
Abstract
In this chapter we address factors that may bias experiments and impact results unless they are controlled for. This apply for factors in the environment that the animal interacts with to assure optimal homeostasis or to fulfil basic needs. It also includes intrinsic properties of the animal themselves that should be taken into consideration when designing studies and applying results from animal research.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
- Therioepistemology
- Environment
- Sex
- Circadian rhythm
- Hormones
- Reproduction
- Enrichment
- Diet
- Water
- Noise
- Vibration
- Temperature
- Humidity
1 Introduction
Animals used in research must cope with the environment we provide them, and their homeostatic system must adjust to changes in this environment to maintain optimal physiology. Changes in this environment cause the animal to adapt to new conditions by physiologic accommodation. This response may affect experimental outcomes, be a confounding variable or a source of variation. In order to obtain reliable, meaningful results, an attempt should be made to understand and account for all known biological, environmental and social factors when conducting experiments involving animals. This may mean changing experimental designs to maximise variation [1] or listing a wide variety of environmental and animal information in supplemental materials [2]. This chapter will focus on biological and environmental factors that affect animal physiology and thereby also output of experiments. These are also factors useful to assess in comparing studies and addressing questions of non-reproducibility.
In chapter “Rodent Genetics”, the authors of that chapter address the genetics of rodents and how this can have an impact on the reproducibility of animal experiments. Even though we will not consider genetics in this chapter, the factors we will discuss here are dependent and related to the genome of the animal. What is more, there is also an interdependence between the genome, the microbiome (addressed in chapter “Microbiology and Microbiome”) and the other factors described in this chapter [3,4,5,6]. We cannot separate them, and they need to be treated together as potential causes of variability leading to irreproducible animal research if not taken into account in the experimental design (see also Part II in this book dealing with statistics and experimental design).
1.1 Therioepistemology
In 2017, Garner and collaborators introduced the term therioepistemology to describe the study of how knowledge is gained from animal research. They coined the word from the study of the theory of knowledge and the mechanisms by which rational inference is formed, known as epistemology, and the prefix therio, of animals [2]. They proposed six questions to help address the problem of those factors in animal research that we cannot control but should acknowledge instead of ignoring them [2]:
-
1.
What features of model biology are ignored? Physiological, anatomical and behavioural particularities of the animal model will differ from humans; nevertheless, they are consistently ignored. We have dealt with some in this chapter but also in chapters “Rodent Genetics” and “Microbiology and Microbiome” of this book.
-
2.
What features of human biology are ignored? Sometimes it is not the characteristics of the animal model that are ignored but the human condition in itself; thus, we try to replicate human pathologies using genetically modified mice that can only replicate partially a molecular pathway or mimic a specific aspect of a much broader human pathology.
-
3.
What features of the measures are ignored? Part of this problem is how to deal with type I errors, which is consistently ignored throughout the animal research literature. The reader can find more on this question on chapters “Statistical Tests and Sample Size Calculations”, “Design of Experiments” and “Scholarly Publishing and Scientific Reproducibility”, dealing with statistics and experimental design.
-
4.
What features of background methodology and husbandry are ignored? This relates to the well-known justification of the historical standardisation, i.e. if a model works, why change the conditions, even though there could potentially be refinements and improvements made? The answer will be that if a model stops working under different experimental backgrounds, it lacks external validity, and, thus, it will not translate to humans.
-
5.
What animal well-being issues are ignored? Preclinical research should be treated exactly the same as clinical research, i.e. the animal should be seen as a patient, not as a reagent. The reader is also referred to chapter “Systematic Reviews” in this book.
-
6.
What principles of experimental design and statistics are ignored? In general, preclinical research lacks proper experimental design and statistics. The reader is referred to chapters “Statistical Tests and Sample Size Calculations”, “Design of Experiments” and “Scholarly Publishing and Scientific Reproducibility” for more on this.
1.2 Standardisation of the Environment
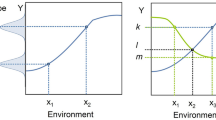
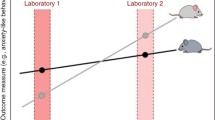
It has been shown that environmental standardisation will increase test sensitivity, which is the proportion of correctly identified data, and reduce the variation in the obtained data, but, as Richter et al. [1] questioned, will these lead to an increase in reproducibility? The answer is no, as they found in a set of experiments with 18 standardised replicated cohorts and 18 heterogenised replicated cohorts. Through heterogenisation, they better understood the systematic variation of the experimental conditions. Environmental standardisation resulted in poorer reproducibility and introduced a systematic source of false-positive results above that expected by chance alone. This work was criticised because the conclusions were based on a retrospective analysis. The authors went on to do a prospective study and also found that standardised experiments increased test sensitivity at the expense of external validity, i.e. the applicability of a result to other conditions, populations or species [7]. For this study, they used 3 behavioural tests on 256 female mice from 2 different strains, C57BL/6 and BALB/c, within 4 standardised and 4 heterogenised cohorts taking 36 different behavioural measures. They confirmed their previous findings [1] and that even simple forms of heterogenisation may guarantee robust results across experiments [7]. This is something that Michael Festing had already proposed, in terms of the genome, 30 years earlier. In 1980, he published his first proposition to change the way toxicology was carried out in outbred stocks by using instead several inbreds of F1 hybrid strains in a factorial experiment design [8].
2 The Animal Sex
Sex is an important variable to consider, not only for the obvious reason that several physiological processes are different between sexes but also because in a majority of manuscripts either the sex of the animals is not reported or only one sex is used. This is a major source of reduced external validity of those studies. When sex is reported, it is readily apparent there is a generalised male bias across preclinical research disciplines, and this has been criticised for several years. In 2011, Beery et al. [9] reported male bias in eight out of ten fields analysed, including pharmacology, endocrinology, animal behaviour, behavioural physiology, neuroscience, general biology, zoology and physiology. They also found a female skew in the fields of reproduction and immunology, although in this last field less than 40% of manuscripts reported the sex of the animals used [9]. The reasons for omitting one sex, mainly females, in preclinical studies have been poorly justified, consisting of citation of confounding hormonal variations during the oestrus cycle [10, 11], a reduction in statistical power by the introduction of a second sex [10] and historical reasons [11]. This exclusion of one sex in preclinical research and the consequent inadequate analysis has also been cited as a reason for the lack of reproducibility in preclinical research [11]. On the 25th of January 2016, the National Institutes of Health (NIH) implemented the requirement to consider sex as a biological variable within their grant submissions [12]. Nevertheless, some researchers have protested this policy, arguing that to adequately design experiments using both sexes will be more expensive and space-intensive [13] and will result in unnecessary duplication and slow the progress due to more workload [14]. If we, as scientists, are truly serious about moving forwards from a lack of reproducibility, these arguments are both specious and detrimental. Clearly, both sexes need to be included in scientific endeavours, and using just one sex based on those arguments will hinder not only reproducibility but also translational research. As Cara Tannenbaum et al. [15] clearly expressed, “researchers and peer reviewers are being asked to thoughtfully consider whether a single-sex study is justified when research results are to be applied to both sexes”. She proposed a set of questions for peer reviewers to consider that we have included here (Table 1).
Physiological processes which are different between sexes include pain and its control, which clearly affects not just pain research but any research which will cause some degree of pain that must be relieved or it will otherwise interfere with the aim of the study. In 2011, Robert E. Sorge, then working in the laboratory of Jeffrey S. Mogil, made a seminal discovery in pain research, namely, that the afferent pain pathway is different in female and male mice [16]. Whereas in male mice, microglia have a major role in pain sensitivity, they realised by working in both sexes that in female mice T cells, instead of microglia, are the preferred afferent pathway. What is more, they discovered that testosterone is the switch that allows the use of microglia instead of T cells by using castrated males and intact females [17]. Since then other discoveries have been made to explain, for example, the attenuated response to morphine observed in females [18]. Thus, unless laboratories were using both sexes, preclinical research on pain would reach very different outcomes depending on the sex of animals used. Memory research is another field of neuroscience that has been shown to be biased because of the use of only male animals [19]. It has been demonstrated that females show a more prominent basal amygdala activation compared with hippocampus activation in males during memory retrieval [20]. In cardiovascular research, we now know that the ability of ventricular myocytes to contract declines with age in male rodents more than in female rodents [21]. There are also well-documented differences in haematology and biochemical analytes between sexes of the same species and even the same strain [22].
2.1 Circadian Rhythm
Circadian rhythm has a significant impact in animal physiology [23, 24]. Circadian rhythms are generated by solar time, with photons impacting the cells in the retina, which in turn send electric signals to the suprachiasmatic nucleus (SCN) within the hypothalamus through the retino-hypothalamic tract. These electric signals will cause SCN neurons spontaneously firing. Signals from the SCN will travel then to the hypothalamus, cortex, brain stem and the different circadian clocks around the body [25]. The retinal evoked firing in the SCN will only persist for the duration of the light pulse [25]. This is the mechanistic reason for the physiological changes evoked by light of enough intensity during the night part of the cycle, which have been documented to occur after a light pulse of less than a second.
Adaption to seasonal changes is an important quality for survival and reproduction, and this quality is deeply conserved in animals even after many generations in captivity. Light, and change in daylight, is an important regulator of the reproduction cycle of many species, and disturbances in light cycle may be responsible for drops in reproductive performance [26]. Standardised light regimes are commonly used to control circadian rhythm variations. Attention should also be paid to indirect light coming through inspection windows, light leakage around doors and daylight in adjacent corridors [27]. Furthermore, attention must be paid to light exposure when animals are brought from animal holding rooms to specially equipped laboratories (imaging, telemetry, behavioural suites, etc.). Laboratories built for humans are usually equipped with windows to allow daylight in and also with a higher light intensity than the one found in the animal holding rooms. The European Guidelines for the Accommodation and Care of Animals Used for Experimental and Other Scientific Purposes (ETS 123) [28] defines standards for light/dark cycle in rodent facilities as typically 12 hours of dark and 12 hours of light, but also other light regimes with longer or shorter day periods are used depending on the species [29]. Artificial induction of seasonal change has been accomplished by modulation of light/dark cycles to expedite or delay developmental stages and reproduction performance [30].
Rodents are nocturnal animals and normally sleep during the daytime. Received wisdom states that frequent sleep disturbance during daytime due to daily routines in the animal facility may cause stress and sleep deprivation in the animals, although this may not be true [31]. Even though efforts have been made to standardise light/dark cycles in duration, few labs keep their animals on reverse day/night cycles, and most experiments are still performed during daytime (since humans trend to be diurnal) [32]. The observation of clinical signs and animal welfare assessments is difficult to perform during daylight hours as animals do not express normal night-time activity levels when they rest. ETS 123 therefore recommends some observation of animals under frequencies of red light undetectable to the rodents, as this is not perceived as bright daylight by rodents [33]. However, this has recently been challenged in an article by Niklaus and collaborators [34] where they claimed that rats are sensitive to light wavelengths longer than 620 nm, reaching opposite results to previous work by De Farias Rocha and collaborators [35], by using a different experimental setup. The question, thus, remains certainly open, but more work needs to be done to support one or the other.
Animal facilities have, more and more, automated means of controlling light/dark periods and light intensity within the room. Thus, we rely on systems such as building management systems software to keep the room environment within the regulatory limits. These systems sometimes fail, and unless there are strong processes in place, there may be a gap of hours or days before the staff working in the facility realise there is a problem. One of the more common failures is constant light exposure due to a failure to start the dark cycle. Mice will increase body weight and become insulin resistant with constant light exposure [36]. A similar effect has been observed in rats, with constant light reducing glucose-stimulated insulin secretion due to a disruption in the pancreatic beta cell circadian clock [37] and accelerating the development of diabetes in transgenic rats for human islet amyloid polypeptide [38].
2.2 Light and the Laboratory Mouse
Light quality (referring here to the spectral composition of light), and its influence in the circadian regulation, is not generally considered by current regulations [28, 39]. However, this can be an important source of variability between different laboratories, especially those working with certain mouse strains or with rats. Thus, most animal facilities do not pay due attention to this important factor, apart from controlling the intensity to avoid rodent light retinopathy and to establish a constant light/dark cycle throughout the year [40]. Research has found that the more light in the 465–485 nm wavelength, which is the blue colour of the visible light spectrum, the better the animal health and welfare compared to cool white fluorescent (CWF) light [41]. Melatonin has been shown to be six- to sevenfold higher in rodents under this blue-appearing wavelength than with CWF lighting [41, 42]. There is now consensus on how to quantify and report light stimuli in experimental studies [43, 44], which should be used to harmonise reporting and thus improve the reproducibility of experimental work between laboratories [41].
2.3 Age and Developmental Stage of the Animal
Studies using animals often use animals at one age, or body weight group, so the external validity of such studies is limited to that age or body weight. This can be critical when other researchers try to reproduce the experiments, especially when the publication does not provide any details of the age or life stage of the animals used. Not only that, but when interpreting the results, the difference found in the data might be the consequence of a normal age-related maturity process taking place in the particular age range chosen for the study like puberty or the beginning of feeding in fish larvae.
The Jackson Laboratory conducted a study with 31 different inbred strains and published data on median lifespans and circulating IGF1 levels at 6, 12 and 18 months for the first cohort of 32 females and 32 males of each strain [45]. They documented that males from C57BL/6 J or 129S1/SvImJ lived twice as long as males from FVB/NJ. These are just three of the inbred strains in which genetically modified mice have been produced worldwide and are used in most, if not all, of the studies using genetically modified mice. Thus, studies using animals in these different backgrounds, but the same genetic modification should take into account the differences in lifespan, which may account for maturation or aging processes that may affect the experimental data.
There are also many examples in the recent literature on how different organs and tissues will mature, and thus change, with age. For example, the developing spleen is an active haematopoietic centre from approximately day 15 of gestation until several weeks after birth in mice, rather than a secondary lymphoid organ as in the adult [46]. In humans, the experimental evidence is that the haematopoietic stem cells (HSCs) naturally migrate back and forth from the bone marrow periodically [47]. In vertebrates, the origins of the haematopoietic tissue are non-singular, with a shifting source and localisation over time. In total, the haematopoietic system is composed of HSCs, multiple terminally differentiated lineages and multiple intermediate committed progenitors [48]. In mice, the haematology goes through various stages as the animal ages. Thus, erythrocyte morphology in the young mouse is quite variable, and there is also a larger count of reticulocytes than in the adult mouse. Leukocytes, on the other hand, have a low count at birth to only reach adult numbers by 6–7 weeks of age. Depending on the mouse strain used, some haematological changes associated with age will be more pronounced. Another aging change in the haematology of mice is a reduction in the haematocrit due to a plasma expansion, which occurs with age and is often misinterpreted as anaemia [49, 50]. Table 2 shows normal haematological reference intervals for different inbred strains at 9 weeks of age [51]. These age-related changes have also been shown to occur in 26 biochemical analytes [22]. In 2008, Mazzaccara et al. [22] examined three mouse strains, C57BL/6 J, 129SV/Ev and C3H/HeJ, and showed that most of the biochemical analytes analysed differed according to age. They also evaluated five haematology parameters of which red blood cell counts, haemoglobin, haematocrit and platelet counts increased with the animal’s age only in C57BL/6 J mice [22] (for an excellent review on mouse haematology, we refer the reader to The Mouse in Biomedical Research, Volume III: Normative Biology, Husbandry and Models) [52].
Aging is another variable affecting studies using animal models. If the age of mice used in the study is not documented, results may be irreproducible. The mouse cochlea, for example, continues to mature during the first 2 weeks of life [53], and some strains of mice carry alleles causing age-related hearing loss [54, 55]. In the central nervous system, it has also been shown that pain modulation changes with age in rats from a facilitation of spinal pain transmission before day 21 of age to both facilitation and inhibition after 28 days of age [56,57,58]. Also, the heart changes with age and activity. There is epicardial fat deposition and aortic valve calcifications in older guinea pigs and rats [21]. There is also evidence for atrial hypertrophy and dilation in older rodents, and left ventricular wall thickness increases with age in older rats and mice [21]. There is also strong evidence that the heart’s responsiveness to β-adrenergic stimulation declines with age in animals [21]. In aged zebrafish, myocyte hypertrophy, increased ventricular density and fibrosis, valvular lesions and reductions in coronary vasculature have been described [59] (for an excellent review on age-associated changes in zebrafish, see Stoyek MR and collaborators review [59]).
2.4 Hormones and Reproduction
Reproductive performance and animal activity are very much influenced by circadian rhythm, and the reader is advised to read the Circadian Rhythm section for more information.
The mammalian nose contains the main olfactory epithelium, the septal organ of Masera, the vomeronasal organ (VNO) and the Grueneberg ganglion; all are related to olfactory functions including social communications. The major player in social communication through olfactory signals is the VNO, which collects information from the environment through the nose and vomeronasal duct, feeding the vomeronasal sensory neurons (VSNs) with chemicals taken up by the approximately 300 different vomeronasal receptors. The VSN axon passes through the ethmoid cribriform plate to access the accessory olfactory bulb, and from here reaches the amygdala and the hypothalamus [60]. The main olfactory system is also involved in eliciting behaviour from olfactory cues, together with the accessory olfactory system, and they both will interact at different levels in the CNS: olfactory bulbs, amygdala and hypothalamus [60]. The VNO conveys information about pheromones, which are anonymous signals, not being used to identify individuals; predators, the kairomones of Wyatt [61]; prey; and individual identity and may also identify pathogenic states in mice [60]. These olfactory cues in mice are very strongly shaped by a specific set of polymorphic communication proteins that has evolved to provide a distinctive signal of identity: the major urinary proteins (MUPs) [62]. These MUPs are a group of 18–20 kDa lipocalins involved in mouse chemical signalling, synthesised, in their majority, in the liver for excretion in the urine [63]. These MUPs are encoded by a cluster of 21 major urinary protein (MUP) genes on mouse chromosome 4 [62] and released at a high concentration in mouse urine. These genes are rearranged and expressed in a combinatorial form, particularly to each individual in a non-inbred population, and also discriminated through a set of vomeronasal sensory neurons using a combinatorial coding strategy [62]. These proteins are known to bind and slowly release volatile pheromones [63]. Some are involved in male aggression and attraction to females, like MUP20 [60]. There are also 38 exocrine gland-secreting peptide (ESP) genes; some of their translated products are involved in stopping male sexual behaviour (ESP22), and some in starting female lordosis (ESP1) [60].
Urine marking plays an important role in communication between female mice as well [64]. Some of the odours will have signalling effects, i.e. these odours will change the behaviour of other mice, whereas others will have primer effects, i.e. these odours will change the physiology of other mice. This primer effect is the cause of well-known reproductive effects in mice. Female mouse urine is known to contain pheromones and inhibit the reproductive physiology of other females under conditions linked to competition for reproductive opportunities, such as overcrowding. This is known as the Lee-Boot effect, which is a prolongation of the oestrus cycle in group-housed females [65, 66]. The key compound causing the Lee-Boot effect is 2,5-dimethylpyrazine [67], and its excretion is at its peak during metestrus. This compound has also been found to have a negative effect on male mice by depressing the maturation of reproductive organs and the level of immunocompetence [68]. If females are exposed to male urine pheromones, there is an induction of oestrus, a shortening of the oestrous cycle, and oestrus synchronisation of female mice; together this constellation of effects is known as the Whitten effect [69, 70]. Pregnancy failure, known as the Bruce effect [71, 72], is a phenomenon where pregnant rodents terminate their pregnancy after being exposed to the scent of an unfamiliar male. This occurs when female mice in early pregnancy are exposed to odour from an unfamiliar male at the same time as twice daily surges in their prolactin levels. This is stimulated by differences in low-molecular-weight urinary components that include MHC peptides or by differences in the amount of exocrine gland-secreting peptide 1 in male tear fluids, compared to the remembered stud male [62]. The unfamiliar male scent will trigger an increase of dopamine in the hypothalamus and a decrease of prolactin secretion from the anterior pituitary gland resulting in a subsequent decrease in progesterone, which is essential to maintain pregnancy, and the female returns to oestrus within a week [68]. Another primer effect is the accelerated onset of puberty in females exposed to male odours during their prepubertal period, also known as the Vandenbergh effect [73]. In this case, there has been found a correlation between the exposure to male odours and an activation of the posteroventral medial amygdala, posterodorsal medial amygdala, the anterior cortical nucleus of the amygdala, the medial preoptic area and the ventromedial nucleus, showing that these areas are differentially sensitive to intact male odours [74]. In both the Whitten and the Vandenbergh effects, several compounds have been potentially found to have an effect. These compounds have all a strong affinity to MUPs in male mice [75]. Male mouse odours have also a primer effect in male mice. The odour of dominant male mice suppresses sperm motility in subordinates [76].
The reader is directed to Sachiko Koyama’s excellent review for more information about the effects of primer marking in mice [68].
2.5 Handling
In 2010, Jane Hurst and Rebecca West published the results of a study that has had a profound impact in the husbandry of laboratory mice [77]. Briefly, they showed that picking up mice using tunnels or the open hand led to a voluntary approach from the animals, low anxiety and acceptance of physical restraint [77]. They also showed that picking them up by the tail induced aversion and high anxiety [77]. In a series of later publications, Jane Hurst and Kelly Gouveia demonstrated that mice do not even have to be familiar with the tunnel, although previous familiarisation helped in an outbred stock, for the anxiety levels to be reduced [78]; that tail handled mice performed poorly in behavioural studies, and this was only slightly improved by prior familiarisation [79]; that mice handled by tunnel explored readily and showed robust responses to test stimuli regardless of prior familiarisation or stimulus location [79]; that very brief handling (just 2 s) was sufficient to familiarise mice with tunnel handling, even when experienced only during cage cleaning [80]; and that experience of repeated immobilisation and subcutaneous injection did not reverse the positive effects of tunnel handling [80]. In spite of all this evidence, there are still laboratories and facilities picking up mice by the base of the tail, some of them even used sterile forceps, as a recent article by Henderson and collaborators [81] has shown. The group sent an online survey worldwide and received 390 complete responses to eight questions addressing the uptake of these non-aversive methods for handling mice. Even though most of the participants were aware of the benefit of using non-aversive handling methods, just 18% of them were using these methods exclusively, with 43% using a combination of non-aversive methods and tail handling and a 35% using only tail handling methods despite all the evidence against this [81]. The authors of this chapter speculate that this failure of uptake is due to concerns about transmission of infectious disease, the resistance of researchers to change that might affect “historical data” and concerns about disruption of established routines.
3 The Environment
3.1 Primary Enclosure (Cage, Pen and Tank)
3.1.1 Size of the Primary Enclosure
The European Union and the United States have defined minimum standards for enclosure dimensions and space allowances for housing research animals in their regulations and guidelines, which are rather similar [28, 39]. These usually define a minimum area (in m2 or cm2) per animal of a certain weight and state that all animals should be able to assume normal body postures. For example, the ethological needs of mice include resting, grooming, exploration, gnawing, nesting, hiding and social interaction, so at a minimum, consideration should be given to cage designs which allow performance of these behaviours. Both sets of regulations established a “one-size-fits-all” paradigm that does not necessarily correspond to the wide range of different breeds, stocks or strains used in the laboratory. What is more, there were no studies to sustain these arbitrary standards at the time of their publication. Even though mice are highly motivated to work for incremental space, there are no biological markers that will clearly indicate a negative effect with reduced space allocation. Attention should be paid to qualitative space, where animals can display the full behavioural repertoire, rather than to quantitative space, simply assigning a minimum area per animal [82].
The three-dimensional design of the primary enclosure is less well defined, though it has been shown that opportunity to use three-dimensional space is important for the development of the brain [83]. For many species, using both horizontal and vertical spaces provided is a natural behaviour. Implementation of “enrichment” programmes may meet some of the demands, but it is important that animals’ natural needs and behaviour are the focus of enrichment programmes and that any enrichment is consistently applied.
3.1.2 Enrichment of the Primary Enclosure
Enrichment of the barren cage environment to meet animals’ basic needs is now the default way to house research animals [28, 84] and is also regarded as refinement of animal research with the expectation to continuously refine enriched housing conditions based on updated information. Research animals are typically housed under conditions very different from their natural habitat and with limited opportunities to express normal behaviours. Such conditions impose constraints on behaviour and brain development and result in altered brain functions [85]. It has been shown that 2½-week-old rat pups already have a rudimentary map of space [86]. Histological examination of the brains of animals exposed to either a complex (“enriched”) environment compared to unenriched controls has revealed experience-induced morphological plasticity in the brain through life [83]. André and collaborators [87] checked 164 physiological parameters under three different conditions: no environmental enrichment, nesting and nesting and shelter. They found that nesting material and shelters may be used to improve animal welfare without impairment of experimental outcome or loss of comparability to previous data collected under barren housing conditions. These results and conclusions contrast with the ones obtained by Macri and collaborators in 2013 [88], where they claim that some effects of the synthetic compound JWH-018, a potent cannabinoid receptor agonist [89], are environmentally mediated. However, this article has many experimental design flaws potentially leading to bias, i.e. experimental groups differed in their conditions, the authors do not report randomisation or blinding, there is no indication of sample size calculations and they report the mean and standard error of the mean instead of the mean and the confidence interval [90]. Keeping animals without the ability to support their basic behavioural needs leads to suffering and distress that may as well be a confounding factor in experimental work. Animals may respond individually with either stereotypic behaviour, aggression, depression, self-mutilation or other maladaptive behaviour and their response cannot be standardised. It is important that responsible bodies (AWB, IACUC, AWERB or equal) develop and update enrichment programmes that take both animal welfare and scientific considerations into account.
3.1.3 Animal Position in the Room
The position of the cage in the rack and the rack within the room may affect a study as light and worker motion is not equally distributed in the room, and this may affect animal behaviour and results [91,92,93]. Light intensity will vary significantly between the top shelf, usually more brightly lit, and the cages on the bottom row, which are typically much dimmer. We know that mice find brightly lit, elevated spaces aversive, and this may influence results [94, 95]. In addition, the order of cage handling may bias results, e.g. all cages of one group are placed on the top row and always treated or measured early in the day, while those in a different group are placed at the bottom and are treated at the end. To assure proper randomisation in a study, cage position in the rack should also be randomised [96, 97], and in addition rotation of cage position in the rack during a study is recommended to avoid induction of systematic failure because of cage position.
3.2 Diet
Laboratory animals are fed a wide variety of diets, differing between laboratories and commercial companies. These diets, referred to as standard or regular diets, are made with natural ingredients such as soybean meal, alfalfa, fish meal and animal by-products, have variable nutritional content between batches and contain biologically active components such as phytoestrogens and toxic heavy metals such as arsenic [98, 99]. Diet ingredients cover the minimum requirements for the species and life stage and can be manufactured in various ways [100]. It is important that the chosen diet does not negatively influence the experiments, for example, by containing antinutrients or hormone-mimicking or blocking substances. Also, ingredients should be free of any chemicals, toxins, heavy metals or microbes, and documentation should be available from the producer on the quality analyses performed on different batches. The origin of dietary ingredients, processing and storing will impact the quality of the food and thereby the animal and the experiments. We know, for example, that the total isoflavones of soybeans vary within variety, locations and, over time, even when grown in the same location [101]. To avoid uncontrolled variation in models, the same batch of diet should be used throughout a study, and if it is a long-term study, the diet should be one lot (or mixed at the beginning of the study) and should be frozen and thawed for each feeding. Diet past its expiration date should never be used. Not only is degradation of the nutritional value of the diet a concern, but mould or bacterial overgrowth might also occur. The diet should be stored according to manufacturer’s recommendations, and a pest control programme should be in place to avoid compromise of diet by infectious agents carried by pests.
The composition of the major nutritional ingredients, carbohydrates, fat and proteins may have significant impact on animal studies. Prebiotic properties of dietary fermentable oligo-, di-, mono-saccharides and polyols (FODMAPs) are known to have a profound influence on the microbiota [102]. Fatty acids play an important role in inflammatory cascades and oxidative stress, and the source of fatty acids in the diet has immunomodulating effects [103]. Antioxidants are added to the food to avoid nutritional degradation and to avoid rancidity of the diet. The level of fatty acids in a diet influences the need for antioxidants, and fatty diet in general has shorter lifetime and must be stored under more strict conditions. The source of the protein may also impact nutritional content. In general, animal protein sources supply essential amino acids; however a combination of plant protein sources can also supply all essential amino acids. In 2005, Mattson et al. [104] demonstrated that the source of protein and fat in a diet affected the mean arterial pressure in salt-sensitive rats. Food sources, fatty acid and amino acid composition, and interaction with genetic predisposition to develop certain diseases, is an important field of research to better understand mechanism of metabolic diseases like diabetes [105].
For some animal models, food must be sterilised to be sure it is free of infectious agents. Heat sterilising via autoclave is one option. However, autoclaving has a detrimental effect on nutritional content, and only diets formulated to withstand this loss of nutrients should be used if autoclaving is necessary. Also, though autoclaving kills vegetative microbes and spores, it does not necessarily degrade or inactivate heat-stable bacteria toxins or products [106], so the same strict quality requirements for dietary sources still apply. Irradiation, usually via large-scale cobalt 60 gamma rays or electronic beam (E-beam) sources, can be used as an alternative to sterilise the diet when the risk of microbial contamination is an issue. Irradiation has a less negative impact on nutritional value [107].
Ad libitum feeding is the most common feeding regime in rodents; however as opportunities for activity in the rodent cage are low, there is a risk of overfeeding and obesity with subsequent welfare issues and obesity-related complications [108]. Food restriction should therefore be considered for long-term studies [109].
3.2.1 Phytoestrogens
Human clinical and nutritional interest in phytoestrogens during the last decade of the twentieth century led to publications illustrating that natural phytoestrogens present in commercial rodent diets could interfere with some research [110,111,112,113]. This is particularly true of steroid research due to the nature of these molecules. Coumestrol is the major phytoestrogen in alfalfa, which is a component of some commercial rodent diets. This phytoestrogen, which binds to the oestrogen receptor (ER), has been shown to alter the reproductive development of rats [110,111,112]. Isoflavone phytoestrogens, which are present in soy, are structurally similar to 17β-oestradiol and, thus, may bind to oestrogen receptors and have both oestrogenic and antiestrogenic activities [114], leading to induced alterations of normal physiological processes that may interfere with the research question being addressed. Alfalfa and soya are the major natural ingredients responsible for the isoflavone content of the current rodent diets. However, some laboratories have recently found no evidence of a soya-based diet influencing the results of behavioural, reproductive or welfare parameters in C57BL/6NCrl mice [115]. This group also found less despair behaviour in the forced swim test in the soy-free group and sexually dimorphic cognitive behaviour with the soy-containing standard diet [115]. Nevertheless, we know that the phytoestrogens present in the diet can influence oestrogenic studies, toxicology programs and carcinogenic studies [110,111,112,113, 116]. We also know that the total isoflavone content in soya varies with the variety and the location where it is grown [101, 117].
Rodent feed manufacturers offer not only phytoestrogen-free diets but also grain-based diets, free of any animal origin protein. This movement, towards an animal protein-free rodent diet, had its origins in the public health crisis related to “mad cow” disease, bovine spongiform encephalopathy (BSE), which was linked to a fatal brain disease in humans called variant Creutzfeldt-Jakob disease (vCJD) in the 1990s [118, 119]. Fishmeal has also been linked with confounding results in chemical toxicity and carcinogenicity studies conducted for the National Toxicology Program (NTP) due to nitrosamines and heavy metal content [120].
For carnivorous species, replacement of animal origin ingredients by plant-based ingredients can cause pathological conditions like soybean-induced enteritis in Atlantic salmon [121, 122].
A cautionary tale on how diet can affect experimental outcomes is illustrated by the Dahl salt-sensitive inbred rat (SS/Jr). In 2016, Margaret Zimmerman and Sarah Lindsey raised awareness over recurrent inconsistencies of the SS/Jr [123,124,125]. Jane Reckelhoff’s group found inconsistencies when testing the same strain from the same vendor (SS/JrHsd) but with different diets in different years [124, 126]. In 2010, the SS/JrHsd rats were given purified AIN-76A diet (American Institute of Nutrition formulated purified diet) [127, 128], which used refined ingredients (casein, DL-methionine, sucrose, corn starch, corn oil, cellulose, mineral mix AIN-76 (170915), vitamin mix AIN-76A (40077), choline bitartrate and ethoxyquin and antioxidant) and does not contain alfalfa, whereas, in 2016, SS/JrHsd rats were given Teklad 7034, a fixed formula diet with a different nutrient composition that does not used refined ingredients and contains alfalfa and soybean, two known sources of phytoestrogens. Thus, as illustrated by the Dalmasso and collaborators, the differences between a purified diet and a natural ingredient diet may have accounted for the variability seen in the model. Or, as pointed out by Zimmerman and Lindsey, through a revision of their own experiments with the model, other uncontrolled factors may have accounted for these differences. In any case, this shows how a well-characterised model that has worked consistently over decades can start producing inconsistent data, which may lead to wrong conclusions and thus lead scientists on equivocal pathways.
The diet provided to experimental animals can also affect the microbiota, but for that, we refer the reader to chapter “Microbiology and Microbiome” on microbiology and microbiome.
3.3 Water
As long as a diet of natural ingredients is used, normal municipality drinking water from the tap may be suitable for research animals. However, for immunocompromised animals or in studies where strict dietary control is crucial, other sources may be better, and for fish, water treatment is necessary. Autoclaving water will kill microbes and reduce the risk of water-born infections in immunocompromised animals. However, autoclaving does not take away chemical substances or heat-stable toxins. Chlorification or acidification is also used to reduce microbiological growth in the drinking water. However, this can impact the taste of the water and water consumption. Depending on the filter technology, mechanical filtration takes out organic material, and charcoal filtration may take out smell, taste and some chemical substances. In reverse osmosis (RO) treatments of water, a partially permeable membrane is used to remove ions, unwanted molecules and larger particles from the raw water source. Regardless of the source, water should be regularly monitored at the facility level for contaminants, both organic and inorganic [129].
Water is usually provided ad libitum in water bottle or in a centralised watering system. If animals are offered fruit, gels or other water-containing diet in addition to kibble or pellets, this might influence their overall drinking water intake. If test substances are provided per os in the drinking water, it might be necessary to control the water intake to be sure animals get the exact amount of test substance.
3.4 Noise and Vibration
The animal facility acoustic environment was mostly ignored by managers, regulators, scientists, architects, engineers and designers in the past. As recent research has shown, personnel involved with animal facilities recognised that control of environmental factors, such as noise, in animal facilities is important to ensure consistent responses to experimental procedures [130]. It is also well known that construction noise (range 70–90 dBA) affects different reproductive parameters in mice [131]. In Europe, the Commission Recommendation of 18 June 2007 on guidelines for the accommodation and care of animals used for experimental and other scientific purposes (2007/526/EC), under “The Environment and Its Control” section, in its point 2.5 recognises that “noise can be a disturbing factor for animals” and gives vague recommendations on what should be an ideal acoustic environment and what should be avoided, both in terms of actual noise and design of the facilities [28]. By contrast, noise and vibration are dealt with by the Guide for the Care and Use of Laboratory Animals extensively in Chapter 3, “Environment, Housing and Management”, for terrestrial and aquatic animals, and in Chapter 5, “Physical Plant” [39]. Measurements are routinely taken to monitor ventilation, temperature, humidity and lighting and adequate ranges given for them. However, the acoustic environment is often given relatively little consideration mainly due to the difficulty in consistent noise measurements, the differences in audible frequencies to different species and the unknown limits where there is an effect on the normal physiology of animals.
Some groups advocate for a more detailed definitions of what is a safe acoustic environment for animals in research. The first question we need to address is: what is sound? Sound, in physics terms, is a vibration that propagates through a transmission medium, like air or water, as a pressure wave that is audible. At the reception point, i.e. the ears in humans and other vertebrates, pressure and time are the two elements that will describe every sound we hear. We will see when we read about noise that most of the research refers to sound pressure level. Sound pressure will be the deviation from the atmospheric pressure caused by a sound wave. Sound pressure level is the ratio of the absolute sound pressure and a reference level, usually the threshold of hearing, or the lowest intensity of sound that can be heard by most people. The decibel (dB) is commonly used as the unit for sound pressure level, being the ratio of sound pressure on a logarithmic scale.
Different cardiovascular parameters have already been shown to be affected by noise; environmental noise causes a number of changes in laboratory animals: increase of blood pressure in cats, rats, rhesus monkeys and macaque monkeys [132,133,134,135,136,137]; increase in heart rate in desert mule deer and rats [138]; increase in vasoconstriction in rats [139,140,141]; increase in respiratory rates and adrenocorticotropin hormone in cats [142]; hypertension [143, 144]; cardiac hypertrophy [144]; changes in electrolyte metabolism [145]; reduced body weight [146,147,148]; increased adrenal weight [144]; altered tumour resistance and immune response [149]; slower wound healing [148]; changes in oestrous cycles, increased weight of uterus and ovaries, spontaneous lactation, decreased fertility and termination of pregnancy [150]; and embryonic abnormalities [144]. Mice stressed by sound during pregnancy also produce offspring with poor learning ability [151]. Some researchers in contrast have found that noise induces no change in blood pressure in the rat [152]. Noise has also been associated with a change in sleeping patterns in humans [153]. Sanchez-Morgado and collaborators have also studied the effects of construction noise on different biochemical parameters, heart rate and arterial blood pressure in mice and found that there is an increase in systolic and diastolic blood pressure; in the pulse in males more than in females; in cholesterol, triglycerides and LDL in males; and that males are more affected by noise than females [154]. Researchers working with mice in noisy environments should be aware of variations that could mask valid experimental results.
Water is an excellent medium for transfer of acoustic energy without major attenuation. For aquatic animals the perception of “sound” is not limited to “hearing” by ears, but also – depending on the species – involves the lateral line and the gas bladder, making “hearing” more complex involving near-field and far-field signals. The ability to discriminate sounds of interest from background noise also varies between species so that fish classified as auditory specialists are of greater risk of suffering from hearing loss than auditory generalists [155].
Vibration is the periodic back-and-forth motion of the particles of an elastic body or medium, commonly resulting when almost any physical system is displaced from its equilibrium condition and allowed to respond to the forces that tend to restore equilibrium. Thus, sound is generated by vibrating structures, and sound can also cause vibration of structures. We can see that sound and vibration are intimately related. Nevertheless, contrary to sound, we have little data on vibration and its effects on laboratory animals [156].
The reader is referred to other more in-depth reviews on the hearing of laboratory animals like the one published in 2005 by John Turner et al. [157].
3.5 Temperature
The thermoneutral zone (TNZ) is defined as the range of ambient temperature at which temperature regulation is achieved only by control of sensible heat loss, i.e. without regulatory changes in metabolic heat production (rate of transformation of chemical energy into heat in an organism) or evaporative heat loss. The TNZ will therefore be different when insulation, posture and basal metabolic rate (BMR) vary. In mice, the thermoneutral zone is approximately between 29.6 °C and 30.5 °C, although this varies with the strain, age, sex and activity level. The preferred temperature – which can be defined as the temperature where animals will choose to stay when a range of temperatures is given as a choice – varies in mice depending on behaviour, strain, time of the day, age and sex [158,159,160,161,162]. Generally, it has been found that the 26 °C to 29 °C range is preferred for sleep by both sexes in all mouse strains [160]. Rodents adjust to changes in temperature by adjusting metabolism, so temperature control is extremely important as it has a major impact on experimental results [163]. The European Guidelines for the Accommodation and Care of Animals Used for Experimental and other Scientific Purposes defines standard temperatures for animal housing in Europe [28]. These ranges are measured at the room level and are typically lower than those preferred by the animals. Nevertheless, these temperature ranges are a compromise between the animals’ needs and what employees can tolerate as their working environment. Thus, there is a thermal stress associated with the current recommended temperature ranges [28, 39] for many animals. Fischer and collaborators [164] have found that mouse metabolism more closely resembles human metabolism at the thermoneutral zone. This is something important to consider for scientists working in human metabolism and using the mouse as a model. As they established, “at any temperature below thermoneutrality, mice metabolism exceeds the human equivalent: Mice under standard conditions display energy expenditure 3.1 times basal metabolism”, whereas “humans usually display average metabolic rates of about 1.6 times basal metabolic rate” [164].
The term thermoneutral zone does not apply to ectotherms [165]. For poikilothermic animals the temperature will have an intrinsic impact on growth, development and behaviour.
3.6 Humidity
Recommendations for humidity levels of rodent housing rooms can be found in the European Guidelines for the Accommodation and Care of Animals Used for Experimental and Other Scientific Purposes [28]. Values for rodents are in general higher than office areas so humidification of the ventilated air is usually necessary. Fluctuation of humidity is stressful for the animals and should therefore be kept within a defined range. Too low relative humidity has been discussed as a cause of the condition “ringtail” in rodents [166], respiratory issue and reproductive problems (pup eating). Too high humidity may cause hygienic problems with microbial growth. Different housing systems can affect the humidity depending on the ventilation range and quality of the ventilated air.
4 Final Word
In any animal research effort, many factors may come into play. Control and standardisation of all variables are impossible on either the scientist or the animal side. Uncontrolled variables will have an influence on the final data, and the acknowledgement of this fact can open new avenues of inquiry or aid other scientists in answering their questions. It is, therefore, important to provide all the information we have in our published manuscripts so others can, at least, try to reproduce our conditions when attempting to reproduce results or explain conflicting results in their facility. Ideally, scientists would only publish robust data with external validity, but we understand this is an incremental goal that requires many small steps and changes to the scientific endeavour. After reading this chapter, we hope scientists will be more aware of how the environment and the animal may affect their research outcome. If the reader stops to think carefully about their next experiment and these influences, we will have attained our goal in writing this chapter. There are other chapters within this book that will help both seasoned researchers and early-career scientists tackle different issues related to animal-based research. Therefore, we encourage the reader to peruse those chapters and obtain a much better overview of these factors.
References
Richter SH, Garner JP, Würbel H. Environmental standardization: cure or cause of poor reproducibility in animal experiments? Nat Methods. 2009;6:257–61. https://doi.org/10.1038/nmeth.1312.
Garner JP, Gaskill BN, Weber EM, et al. Introducing therioepistemology: the study of how knowledge is gained from animal research. Lab Anim (NY). 2017;46:103–13. 2017/03/23. https://doi.org/10.1038/laban.1224.
Bubier JA, Philip VM, Quince C, et al. A microbe associated with sleep revealed by a novel systems genetic analysis of the microbiome in collaborative cross mice. Genetics. 2020;214:719–33. 2020/01/04. https://doi.org/10.1534/genetics.119.303013.
Deloris Alexander A, Orcutt RP, Henry JC, et al. Quantitative PCR assays for mouse enteric flora reveal strain-dependent differences in composition that are influenced by the microenvironment. Mamm Genome. 2006;17:1093–104. 2006/11/09. https://doi.org/10.1007/s00335-006-0063-1.
Goodrich JK, Waters JL, Poole AC, et al. Human genetics shape the gut microbiome. Cell. 2014;159:789–99. 2014/11/25. https://doi.org/10.1016/j.cell.2014.09.053.
Jacobs JP, Braun J. Immune and genetic gardening of the intestinal microbiome. FEBS Lett. 2014;588:4102–11. 2014/03/13. https://doi.org/10.1016/j.febslet.2014.02.052.
Richter SH, Garner JP, Auer C, et al. Systematic variation improves reproducibility of animal experiments. Nat Methods. 2010;7:167–8. https://doi.org/10.1038/nmeth0310-167.
Festing MF. The choice of animals in toxicological screening: inbred strains and the factorial design of experiment. Acta Zool Pathol Antverp. 1980;117–31. 1980/10/01.
Beery AK, Zucker I. Sex bias in neuroscience and biomedical research. Neurosci Biobehav Rev. 2011;35:565–72. https://doi.org/10.1016/j.neubiorev.2010.07.002.
Beery AK. Inclusion of females does not increase variability in rodent research studies this review comes from a themed issue on sex and gender. Curr Opin Behav Sci. 2018;23:143–9. https://doi.org/10.1016/j.cobeha.2018.06.016.
Clayton JA, Collins FS. Policy: NIH to balance sex in cell and animal studies. Nature. 2014;509:282–3. https://doi.org/10.1038/509282a.
NOT-OD-15-102. Consideration of sex as a biological variable in NIH-funded research. 2015.
Richardson SS, Reiches M, Shattuck-Heidorn H, et al. Opinion: focus on preclinical sex differences will not address women’s and men’s health disparities. Proc Natl Acad Sci. 2015;112:13419–20. https://doi.org/10.1073/PNAS.1516958112.
Klein SL, Schiebinger L, Stefanick ML, et al. Opinion: sex inclusion in basic research drives discovery. Proc Natl Acad Sci U S A. 2015;112:5257–8. https://doi.org/10.1073/pnas.1502843112.
Tannenbaum C, Schwarz JM, Clayton JA, et al. Evaluating sex as a biological variable in preclinical research: the devil in the details. Biol Sex Differ. 2016;7:13. https://doi.org/10.1186/s13293-016-0066-x.
Sorge RE, LaCroix-Fralish ML, Tuttle AH, et al. Spinal cord toll-like receptor 4 mediates inflammatory and neuropathic hypersensitivity in male but not female mice. J Neurosci Off J Soc Neurosci. 2011;31:15450–4. https://doi.org/10.1523/JNEUROSCI.3859-11.2011.
Sorge RE, Mapplebeck JCS, Rosen S, et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat Neurosci. 2015;18:1081–3. https://doi.org/10.1038/nn.4053.
Doyle HH, Eidson LN, Sinkiewicz DM, et al. Sex differences in microglia activity within the periaqueductal gray of the rat: a potential mechanism driving the dimorphic effects of morphine. J Neurosci Off J Soc Neurosci. 2017;37:3202–14. https://doi.org/10.1523/JNEUROSCI.2906-16.2017.
Tronson NC. Focus on females: a less biased approach for studying strategies and mechanisms of memory this review comes from a themed issue on sex and gender ScienceDirect. Curr Opin Behav Sci. 2018;23:92–7. https://doi.org/10.1016/j.cobeha.2018.04.005.
Keiser AA, Turnbull LM, Darian MA, et al. Sex differences in context fear generalization and recruitment of Hippocampus and amygdala during retrieval. Neuropsychopharmacology. 2017;42:397–407. https://doi.org/10.1038/npp.2016.174.
Keller KM, Howlett SE. Sex differences in the biology and pathology of the aging heart. Can J Cardiol. 2016;32:1065–73. https://doi.org/10.1016/J.CJCA.2016.03.017.
Mazzaccara C, Labruna G, Cito G, et al. Age-related reference intervals of the Main biochemical and hematological parameters in C57BL/6J, 129SV/EV and C3H/HeJ mouse strains. PLoS One. 2008;3:e3772. https://doi.org/10.1371/journal.pone.0003772.
Berson DM, Dunn FA, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295:1070–3. 2002/02/09. https://doi.org/10.1126/science.1067262.
Hanifin JP, Brainard GC. Photoreception for circadian, neuroendocrine, and neurobehavioral regulation. J Physiol Anthropol. 2007;26:87–94. 2007/04/17. https://doi.org/10.2114/jpa2.26.87.
Hastings MH, Maywood ES, Brancaccio M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat Rev Neurosci. 2018;19:453–69. https://doi.org/10.1038/s41583-018-0026-z.
Furudate S, Takahashi A, Takagi M, et al. Delayed persistent estrus induced by continuous lighting after inadequate acclimation in rats. Exp Anim. 2005;54:93–5. 2005/02/24. https://doi.org/10.1538/expanim.54.93.
Bedrosian TA, Vaughn CA, Galan A, et al. Nocturnal light exposure impairs affective responses in a wavelength-dependent manner. J Neurosci. 2013;33:13081–7. 2013/08/09. https://doi.org/10.1523/JNEUROSCI.5734-12.2013.
European C. Commission recommendation of 18 June 2007 on guidelines for the accommodation and care of animals used for experimental and other scientific purposes (2007/526/EC). 2007. Brussels.
Joyner CP, Myrick LC, Crossland JP, et al. Deer mice as laboratory animals. ILAR J. 1998;39:322–30. 2001/06/15. https://doi.org/10.1093/ilar.39.4.322.
Banerjee S, Chaturvedi CM. Testicular atrophy and reproductive quiescence in photorefractory and scotosensitive quail: involvement of hypothalamic deep brain photoreceptors and GnRH-GnIH system. J Photochem Photobiol B. 2017;175:254–68. 2017/09/20. https://doi.org/10.1016/j.jphotobiol.2017.09.005.
Robinson-Junker A, O’Hara B, Durkes A, et al. Sleeping through anything: the effects of unpredictable disruptions on mouse sleep, healing, and affect. PLoS One. 2019;14:e0210620. 2019/02/01. https://doi.org/10.1371/journal.pone.0210620.
Hawkins P, Golledge HDR. The 9 to 5 rodent – time for change? Scientific and animal welfare implications of circadian and light effects on laboratory mice and rats. J Neurosci Methods. 2018;300:20–5. 2017/05/16. https://doi.org/10.1016/j.jneumeth.2017.05.014.
European Parliament and Council of Europe. Council directive 86/609/EEC of 24 November 1986 on the approximation of laws, regulations and administrative provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes. Strasbourg: Official Journal of the European Communities, 1986, p. 28.
Niklaus S, Albertini S, Schnitzer TK, et al. Challenging a myth and misconception: red-light vision in rats. Animals (Basel). 2020;10. 2020/03/07. https://doi.org/10.3390/ani10030422.
Rocha FA, Gomes BD, Silveira LC, et al. Spectral sensitivity measured with electroretinogram using a constant response method. PLoS One. 2016;11:e0147318. 2016/01/23. https://doi.org/10.1371/journal.pone.0147318.
Coomans CP, van den Berg SA, Lucassen EA, et al. The suprachiasmatic nucleus controls circadian energy metabolism and hepatic insulin sensitivity. Diabetes. 2013;62:1102–8. 2013/01/01. https://doi.org/10.2337/db12-0507.
Qian J, Block GD, Colwell CS, et al. Consequences of exposure to light at night on the pancreatic islet circadian clock and function in rats. Diabetes. 2013;62:3469–78. https://doi.org/10.2337/db12-1543.
Gale JE, Cox HI, Qian J, et al. Disruption of circadian rhythms accelerates development of diabetes through pancreatic beta-cell loss and dysfunction. J Biol Rhythms. 2011;26:423–33. 2011/09/17. https://doi.org/10.1177/0748730411416341.
The National Research C. Guide for the care and use of laboratory animals. Washington, DC: National Academies Press; 2011. p. 246.
Peirson SN, Brown LA, Pothecary CA, et al. Light and the laboratory mouse. J Neurosci Methods. 2018;300:26–36. 2017/04/18. https://doi.org/10.1016/j.jneumeth.2017.04.007.
Dauchy RT, Blask DE, Hoffman AE, et al. Influence of daytime LED light exposure on circadian regulatory dynamics of metabolism and physiology in mice. Com Med. 2019;69:350–73. https://doi.org/10.30802/AALAS-CM-19-000001.
Dauchy RT, Wren-Dail MA, Hoffman AE, et al. Effects of daytime exposure to light from blue-enriched light-emitting diodes on the nighttime melatonin amplitude and circadian regulation of rodent metabolism and physiology. Comp Med. 2016;66:373–83. 2016/10/26
Lucas RJ, Brainard GC, Berson DM, et al. Report on the first international workshop on circadian and neurophysiological photometry, 2013. The Commission Internationale de l’Eclairage; 2015.
Lucas RJ, Peirson SN, Berson DM, et al. Measuring and using light in the melanopsin age. Trends Neurosci. 2014;37:1–9. 2013/11/25. https://doi.org/10.1016/j.tins.2013.10.004.
Yuan R, Tsaih S-W, Petkova SB, et al. Aging in inbred strains of mice: study design and interim report on median lifespans and circulating IGF1 levels. Aging Cell. 2009;8:277–87. https://doi.org/10.1111/j.1474-9726.2009.00478.x.
Holladay SD, Blaylock BL. The mouse as a model for developmental immunotoxicology. Hum Exp Toxicol. 2002;21:525–31. https://doi.org/10.1191/0960327102ht292oa.
Rieger MA, Schroeder T. Hematopoiesis. Cold Spring Harb Perspect Biol. 2012;4. https://doi.org/10.1101/CSHPERSPECT.A008250.
Schmitt CE, Lizama CO, Zovein AC. From transplantation to transgenics: mouse models of developmental hematopoiesis. Exp Hematol. 2014;42:707–16. https://doi.org/10.1016/J.EXPHEM.2014.06.008.
Loeffler M, Pantel K. A mathematical model of erythropoiesis suggests an altered plasma volume control as cause for anemia in aged mice. Exp Gerontol. 1990;25:483–95.
Boggs DR, Patrene KD. Hematopoiesis and aging III: Anemia and a blunted erythropoietic response to hemorrhage in aged mice. Am J Hematol. 1985;19:327–38. https://doi.org/10.1002/ajh.2830190403.
Sorzano COS, Sánchez-Morgado JM. Normal haematological reference intervals for different inbred strains at 9 weeks of age. Dublin: Trinity College Dublin; 2020.
Fox JG. The mouse in biomedical research. Volume III, Normative biology, husbandry and models. 2nd ed. Amsterdam/Boston: Elsevier; 2007.
Walters BJ, Zuo J. Postnatal development, maturation and aging in the mouse cochlea and their effects on hair cell regeneration. Hear Res. 2013;297:68–83. https://doi.org/10.1016/j.heares.2012.11.009.
Johnson KR, Zheng QY, Erway LC. A major gene affecting age-related hearing loss is common to at least ten inbred strains of mice. Genomics. 2000;70:171–80. 2000/12/09. https://doi.org/10.1006/geno.2000.6377.
Kane KL, Longo-Guess CM, Gagnon LH, et al. Genetic background effects on age-related hearing loss associated with Cdh23 variants in mice. Hear Res. 2012;283:80–8. 2011/12/06. https://doi.org/10.1016/j.heares.2011.11.007.
Hathway GJ, Koch S, Low L, et al. The changing balance of brainstem-spinal cord modulation of pain processing over the first weeks of rat postnatal life. J Physiol. 2009;587:2927–35. https://doi.org/10.1113/jphysiol.2008.168013.
Kwok CHT, Devonshire IM, Imraish A, et al. Age-dependent plasticity in endocannabinoid modulation of pain processing through postnatal development. Pain. 2017;158:2222–32. https://doi.org/10.1097/j.pain.0000000000001027.
Schwaller F, Kanellopoulos AH, Fitzgerald M. The developmental emergence of differential brainstem serotonergic control of the sensory spinal cord. Sci Rep. 2017;7. https://doi.org/10.1038/S41598-017-02509-2.
Stoyek MR, Rog-Zielinska EA, Quinn TA. Age-associated changes in electrical function of the zebrafish heart. Prog Biophys Mol Biol. 2018;138:91–104. https://doi.org/10.1016/j.pbiomolbio.2018.07.014.
Mohrhardt J, Nagel M, Fleck D, et al. Signal detection and coding in the accessory olfactory system. Chem Senses. 2018;43:667–95. https://doi.org/10.1093/chemse/bjy061.
Wyatt TD. Pheromones. Curr Biol. 2017;27:R739–43. https://doi.org/10.1016/j.cub.2017.06.039.
Roberts SA, Prescott MC, Davidson AJ, et al. Individual odour signatures that mice learn are shaped by involatile major urinary proteins (MUPs). BMC Biol. 2018;16:48. 2018/04/29. https://doi.org/10.1186/s12915-018-0512-9.
Nodari F, Hsu F-F, Fu X, et al. Sulfated steroids as natural ligands of mouse pheromone-sensing neurons. J Neurosci. 2008;28:6407–18. https://doi.org/10.1523/jneurosci.1425-08.2008.
Stockley P, Bottell L, Hurst JL. Wake up and smell the conflict: odour signals in female competition. Philos Trans R Soc Lond B Biol Sci. 2013;368:20130082. 2013/10/30. https://doi.org/10.1098/rstb.2013.0082.
Van Der Lee S, Boot LM. Spontaneous pseudopregnancy in mice. II. Acta Physiol Pharmacol Neerl. 1956;5:213–5. 1956/12/01
Van Der Lee S, Boot LM. Spontaneous pseudopregnancy in mice. Acta Physiol Pharmacol Neerl. 1955;4:442–4. 1955/11/01
Ma W, Miao Z, Novotny MV. Role of the adrenal gland and adrenal-mediated chemosignals in suppression of estrus in the house mouse: the lee-boot effect revisited. Biol Reprod. 1998;59:1317–20. 1998/11/26. https://doi.org/10.1095/biolreprod59.6.1317.
Koyama S. Primer effects by conspecific odors in house mice: a new perspective in the study of primer effects on reproductive activities. Horm Behav. 2004;46:303–10. https://doi.org/10.1016/j.yhbeh.2004.03.002.
Whitten WK. Modification of the oestrous cycle of the mouse by external stimuli associated with the male. J Endocrinol. 1956;13:399–404. 1956/07/01. https://doi.org/10.1677/joe.0.0130399.
Whitten WK. Modification of the oestrous cycle of the mouse by external stimuli associated with the male; changes in the oestrous cycle determined by vaginal smears. J Endocrinol. 1958;17:307–13. 1958/09/01. https://doi.org/10.1677/joe.0.0170307.
Bruce HM. An exteroceptive block to pregnancy in the mouse. Nature. 1959;184:105. 1959/07/11. https://doi.org/10.1038/184105a0.
Parkes AS, Bruce HM. Olfactory stimuli in mammalian reproduction. Science. 1961;134:1049–54. 1961/10/13. https://doi.org/10.1126/science.134.3485.1049.
Vandenbergh JG. Male odor accelerates female sexual maturation in mice. Endocrinology. 1969;84:658–60. 1969/03/01. https://doi.org/10.1210/endo-84-3-658.
Szymanski LA, Keller M. Activation of the olfactory system in response to male odors in female prepubertal mice. Behav Brain Res. 2014;271:30–8. https://doi.org/10.1016/j.bbr.2014.05.051.
Bacchini A, Gaetani E, Cavaggioni A. Pheromone binding proteins of the mouse, Mus musculus. Experientia. 1992;48:419–21. 1992/04/15. https://doi.org/10.1007/bf01923448.
Koyama S, Kamimura S. Effects of vomeronasal organ removal on the sperm motility in male mice. Zoolog Sci. 2003;20:1355–8. 2003/11/19. https://doi.org/10.2108/zsj.20.1355.
Hurst JL, West RS. Taming anxiety in laboratory mice. Nat Methods. 2010;7:825–6. 2010/09/14. https://doi.org/10.1038/nmeth.1500.
Gouveia K, Hurst JL. Reducing mouse anxiety during handling: effect of experience with handling tunnels. PLoS One. 2013;8:e66401. 2013/07/11. https://doi.org/10.1371/journal.pone.0066401.
Gouveia K, Hurst JL. Optimising reliability of mouse performance in behavioural testing: the major role of non-aversive handling. Sci Rep. 2017;7:44999. 2017/03/23. https://doi.org/10.1038/srep44999.
Gouveia K, Hurst JL. Improving the practicality of using non-aversive handling methods to reduce background stress and anxiety in laboratory mice. Sci Rep. 2019;9:20305. https://doi.org/10.1038/s41598-019-56860-7.
Henderson LJ, Smulders TV, Roughan JV. Identifying obstacles preventing the uptake of tunnel handling methods for laboratory mice: An international thematic survey. PLoS One. 2020;15:e0231454. 2020/04/15. https://doi.org/10.1371/journal.pone.0231454.
Whittaker AL, Howarth GS, Hickman DL. Effects of space allocation and housing density on measures of wellbeing in laboratory mice: a review. Lab Anim. 2012;46:3–13. https://doi.org/10.1258/la.2011.011049.
Markham JA, Greenough WT. Experience-driven brain plasticity: beyond the synapse. Neuron Glia Biol. 2004;1:351–63. 2006/08/22. https://doi.org/10.1017/s1740925x05000219.
European Parliament and Council of Europe. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Strasbourg: Official Journal of the European Union; 2010.
Wurbel H. Ideal homes? Housing effects on rodent brain and behaviour. Trends Neurosci. 2001;24:207–11. 2001/03/16. https://doi.org/10.1016/s0166-2236(00)01718-5.
Langston RF, Ainge JA, Couey JJ, et al. Development of the spatial representation system in the rat. Science. 2010;328:1576–80. 2010/06/19. https://doi.org/10.1126/science.1188210.
André V, Gau C, Scheideler A, et al. Laboratory mouse housing conditions can be improved using common environmental enrichment without compromising data. PLoS Biol. 2018;16:e2005019. https://doi.org/10.1371/journal.pbio.2005019.
Macri S, Ceci C, Altabella L, et al. The Directive 2010/63/EU on animal experimentation may skew the conclusions of pharmacological and behavioural studies. Sci Rep. 2013;3:2380. 2013/08/09. https://doi.org/10.1038/srep02380.
Atwood BK, Huffman J, Straiker A, et al. JWH018, a common constituent of ‘Spice’ herbal blends, is a potent and efficacious cannabinoid CB receptor agonist. Br J Pharmacol. 2010;160:585–93. 2010/01/27. https://doi.org/10.1111/j.1476-5381.2009.00582.x.
Motulsky H. Intuitive biostatistics. A nonmathematical guide to statistical thinking. 4th ed. New York: Oxford University Press; 2018. p. 568.
Gaskill BN, Garner JP. Stressed out: providing laboratory animals with behavioral control to reduce the physiological effects of stress. Lab Anim (NY). 2017;46:142–5. 2017/03/23. https://doi.org/10.1038/laban.1218.
Greenman DL, Bryant P, Kodell RL, et al. Relationship of mouse body weight and food consumption/wastage to cage shelf level. Lab Anim Sci. 1983;33:555–8. 1983/12/01
Greenman DL, Kodell RL, Sheldon WG. Association between cage shelf level and spontaneous and induced neoplasms in mice. J Natl Cancer Inst. 1984;73:107–13. 1984/07/01
Ader DN, Johnson SB, Huang SW, et al. Group size, cage shelf level, and emotionality in non-obese diabetic mice: impact on onset and incidence of IDDM. Psychosom Med. 1991;53:313–21. 1991/05/01. https://doi.org/10.1097/00006842-199105000-00005.
Okva K, Nevalainen T, Pokk P. The effect of cage shelf on the behaviour of male C57BL/6 and BALB/c mice in the elevated plus maze test. Lab Anim. 2013;47:220–2. 2013/06/14. https://doi.org/10.1177/0023677213489280.
Young SS. Are there local room effects on hepatic tumors in male mice? An examination of the NTP eugenol study. Fundam Appl Toxicol. 1987;8:1–4. 1987/01/01
Young SS. A blind reanalysis of a random subset of NCI bioassay studies: agreement between rats and mice. Fundam Appl Toxicol. 1989;12:232–41. 1989/02/01
Kozul CD, Nomikos AP, Hampton TH, et al. Laboratory diet profoundly alters gene expression and confounds genomic analysis in mouse liver and lung. Chem Biol Interact. 2008;173:129–40. https://doi.org/10.1016/J.CBI.2008.02.008.
Brown NM, Setchell KDR. Animal models impacted by phytoestrogens in commercial chow: implications for pathways influenced by hormones. Lab Investig. 2001;81:735–47. https://doi.org/10.1038/labinvest.3780282.
Carter RL, Lipman NS. Feed and bedding. In: Weichbrod RH, Thompson GAH, et al., editors. Management of animal care and use programs in research, education, and testing. Boca Raton: CRC Press; 2018. p. 639–54.
Eldridge AC. Determination of isoflavones in soybean flours, protein concentrates, and isolates. J Agric Food Chem. 1982;30:353–5. https://doi.org/10.1021/jf00110a035.
Tuck CJ, Caminero A, Jimenez Vargas NN, et al. The impact of dietary fermentable carbohydrates on a postinflammatory model of irritable bowel syndrome. Neurogastroenterol Motil. 2019;31:e13675. 2019/07/11. https://doi.org/10.1111/nmo.13675.
Li M, van Esch B, Wagenaar GTM, et al. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur J Pharmacol. 2018;831:52–9. 2018/05/12. https://doi.org/10.1016/j.ejphar.2018.05.003.
Mattson DL, Meister CJ, Marcelle ML. Dietary protein source determines the degree of hypertension and renal disease in the Dahl salt-sensitive rat. Hypertension. 2005;45:736–41. https://doi.org/10.1161/01.HYP.0000153318.74544.cc.
Svard J, Rost TH, Sommervoll CEN, et al. Absence of the proteoglycan decorin reduces glucose tolerance in overfed male mice. Sci Rep. 2019;9:4614. 2019/03/16. https://doi.org/10.1038/s41598-018-37501-x.
Hrncir T, Stepankova R, Kozakova H, et al. Gut microbiota and lipopolysaccharide content of the diet influence development of regulatory T cells: studies in germ-free mice. BMC Immunol. 2008;9:65. 2008/11/08. https://doi.org/10.1186/1471-2172-9-65.
Caulfield CD, Cassidy JP, Kelly JP. Effects of gamma irradiation and pasteurization on the nutritive composition of commercially available animal diets. J Am Assoc Lab Anim Sci. 2008;47:61–6. 2008/12/04
Martin B, Ji S, Maudsley S, et al. “Control” laboratory rodents are metabolically morbid: why it matters. Proc Natl Acad Sci U S A. 2010;107:6127–33. https://doi.org/10.1073/pnas.0912955107.
Hubert MF, Laroque P, Gillet JP, et al. The effects of diet, ad Libitum feeding, and moderate and severe dietary restriction on body weight, survival, clinical pathology parameters, and cause of death in control Sprague-Dawley rats. Toxicol Sci. 2000;58:195–207. 2000/10/29. https://doi.org/10.1093/toxsci/58.1.195.
Whitten PL, Lewis C, Russell E, et al. Potential adverse effects of phytoestrogens. J Nutr. 1995;125:771S–6S. https://doi.org/10.1093/jn/125.3_suppl.771s.
Whitten PL, Russell E, Naftolin F. Effects of a normal, human-concentration, phytoestrogen diet on rat uterine growth. Steroids. 1992;57:98–106. https://doi.org/10.1016/0039-128X(92)90066-I.
Whitten PL, Naftolin F. Effects of a phytoestrogen diet on estrogen-dependent reproductive processes in immature female rats. Steroids. 1992;57:56–61. https://doi.org/10.1016/0039-128X(92)90029-9.
Ashby J, Tinwell H, Soames A, et al. Induction of hyperplasia and increased DNA content in the uterus of immature rats exposed to coumestrol. Environ Health Perspect. 1999;107:819–22. https://doi.org/10.1289/ehp.99107819.
Dixon RA. Phytoestrogens. Annu Rev Plant Biol. 2004;55:225–61. https://doi.org/10.1146/annurev.arplant.55.031903.141729.
Mallien AS, Soukup ST, Pfeiffer N, et al. Effects of soy in laboratory rodent diets on the basal, affective, and cognitive behavior of C57BL/6 mice. J Am Assoc Lab Anim Sci. 2019;58:532–41. https://doi.org/10.30802/AALAS-JAALAS-18-000129.
Thigpen JE, Setchell KD, Goelz MF, et al. The phytoestrogen content of rodent diets. Environ Health Perspect. 1999;107:A182–3. https://doi.org/10.1289/ehp.107-1566530.
Eldridge AC, Kwolek WF. Soybean isoflavones: effect of environment and variety on composition. J Agric Food Chem. 1983;31:394–6. https://doi.org/10.1021/jf00116a052.
Collinge J, Sidle KCL, Meads J, et al. Molecular analysis of prion strain variation and the aetiology of ‘new variant’ CJD. Nature. 1996;383:685–90. https://doi.org/10.1038/383685a0.
Will RG, Ironside JW, Zeidler M, et al. A new variant of Creutzfeldt-Jakob disease in the UK. Lancet. 1996;347:921–5. https://doi.org/10.1016/S0140-6736(96)91412-9.
Rao GN, Knapka JJ. Contaminant and nutrient concentrations of natural ingredient rat and mouse diet used in chemical toxicology studies. Fundam Appl Toxicol. 1987;9:329–38. https://doi.org/10.1016/0272-0590(87)90055-8.
Knudsen D, Jutfelt F, Sundh H, et al. Dietary soya saponins increase gut permeability and play a key role in the onset of soyabean-induced enteritis in Atlantic salmon (Salmo salar L.). Br J Nutr. 2008;100:120–9. 2008/01/03. https://doi.org/10.1017/S0007114507886338.
Refstie S, Korsøen ØJ, Storebakken T, et al. Differing nutritional responses to dietary soybean meal in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Aquaculture. 2000;190:49–63. https://doi.org/10.1016/S0044-8486(00)00382-3.
Zimmerman MA, Lindsey SH. Inconsistent blood pressure phenotype in female Dahl salt-sensitive rats. Am J Physiol Renal Physiol. 2016;311:F1391–2. https://doi.org/10.1152/ajprenal.00454.2016.
Dalmasso C, Maranon R, Patil C, et al. 20-HETE and CYP4A2 ω-hydroxylase contribute to the elevated blood pressure in hyperandrogenemic female rats. Am J Physiol Renal Physiol. 2016;311:F71–7. https://doi.org/10.1152/ajprenal.00458.2015.
Gillis EE, Williams JM, Garrett MR, et al. The Dahl salt-sensitive rat is a spontaneous model of superimposed preeclampsia. Am J Phys Regul Integr Comp Phys. 2015;309:R62–70. https://doi.org/10.1152/ajpregu.00377.2014.
Sartori-Valinotti JC, Venegas-Pont MR, LaMarca BB, et al. Rosiglitazone reduces blood pressure in female Dahl salt-sensitive rats. Steroids. 2010;75:794–9. https://doi.org/10.1016/J.STEROIDS.2009.10.010.
Report of the American Institute of Nutrition Ad Hoc Committee on Standards for Nutritional Studies. J Nutr. 1977;107:1340–8. https://doi.org/10.1093/jn/107.7.1340.
Bieri JG. Second report of the ad hoc committee on standards for nutritional studies. J Nutr. 1980;110:1726. https://doi.org/10.1093/jn/110.8.1726.
Allen ED, Czarra EF, De Tolla L. Water quality and water delivery systems. In: Weichbrod RH, GAH T, Norton JN, editors. Management of animal care and use programs in research, education, and testing. Boca Raton: CRC Press; 2018. p. 655–73.
Baldwin AL, Schwartz GE, Hopp DH. Are investigators aware of environmental noise in animal facilities and that this noise may affect experimental data? J Am Assoc Lab Anim Sci. 2007;46:45–51. 2007/01/06
Rasmussen S, Glickman G, Norinsky R, et al. Construction noise decreases reproductive efficiency in mice. J Am Assoc Lab Anim Sci. 2009;48:363–70. 2009/08/06
Hudak WJ, Buckley JP. Production of hypertensive rats by experimental stress. J Pharm Sci. 1961;50:263–4. https://doi.org/10.1002/JPS.2600500321.
Buckley JP, Smookler HH. Cardiovascular and biochemical effects of chronic intermittent neurogenic stimulation. In: Welch BL, Welch AS, editors. Physiological effects of noise: based upon papers presented at an international symposium on the extra-auditory physiological effects of audible sound, held in Boston, Massachusetts, December 28–30, 1969, in conjunction with the annual meeting of the American Association for the Advancement of Science. Boston: Springer; 1970. p. 75–84.
Ising HHUM. Endocrine and cardiovascular effects of noise. Rockville: American Speech, Language, and Hearing Association; 1980.
Peterson EA, Augenstein JS, Tanis DC, et al. Noise raises blood pressure without impairing auditory sensitivity. Science. 1981;211:1450–2. 1981/03/27. https://doi.org/10.1126/science.7466404.
Peterson EA, Augenstein JS, Hazelton CL, et al. Some cardiovascular effects of noise. J Aud Res. 1984;24:35–62. 1984/01/01
Peterson EA, Haselton CL, Augenstein JS. Daily noise duration influences cardiovascular responses. J Aud Res. 1984;24:69–86. 1984/04/01
Weisenberger ME, Krausman PR, Wallace MC, et al. Effects of simulated jet aircraft noise on heart rate and behavior of desert ungulates. J Wildl Manag. 1996;60(1):52–61.
Borg E. Peripheral vasoconstriction in the rat in response to sound. III. Dependence of pause characteristics in continuous noise. Acta Otolaryngol. 1978;86:155–9. 1978/09/01. https://doi.org/10.3109/00016487809124732.
Borg E. Peripheral vasoconstriction in the rat in response to sound. II. Dependence on rate of change of sound level. Acta Otolaryngol. 1978;85:332–5. 1978/05/01. https://doi.org/10.3109/00016487809121460.
Borg E. Peripheral vasoconstriction in the rat in response to sound. I. Dependence on stimulus duration. Acta Otolaryngol. 1978;85:153–7. 1978/03/01. https://doi.org/10.3109/00016487809111921.
Kristensen MP, Rector DM, Poe GR, et al. Activity changes of the cat paraventricular hypothalamus during stressor exposure. Neuroreport. 2004;15:43–8. 2004/04/27. https://doi.org/10.1097/00001756-200401190-00010.
Rosecrans JA, Watzman N, Buckley JP. The production of hypertension in male albino rats subjected to experimental stress. Biochem Pharmacol. 1966;15:1707–18. https://doi.org/10.1016/0006-2952(66)90078-5.
Geber WF. Cardiovascular and teratogenic effects of chronic intermittent noise stress. In: Welch BL, Welch AS, editors. Physiological effects of noise: based upon papers presented at an international symposium on the extra-auditory physiological effects of audible sound, held in Boston, Massachusetts, December 28–30, 1969, in conjunction with the annual meeting of the American Association for the Advancement of Science. Boston: Springer; 1970. p. 85–90.
Lockett MF. Effects of sound on endocrine function and electrolyte excretion. In: Welch BL, Welch AS, editors. Physiological effects of noise: based upon papers presented at an international symposium on the extra-auditory physiological effects of audible sound, held in Boston, Massachusetts, December 28–30, 1969, in conjunction with the annual meeting of the American Association for the Advancement of Science. Boston: Springer; 1970. p. 21–41.
Am S, As W, Bradshaw M, et al. Endocrine changes due to auditory stress. Acta Endocrinol. 1959;31. https://doi.org/10.1530/ACTA.0.XXXI0405
Fink GB, Iturrian WB. Influence of age, auditory conditioning, and environmental noise on sound-induced seizures and seizure threshold in mice. In: Welch BL, Welch AS, editors. Physiological effects of noise: based upon papers presented at an international symposium on the extra-auditory physiological effects of audible sound, held in Boston, Massachusetts, December 28–30, 1969, in conjunction with the annual meeting of the American Association for the Advancement of Science. Boston: Springer; 1970. p. 211–26.
Wysocki AB. The effect of intermittent noise exposure on wound healing. Adv Wound Care J Prev Heal. 1996;9:35–9.
Jensen MM, Rasmussen AF. Audiogenic stress and susceptibility to infection. In: Welch BL, Welch AS, editors. Physiological effects of noise: based upon papers presented at an international symposium on the extra-auditory physiological effects of audible sound, held in Boston, Massachusetts, December 28–30, 1969, in conjunction with the annual meeting of the American Association for the Advancement of Science. Boston: Springer US; 1970. p. 7–19.
Zondek B, Wolstenholme GE, O’Connor M. Effects of external stimuli on reproduction. In honour of Professor B. Zondek. London: Churchill; 1967. p. 4–19.
Barlow SM. Teratogenic effects of restraint, cold and audiogenic stress in mice and rats. London: University of London; 1972.
Borg E, Moller AR. Noise and blood pressure: effect of lifelong exposure in the rat. Acta Physiol Scand. 1978;103:340–2. 1978/07/01. https://doi.org/10.1111/j.1748-1716.1978.tb06223.x.
Snyder-Halpern R. The effect of critical care unit noise on patient sleep cycles. Ccq. 1985;7:41–51. 1985/02/09
Gutierrez Llaneza S, López Romero P, García Camacho M, et al. Effects of construction noise on mouse cardiovascular parameters: arterial blood pressure. In: FELASA meeting Helsinki, Finland, June 14–17, 2010
Jobling M. Fish in aquaculture environments. In: Huntingford F, Jobling M, Kadri S, editors. Aquaculture and behavior. Ames: Blackwell Publishing Ltd.; 2012. p. 36–64.
Reynolds RP, Li Y, Garner A, et al. Vibration in mice: a review of comparative effects and use in translational research. Anim Mod Exp Med. 2018;1:116–24. https://doi.org/10.1002/ame2.12024.
Turner JG, Parrish JL, Hughes LF, et al. Hearing in laboratory animals: strain differences and nonauditory effects of noise. Comp Med. 2005;55:12–23.
Ogilvie DM, Stinson RH. The effect of age on temperature selection by laboratory mice (mus musculus). Can J Zool. 1966;44:511–7. https://doi.org/10.1139/z66-055.
Eedy JW, Ogilvie DM. The effect of age on the thermal preference of white mice (Mus musculus) and gerbils (Meriones unguiculatus). Can J Zool. 1970;48:1303–6. https://doi.org/10.1139/z70-221.
Gaskill BN, Gordon CJ, Pajor EA, et al. Heat or insulation: behavioral titration of mouse preference for warmth or access to a nest. PLoS One. 2012;7:e32799. https://doi.org/10.1371/journal.pone.0032799.
Gaskill BN, Rohr SA, Pajor EA, et al. Working with what you’ve got: changes in thermal preference and behavior in mice with or without nesting material. J Therm Biol. 2011;36:193–9. https://doi.org/10.1016/j.jtherbio.2011.02.004.
Gaskill BN, Rohr SA, Pajor EA, et al. Some like it hot: mouse temperature preferences in laboratory housing. Appl Anim Behav Sci. 2009;116:279–85. https://doi.org/10.1016/j.applanim.2008.10.002.
Baumans V. The laboratory mouse. In: Hubrecht RC, Kirkwood J, editors. The UFAW handbook on the care and management of laboratory and other research animals. Oxford: Wiley; 2010. p. 276–310.
Fischer AW, Cannon B, Nedergaard J. Optimal housing temperatures for mice to mimic the thermal environment of humans: an experimental study. Mol Metab. 2018;7:161–70. 2017/11/11. https://doi.org/10.1016/j.molmet.2017.10.009.
IUPS Thermal Commission. Glossary of terms for thermal physiology. Third ed. Revised by The Commission for Thermal Physiology of the International Union of Physiological Sciences. Jap J Physiol. 2001;51:245–80.
Crippa L, Gobbi A, Ceruti RM, et al. Ringtail in suckling Munich Wistar Fromter rats: a histopathologic study. Comp Med. 2000;50:536–9. 2000/12/01
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Sánchez-Morgado, J.M., Brønstad, A., Pritchett-Corning, K. (2021). Animal and Environmental Factors That Influence Reproducibility. In: Sánchez Morgado, J.M., Brønstad, A. (eds) Experimental Design and Reproducibility in Preclinical Animal Studies . Laboratory Animal Science and Medicine, vol 1. Springer, Cham. https://doi.org/10.1007/978-3-030-66147-2_3
Download citation
DOI: https://doi.org/10.1007/978-3-030-66147-2_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-66146-5
Online ISBN: 978-3-030-66147-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)