FGF-2 does not affect wound healing or reproduction. Immunopharmacol Immunotoxicol. 2004;26(1):29–41. https://doi.org/10.1081/iph-120029942.
McNeel DG, et al. Phase I trial of a monoclonal antibody specific for alphavbeta3 integrin (MEDI-522) in patients with advanced malignancies, including an assessment of effect on tumor perfusion. Clin Cancer Res. 2005;11(21):7851–60. https://doi.org/10.1158/1078-0432.CCR-05-0262.
Huang FY, et al. Bacterial surface display of endoglin by antigen 43 induces antitumor effectiveness via bypassing immunotolerance and inhibition of angiogenesis. Int J Cancer. 2014;134(8):1981–90. https://doi.org/10.1002/ijc.28511.
Jarosz M, et al. Therapeutic antitumor potential of endoglin-based DNA vaccine combined with immunomodulatory agents. Gene Ther. 2013;20(3):262–73. https://doi.org/10.1038/gt.2012.28.
Fernández Lorente A, et al. Effect of blockade of the EGF system on wound healing in patients vaccinated with CIMAvax® EGF. World J Surg Oncol. 2013;11:275. https://doi.org/10.1186/1477-7819-11-275.
García B, et al. Effective inhibition of the epidermal growth factor/epidermal growth factor receptor binding by anti-epidermal growth factor antibodies is related to better survival in advanced non-small-cell lung cancer patients treated with the epidermal growth factor. Clin Cancer Res. 2008;14(3):840–6. https://doi.org/10.1158/1078-0432.CCR-07-1050.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Milosevic, V., Edelmann, R.J., Fosse, J.H., Östman, A., Akslen, L.A. (2022). Molecular Phenotypes of Endothelial Cells in Malignant Tumors. In: Akslen, L.A., Watnick, R.S. (eds) Biomarkers of the Tumor Microenvironment. Springer, Cham. https://doi.org/10.1007/978-3-030-98950-7_3
Download citation
DOI: https://doi.org/10.1007/978-3-030-98950-7_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-98949-1
Online ISBN: 978-3-030-98950-7
eBook Packages: MedicineMedicine (R0)