
Abstract
Spatial dynamics can promote the evolution of cooperation. While dispersal processes have been studied in simple evolutionary games, real-world social dilemmas are much more complicated. When the investment is low, for example, every additional unit of investment may substantially raise the public goods. However, the effect vanishes as the number of investments increases. Such nonlinear public goods are the norm in a variety of social as well as biological systems. Therefore, we investigate the effect of the nonlinearity on the evolution of cooperation. We show how the nonlinearity in payoffs, resulting in synergy or discounting of public goods, can alter the return on the cooperative investments compared to the linear game. The alteration affects the resulting spatial patterns, not just quantitatively, but in some cases, drastically changing the outcomes. Notably, in cases where a linear game would lead to extinction, synergy can support the coexistence of cooperators and defectors. The eco-evolutionary trajectory can thus be qualitatively different in cases on nonlinear social dilemmas.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others

1 Introduction
The most significant impact of evolutionary game theory has been in the field of social evolution. When an individual’s action results in a conflict between the individual and the group benefit, a social dilemma arises. Social dilemmas can be captured by the two-player prisoners dilemma game [6] and its multiplayer version, the public goods game [8, 17, 24]. The domain of public goods games ranges from behavioural economists, cognitive scientists, psychologists, to biologists given the ubiquity of multiplayer interactions in nature. Situations impossible in two-player games can occur in multiplayer games, which can lead to drastically different evolutionary outcomes [7, 14, 35, 39, 44, 52].
In public goods games, while cooperation raises the group benefit, cooperators themselves get less benefit than defectors. The group benefit typically increases linearly with the number of cooperators in the group. However, in the context of helping behaviour, reference [10] discusses a case where each additional cooperator in the group provides more benefit than the previous (superadditivity of benefit). The approach has been further generalised using a particular nonlinear function where the additional cooperators can provide not only more (synergy) but also less (discounting) benefit than the previous cooperator [20]. The study [5] presents an excellent review of the use and importance of nonlinear public goods game.
The nonlinear public goods game as proposed in [20] has been extended in [13] to include population dynamics. In ecological public goods games, the total density of cooperators and defectors changes, effectively changing the interaction group size. Changes in group size have been shown to result in a stable coexistence of cooperators and defectors [19, 30, 38]. A spatial version of ecological public goods games, where multiple populations of cooperators and defectors are present on a lattice and connected by diffusion, can promote cooperation [53]. The spread of cooperation, in such a case, is possible by a variety of pattern-forming processes.
They use of spatially extended system in different forms such as grouping, explicit space and deme structures, and other ways of limiting interactions have been studied for long [18, 33, 40, 45, 47, 55]. In particular, in [29], the authors provide conditions for strategy selection in nonlinear games about population structure coefficients. The study cited above by [53] while incorporating ecological dynamics focusses solely on linear public goods games.
Previously we have combined a linear social dilemma with density-dependent diffusion coefficients [12, 37]. Including a dynamic diffusion coefficient comes closer to analysing real movements seen across species from bacteria to humans [16, 23, 27, 28, 32, 34, 43]. Incorporating aspects of ecological games as in [19], spatial dynamics per [53] and nonlinear social dilemmas from [13] we develop our previous approach in this study to nonlinear social dilemmas.
We begin by introducing nonlinearity in the payoff function of the social dilemma, including population dynamics. Then we include simple diffusion dynamics and analyse the resulting spatial patterns. For the parameter set comprising of the diffusion coefficients and the multiplication factor, we can observe the extinction, heterogenous or homogenous non-extinction patterns. Under certain simplifying assumptions, we can also characterise the stability of the fixed point and discuss the dynamics of the Hopf-bifurcation transition and the phase boundary between heterogenous- and homogenous-patterned phases. Overall, our results suggest that the spatial patterns while remaining in the same regions relative to each other in the parameter space, synergy and discounting effects shift the boundaries including the phase boundary between extinction and surviving phases. For synergy, the extinction region shrinks as the effective benefit increases resulting in an increased possibility of cooperator persistence. For discounting, however, the extinction region expands. Crucially, the change in the extinction region is not symmetric for synergy and discounting. The above asymmetry is due to the asymmetries in the nonlinear function that we employ for calculating the benefit. The development will help contrast the results with the work of [53] and relates our work to realistic public goods scenarios where the contributions often have a nonlinear impact [9].
2 Model and Results
2.1 Nonlinear Public Goods Game
Complexity of evolutionary games increases as we move from two-player games to multiplayer games [14]. A similar trend ensues as we move from linear public goods games to nonlinear payoff structures [5]. One of the ways of moving from linear to nonlinear multiplayer games is given in [20]. To introduce this method in our notational form, we will first derive the payoffs in a linear setting.
In the classical version of the public goods game (PGG), the cooperators invest c to the common pool while the defectors contribute nothing. The value of the pool increases by a certain multiplication factor r, \(1<r<N\), where N is the group size. The amplified returns are equally distributed to all the N players in the game. For such a setting, the payoffs for cooperators and defectors are given by
where m is the number of cooperators in the group. The nonlinearity in the payoffs can be introduced by the parameter \(\varOmega \) as in [20],
If \(\varOmega >1\) every additional cooperator contributes more than the previous, thus providing a synergistic effect. If \(\varOmega <1\) then every additional cooperator contributes less than the previous, thus saturating the benefits and providing a discounting effect. The linear version of the PGG can be recovered by setting \(\varOmega = 1\).
As in [19] besides the evolutionary dynamics (change in the frequency of cooperators over time), we are also interested in the ecological dynamics (change in the population density over time). This system analysed in [19, 21] is briefly re-introduced in our notation for later extension. We characterise the densities of cooperators and defectors in the population as u and v. Thus \(0\le u+v \le 1\) and the empty space is given by \(w = 1-u-v\). Low population density means that it is hard to encounter other individuals and accordingly hard to interact with them. Hence, the group size N, the maximum group size, in this case, is not always reachable. Instead, S individuals form an interacting group. With fixed N the interacting group size S is bounded, \(S \le N\), and the probability p(S; N) of interacting with \(S-1\) individuals is depending on the total population density \(u+v=1-w\). When we consider the focal individual, the probability p(S; N) of interacting with \(S-1\) individuals among a maximum group of size \(N-1\) individuals (excluding the focal individual),
Then, the average payoffs for defectors and cooperators, \(f_D\) and \(f_C\), are given as
where \(\overline{P_D}(S)\) and \(\overline{P_C}(S)\) are the expected payoffs for defectors and cooperators at a given S. The sum for the group sizes S starts at two as for a social dilemma there need to be at least two interacting individuals.
To derive the expected payoffs, we first need to assess the probability of having a certain number of cooperators m in a group of size \(S-1\) which is given by \(p_c(m;S)\),
Thus, the payoffs in Eq. (2) are weighted with the probability of there being m cooperators, giving us the expected payoffs,
The average payoffs \(f_D\) and \(f_C\) are thus given by
where the investment cost has been set to \(c=1\) without loss of generality. Again, the linear version of the PGG can be recovered by setting \(\varOmega = 1\),
2.2 Spatial Nonlinear Public Goods Games
For tracing the population dynamics, we are interested in the change in the densities of cooperators and defectors over time. Both cooperators and defectors are assumed to have a baseline birth rate of b and death rate of d. Growth is possible only when there is empty space available, i.e. \(w>0\). We track the densities of cooperators and defectors by an extension of the replicator dynamics [19, 22, 48],
To include spatial dynamics in the above system, we assume that a population of cooperators and defectors resides in a given patch. Game interactions only occur within patches, and the individuals can move adjacent patches. The patches live in a two-dimensional space connected in the form of a regular lattice. Taking a continuum limit, we obtain the differential equations with constant diffusion coefficients for cooperators \(D_c\) and defectors \(D_d\),
At the boundaries, there is no in- and out-flux. As in classical activator-inhibitor systems, the different ratio of the diffusion coefficients \(D=D_d/D_c\) can generate various patterns from coexistence, extinction as well as chaos [53].
Nonlinearity in PGG is implemented by \(\varOmega \ne 1\). Previous work shows that the introduction of \(\varOmega \) is enriching the dynamics [13, 20]. Synergy (\(\varOmega >1\)) enhances cooperation while discounting (\(\varOmega <1\)) suppresses it. Accordingly, synergy and discounting with a multiplication factor r can map into the linear game with the higher or lower multiplication factor \(r'\), respectively: \(r'>r\) for synergy and \(r'<r\) for discounting. We call \(r'\) as the effective multiplication factor. As shown in Fig. 1, for synergy effect (\(\varOmega =1.1\)), we can find a chaotic coexistence of cooperators and defectors. The same parameter for a linear case (\(\varOmega = 1.0\)) resulted in total extinction of the population [53]. In the linear case, chaotic patterns were observed for r values larger than that of extinction patterns. Thus, our observation implies the mechanism of how synergy works by effectively increasing r value.
The change in the resulting patterns due to synergy or discounting is not limited to extinction or chaos but is a general feature of the nonlinearity in payoffs. To illustrate this change, we show how a stable pattern under linear PGG (\(\varOmega = 1\)) can change the shape under discounting or synergy in Fig. 2. Such changes in the final structure happen all over the parameter space. To confirm this tendency, we examine the spatial patterns for various parameters and find five phases, same as in the linear PGG case [53] but now with shifted phase boundaries (see Fig. 3). The effective multiplication factor \(r'\) increases with an increasing \(\varOmega \), and thus the location of the Hopf bifurcation also shifts. As a result of shifting \(r_{\mathrm {Hopf}}\), extinction region is reduced in the parameter space with synergy effect. We thus focus our attention on the Hopf-bifurcation point \(r_{\mathrm {Hopf}}\).

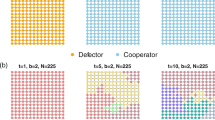
Pattern formation on the two-dimensional square lattice. We observe the chaotic pattern for \(\varOmega =1.1\) (synergy effect) where extinction comes out with \(\varOmega =1\) [53]. Mint green and Fuchsia pink colours represent the cooperator and defector densities, respectively. For a full explanation of the colour scheme, we refer to the appendix. Black indicates no individual on the site, whereas blue appears when the ratio of cooperators and defectors is the same. For a system of size L, initially, a disc with radius L/10 at the centre is occupied by cooperator and defector with densities 0.1. We use multiplication factor \(r=2.2\) and diffusion coefficient ratio \(D=2\). Throughout the paper, for simulations, we used the system size \(L=283\), \(dt=0.1\) and \(dx=1.4\) with the Crank–Nicolson algorithm

Synergy and discounting effects on pattern formation. We get the different patterns under discounting and synergy effects distinct from the linear PGG game at a given the same parameter set. While diffusion-induced instability is observed in the linear PGG, the discounting effect makes diffusion-induced coexistence pattern implying that the discounting effect makes the Hopf-bifurcation point shift to the larger value. Under the synergy effect, on the contrary, we obtain the opposite trend observing the homogenous coexistence pattern. In the linear PGG, the homogenous patterns are observed in higher multiplication factor r, implying the shift of \(r_{\mathrm {Hopf}}\) to the smaller value under the synergy effect

Spatial patterns and corresponding phase diagram for \(\varOmega =1.1\). There are five phases (framed using different colours), extinction (black), chaos (blue), diffusion-induced coexistence (red), diffusion-induced instability (green) and homogeneous coexistence (orange). The Hopf-bifurcation point \(r_{\mathrm {Hopf}}\simeq 2.2208\) and the boundary between diffusion-induced instability and homogeneous coexistence are analytically calculated, while the other boundaries are from the simulation results. All boundaries and \(r_{\mathrm {Hopf}}\) shift to the left, indicating that the multiplication factor r with the synergy maps into the higher multiplication factor \(r'\) in the linear game

Hopf-bifurcation points in \(\varOmega \) and shift of the phase boundary. a The Hopf-bifurcation point \(r_{\mathrm {Hopf}}\) for various \(\varOmega \) (solid line with points). Synergy (\(\varOmega >1\)) decreases \(r_{\mathrm {Hopf}}\) while discounting (\(\varOmega <1\)) increases \(r_{\mathrm {Hopf}}\). By decreasing \(r_{\mathrm {Hopf}}\), the surviving region is extended in the parameter space. The solid line without points is a tangential line at \(\varOmega =1\). b The phase boundaries between diffusion-induced instability and homogeneous coexistence phases are also examined for various \(\varOmega \). Since \(r_{\mathrm {Hopf}}\) increases with a decreasing \(\varOmega \), the boundaries also move to the right
2.2.1 Hopf Bifuraction in Nonlinear PGG
We find the Hopf-bifurcation point \(r_{\mathrm {Hopf}}\) for various \(\varOmega \) values using Eq. (7). The effective multiplication factor \(r'\) increases as \(\varOmega \) increases, and thus \(r_{\mathrm {Hopf}}\) is monotonically decreasing with \(\varOmega \) as in Fig. 4a. The tangential line at \(\varOmega =1\) is drawn for comparing the effects of synergy and discounting. If we focus on the differences between the tangent and \(r_{\mathrm {Hopf}}\) line, synergy changes \(r_{\mathrm {Hopf}}\) more dramatically than discounting. Synergy and discounting effects originate from \(1+(1\pm \Delta \varOmega )+(1\pm \Delta \varOmega )^2+\cdots +(1\pm \Delta \varOmega )^{m-1}\) in Eq. (2), where \(\Delta \varOmega >0\) and plus and minus signs for synergy and discounting, respectively. Straightforwardly, the difference between 1 and \((1+\Delta \varOmega )^k\) is larger than that of \((1-\Delta \varOmega )^k\) for \(k>2\). Hence, the nonlinear PGG itself gives different \(\Delta r_{\mathrm {Hopf}}\) for the same \(\Delta \varOmega \).
2.2.2 Criterion for Diffusion-Induced Instability
Since \(\varOmega \) changes \(r'\) value, the phase boundary also moves. By using the linear stability analysis, we find phase boundaries between diffusion-induced instability and homogeneous coexistence phases in r-D space shown in Fig. 4b. To do that, we introduce new notations, and two reaction–diffusion equations in Eq. (10) can be written as
with density vector \(\mathbf{u} =(u, v)^T\) and matrix \(\mathbf{D} =\left( \begin{array}{ll}D_c&{}0 \\ 0&{}D_d \end{array}\right) \). Elements of the vector \(\mathbf{R} (\mathbf{u} )=\left( \begin{array}{ll}g(u, v) \\ h(u,v) \end{array}\right) \) indicate reaction terms for each density which is the second terms in Eq. (10). Without diffusion, the differential equations have homogeneous solution \(\mathbf{u} _0=(u_0, v_0)^T\) where \(g(u_0, v_0)=h(u_0, v_0)=0\). We assume that the solution is a fixed point, and examine its stability under diffusion.
If we consider small perturbation \(\tilde{\mathbf{u }}\) from the homogeneous solution, \(\mathbf{u} \cong \mathbf{u} _0+\tilde{\mathbf{u }}\), we get the relation,
where \(\mathbf {J}=(\partial \mathbf {R}/\partial \mathbf {u})_{\mathbf {u_0}} \equiv \left. \left( \begin{array}{ll}g_u&{}g_v \\ h_u&{} h_v \end{array}\right) \right| _{\mathbf {u_0}}\). Subscripts of the g and h mean partial derivative of that variable, e.g. \(g_u\) means \(\partial g/\partial u\). Decomposing \(\tilde{\mathbf {u}}=\sum _k \mathbf {a}_k e^{ik\mathbf {r}}\) based on propagation wave number k gives us relation \(\dot{\mathbf {a}_k}=\mathbf {B}\mathbf {a}_k\) where \(\mathbf {B}\equiv \mathbf {J}-k^2\mathbf {D}\). Therefore, the stability of the homogeneous solution can be examined by the matrix \(\mathbf{B} \). Note that \(\mathrm {Tr}(\mathbf{B} )<0\) is guaranteed because \(\mathrm {Tr}(\mathbf{J} )<0\). Hence, if the determinant of \(\mathbf{B} \) is smaller than zero \([\det (\mathbf{B} )<0]\), it means one of the eigenvalues of the matrix \(\mathbf{B} \) is positive. The homogeneous solution becomes unstable and Turing patterns appear.
The condition for \(\det (\mathbf{B} )<0\) is given by
It can be rewritten as following form:
With our model parameters this inequality is equivalent to
where \(u^*\) and \(v^*\) are values at the fixed point. The symbols indicate
with \(\partial _x y = \frac{\partial y}{\partial x}\). If the above criterion is satisfied, the stable fixed point predicted without diffusion becomes unstable with diffusion (Fig. 5).

Schematic figure for expected shift of phase boundaries. According to the change of \(r_{\mathrm {Hopf}}\), over all phase boundaries may shift together at the same direction. As we have seen in Fig. 4b, the phase boundaries with \(r_{\mathrm {Hopf}}\) move to the right with discounting effect and move to the left with synergy effect, respectively. Accordingly, the surviving region in the parameter space expands with synergy effect while it shrinks with discounting effect
3 Discussion
Linear public goods game is a useful approximation of the real nonlinearities in social dilemmas from microbes to macro-life [15, 36, 49] with applications such as in antibiotic resistance [25] as well as cancer [1]. However, taking nonlinearities into account might show different resulting outcomes from naive expectations [13]. Especially, nonlinearities in interactions have a profound effect in ecology when it comes to fecundity and avoiding predation [56, 57]. In this manuscript, we have extended the analysis of spatial public goods games beyond the traditional linear public goods games.
The benefits, in our case, accumulate in a nonlinear fashion in the number of cooperators in the group. Each cooperator can provide a larger benefit than the last one as the number of cooperators increases (resulting in a synergy), or each cooperator provides a smaller benefit than the previous one (thus leading to discounting) [20]. Such an extension to public goods game was proposed very early on by [10]. Termed as superadditivity in benefits, extending from this particular model framework, we can visualise nonlinearities in costs as well, a concept not yet dealt with. Again, such economies of scale [9] can be justified in both bacterial and human interactions as proxies for quorum sensing (or quenching) or accruing of wealth (or austerity) [3, 4, 40].
We show that including such nonlinearities in the benefit function affects the effective rate of return from the public goods game, irrespective of the types of diffusion dynamics. With spatial dynamics, synergy increases the effective rate of return on the investment and expands the region in the parameter space where survival of the population is possible. This itself may make cooperation a favourable strategy. Besides the trivial observation that synergy helps cooperators, we show that as we move symmetrically away from the linear case towards more synergy or discounting, the change in the eventual dynamics is not symmetric. It would be interesting to check if the asymmetry holds for different designs of benefit functions.
We used the particular functional form of the benefit function, including nonlinearities in payoffs [20]. However, there are various ways of including nonlinearities in the benefit function [4, 7, 40]. The model considered in [40] extends the results of [20] to games between relatives. Furthermore, [40] has described the relationships between different nonlinear social dilemma models with a variety of benefit functions. Also these nonlinear social dilemmas have been analysed in a structured population [29, 40, 41]. However, previous studies have focused on the approach presented in [46], which provide a criterion for strategy selection rather than explicitly positioning the populations on a grid and including diffusion. When studying games in structured populations, often a network structure is considered [2, 42]. The role of network connectivity is determined to be critical for the eventual evolutionary outcome [41, 50, 51] and some structures can result in hindering the evolution of cooperation as well [26]. In contrast, our approach focuses more on the ecological framework but not in network structures. We take into account not only the changes in frequencies of cooperators and defectors but also the population dynamics, which is usually missed in a network approach. While both approaches make evolutionary games ecologically explicit, the models are thoroughly different in their setup and implementation.

The exact colour scheme developed for colouring the patterns. Each patch in a pattern is coloured using this palette by choosing the corresponding f and \(\rho \) values. For brightness we used Eq. (17) with \(a=15\)
The importance of including ecology in evolutionary games has been known for long, but the complexity that it generates has prevented it from garnering widespread attention [11]. Seasonal variations in the rate of return radically change the selection pressures on cooperation and defection. Changes in the ecology may not feedback directly to the frequencies of cooperators and defectors but on to a variable in benefit–cost functions. If the variable affects the frequency of cooperators and defectors (or even the group size) in a nonlinear fashion, then the results are not trivial [13, 38]. Thus, even a simple connection between evolutionary and ecological dynamics may already generate rich dynamics [31, 54], and the feedback between the two is often already convoluted. Similar to [12, 37], it is possible to include feedback between the population dynamics and diffusion here, but together with a nonlinear social dilemma, we envision that the formal analysis and the computational implementation will be a considerable challenge.
References
Aktipis A (2016) Principles of cooperation across systems: from human sharing to multicellularity and cancer. Evolutionary Applications 9(1):17–36
Allen B, Lipper G, Chen YT, Fotouhi B, Nowak MA, Yau ST (2017) Evolutionary dynamics on any population structure. Nature 544:227–230
Archetti M (2009) Cooperation as a volunteer’s dilemma and the strategy of conflict in public goods games. Journal of Evolutionary Biology 11:2192–2200
Archetti M, Scheuring I (2011) Co-existence of cooperation and defection in public goods games. Evolution 65(4):1140–1148
Archetti M, Scheuring I (2012) Review: Game theory of public goods in one-shot social dilemmas without assortment. Journal of Theoretical Biology 299(0):9–20
Axelrod R (1984) The evolution of cooperation. Basic Books, New York, NY
Bach LA, Helvik T, Christiansen FB (2006) The evolution of \(n\)-player cooperation - threshold games and ESS bifurcations. Journal of Theoretical Biology 238:426–434
Binmore KG (1994) Playing fair: game theory and the social contract. MIT Press, Cambridge
Dawes RM, Orbell JM, Simmons RT, Van De Kragt AJC (1986) Organizing groups for collective action. The American Political Science Review 80(4):1171–1185
Eshel I, Motro U (1988) The Three Brothers’ Problem: Kin Selection with More than One Potential Helper. 1. The Case of Immediate Help. The American Naturalist 132(4):550–566
Estrela S, Libby E, Van Cleve J, Débarre F, Deforet M, Harcombe WR, Peña J, Brown SP, Hochberg ME (2018) Environmentally Mediated Social Dilemmas. Trends in Ecology & Evolution 34(1):6–18
Funk F, Hauert C (2019) Directed migration shapes cooperation in spatial ecological public goods games. PLOS Computational Biology 15(8):1–14
Gokhale CS, Hauert C (2016) Eco-evolutionary dynamics of social dilemmas. Theoretical Population Biology 111:28–42
Gokhale CS, Traulsen A (2010) Evolutionary games in the multiverse. Proceedings of the National Academy of Sciences USA 107:5500–5504
Gore J, Youk H, van Oudenaarden A (2009) Snowdrift game dynamics and facultative cheating in yeast. Nature 459:253–256
Grauwin S, Bertin E, Lemoy R, Jensen P (2009) Competition between collective and individual dynamics. Proceedings of the National Academy of Sciences of the United States of America 106(49):20,622–20,626
Hardin G (1968) The tragedy of the commons. Science 162:1243–1248
Hauert C, Imhof L (2012) Evolutionary games in deme structured, finite populations. Journal of Theoretical Biology 299:106–112
Hauert C, Holmes M, Doebeli M (2006a) Evolutionary games and population dynamics: maintenance of cooperation in public goods games. Proceedings of the Royal Society B 273:2565–2570
Hauert C, Michor F, Nowak MA, Doebeli M (2006b) Synergy and discounting of cooperation in social dilemmas. Journal of Theoretical Biology 239:195–202
Hauert C, Yuichiro Wakano J, Doebeli M (2008) Ecological public goods games: cooperation and bifurcation. Theoretical Population Biology 73:257–263
Hofbauer J, Sigmund K (1998) Evolutionary Games and Population Dynamics. Cambridge University Press, Cambridge, UK
Kawasaki K, Mochizuki A, Matsushita M, Umeda T, Shigesada N (1997) Modeling spatio-temporal patterns generated by bacillus subtilis. Journal of Theoretical Biology 188:177–185
Kollock P (1998) Social dilemmas: The anatomy of cooperation. Annual Review of Sociology 24:183–214
Lee HH, Molla MN, Cantor CR, Collins JJ (2010) Bacterial charity work leads to population-wide resistance. Nature 467(7311):82–85
Li A, Broom M, Du J, Wang L (2016) Evolutionary dynamics of general group interactions in structured populations. Physical Review E 93(2):022,407
Loe LE, Mysterud A, Veiberg V, Langvatn R (2009) Negative density-dependent emigration of males in an increasing red deer population. Proc R Soc B 276:2581–2587
Lou Y, Martínez S (2009) Evolution of cross-diffusion and self-diffusion. Journal of Biological Dynamics 3(4):410–429
McAvoy A, Hauert C (2016) Structure coefficients and strategy selection in multiplayer games. Journal of Mathematical Biology pp 1–36
McAvoy A, Fraiman N, Hauert C, Wakeley J, Nowak MA (2018) Public goods games in populations with fluctuating size. Theoretical Population Biology 121:72–84, https://doi.org/10.1016/j.tpb.2018.01.004
McNamara JM (2013) Towards a richer evolutionary game theory. Journal of The Royal Society Interface 10:20130,544
Ohgiwari M, Matsushita M, Matsuyama T (1992) Morphological changes in growth phenomena of bacterial colony patterns. J Phys Soc Jap 61:816–822
Ohtsuki H, Pacheco J, Nowak MA (2007) Evolutionary graph theory: Breaking the symmetry between interaction and replacement. Journal of Theoretical Biology 246:681–694
Okubo A, Levin SA (1980) Diffusion and Ecological Problems: Mathematical Models. Springerr-Verlag
Pacheco JM, Santos FC, Souza MO, Skyrms B (2009) Evolutionary dynamics of collective action in n-person stag hunt dilemmas. Proceedings of the Royal Society B 276:315–321
Packer C, Ruttan L (1988) The evolution of cooperative hunting. The American Naturalist 132:159–198
Park HJ, Gokhale CS (2019) Ecological feedback on diffusion dynamics. Journal of the Royal Society Open Science 6:181,273
Peña J (2012) Group size diversity in public goods games. Evolution 66:623–636
Peña J, Lehmann L, Nöldeke G (2014) Gains from switching and evolutionary stability in multi-player matrix games. Journal of Theoretical Biology 346:23–33
Peña J, Nöldeke G, Lehmann L (2015) Evolutionary dynamics of collective action in spatially structured populations. Journal of Theoretical Biology 382:122–136
Peña J, Wu B, Arranz J, Traulsen A (2016) Evolutionary games of multiplayer cooperation on graphs. PLoS Computational Biology 12(8):1–15
Santos FC, Pacheco JM, Lenaerts T (2006) Evolutionary dynamics of social dilemmas in structured heterogeneous populations. Proceedings of the National Academy of Sciences USA 103:3490–3494
Shigesada N, Kawasaki K, Teramoto E (1979) Spatial segregation of interacting species. Journal of Theoretical Biology 79(1):83–99
Souza MO, Pacheco JM, Santos FC (2009) Evolution of cooperation under n-person snowdrift games. Journal of Theoretical Biology 260:581–588
Tarnita CE, Antal T, Ohtsuki H, Nowak MA (2009a) Evolutionary dynamics in set structured populations. Proceedings of the National Academy of Sciences USA 106:8601–8604
Tarnita CE, Ohtsuki H, Antal T, Fu F, Nowak MA (2009b) Strategy selection in structured populations. Journal of Theoretical Biology 259:570–581
Tarnita CE, Wage N, Nowak MA (2011) Multiple strategies in structured populations. Proceedings of the National Academy of Sciences USA 108:2334–2337
Taylor PD, Jonker LB (1978) Evolutionarily stable strategies and game dynamics. Mathematical Biosciences 40:145–156
Turner PE, Chao L (1999) Prisoner’s Dilemma in an RNA virus. Nature 398:441–443
van Veelen M, Nowak MA (2012) Multi-player games on the cycle. Journal of Theoretical Biology 292:116–128
van Veelen M, García J, Rand DG, Nowak MA (2012) Direct reciprocity in structured populations. Proceedings of the National Academy of Sciences USA 109:9929–9934
Venkateswaran VR, Gokhale CS (2019) Evolutionary dynamics of complex multiple games. Proceedings of the Royal Society B: Biological Sciences 286(1905):20190,900
Wakano JY, Nowak MA, Hauert C (2009) Spatial dynamics of ecological public goods. Proceedings of the National Academy of Sciences USA 106:7910–7914
Weitz JS, Eksin C, Paarporn K, Brown SP, Ratcliff WC (2016) An oscillating tragedy of the commons in replicator dynamics with game-environment feedback. Proceedings of the National Academy of Sciences of the United States of America 113(47):E7518–E7525
Wright S (1930) The genetical theory of natural selection. Journal of Heredity 21:349–356
Wrona FJ, Jamieson Dixon RW (1991) Group Size and Predation Risk: A Field Analysis of Encounter and Dilution Effects. The American Naturalist 137(2):186–201
Zöttl M, Frommen JG, Taborsky M (2013) Group size adjustment to ecological demand in a cooperative breeder. Proceedings of the Royal Society B: Biological Sciences 280(1756):20122,772
Acknowledgments
We thank Christoph Hauert for comments and suggestions in improving an early version of the manuscript. The authors thank the constructive comments of the reviewers. Both authors acknowledge generous support from the Max Planck Society.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendix
Appendix
1.1 Colour Coding
Similar to the colour coding used in [37] we use mint green (colour code: #A7FF70) and Fuchsia pink (colour code: #FF8AF3) colours for denoting the cooperator and defector densities, respectively, for each type. The colour spectrum and saturation is determined by the ratio of cooperators to defectors which results in the Maya blue colour for equal densities of cooperators and defectors. For convenience, we use HSB colour space which is a cylindrical coordinate system \((r, \theta , h)=\) (saturation, hue, brightness). The radius of circle r indicates saturation or the colour whereas \(\theta \) helps us transform the RGB space to HSB. The total density of the population \(\rho =u+v\) is represented by the brightness h of the colour. For better visualisation, we formulate the brightness h as
where a control parameter a \((>-1\) and \(\ne 0)\) (see Fig. 6). The complete colour scheme so developed passes the standard tests for colour blindness.
Rights and permissions
Copyright information
© 2020 The Editor(s) (if applicable) and The Author(s), under exclusive license to Springer Nature Switzerland AG
About this paper

Cite this paper
Gokhale, C.S., Park, H.J. (2020). Eco-evolutionary Spatial Dynamics of Nonlinear Social Dilemmas. In: Ramsey, D.M., Renault, J. (eds) Advances in Dynamic Games. Annals of the International Society of Dynamic Games, vol 17. Birkhäuser, Cham. https://doi.org/10.1007/978-3-030-56534-3_8
Download citation
DOI: https://doi.org/10.1007/978-3-030-56534-3_8
Published:
Publisher Name: Birkhäuser, Cham
Print ISBN: 978-3-030-56533-6
Online ISBN: 978-3-030-56534-3
eBook Packages: Mathematics and StatisticsMathematics and Statistics (R0)