
Abstract
Ischemia-reperfusion injury (IRI) is an important problem in transplantation when blood flow is restored in damaged organs resulting in significant morbidity and mortality in recipient. Liver grafts from donation after circulatory death (DCD) have a more prolonged warm ischemia time that can result in decreased postoperative liver function and increased bile duct injury.
Liver IRI induces damage of hepatocytes and liver sinusoidal endothelial cells (SECs), which is regulated by several molecular pathways. During the ischemic period, there is a metabolic disturbance with adenosine triphosphate (ATP) depletion of cells and lactic acidosis, which induces mitochondrial dysfunction and production of reactive oxygen species (ROS). After reperfusion and restoration of oxygen supply, ROS and damage-associated molecular patterns (DAMPs) are produced. Damaged and activated SEC and liver macrophages (Kupffer cells) promote neutrophil migration and platelet activation by releasing pro-inflammatory mediators such as cytokines and chemokines. Migrated neutrophils secrete ROS, which further promotes cell death. Activated platelets induce a reduction of microcirculation by vasoconstriction and thrombosis in the sinusoid.
In the clinical setting, three main therapeutic strategies can be applied to protect the DCD liver graft from ischemic damage and to improve postoperative function: (1) minimizing the ischemia time, (2) providing thrombolytic therapy during the transplant procedure, and (3) carrying out in situ and ex situ machine perfusion. Especially, the ex situ machine perfusion is a promising approach for the protection from IRI with improved hepatic function and a reduction of biliary injury.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Ischemia-reperfusion injury
- Donation after circulatory death
- Liver sinusoidal endothelial cells
- Hepatocytes
- Warm ischemia time
- Cold ischemia time
- Ischemic-type biliary lesions
- Ischemic cholangiopathy
- Ex vivo machine perfusion
- Normothermic ex situ machine perfusion
- Hypothermic ex situ machine perfusion
- Normothermic regional machine perfusion
Introduction
Ischemia-reperfusion injury (IRI) is common during transplantation when blood flow is restored and oxygen and nutrients are returned to the liver following ischemic injury. Although donation after circulatory death (DCD) is one important strategy to expand the donor pool, it is associated with severe reperfusion injury. Liver grafts from DCD donors are exposed to the agonal phase during donation resulting in an additional warm ischemia time with insufficient blood supply.
Liver IRI is regulated by several molecular pathways. Reperfusion injury results in significant changes in hepatocytes and liver sinusoidal endothelial cells (SECs). The prolonged ischemic period results in a depletion of adenosine triphosphate (ATP) with an activation of mediators of apoptosis and necrosis in liver cells. After reperfusion, neutrophils and liver macrophages (Kupffer cells) are activated in damaged livers, which amplify IRI by secretion of paracrine and autocrine signals, such as reactive oxygen species (ROS), lipid peroxidation, and damage-associated molecular patterns (DAMPs) [1].
In this chapter, we will focus on mechanisms of IRI in hepatocytes and bile ducts and discuss therapeutic strategies targeted on molecular mechanism of IRI in liver transplantation using DCD donors.
Molecular Mechanisms of IRI: Ischemic Period
According to the revised Maastricht classification in 2013 [2], DCD transplantation was categorized into two major types: controlled DCD (cDCD) and uncontrolled DCD (uDCD).
In cDCD, during the agonal phase, the oxygen saturation and the blood pressure are decreasing following withdrawal of life-sustaining therapies (WLST), and donor death is declared 2–5 minutes after a no-touch period [3]. Following death declaration, cold flush or regional perfusion is performed, and the organs are procured. Warm ischemia time has been variably defined, but it is necessary to consider the agonal phase (from WLST to cardiac arrest) as a relative ischemia time. A retrospective study in five major liver transplant centers determined that functional DWIT with SpO2 ≤ 60% is an important predictive parameter for postoperative complications in DCD liver transplantation [4].
In uDCD, the donor underwent an unexpected cardiac arrest outside the hospital with unsuccessful cardiopulmonary resuscitation before determination of death. A prolonged time period exists between cardiac arrest and arrival at the hospital prior to death declaration. The extent of the ischemia is more uncertain in uDCD, making the post-transplant severity of IRI difficult to predict.
Prolonged warm ischemic injury of more than 30 minutes is a well-known risk factor for post-transplant liver failure [5,6,7,8,9,10]. During ischemia, the cell death is mainly caused by metabolic disturbances [11]. Depletion of oxygen causes cell hypoxia that results in an inhibition of the electron transport in the respiratory chain and a decrease in intracellular ATP levels. ATP-dependent ion channels such as Na+/K+ adenosine triphosphatase (ATPase), Na+/H+ exchanger, and Ca channels start to fail, which induces depolarization of the cell membrane with accumulation of intracellular Na+ and Ca2+ and cellular edema. This activates proteases, lipases, phospholipases, and ATPases promoting hepatic apoptosis and necrosis. At the same time, anaerobic respiration induced by insufficient oxygenation supply causes lactic acidosis that further activates intracellular proteases. The increase of Ca2+ influx and accumulation of adenosine diphosphate (ADP), adenosine monophosphate (AMP), and phosphate in hepatocyte lead to mitochondrial membrane permeability transition (MMPT) [12]. MMPT induces mitochondrial swelling and allows soluble molecules with a molecular weight of less than 1500 kDa to pass through the “ionic mega-channels” of the mitochondrial membrane and further enhances the liver damage [13]. Furthermore, warm ischemia decreases phospholipid cardiolipin (diphosphatidylglycerol), which is an essential predominant mitochondrial phospholipid and increases oxidized form of cardiolipin in hepatocyte [14]. These pathways cause mitochondrial dysfunction and promote cell death (Fig. 6.1).

Mechanism of cell damage in ischemic period. Abbreviation: ROS reactive oxygen species, DAMPs damage-associated molecular patterns, ATP adenosine triphosphate, ADP adenosine diphosphate
After procurement with an organ preservation solution, the liver is stored on ice (at 4 degrees Celsius). This second ischemic phase is called cold ischemia time (CIT) and is associated with cold ischemic injury until liver is successfully reperfused in the recipient. During this time, liver metabolism is reduced, and ATP stores within cells are depleted less rapidly [15]. SEC is sensitive against cold storage [16, 17]. ATP depletion during the ischemic phase in SEC induces not only mitochondrial dysfunction but also actin-fiber disassembly [18] and the release of matrix metalloproteinases (MMP-2, MMP-9) [19]. This results in an expression of von Willebrand factor (vWF) and P-selectin on the endothelial cell surface, which promotes thrombosis after reperfusion [20].
Molecular Mechanism of IRI: Reperfusion Period
While the warm and cold ischemic phases condition the liver cells to preservation injury, it is the reperfusion phase when the apoptotic and necrotic pathways are executed, and the cell death occurs. Reperfusion increases the intracellular Ca2+ concentration and the production of reactive oxygen species (ROS) by several pathways such as neutrophil migration and inflammatory cytokines and chemokines such as TNF-α, IL-1β, IFN-γ, and IL-12 (Fig. 6.2). This leads to irreversible cellular and mitochondrial changes and cell death.

Main mechanism of ischemia-reperfusion injury. Abbreviation: SECs Liver sinusoidal endothelial cells, ROS reactive oxygen species, DAMPs damage-associated molecular patterns, TNF-α tumor necrosis factor-α, IL-1β interleukin-1β, IFN-γ interferon-γ, IL-12 interleukin-12, TXA2 thromboxane A2, NO nitric oxide, PAI-1 plasminogen activator inhibitor-1, TGF-β transforming growth factor-β
Reperfusion injury involves numerous parenchymal cells as well as non-parenchymal cells that interact in a network of simultaneous events prompting pro-inflammatory change and cell injury.
Hepatocytes develop cell swelling, lactic acidosis, and mitochondrial dysfunction induced by ATP depletion and hypoxia during the ischemic phase. After reperfusion, the surplus oxygen is not used in the respiratory chain which results in the generation of oxygen free radicals that lead to cell death [21]. In addition, damaged hepatocytes secrete DAMPs such as HMGB-1, histone/DNA, and ATP to activate Kupffer cells and neutrophils as a sterile inflammation [22]. These productions of ROS and DAMPs promote more severe damage of hepatocytes.
Sinusoidal endothelial cells (SECs) play a key role to control sinusoidal blood flow, oxygen supply, and delivery of nutrients for liver tissue by regulating vascular tone [23]. SEC injury gives rise to cell swelling as well as detachment. Mitochondrial injury results in decreased NO (nitric oxide) production and depletion of NO stores. The balance between the vasorelaxation effect of NO and the vasoconstrictor effects of TXA2 (thromboxane A2) from platelet becomes disturbed, which leads to an increase of the vascular tone and decrease of the hepatic blood flow [24, 25]. In addition, activated SECs express P-selectin which enhances platelet adhesion and activation. Adhesion of platelet further promotes cell death and decreases sinusoidal microcirculation by inducing congestion and reducing flow [26, 27].
Kupffer cells play a central role in the pro-inflammatory cascade after reperfusion. Under normal circumstances in the absence of preservation injury, Kupffer cells present circulating antigens from the blood to T cells and induce tolerogenic T cells to produce anti-inflammatory cytokines (IL-10) [28]. In contrast, during IRI, Kupffer cells recognize DAMPs from hepatocytes and SEC through Toll-like receptors 3, 4, and 9 and secrete pro-inflammatory cytokines such as TNF-α, IL-1β, IFN-γ, and IL-12. These mediators induce neutrophil migration to the liver and the release of ROS from neutrophils and promote platelet adhesion on SEC [29,30,31].
Neutrophils are main actors during IRI. After reperfusion, the complement system is activated and enhances production of complement protein 3a (C3a), complement protein 5a (C5a), and the membrane attack complex (MAC). This complement activation leads to the recruitment of pro-inflammatory cells including neutrophils to the damaged liver and promotes in cell death [32, 33]. In the liver, neutrophils detect chemokines such as CXCL1 and CXCL2 secreted by activated Kupffer cells, which guides them into the sinusoids [34]. Chemokines also bind to glycosoaminoglycans on the vascular surface of SEC. When neutrophils reach the SEC, chemokine-chemokine receptor interactions activate the integrins. Neutrophils bind SEC through integrin CD11b/CD18a (Mac-1) on the neutrophils and intracellular adhesion molecule-1 (ICAM-1) on SEC [35]. Neutrophils also respond to inflammatory signals (DAMPs) in the liver, such as high-mobility group box 1 (HMBG-1) and DNA fragment released from injured hepatocytes. These substances enhance the production of ROS from neutrophils through DAMP receptors including Toll-like receptor (TLR) [36]. DNA fragments activate TLR9 on neutrophils, which plays a significant role in neutrophil migration, activation, and production of ROS [37]. Damaged hepatocytes release the HMGB-1, which activates TLR4 and amplifies hepatic injury [38]. This cascade causes further migration of inflammatory cells, and liver tissue damage creates a positive feedback loop [39].
Platelets have been recognized as important players within the hepatic reperfusion injury cascade. Activated Kupffer cells by DAMPs from hepatocytes and SEC release TNF-α. TNF-α induces the P-selectin on SEC and promotes platelet adhesion and activation [40]. This leads to microthrombosis in sinusoid and induces apoptosis of SEC [27]. On the other hand, SECs express CD39 (ectonucleoside triphosphate diphosphohydrolase-1 (ENTPD1)) on the luminal side, which is a regulator of ATP and ADP in platelets. When SECs are injured, CD39 activity decreases and ADP increases in the extracellular environment. ADP is a key inducer of platelet aggregation, and platelets are activated [41]. Furthermore, damaged SECs can result in endothelial fenestrations allowing platelets to enter the space of Disse. Platelets attach to the collagen type III in space of Disse and aggregate, which is called “extravasated platelet aggregation” [42]. Activated platelets release negative mediators, such as thromboxane A2 (TXA2) [24], serotonin [27], plasminogen activator inhibitor-1 (PAI-1) [43], and TGF-β [44]. TXA2 and secretin can induce portal hypertension, while PAI-1 and TGF-β promote hepatic fibrosis and suppression of liver regeneration.
Therapeutic Strategies
Minimizing Ischemia Times
It is important to realize that minimizing ischemia is a low-cost and highly effective way to reduce preservation injury in liver grafts from DCD donors. Warm ischemia has severe effects in DCD grafts, but the length of warm ischemia does not linearly correlate with the severity of injury [45]. Prolonged CIT of more than 8 hours is a risk factor for graft failure and ischemic cholangiopathy [6,7,8,9, 46,47,48]. To shorten the cold ischemia time, some institutes start the recipient hepatectomy prior to the procurement team’s return when using grafts from DCD donors [7, 49]. Other possible strategies to minimize warm ischemic injury are ante-mortem procedures such as donor anticoagulation, administration of vasodilators, and femoral cannulation for regional perfusion. Limitations include legal and cultural restrictions as well as the limited scientific evidence of the beneficial effects of antemortem strategies on postoperative outcomes [50].
Thrombolytic Therapy
Biliary complications are common after DCD liver transplantation. Ischemic-type biliary lesions (ITBLs) and ischemic cholangiopathy (IC) occur in 12% to 50% of DCD transplantations resulting frequently in graft loss [7, 10, 51, 52]. These biliary complications are thought to be caused by insufficient arterial blood supply of the intra- and extrahepatic bile ducts. While the liver parenchyma receives the dual blood flow from the hepatic artery and the portal vein, the blood supply for bile duct comes only from hepatic artery via peribiliary vascular plexus [53]. Dries et al. analyzed biliary injury of 128 liver transplants including 29 from DCD donors. The authors demonstrated that 92% of the bile epithelium was injured at the end of cold storage with a luminal epithelium loss >50%. In addition, the peribiliary glands which promote biliary regeneration were damaged in 57% of the superficial periluminal side and 18% in deep bile duct wall. Furthermore, the mural stroma necrosis, vascular injury, intramural bleeding, and inflammation worsened after reperfusion [54].
To dissolve the microthrombi and to obtain sufficient blood flow in biliary microcirculation, some transplant programs used thrombolytic therapy during back-table preparation or implantation of the liver graft. Hashimoto et al. reported their experience with the tissue plasminogen activator (tPA) flush on the back table for 22 DCD liver grafts [55]. Several other groups reported the use of tPA during DCD liver transplantation, and a systematic review indicated that thrombolytic therapies in DCD liver transplantation statistically decreased ITBLs and retransplantation rate and improved 1-year graft survival without the risk of increasing blood transfusion [53]. However, the efficacy of thrombolytic therapy is still controversial in the absence of randomized controlled trials and the differences within the tPA injection protocols. In addition, there is a significant variation in functional DWIT and CIT between studies, which makes the comparison of the different trials difficult [56]. This topic is covered in more depth in Chap. 8, “tPA and Thrombolytic Therapy.”
Machine Perfusion
Machine perfusion (MP) is a novel strategy for preservation of DCD grafts. Machine perfusion can be performed as in situ and ex situ machine perfusion. Ex situ machine perfusion has been performed at physiologic temperatures (warm perfusion) with oxygen and nutrition, while cold (4 °C degrees) ex situ machine perfusion has been developed with or without oxygen.
Ex situ MP is currently categorized into three groups: post-static cold storage (SCS) MP , replacing cold storage with MP , and ischemia-free liver transplantation without warm or cold ischemic preservation [57]. In post-SCS MP, liver graft is perfused after cold storage and transport of the liver graft to the recipient hospital. With preservation MP, the perfusion starts at the donor hospital after cold flush and continues until transplantation. Ischemia-free liver transplantation is a novel method to connect the perfusion machine to the donor vessels and continue perfusion until reperfusion without any ischemia. In clinical settings, perfusate temperature and perfusate type differ between each perfusion strategy. Two different temperature settings have been used during machine perfusion: hypothermic MP (HMP , 0–12 degrees) and normothermic MP (NMP , 35–38 degrees). Although each perfusion has its own protective effects against IRI, the basic merits of MP in both settings are decreased preservation injury, graft assessment, and graft reconditioning, compared with SCS.
Hypothermic Oxygenated Ex Situ Machine Perfusion
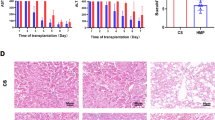
Oxygenated HMP increases ATP and attenuates the inflammatory IRI cascade compared with static cold storage. Oxygenated HMP reduces mitochondrial injury [58] and improves ATP storage during preservation [15]. Furthermore, compared with SCS group, HMGB-1, which is one of DAMPs and representative of nuclear damage, was lower, Kupffer cell activation was suppressed, and expression of vWF on LSECs was significantly decreased in HMP group [58]. In a matched control clinical trial, Schlegel et al. demonstrated that liver grafts from DCD donors with oxygenated HMP had significantly lower graft loss at 5 years after transplantation (HMP-DCD 8% vs SCS-DCD 32%) [59]. Oxygenated HMP in DCD liver grafts also decreased biliary injury after transplantation by reducing biliary fibrosis with less activated myofibroblasts compared with SCS-preserved grafts [60]. In a clinical trial, Rijn et al. demonstrated that oxygenated HMP-DCD liver transplantation reduced IRI of the bile duct when compared with DCD-SCS controls, with less mural stroma necrosis and better preservation of periluminal peribiliary glands after reperfusion [61, 62]. This was associated with a significantly lower rate of graft loss by ischemic cholangiopathy (HMP-DCD 0% vs SCS-DCD 10%) [59] (Fig. 6.3a).

Clinical evidence of ex vivo machine perfusion for DCD liver graft. (a) Hypothermic ex situ perfusion. (b) Normothermic ex situ perfusion. ∗1 1 liver was not perfused because of portal vein torsion. (c) Normothermic regional perfusion. Abbreviation: WIT warm ischemia time, CIT cold ischemia time, PNF primary non-function, DCD donation after cardiocirculatory death, DBD donor after brainstem death, HMP hypothermic machine perfusion, NMP normothermic machine perfusion, NRP normothermic regional perfusion, HA hepatic artery, PV portal vein, NA not available
Normothermic Ex Situ Machine Perfusion (NMP)
The liver is metabolically active during normothermic ex situ perfusion, which offers the opportunity to assess the hepatocyte and cholangiocyte viability. Aminotransferase levels in the perfusate can be determined as hepatocyte injury maker. As hepatocellular functional parameters, lactate clearance, bile volume and quality (bile pH, bicarbonate and glucose levels), and glucose consumption can be measured [63]. Several markers during normothermic ex situ perfusion were reported to be associated with post-transplant primary non-function liver. Mergental et al. defined viability criteria during NMP. These viability criteria consisted of lactate clearance, pH maintenance, bile production, vascular flow patterns, and liver macroscopic appearance based on data of human discarded livers [64].
NMP has been demonstrated to replenish ATP levels in hepatocyte [66,67,68,68], significantly lower aminotransferase after transplantation, and result in better survivals in pig DCD liver transplant models [69]. Recently, Jassem demonstrated that NMP leads to an upregulation of gene expression of tissue regeneration and platelet function and a reduced expression of immune-related genes. NMP induces regulatory T cells and reduces the proportion of CD4-positive T cells producing IL-2, IL-4, IFN-γ, and IL-17 and CD8-positive T cells producing IFN-γ. This results in a suppression of neutrophil infiltration and reduction of parenchymal cell death compared with SCS [70]. Nasralla et al. reported the first randomized trial of NMP with 220 human livers including 53 DCD livers. They demonstrated lower level of graft injury (peak AST NMP 488.1 vs SCS 964.9 IU/L), lower discarded rate (NMP 11.7% vs SCS 24.1%), and lower rate of early allograft dysfunction (NMP 10.2% vs SCS 29.9%) [71].
NMP also decreases biliary IRI and promotes bile regeneration in DCD liver grafts. NMP-DCD livers showed mild epithelial injury, while SCS-DCD showed diffuse epithelial injury in extrahepatic duct and the peribiliary gland. Furthermore, Ki-67 staining revealed active cholangiocyte regeneration in NMP-DCD livers in the bile duct lumen and superficial and deep peribiliary gland, whereas Ki-67 staining was absent in SCS-DCD [72].
As a new type of perfusion, Boteon et al. demonstrated that a combined perfusion of HMP and NMP (2-hour HMP and 4-hour NMP) had 1.77 times higher ATP levels and lower tissue expression markers of oxidative injury (4-hydroxynonenal) and inflammation (CD11b, vascular cell adhesion molecule) compared with 6-hour NMP in ten human discarded livers (DCD 70%) [73] (Fig. 6.3b).
Normothermic In Situ Regional Perfusion (NRP)
Normothermic in situ regional perfusion was developed to assess the organ function in cDCD and uDCD prior to organ excision in the donor. NRP restarts blood flow to the abdominal organs after death declaration via extracorporeal membrane oxygenation (ECMO) prior to the graft cooling. Watson et al. compared NRP-DCD (n = 43) with non-NRP-DCD (n = 187) liver transplantation. The NRP-DCD group had decreased liver injury (peak ALT; 633 vs 1154 IU/L), lower early allograft dysfunction rate (3.5% vs 5.0%), and lower IC (0% vs 27%) [74]. Hessheimer et al. reported that NRP group showed significantly lower ITBLs (2% vs 13%) and lower graft loss (12% vs 24%) compared with super-rapid recovery group [75] (Fig. 6.3c).
Conclusion
IRI in liver transplantation is induced by a simultaneous activation of parenchymal and non-parenchymal cells within the liver. In liver grafts from DCD donors, the prolonged ischemia times are a crucial factor for postoperative liver function and bile duct injury. To reduce graft injury and improve post-transplant graft function, minimizing WIT and CIT is critical. In addition, novel preservation methods, such as cold and warm ex situ perfusion, as well as in situ regional perfusion, are promising approaches to improve reperfusion injury in DCD grafts. Currently, several organ perfusion settings demonstrated feasibility and improved results in transplantation with DCD grafts. It is expected that future research will result in the development of new targeted drugs for more effective protection against IRI and reconditioning of grafts from DCD donors in the future.
Abbreviations
- cDCD:
-
Controlled DCD
- DAMPs:
-
Damage-associated molecular patterns
- DCD:
-
Donation after circulatory death
- HMBG-1:
-
High-mobility group box 1
- HMP:
-
Hypothermic ex situ machine perfusion
- IC:
-
Ischemic cholangiopathy
- IRI:
-
Ischemia-reperfusion injury
- ITBLs:
-
Ischemic-type biliary lesions
- MP:
-
Machine perfusion
- NMP:
-
Normothermic ex situ machine perfusion
- NRP:
-
Normothermic regional machine perfusion
- ROS:
-
Reactive oxygen species
- SECs:
-
Liver sinusoidal endothelial cells
- TLR:
-
Toll-like receptor
- tPA:
-
Tissue plasminogen activator
- uDCD:
-
Uncontrolled DCD
References
Dar WA, Sullivan E, Bynon JS, Eltzschig H, Ju C. Ischaemia reperfusion injury in liver transplantation: cellular and molecular mechanisms. Liver Int. 2019;39(5):788–801.
Thuong M, Ruiz A, Evrard P, et al. New classification of donation after circulatory death donors definitions and terminology. Transplant Int. 2016;29(7):749–59.
Smith M, Dominguez-Gil B, Greer DM, Manara AR, Souter MJ. Organ donation after circulatory death: current status and future potential. Intensive Care Med. 2019;45(3):310–21.
Coffey JC, Wanis KN, Monbaliu D, et al. The influence of functional warm ischemia time on DCD liver transplant recipients’ outcomes. Clin Transpl 2017; 31(10). doi: 10.1111/ctr.13068.
Foley DP, Fernandez LA, Leverson G, et al. Donation after cardiac death: the University of Wisconsin experience with liver transplantation. Ann Surg. 2005;242(5):724–31.
de Vera ME, Lopez-Solis R, Dvorchik I, et al. Liver transplantation using donation after cardiac death donors: long-term follow-up from a single center. Am J Transplant. 2009;9(4):773–81.
Foley DP, Fernandez LA, Leverson G, et al. Biliary complications after liver transplantation from donation after cardiac death donors: an analysis of risk factors and long-term outcomes from a single center. Ann Surg. 2011;253(4):817–25.
Mathur AK, Heimbach J, Steffick DE, Sonnenday CJ, Goodrich NP, Merion RM. Donation after cardiac death liver transplantation: predictors of outcome. Am J Transplant. 2010;10(11):2512–9.
Hong JC, Yersiz H, Kositamongkol P, et al. Liver transplantation using organ donation after cardiac death: a clinical predictive index for graft failure-free survival. Arch Surg (Chicago, Ill: 1960). 2011;146(9):1017–23.
Abt P, Crawford M, Desai N, Markmann J, Olthoff K, Shaked A. Liver transplantation from controlled non-heart-beating donors: an increased incidence of biliary complications. Transplantation. 2003;75(10):1659–63.
Nakazato PCG, Victorino JP, Fina CF, et al. Liver ischemia and reperfusion injury. Pathophysiology and new horizons in preconditioning and therapy. Acta cirurgica brasileira. 2018;33(8):723–35.
Abu-Amara M, Yang SY, Tapuria N, Fuller B, Davidson B, Seifalian A. Liver ischemia/reperfusion injury: processes in inflammatory networks--a review. Liver Transplant. 2010;16(9):1016–32.
Brass CA, Roberts TG. Hepatic free radical production after cold storage: Kupffer cell-dependent and -independent mechanisms in rats. Gastroenterology. 1995;108(4):1167–75.
Martens JC, Keilhoff G, Halangk W, et al. Lipidomic analysis of molecular cardiolipin species in livers exposed to ischemia/reperfusion. Mol Cell Biochem. 2015;400(1–2):253–63.
Compagnon P, Levesque E, Hentati H, et al. An oxygenated and transportable machine perfusion system fully rescues liver grafts exposed to lethal ischemic damage in a pig model of DCD liver transplantation. Transplantation. 2017;101(7):e205–e13.
Imamura H, Brault A, Huet PM. Effects of extended cold preservation and transplantation on the rat liver microcirculation. Hepatology (Baltimore, Md). 1997;25(3):664–71.
Clavien PA, Harvey PR, Strasberg SM. Preservation and reperfusion injuries in liver allografts. An overview and synthesis of current studies. Transplantation. 1992;53(5):957–78.
Upadhya GA, Topp SA, Hotchkiss RS, Anagli J, Strasberg SM. Effect of cold preservation on intracellular calcium concentration and calpain activity in rat sinusoidal endothelial cells. Hepatology (Baltimore, Md). 2003;37(2):313–23.
Upadhya AG, Harvey RP, Howard TK, Lowell JA, Shenoy S, Strasberg SM. Evidence of a role for matrix metalloproteinases in cold preservation injury of the liver in humans and in the rat. Hepatology (Baltimore, Md). 1997;26(4):922–8.
Upadhya GA, Strasberg SM. Platelet adherence to isolated rat hepatic sinusoidal endothelial cells after cold preservation. Transplantation. 2002;73(11):1764–70.
Nardo B, Caraceni P, Pasini P, et al. Increased generation of reactive oxygen species in isolated rat fatty liver during postischemic reoxygenation. Transplantation. 2001;71(12):1816–20.
Tsung A, Klune JR, Zhang X, et al. HMGB1 release induced by liver ischemia involves toll-like receptor 4 dependent reactive oxygen species production and calcium-mediated signaling. J Exp Med. 2007;204(12):2913–23.
Poisson J, Lemoinne S, Boulanger C, et al. Liver sinusoidal endothelial cells: physiology and role in liver diseases. J Hepatol. 2017;66(1):212–27.
Ruan Z, Shibamoto T, Shimo T, et al. Effects of platelet-activating factor and thromboxane A2 on isolated perfused guinea pig liver. Prostaglandins Other Lipid Mediat. 2004;73(1–2):73–85.
Kageyama S, Yagi S, Tanaka H, et al. Graft reconditioning with nitric oxide gas in rat liver transplantation from cardiac death donors. Transplantation. 2014;97(6):618–25.
Massberg S, Enders G, Leiderer R, et al. Platelet-endothelial cell interactions during ischemia/reperfusion: the role of P-selectin. Blood. 1998;92(2):507–15.
Sindram D, Porte RJ, Hoffman MR, Bentley RC, Clavien PA. Platelets induce sinusoidal endothelial cell apoptosis upon reperfusion of the cold ischemic rat liver. Gastroenterology. 2000;118(1):183–91.
You Q, Cheng L, Kedl RM, Ju C. Mechanism of T cell tolerance induction by murine hepatic Kupffer cells. Hepatology (Baltimore, Md). 2008;48(3):978–90.
Toki Y, Takenouchi T, Harada H, et al. Extracellular ATP induces P2X7 receptor activation in mouse Kupffer cells, leading to release of IL-1beta, HMGB1, and PGE2, decreased MHC class I expression and necrotic cell death. Biochem Biophys Res Commun. 2015;458(4):771–6.
Witthaut R, Farhood A, Smith CW, Jaeschke H. Complement and tumor necrosis factor-alpha contribute to Mac-1 (CD11b/CD18) up-regulation and systemic neutrophil activation during endotoxemia in vivo. J Leukoc Biol. 1994;55(1):105–11.
Perry BC, Soltys D, Toledo AH, Toledo-Pereyra LH. Tumor necrosis factor-alpha in liver ischemia/reperfusion injury. J Invest Surg. 2011;24(4):178–88.
Marshall KM, He S, Zhong Z, Atkinson C, Tomlinson S. Dissecting the complement pathway in hepatic injury and regeneration with a novel protective strategy. J Exp Med. 2014;211(9):1793–805.
Diepenhorst GM, van Gulik TM, Hack CE. Complement-mediated ischemia-reperfusion injury: lessons learned from animal and clinical studies. Ann Surg. 2009;249(6):889–99.
Sarris M, Masson JB, Maurin D, et al. Inflammatory chemokines direct and restrict leukocyte migration within live tissues as glycan-bound gradients. Current biology: CB. 2012;22(24):2375–82.
Jaeschke H, Farhood A, Bautista AP, Spolarics Z, Spitzer JJ, Smith CW. Functional inactivation of neutrophils with a Mac-1 (CD11b/CD18) monoclonal antibody protects against ischemia-reperfusion injury in rat liver. Hepatology (Baltimore, Md). 1993;17(5):915–23.
Huang H, Tohme S, Al-Khafaji AB, et al. Damage-associated molecular pattern-activated neutrophil extracellular trap exacerbates sterile inflammatory liver injury. Hepatology (Baltimore, Md). 2015;62(2):600–14.
Bamboat ZM, Balachandran VP, Ocuin LM, Obaid H, Plitas G, DeMatteo RP. Toll-like receptor 9 inhibition confers protection from liver ischemia-reperfusion injury. Hepatology (Baltimore, Md). 2010;51(2):621–32.
Tsung A, Sahai R, Tanaka H, et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med. 2005;201(7):1135–43.
Oliveira THC, Marques PE, Proost P, Teixeira MMM. Neutrophils: a cornerstone of liver ischemia and reperfusion injury. Lab Invest. 2018;98(1):51–62.
Weller A, Isenmann S, Vestweber D. Cloning of the mouse endothelial selectins. Expression of both E- and P-selectin is inducible by tumor necrosis factor alpha. J Biol Chem. 1992;267(21):15176–83.
Enjyoji K, Sevigny J, Lin Y, et al. Targeted disruption of cd39/ATP diphosphohydrolase results in disordered hemostasis and thromboregulation. Nat Med. 1999;5(9):1010–7.
Miyashita T, Nakanuma S, Ahmed AK, et al. Ischemia reperfusion-facilitated sinusoidal endothelial cell injury in liver transplantation and the resulting impact of extravasated platelet aggregation. Eur Surg: ACA: Acta chirurgica Austriaca. 2016;48:92–8.
Watanabe K, Togo S, Takahashi T, et al. PAI-1 plays an important role in liver failure after excessive hepatectomy in the rat. J Surg Res. 2007;143(1):13–9.
Ueda S, Yamanoi A, Hishikawa Y, Dhar DK, Tachibana M, Nagasue N. Transforming growth factor-beta1 released from the spleen exerts a growth inhibitory effect on liver regeneration in rats. Lab Invest. 2003;83(11):1595–603.
Paterno F, Guarrera JV, Wima K, et al. Clinical implications of donor warm and cold ischemia time in donor after circulatory death liver transplantation. Liver Transpl. 2019 Sep;25(9):1342–52.
Abt PL, Desai NM, Crawford MD, et al. Survival following liver transplantation from non-heart-beating donors. Ann Surg. 2004;239(1):87–92.
Skaro AI, Jay CL, Baker TB, et al. The impact of ischemic cholangiopathy in liver transplantation using donors after cardiac death: the untold story. Surgery. 2009;146(4):543–52; discussion 52–3
Meurisse N, Vanden Bussche S, Jochmans I, et al. Outcomes of liver transplantations using donations after circulatory death: a single-center experience. Transplant Proc. 2012;44(9):2868–73.
Kubal C, Mangus R, Fridell J, et al. Optimization of perioperative conditions to prevent ischemic cholangiopathy in donation after circulatory death donor liver transplantation. Transplantation. 2016;100(8):1699–704.
Dikdan GS, Mora-Esteves C, Koneru B. Review of randomized clinical trials of donor management and organ preservation in deceased donors: opportunities and issues. Transplantation. 2012;94(5):425–41.
Maheshwari A, Maley W, Li Z, Thuluvath PJ. Biliary complications and outcomes of liver transplantation from donors after cardiac death. Liver Transplant. 2007;13(12):1645–53.
Taner CB, Bulatao IG, Willingham DL, et al. Events in procurement as risk factors for ischemic cholangiopathy in liver transplantation using donation after cardiac death donors. Liver Transplant. 2012;18(1):100–11.
Jayant K, Reccia I, Virdis F, Shapiro AMJ. Systematic review and meta-analysis on the impact of thrombolytic therapy in liver transplantation following donation after circulatory death. J Clin Med. 2018;7(11):pii: E425.
op den Dries S, Westerkamp AC, Karimian N, et al. Injury to peribiliary glands and vascular plexus before liver transplantation predicts formation of non-anastomotic biliary strictures. J Hepatol. 2014;60(6):1172–9.
Hashimoto K, Eghtesad B, Gunasekaran G, et al. Use of tissue plasminogen activator in liver transplantation from donation after cardiac death donors. Am J Transplant. 2010;10(12):2665–72.
Chan EY, Olson LC, Kisthard JA, et al. Ischemic cholangiopathy following liver transplantation from donation after cardiac death donors. Liver Transplant. 2008;14(5):604–10.
de Meijer VE, Fujiyoshi M, Porte RJ. Ex situ machine perfusion strategies in liver transplantation. J Hepatol. 2019;70(1):203–5.
Schlegel A, de Rougemont O, Graf R, Clavien PA, Dutkowski P. Protective mechanisms of end-ischemic cold machine perfusion in DCD liver grafts. J Hepatol. 2013;58(2):278–86.
Schlegel A, Muller X, Kalisvaart M, et al. Outcomes of DCD liver transplantation using organs treated by hypothermic oxygenated perfusion before implantation. J Hepatol. 2019;70(1):50–7.
Schlegel A, Graf R, Clavien PA, Dutkowski P. Hypothermic oxygenated perfusion (HOPE) protects from biliary injury in a rodent model of DCD liver transplantation. J Hepatol. 2013;59(5):984–91.
van Rijn R, Karimian N, Matton APM, et al. Dual hypothermic oxygenated machine perfusion in liver transplants donated after circulatory death. Br J Surg. 2017;104(7):907–17.
van Rijn R, van Leeuwen OB, Matton APM, et al. Hypothermic oxygenated machine perfusion reduces bile duct reperfusion injury after transplantation of donation after circulatory death livers. Liver Transplant. 2018;24(5):655–64.
Watson CJE, Kosmoliaptsis V, Pley C, et al. Observations on the ex situ perfusion of livers for transplantation. AM J Transplant. 2018;18(8):2005–20.
Mergental H, Stephenson BTF, Laing RW, et al. Development of clinical criteria for functional assessment to predict primary nonfunction of high-risk livers using normothermic machine perfusion. Liver Transplant. 2018;24(10):1453–69.
Bral M, Gala-Lopez B, Bigam D, et al. Preliminary single-center Canadian experience of human normothermic ex vivo liver perfusion: results of a clinical trial. Am J Transplant. 2017;17(4):1071–80.
Ravikumar R, Jassem W, Mergental H, et al. Liver transplantation after ex vivo normothermic machine preservation: a phase 1 (first-in-man) clinical trial. Am J Transplant. 2016;16(6):1779–87.
Selzner M, Goldaracena N, Echeverri J, et al. Normothermic ex vivo liver perfusion using steen solution as perfusate for human liver transplantation: first North American results. Liver Transplant. 2016;22(11):1501–8.
Xu H, Berendsen T, Kim K, et al. Excorporeal normothermic machine perfusion resuscitates pig DCD livers with extended warm ischemia. J Surg Res. 2012;173(2):e83–8.
Brockmann J, Reddy S, Coussios C, et al. Normothermic perfusion: a new paradigm for organ preservation. Ann Surg. 2009;250(1):1–6.
Jassem W, Xystrakis E, Ghnewa YG, et al. Normothermic Machine Perfusion (NMP) inhibits proinflammatory responses in the liver and promotes regeneration. Hepatology (Baltimore, Md). 2019;70(2):682–95.
Nasralla D, Coussios CC, Mergental H, et al. A randomized trial of normothermic preservation in liver transplantation. Nature. 2018;557(7703):50–6.
Liu Q, Nassar A, Farias K, et al. Sanguineous normothermic machine perfusion improves hemodynamics and biliary epithelial regeneration in donation after cardiac death porcine livers. Liver Transplantation. 2014;20(8):987–99.
Boteon YL, Laing RW, Schlegel A, et al. Combined hypothermic and normothermic machine perfusion improves functional recovery of extended criteria donor livers. Liver Transpl. 2018 Dec;24(12):1699–715.
Watson CJE, Hunt F, Messer S, et al. In situ normothermic perfusion of livers in controlled circulatory death donation may prevent ischemic cholangiopathy and improve graft survival. Am J Transplant. 2019;19(6):1745–58.
Hessheimer AJ, Coll E, Torres F, et al. Normothermic regional perfusion vs. super-rapid recovery in controlled donation after circulatory death liver transplantation. J Hepatol. 2019;70(4):658–65.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Goto, T., Selzner, M. (2020). Ischemia-Reperfusion Injury and Therapeutic Strategy in Donation After Circulatory Death Liver Transplantation. In: Croome, K., Muiesan, P., Taner, C. (eds) Donation after Circulatory Death (DCD) Liver Transplantation. Springer, Cham. https://doi.org/10.1007/978-3-030-46470-7_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-46470-7_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-46469-1
Online ISBN: 978-3-030-46470-7
eBook Packages: MedicineMedicine (R0)