
Abstract
The high content of heavy metals (e.g., Cd, Zn, Cu, Co, Cr, Ni, Hg, and Pb) in the soil has harmful effects on plant development and growth and crop yield. The remediation of these pollutants from the environment especially from the contaminated soil and water is an important task for environmentalists and agricultural scientists in order to make our precious soil and water resources clean and reusable for our agricultural industry and for safe human consumption. It is important to develop new and less expensive technologies to remediate contaminated areas and eliminate the potentially deleterious effects of these contaminants which may pose serious problems to our food quality and health if not addressed properly. Phytoremediation, an in situ remediation technique, could be one of the best approaches through which toxic metals can be extracted from the polluted soils and waters. However, it is a relatively new technology, and there are not much data available in order to understand the molecular mechanism of uptake, transport, and accumulation of the toxic metals/metalloids in the harvestable parts of the plants. Recent research has shown great advances in the understanding of key mechanisms responsible for heavy metal detoxification, transport from root to shoot and accumulation in the shoots and leaves of the plants. For example, genes of metal transporters, metal ATPases, micro RNAs involved in translocation, and accumulation process have been identified and their role in metal accumulation is being elucidated to some extent. The aim of this review is to provide a close understanding of the molecular mechanisms of heavy metal-induced plant stress response and tolerance based on the current findings related to molecular biology and biotechnology research. Additionally, recent work on metal stress–related microRNAs and genome editing will be highlighted.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The increase in industrialization activities in the developed and developing countries has ultimately increased the global threat to the environment that can cause an unprecedented imbalances in the natural ecosystem. For example, industrial activities like mining, leather tanning industry, textile factories, petrol chemical industry, intensive farming, and other manufacturing industries are the main sources for polluting the environment (Bhargava et al. 2012). Each source of contamination has its own damaging effects to plants, animals, and ultimately human health, but those that add heavy metals to soils and waters are of serious concern due to their persistence in the environment and carcinogenicity to human beings.
The contamination of agricultural soil by heavy metals has become a critical environmental concern due to their potential adverse ecological effects. Such toxic elements are considered as soil pollutants due to their widespread occurrence and their acute and chronic toxic effect on plants grown of such soils (Chaney et al. 2018). Heavy metals are defined as metals with a density higher than 5 gcm−3. Around 43 of 90 naturally occurring elements are heavy metals (Cevher-Keskin et al. 2019; Weast et al. 1988), but only some of them have biological importance (Marschner 2011). Based on their solubility under physiological conditions, 17 heavy metals can be absorbed by living cells and could be important for organisms and ecosystems (Weast et al. 1988). Among these metals, Fe, Mn, Zn, Cu, and Mo are important as micronutrients. V, Co, W, and Cr are toxic elements with high or low importance as trace elements. As, Hg, Ag, Sb, Cd, Pb, and U have no known function as nutrients and seem to be more or less toxic to plants and microorganisms (Weast et al. 1988; Memon et al. 2008). The metal contamination caused by the human activities is far greater than the natural process. For example, it was shown that around 13 times more Cu, 15 times more Cd, 21 times more Zn, and 100 times more Pb were released in the atmosphere by human activities than by natural processes (Campbell et al. 1983). Several laborious and expensive methods have been used to clean up these kinds of contaminants, but no optimal results were obtained. Presently, phytoremediation is being used as an alternative, cost-effective, and environmental friendly method for removing the metal pollutants from contaminated soil and water (van der Ent et al. 2018). Numerous accumulator plant species have been identified to absorb and remove toxic elements, such as cadmium, chromium, lead, arsenic, and variety of radionuclides, from the soils successfully. Phytoextraction , which is one of the category of phytoremediation technology, could be used to extract and remove majority of the toxic metals with unknown biological function (e.g., Cd, Cr, Pb, Co, Ag, Se, Hg) from the contaminated soils (Chaney and Baklanov 2017; Memon et al. 2001).
An interesting breakthrough that has emerged from the comparative physiological and molecular analyses of hyperaccumulators and related nonhyperaccumulators is that most key steps of hyperaccumulation rely on different regulations and gene expressions found in both kinds of plants. In particular, a determinant role in driving the uptake and translocation of heavy metals to leaves and finally sequestration in vacuoles or cell walls is one of the important part of detoxification mechanism in hyperaccumulator plants (Memon et al. 2001; Memon and Yatazawa 1982). The constitutive overexpression of genes encoding transmembrane transporters, such as members of ZIPs, HMAs, MATE, YSL, and MTPs families, has been reported in many of these accumulator plants (Memon 2016; Memon and Schröder 2009).
Recently, phytoremediation researchers have discovered that Indian mustard (Brassica juncea (L.) Czern, AABB genome) can accumulate high levels of metals, including Zn, Cd, Pb, and Se, and B. nigra (BB genome) has been shown a superb Cu accumulator (Cevher-Keskin et al. 2019; Dalyan et al. 2017; Memon and Zahirovic 2014). The metal-accumulating ability of these plants, coupled with the potential to rapidly produce large quantities of shoot biomass, makes these plants ideal for phytoextraction (Memon 2016), and these plant species have received huge attention in phytoremediation field.
2 Phytoremediation
Phytoremediation is an emerging cleanup technology defined as the use of plants to remove or contain or render harmless the contaminants such as toxic metals and organic and radioactive compounds from soils, sediments, and water (Chaney and Baklanov 2017). This technology is environmental friendly and potentially cost-effective. This green technology is unique in way that it takes the advantage of selective uptake capabilities of plant root systems, together with the translocation, bioaccumulation, and contaminant degradation abilities of the entire plant system (Lasat 2002).
However, the ability of plant to accumulate heavy metals varies significantly between species and among varieties within species, as different mechanisms of ion uptake and translocation are operative in each species, based on their genetic, morphological, physiological, and anatomical characteristics. Phytoremediation can be divided into several different categories, such as phytoextraction, phytofiltration, phytostabilization, phytovolatilization, and phytodegradation, depending on the remediation mechanisms (Ali et al. 2013; Memon et al. 2001).
Phytoextraction is also referred as phytoaccumulation, and it takes the approach to remove contaminants from soil without destroying the soil structure and other physical and chemical properties of the soil. The metal ion accumulated in the aerial parts of the plant can be removed to dispose or burnt to recover metals.
Phytofiltration is defined as the use of plants, both land and aquatic, to remove the contaminants from aqueous wastes.
Phytostabilization is generally used to remove the pollutants from the soil by absorbing the pollutants (e.g., toxic metals) through plant roots and keeping them in the rhizosphere, rendering them harmless by preventing them from leaching.
Phytovolatilization involves the use of plants to remove the contaminants from the soil, for example, mercury, selenium, and arsenic, by transforming them into volatile form and volatilize them into the atmosphere.
Phytodegradation is the breakdown of organics pollutants by the use of plants and associated microorganisms (Alkorta and Garbisu 2001). Most of the phytoremediation technologies could be used simultaneously, but the metal removal from the soil depends on its bioavailable form in the soil. The efficient use of phytotechnology depends on the plant species, for example, some plants may have one function, whereas others can have multiple functions of phytoremediation (Lone et al. 2008).
Phytoremediation is considered as environmentally friendly and cost-effective technology for cleaning up the contaminated soils. The phytoextraction is the most common and profitable technique mainly used for extracting heavy metals and radioactive elements from the soil (van der Ent et al. 2018).
There are several important components of phytoextraction which can play an important role in remediating the contaminated soils efficiently, which are as follows: appropriate accumulator plant species, metal availability in the soil and uptake by roots, metal translocation from roots to shoots, and plant tolerance to toxic metals. There are several plant species which are currently proposed for phytoextraction, such as Arabidopsis halleri, Noccaea caerulescens, and Alyssum sps (A. murale, A. lesbiacum, and A. tenium), which can absorb and accumulate high concentration of Zn and Cd in their shoots. However, the remediation capacity of these plant species is limited because of their slow growth and low biomass. Currently, a number of plant species as metal accumulators have been identified within the Brassicaceae family (Pollard et al. 2014). To understand the genomics of these metal accumulator plants, the vast genetic resources and bioinformatics data developed in model plant A. thaliana could be extended to the newly identified metal accumulator species that display traits absent in this model species (Reeves et al. 2017). For developing the new-generation phytoremediation technologies , the data of the multidisciplinary research including high-throughput sequencing, all available -omics data, genetic engineering, plant–microbe interaction, and agricultural and environmental engineering tools could be integrated and be applied for practical purpose in the contaminated areas (Chaney and Baklanov 2017; Memon and Schröder 2009).
3 Plants as Accumulators of Heavy Metals
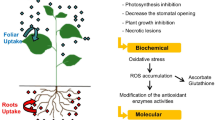
Plant requires at least six major elements (N, P, K, Ca, Mg, and S) and eight minor elements (Cu, Zn, Mn, Fe, B, Cl, Ni, and Mo) for their growth and development. Plant roots take up the elements selectively, with some being preferentially acquired over others (Memon et al. 2008). The uptake and accumulation of heavy metals by plant roots depend on an interrelated network of physiological and molecular mechanisms, such as (i) binding of the metals to extracellular exudates and cell wall components; (ii) movement of the metals from cytoplasm to the vacuolar compartment; (iii) complexation of the metal with various compounds in the cell, such as several amino acids, organic acids, small molecular weight proteins like metallothioneins, and small metal-binding peptides like phytochelatins; (iv) heavy metal-induced antioxidative enzymes; and (v) modification of plant metabolism and quick repair and recover of damaged cell structures (Paunov et al. 2018).
Baker and co-workers (Baker et al. 2000), while working on the vegetation of metalliferous soils, have classified plants into three categories: (i) excluder plants, which prevent the uptake of toxic metals into root cells and keep the metal level in their shoots very low (De Vos and Schat 1991). Excluders can be used in the polluted soils for stabilization and to prevent further spread of contamination due to erosion (Lasat 2002). (ii) Accumulator plants can accumulate huge amount of metals in their aboveground parts when grown at either low or high metal soil concentrations. Accumulators have high metal uptake rate in the roots and can efficiently transport them in the shoots and have high metal accumulation capacity (Pollard et al. 2014). (iii) In indicator plants, internal metal concentration reflects the external levels (McGrath et al. 2002)..
4 Hyperaccumulators
Plants with exceptional metal-accumulating capacity are known as hyperaccumulator plants. The hyperaccumulator plants, which are naturally growing in metal-rich habitats, can accumulate 100- to 1000-fold higher levels of metals than normal plants, since these plants take up two or three orders more of metals from the soil than plant species growing on uncontaminated soils (Lone et al. 2008). Accumulator species are naturally capable of accumulating heavy metals in their aboveground tissues, without developing any toxicity symptoms. They are often endemic to naturally mineralized soils and can accumulate metals at different level depending on metal species. For example, they can accumulate to a level of 0.1% (on leaf dry weight basis) for Ni, Co, Cr, Cu, Al, and Pb; at 1% level for Zn and Mn; and at 0.01% level for Cd and Se (Baker and Brooks 1989; Baker et al. 2000).
Considerable research work has been carried out in identifying the accumulator plant species and their mechanisms of metal uptake and hyperaccumulation. The field of phytotechnology has been revolutionized and got high momentum after the discovery of hyperaccumulator plant species since these plants have excellent capacity to absorb and accumulate metals at levels 50–500 times greater than average plants (Lasat 2000). Hyperaccumulator plants which are generally restricted to metalliferous soils and accumulate metals in their aboveground parts especially in the leaves are classified as “obligate” hyperaccumulators (Reeves et al. 2017). The other type of hyperaccumulator plants which can accumulate metal from both metalliferous and nonmetalliferous soils is classified as “facultative” hyperaccumulators (Pollard et al. 2014). This latter category includes many plant species which can accumulate metal not only from ultramafic soils but also from metal polluted soils. For example, Biscutella laevigata accumulates >1% thallium (Babst-Kostecka et al. 2016), Pteris vittata with up to 2.3% arsenic (Ma et al. 2001), and Phytolacca americana accumulates >1% manganese (Xu et al. 2009).
Around 450 plant species from 45 angiosperms families have been reported as metal hyperaccumulators, which include the members of the Asteraceae, Brassicaceae, Caryophyllaceae, Cyperaceae, Cunoniaceae, Fabaceae, Flacourtiaceae, Lamiaceae, Poaceae, Violaceae, and Euphorbiaceae (Padmavathiamma and Li 2007; Reeves et al. 2017). Interestingly, a large number of accumulator plant species are reported to be in Brassicaceae family especially in the genera Alyssum and Noccaea, wherein accumulation of more than one metal has been reported (Reeves and Baker 2000; Vamerali et al. 2010; Vara Prasad and de Oliveira Freitas 2003; Verbruggen et al. 2009). Pteris vittata (Chinese brake fern) is reported to accumulate up to 95% of the arsenic in its fronds (Ma et al. 2001; Zhang et al. 2002). Noccaea caerulescens (pennycress) is a well-known metal hyperaccumulator which can accumulate large amounts of Zn (39,600 mg/kg) and Cd (1800 mg/kg) without any apparent damage (Basic et al. 2006; Hanikenne and Nouet 2011; Rascio and Navari-Izzo 2011). This diploid and self-pollinating plant can be easily grown under laboratory conditions and is an excellent experimental material for studying the mechanisms of metal uptake, accumulation, and tolerance. Apart from N. caerulescens , Brassica juncea and B. nigra have also been used as a model system to investigate the physiology and biochemistry of metal accumulation in plants especially for Zn, Cd, and Cu (Cevher-Keskin et al. 2019; Memon et al. 2001). These crop plants with high biomass production could be excellent candidates for phytoremediation in coming years.
Memon and co-workers reported several Mn accumulator plant species, which accumulated huge amount of Mn in their leaves. For example, Acanthopanax sciadophylloides Frach. & Sav. (Mn: 4600 ppm), Ilex crenata Thunb. var. paludosa (Mn:1155 ppm), and Clethra barbinervis Sieb. & Zucc. (Mn: 1374 ppm) accumulated very high amount of Mn in their leaves while naturally growing in the forest soil containing very low metal. These plants are possibly facultative Mn accumulators and can grow in both nonmetalliferous and metalliferous soils (Memon et al. 1979; Memon and Yatazawa 1982). Among these Mn accumulator plant species, Acanthopanax sciadophylloides was found to be superb Mn accumulator, achieving >1% Mn in their leaves when growing on soils with only background concentrations of this element (Memon et al. 1979; Memon and Yatazawa 1982). Similarly, Noccaea caerulescens and Arabidopsis halleri were also reported to be facultative Zn accumulators, achieving >1% foliar Zn when growing in soils containing very low Zn concentration (Reeves et al. 2001; Stein et al. 2017). Majority of the accumulator plants are reported to be grown in ultramafic soils and hyperaccumulate Ni, and some of them accumulate Ni, Co, and Mn (Reeves et al. 2018). Many obligate and facultative hyperaccumulator plants have reported to hyperaccumulate several toxic elements, for example, Cd, Cu, Co, Mn, Pb, Zn, Se, Tl, and some rare earth elements (Reeves et al. 2017).
The efficiency of the cleanup of metal contaminated soils by the use of accumulator plants depends on their biomass production as well as on their bioconcentration factor (BCF ratio of metal concentration in the shoot tissue to the soil) (McGrath and Zhao 2003). The BCF varies with plant species and is determined by the capacity of the roots to absorb, accumulate, store, and detoxify metals while maintaining metabolism, growth, and biomass production (Clemens et al. 2002; Gleba et al. 1999; Guerinot and Salt 2001). Hyperaccumulators have a bioconcentration factor greater than 1, sometimes reaching as high as 50–100 (Reeves et al. 2018). The metal bioconcentration factors in nonaccumulator plants is less than 1, which means that they are not able to reduce soil contamination by 50% for longer time (time longer than human lifespan) (Peuke and Rennenberg 2005). Multiple mechanisms are involved for metal tolerance in accumulator plants, such as high cell wall–binding capacity, active transport of metal ions into the vacuolar compartment, and formation of complexes with organic acids, metallothioneins, and/or chelation with phytochelatins (Memon et al. 2001; Memon and Schröder 2009).
There is general consensus that metal hyperaccumulation is an evolutionary adaptation by specific plants which can survive and live in naturally metal-rich habitats that confer on them high metal tolerance qualities, drought tolerance, and protection against herbivores or pathogens (Reeves et al. 2018). The most accepted hypothesis is that the hyperaccumulation character protects the plant against pathogens and herbivores (Boyd and Banzhaf 2007; Galeas et al. 2008; Huitson and Macnair 2003; Martens and Boyd 1994; Noret et al. 2007). However, the detailed mechanisms of metal uptake and tolerances need to be worked out in coming years.
Notable efforts were carried out to assemble a database for hyperaccumulator plants. For example, Environment Canada’s PHYTOREM database and the METALS (metal-accumulating plants) database originally maintained by the Environmental Consultancy, University of Sheffield (ECUS Ltd., Sheffield, UK), were created. The problem in these databases is that not only accumulators were recorded but also other plant species grown in metalliferous soils were included. (Reeves et al. 2017). To circumvent this problem, a new database was established which was confined only with hyperaccumulator plants, and currently, around 700 pint species have been recorded and will eventually increase to 1000–1500 species. The Global Hyperaccumulator Database (www.hyperaccumulators.org) went online in 2015 under the administration of the Center for Mined Land Rehabilitation of the University of Queensland, Brisbane, Australia. This new database gives the information about all known metal and metalloid hyperaccumulator plant species and contains detailed information about the taxonomy, distribution, ecology, collection records, analytical data, and other useful studies related to these species (Reeves et al. 2017). This database is continuously updated and is freely available to all researchers.
5 Brassicaceae
The Brassicaceae (order Brassicales) is the largest family in Brassicales, commonly known as the mustard family, and is composed of approximately 338 genera and 3700 species. Genus Brassica contains around 100 species, including important oilseed crops (e.g., B. napus, B. juncea) and many common vegetable plants such as broccoli, cauliflower, cabbage, Brussels sprouts, radishes, turnip, and various gardening plants (wallflower, sweet alyssum, rock cress, etc.) (Ozturk et al. 2012; Warwick and Black 1991). The Brassica species are closely related to the model plant Arabidopsis thaliana, and their chromosome numbers vary from 2n = 10 to 2n = 38 (Lysak et al. 2005). Some of the Brassica species are diploid (e.g., B. rapa, B. nigra, B. oleracea), and some of them are allotetraploid (e.g., B. napus, B. juncea, B. carinata). The genome of B. rapa is the smallest, at ca. 529 Mb, and B. napus is the largest one, at ca 1132 Mb, in Brassica species represented in the “Triangle U” (Memon and Zahirovic 2014). The genome of both plants has been sequenced, and both sequences and sequence annotations are available in the public domain: http://www.brassicagenome.net/databases.php; https://www.ncbi.nlm.nih.gov/genome/?term=brassica%20napus (Memon 2016; Wang et al. 2016). The annotated Arabidopsis genome sequence can be exploited as a tool for carrying out the comparative analysis of Arabidopsis and Brassica genomes, for example, metal accumulator and tolerant species, B. napus and B. juncea, respectively. The biology of both Arabidopsis and Brassica is similar, and the comparative genetic mapping between species of Brassicaceae showed collinear blocks even though the species differed with respect to genome size, base chromosome number, and ploidy. Around 80–90% homology was found between the exons of putative orthologous genes in Arabidopsis and Brassica, and this clearly indicates that the knowledge from Arabidopsis is highly relevant for gene isolation and characterization in Brassica crops (Ozturk et al. 2012).
6 Brassica Species
The plant species in Brassica are the major source of vegetable oil in the world after palm and soybean oil. These plant species produce high biomass and also accumulate and tolerate high amount of metals in their tissues (Anjum et al. 2013; Kumar et al. 1995). There are six agronomical important species of Brassica which are commercially used for both oil and vegetable production, and among them, three are diploids (B. nigra, B. oleracea, and B. campestris) and the other three are amphidiploids (B. juncea, B. napus, and B. carinata). The genomic variation and relationships between these species have been described in the form of triangle, and in the literature, it is known as triangle of U (Nagaharu 1935; Ozturk et al. 2012). Furthermore, an extensive research work on metal tolerance and accumulation was carried out, and the differential metal accumulation pattern among Brassica species has been observed (Anjum et al. 2013; Diwan et al. 2010).
Several Brassica species have been reported to exhibit higher tolerance toward most of the toxic metals/metalloids, and majority of Brassica species are now known as good accumulators of toxic metals (including Cd, Cu, Ni, Pb, U, Zn) (Kumar et al. 1995; Ozturk et al. 2012), allocating large amounts of majority of these metals into aboveground parts, thus stand second to none in terms of their utility in toxic metal-remediation strategies. Moreover, as several species of Brassica are able to produce significant amounts of biomass (a required trait for phytoremediation) and are adaptable to a range of environmental conditions, there is the potential to develop superior genotypes of Brassica sps for phytoremediation through selection and breeding techniques (Anjum et al. 2012).
Brassica juncea needs special attention in the field of phytoremediation because it is not only a metal tolerant but also accumulates huge amount of several metals in its shoots, including Zn, Cd, and Pb. It is shown that B. juncea accumulates high amount of Cd in the shoots (1450 μg Cd/g dry wt), which is three times more than reported in Brassica napus (555 μg/g dry wt). In addition, this plant also extracts high amount of other metals from soil, such as Pb (28% reduction) and Se (reduced between 13% and 48%) (Szczygłowska et al. 2014). Interestingly, this plant is more effective at removing Zn from soil than Noccaea caerulescens, a known hyperaccumulator of zinc . This is due to the fact that B. juncea produces ten times more biomass than N. caerulescens (Anjum et al. 2013; Szczygłowska et al. 2014). In Southeastern Anatolia, several endemic metal accumulator was discovered, and among them, Brassica nigra Diyarbakir ecotype was found to be Cu accumulator (Cevher-Keskin et al. 2019). When this ecotype was regenerated from callus culture and grown in soil containing 500 uM Cu, the shoots accumulated around 20,000 ug g−1 DW Cu, which was around three times more than in the roots (Memon and Zahirovic 2014; Ozturk et al. 2012).
The high metal accumulation capacity of several plant species from the Brassicaceae family indicates that these accumulator plants especially B. juncea and B. nigra could be important candidates for phytoremediation of Zn, Cu, Cd, and Pd from the contaminated soils (Cevher-Keskin et al. 2019; Dalyan et al. 2017; Kumar et al. 2012; Memon and Zahirovic 2014).
7 Heavy Metal Uptake, Accumulation, and Subcellular Localization
Highly specific and very efficient mechanisms have been developed by plants in order to take up essential micronutrients from the soil, even when present at low quantities. Plant-induced pH changes, redox reactions, and plant-produced chelating agents in the rhizosphere help plants to absorb trace elements even from poor nutrient soils and translocate and store them in vacuole and other organelles. The same mechanism is also involved in the uptake, translocation, and storage of toxic elements, whose chemical properties simulate those of essential elements. Thus, the metal uptake, translocation, and accumulation mechanisms are of much interest in the area of phytoremediation (Memon and Schröder 2009; Tangahu et al. 2011).
B. nigra is shown to be a Cu accumulator (Memon et al. 2008), and our results with microarray analysis showed that many genes especially metal ATPases and other metal transporters were several hundred fold upregulated in the shoots of B. nigra Diyarbakir ecotype when plants were subjected to 500 uM Cu concentration (Memon and Zahirovic 2014). This Diyarbakır ecotype was named as facultative metallophyte because it can grow both at low metal and at very high metal concentration (e.g., 500 uM Cu) in the soil. We used Arabidopsis ATH1 genome array to analyze the gene expression pattern in our facultative accumulator B. nigra Diyarbakir ecotype. The gene expression pattern in the roots and shoots of B. nigra was compared when grown at 0 and 25 uM Cu. The differential response in terms of gene expression in roots and shoots was observed when plants were subjected to 25 uM Cu concentration for 72 h. The response to Cu was much stronger in roots (88 genes showed increased or decreased mRNA levels) than in leaf tissues (24 were up- or downregulated). These genes were identified as metal transporters, signal transduction and metabolism-related genes, and transport facilitation genes. Glutathione pathway-related genes (γ-ECS, PC, etc.) were also identified, and their mRNAs were differentially expressed in root and shoot tissues (Cevher-Keskin et al. 2019).
The range of known transport mechanisms or specialized proteins embedded in the plant cell plasma membrane involved in ion uptake and translocation include (1) proton pumps ATPases that consume energy and generate electrochemical gradients and (2) co- and anti-transporters (proteins that use the electrochemical gradients generated by H+-ATPases to drive the active uptake of ions) and channels (proteins that facilitate the transport of ions into the cell). Each transport mechanism is likely to take up a range of ions. Plant uptake-translocation mechanisms are presumably closely regulated (Tangahu et al. 2011). Most of the crop plants require small amount of micronutrients for their metabolism and normal growth, which generally vary from 10 to 15 ppm. Some other plants which are labelled as “hyperaccumulators” can take up toxic metals at levels in the thousands of ppm without showing any toxicity symptoms. It is intriguing that how these accumulator plants cope with metal toxicity and how they avoid metal toxicity. What are the mechanisms responsible to make these metals in innocuous form? Multiple mechanisms are involved, especially the storage in the vacuole appears to be one of the major mechanism for metal detoxification (Memon et al. 2001; Reeves et al. 2018; Tangahu et al. 2011).
Clarkson and Lüttge (1989) reported that Cu and Zn, Ni and Cd compete for the same membrane carriers. Different chelators are reported to be involved in the translocation of metal cations through the xylem, such as organic acid chelators (e.g., malate, citrate, histidine (Salt et al. 1995; von Wirén et al. 1999), or nicotinamide (Stephan et al. 1996; von Wirén et al. 1999). Since the metal is complexed within a chelate, it can be translocated upward in the xylem without being adsorbed by the high cation exchange capacity of the xylem (von Wirén et al. 1999).
Several metal tolerant and accumulation mechanisms in plants growing in metal contaminated soils have been suggested (Memon 2016; Memon and Schröder 2009). Hyperaccumulation in plants is a part of metal homeostasis network in which metal is efficiently taken by the roots and then transported from the roots to shoots through xylem and then either complexed and sequestered in the subcellular compartments or secreted in the trichomes (Hanikenne and Nouet 2011; Memon and Schröder 2009; Ovečka and Takáč 2014). These accumulator plants have very unique eco-physiological character and have the ability to uptake, transport, and accumulate huge amount of metals in their shoots and leaves and compartmentalize them in the cell wall, vacuole, and other subcompartments in the cytosol in order to keep them away from metabolic activities in the cell (Memon and Schröder 2009; Memon and Yatazawa 1982). To determine the subcellular localization on Mn in the leaves of Acanthopanax sciadophylloides, electroprobe X-ray microprobe analysis was carried out with fresh leaves frozen down in liquid nitrogen. The microdistribution pattern of Mn at subcellular level showed that most of the Mn was deposited in the cell wall and vacuolar compartment of epidermal cells and it was kept away from metabolically active compartments, for example, cytosol, mitochondria, and chloroplast (Fig. 1; Memon and Yatazawa 1984). Interestingly, Mn was predominantly accumulated in the epidermal cells (E) and was almost absent from the cells of stomatal complex (Fig. 2, A. R. Memon, Unpublished data).

Secondary electron image (SEI) showing a line scan profile of a leaf section of tea plant with Mn (Kα radiation) peaks. cy, cytosol; V, vacuole of epidermal cells (E); P, palisade parenchyma cells

Secondary electron (SEI) and Mn X-ray distribution images of the abaxial side of a tea leaf. A, SEI. B, Mn X-ray distribution image. EC, epidermal complex; SC, stomatal complex; S, spongy parenchyma; V, vascular bundle; P, parenchyma cells
The metal distribution and accumulation pattern vary with plant species and the type of element. For example, A. halleri leaves accumulates Zn and Cd more in the mesophyll cells than in the epidermis (Küpper et al. 2000), whereas N. caerulescens accumulates six-fold higher Zn and Cd in epidermis cells than in mesophyll cells (Küpper et al. 1999). Interestingly, B. juncea which is reported to be metal tolerant and accumulator (Dalyan et al. 2017) and is a good candidate for phytoremediation is reported to accumulate 40-fold higher Cd in trichomes compared to the total leaf (Salt et al. 1995). Other hyperaccumulator species like Alyssum lesbiacum accumulate a significant portion of Zn and Ni in leaf trichomes (Reeves et al. 2018).
Cell fractionation analysis with A. sciadophylloides Frach & Sav leaves showed that most of the Mn was present in cell wall and in supernatant, and a very large amount of Mn in the supernatant fraction was found to be bound with organic acid having a molecular weight of approximately 145 g/mol (Memon and Yatazawa 1984). In order to identify the chemical forms of accumulated Mn in the cell, a high-performance liquid chromatography and high-voltage paper electrophoresis analysis were carried with the leaf extracts, and results showed that Mn was chelated with oxalic acid in vacuolar compartment (Memon and Yatazawa 1984). The detoxification mechanism of Mn proposed that Mn2+ is taken up at plasma membrane and binds with malate in cytoplasm, and this Mn-malate complex is transported through the tonoplast membrane into the vacuoles, where Mn dissociates from malate and complexes with oxalate. Here, malate functions as “transport vehicle” through the cytoplasm and oxalate as the “terminal acceptor” in the vacuole (Memon and Schröder 2009; Memon and Yatazawa 1984). Many other mechanisms have been proposed for detoxification and accumulation depending on the metal type and plant species (Isaure et al. 2015; Memon 2016; Rascio and Navari-Izzo 2011). Phytochelatins, metallothioneins, metalloenzymes, metal-activated enzymes, and various channel proteins have been reported to bind and sequester metals (Hanikenne and Nouet 2011; Isaure et al. 2015; Memon 2016).
Most of the current research on genetic, genomic, and transcriptome analysis of transporters and the participation of multiple gene families in response to metal stress have been obtained in a two-model accumulator plant species such as Noccaea caerulescens and Arabidopsis halleri (Verbruggen et al. 2009). The recent development of next-generation sequencing technologies, transcriptomics (microarray analysis, RNA-seq analysis), proteomics, and metabolomics opens up the new avenue to understand the function and regulation of genes, proteins, and metabolites in the cell when encountered high metal concentration in the environment (Verbruggen et al. 2013).
8 Heavy Metal Transporters in Plants
Recently, molecular biology and genetics studies with hyperaccumulators plants have identified several important gene families involved in metal transport and could play a key role in metal tolerance. So far, several classes of metal transporter proteins have been identified in plants, and they fulfil many important functions ranging from metal absorption, transport, sequestration, and storage in specific organelles. They also play an important role in metal homeostasis in plant cell (Memon 2016). Metal transporters are classified into six main groups, which include natural resistant-associated macrophage protein (NRAMP), ZRT/ RT-like protein (ZIP), cation diffusion facilitator (CDF) transporters, yellow stripe-like (YSL) proteins, and P1B-type heavy metal ATPases (HMAs) (Memon 2016; Merlot et al. 2018). Plants grown in polluted or metalliferous soils have evolved several mechanisms in order to deal with toxic quantities of heavy metals, such as Cd, Ni, Cr, Pb, or Hg, or toxic excess of essential elements, such as Zn, Cu, Mn, B, and Co. A fine control of metal homeostasis is required to overcome the toxic and oxidative damage at subcellular level. Thus, it is not surprising that plant can activate several transporters which can function either in excluding metal at the root or in sequestering them at subcellular level in some compartments such as vacuole. Arabidopsis thaliana genome has uncovered several families of transporter genes, which include 15 members of zinc and iron transporters (ZIP), 8 members of cation diffusion facilitator (CDF), 6 members of copper transporters (CTR), 6 members of NRAMP homologues, and 8 members of Cu, Zn/Cd transporting ATPases (Mäser et al. 2001; Merlot et al. 2018) (http://www.cbs.umn.edu/arabidopsis/). In addition, some other transporter families, for example, vacuolar cation proton exchanger (CAX) and ABC transporters, are also involved in metal homeostasis in plant cells (Colangelo and Guerinot 2006; Hall and Williams 2003; Memon 2016; Memon and Schröder 2009; Sarma et al. 2018).
8.1 ZIP Gene Family
ZIP transporters are involved in the transport of four essential micronutrients: Zn, Fe, Mn, and Cu (Cohen et al. 2004; Eide et al. 1996; Grotz et al. 1998; Lin et al. 2009; Pedas et al. 2008; Pence et al. 2000; Wintz et al. 2003). This family is derived from the first defined members that include ZRT- (Zn-regulated carrier) and IRT- (Fe-regulated carrier) like proteins. It has been reported that ZIP family members can also transport heavy metals such as Cd (Zheng et al. 2018). Therefore, the ZIP family can play an important role in the transport of various heavy metals, both essential and nonessential (toxic) (Guerinot 2000; Pence et al. 2000; Rogers et al. 2000). They have been discovered in both dicots and monocots, including Arabidopsis, Medicago (Milner et al. 2013; Stephan et al. 1996; Stephens et al. 2011), rice, maize, and barley (Chen et al. 2008; Li et al. 2013; Stephens et al. 2011; Tiong et al. 2015). Arabidopsis has 15 members of ZIP transporters, but recently, 18 ZIPs from Arabidopsis and 16 ZIPs from rice has been annotated (Ivanov and Bauer 2017). To date, only three members of the ZIP family (AtIRT gene) have been isolated from Arabidopsis. The role and functions of Arabidopsis AtIRT1, AtIRT2, and AtIRT3 have been described in detail (Grotz et al. 1998; Guerinot 2000). Little is known about the functionality of the other 12 ZIP members in Arabidopsis. Recently, a distinct expression pattern of ZIP genes in Arabidopsis and rice was found in response to Cd stress (Zheng et al. 2018). ZIP genes in Arabidopsis were mainly upregulated in roots, while in rice, they were upregulated in shoots when subjected to Cd stress. Most of the characterized ZIP proteins are reported to be targeted to plasma membrane; however, some of them are found to be located in tonoplast or other endomembrane system (Milner et al. 2013; Ricachenevsky and Sperotto 2016). NcZNT1 is an important ZIP-like transporter involved in Zn and Cd hyperaccumulation and tolerance in N. caerulescens (Lin et al. 2016). It is orthologue of AtZIP4 from A. thaliana having 90% cDNA and 87% amino acid identity (Lin et al. 2014). NcZNt1 is differentially expressed between A. thaliana and N. caerulescens due to its differences in cis- and trans-regulatory elements. It is constitutively expressed in the stele of the roots of N. caerulescens and is responsible for xylem-mediated translocation of metals to the shoot (Lin et al. 2016).
8.2 CDF-Type Family
The cation diffusion facilitator (CDF) proteins play an important role in metal homeostasis and tolerance (Mäser et al. 2001). CDFs are membrane bound proteins and transport zinc and other heavy metal ions. They are involved in metal tolerance/resistance by efflux of ions and share a two-modular architecture consisting of a transmembrane domain (TMD) and C-terminal (Ricachenevsky et al. 2013) domain (CTD) that protrude into the cytoplasm (Kolaj-Robin et al. 2015). The first CDF gene in plants is zinc carrier ZAT1 characterized in A. thaliana. It was then renamed to metal tolerance protein 1 (MTP1) (Ricachenevsky et al. 2013). ZAT is expressed primarily throughout the plant, and its expression is enhanced by increase in Zn concentration. These carriers are known to play an important role in tolerating Zn2+, Cd2+, or Co2+ ions in plants, yeast, bacteria and animals (Memon 2016).
8.3 COPT Gene Family
High-affinity Cu carriers (CTRs, COPTs in other organisms other than plants) are essential components of the major pathway for cellular high-affinity Cu uptake. Six members of the putative Cu family (COPT1-6) have been identified in Arabidopsis, and three of them COPT1, COPT2 and COPT6 are localized in plasma membrane (Peñarrubia et al. 2010; Sancenón et al. 2003; Yuan et al. 2011). COPT1 is required in Cu acquisition when Cu availability decreases in the soil (Sancenón et al. 2004; Yamasaki et al. 2009), whereas COPY2 is a cell surface transporter and expressed mostly in all plant parts, especially in roots, young leaves, apical meristems, trichomes, and anthers (Perea-García et al. 2013). COPT5 is reported to be localized in both the tonoplast membrane (Klaumann et al. 2011) and the prevacuolar compartment (Perea-García et al. 2013) and is believed to be involved in intracellular homeostasis. The plasma membrane-localized COPT6 functions in Cu distribution in plants and is expressed in seeds and leaves during Cu deficiency (Perea-García et al. 2013). In Cu deficiency, the increase in transcript levels of COPT1 and COPT2 is activated by the SPL7 transcription factor (Bernal et al. 2012; Perea-García et al. 2013). CTR/COPT family Cu transporters have also been identified in rice (Yuan et al. 2011).
8.4 Nramp Gene Family
Nramp gene family defines a new family of proteins involved in the transport of divalent metal ions. This gene family is highly conserved during evolution and is involved in the transport of metal ions in a wide range of living organisms, including bacteria, yeasts, insects, mammals, and also higher plants (Cellier 2012; Gruenheid et al. 1995). It encodes integral membrane polypeptide containing 10–12 transmembrane domains, 1–2 extracellular loops, and an intracellular domain with structural characteristics of a transporter protein (Bairoch 1993).
NRAMP genes play an important role in the transport several metal ions including Fe, Mn, Zn, and Cd (Nevo and Nelson 2006). In plants, this family was first described in rice, and three Nramp genes (OsNramp 1–3) were reported (Belouchi et al. 1997; Cellier et al. 1995; Hall and Williams 2003). OsNramp3 is expressed in both roots and shoots, whereas OsNramp2 is expressed only in the leaves (Belouchi et al. 1997). In Arabidopsis thaliana , six Nramp genes have been identified and classified into two subfamilies. AtNRAMP1 and AtNRAMP6 comprise the first group, and AtNRAMP2 to AtNRAMP5 belong to the second (Mäser et al. 2001). AtNRAMP1 is localized to both the plasma membrane and intracellular membranes including the Golgi apparatus (Agorio et al. 2017; Cailliatte et al. 2010) and can also transport iron (Fe), manganese (Mn), and cadmium (Cd). AtNRAMP2 is located in trans-Golgi network and is activated in plant roots when subjected to low Mn concentration (Gao et al. 2018). AtNRAMP3 and AtNRAMP4 are required for Mn transport from vacuolar compartment to chloroplast and are localized to the tonoplast membrane. Double mutant of these transporters reduced the function of photosystem II and impaired plant growth (Lanquar et al. 2010). OsNramp5 is involved in Mn uptake and is localized in the plasma membrane of rice roots (Ishimaru et al. 2012). In soybean genome, 13 NRAMP genes have been identified (Qin et al. 2017). Gene expression analysis showed that GmNRAMP is differentially regulated by deficiencies of major elements in the cell like N, P, K, Fe, S and also regulated by the toxicities of Fe, Cu, Cd, and Mn (Qin et al. 2017). Brassica napus genome contains 22 NRAMP transporter genes, and based on sequence identity, these transporters are classified into six subfamilies. Nineteen NRAMP transporters were confirmed by RNA-seq analysis, and 10 NRAMP genes were differentially expressed under Cd exposure (Meng et al. 2017). BnNRAMP1b was constitutively expressed throughout all developmental stages of B. napus plant and can be strongly induced in seedlings exposed to high Cd concentration. BnNRAMP1b was found to be cleaved by miR167, suggesting that it is posttranscriptionally regulated by miR167 in B. napus under Cd stress (Meng et al. 2017). A new rice line with extremely low Cd content has been developed by knocking out OsNramp5 transporter using the CRISPR/cas9 technique (Tang et al. 2017). This transgenic line can grow in Cd-contaminated paddy soils and accumulate very low Cd in their grain (0.05 mg/kg); in contrast, wild-type India rice had high Cd concentration in their grain ranging from 0.33 to 2.90 mg/kg when grown in the same soil (Tang et al. 2017). Excessive Cd in rice grain is a serious problem to health (e.g., itai-itai disease) of those who consume rice as a staple food. These mutant rice lines with very low Cd content in their grains could be useful material to develop new hybrid lines, which could be commercially used in the Cd-contaminated areas, and their grains can be consumed for dietary purpose.
8.5 ABC-Type Family
ATP-binding cassette (or ABC) proteins constitute one of the largest proteins families and are present in all organisms ranging from bacteria to humans (Henikoff et al. 1997). ABC transporters are ATP-driven pumps comprising two nucleotide-binding domains (NBDs) and two transmembrane domains (TMDs). Three structural type ABC transporters have been reported. One is full-type transporter containing two membrane domains (TMD) and two nucleotide-binding domains (NBD) and largely expressed as TMD–NBD–TMD–NBD or NBD–TMD–NBD–TMD. The second one is called as half transporters containing one TMD and one NBD domain and dimerizes in pairs to form virtual homodimers or heterodimers transporters. The third-type transporter contains two NBDs but no TMDs (Verrier et al. 2008). The NBD domain in all structural type of ABC transporters contains some conserved motifs, which are as follows: Walker A, Q-loop, Walker B, D-loop, switch H-loop, and a signature motif (LSGGQ). Each motif has its specific function, for example, the D-loop functions as holding dimers together, the switch H-loop interacts with transmembrane domain, the P-loop (Walker A and B motifs together) binds ATP, the other two loops Q- and H- are important for interaction with the γ-phosphate of the ATP (Davidson et al. 2008; Lane et al. 2016). The signature motif (LSGGQ) is specific to ABC transporter proteins and distinguishes them from ATPases (Davies and Coleman 2000).
In humans , seven to eight subfamilies of ABC transporters have been identified based on their primary sequence and domain structure (Dean and Allikmets 2001). In Arabidopsis and in rice, 130 and 133 ABC transporters, respectively, have been identified (Hwang et al. 2016; Lefèvre et al. 2015). Among them, only a few numbers of transporters have been functionally characterized, and a limited number of transporters have also been identified in other plant species, for example, wheat, maize, and Vitis vinifera (Bhati et al. 2015; Cakir et al. 2013; Kang et al. 2011a; Pang et al. 2013).
Recently, 314 ABC transporters in Brassica napus genome have been identified and are classified into eight subfamilies from A-G and I. The ABCG transporters constitute the largest subfamily with 116 members followed by ABCB (69 members) and ABCC (47 members) (Kang et al. 2011b; Lane et al. 2016; Yan et al. 2017; Yang et al. 2016), and the ABCB and ABCC subfamilies rank second and third with 69 and 47 members, respectively. Most of the ABC genes were validated by RNA sequencing. Among 132 genes which were differentially expressed, 84 genes were significantly expressed by Cd stress (Yan et al. 2017). Recently, AtABCC1 and AtABCC2 have been implicated in phytochelatin-mediated Cd and Hg detoxification by vacuolar sequestration (Park et al. 2012). Furthermore, OsABCB14 is shown to be responsible for Fe homeostasis in rice (Xu et al. 2014), and OsABCG43/PDR5 is induced in rice roots when subjected to Cd stress and may be involved in detoxification of Cd by compartmentalizing Cd into organelles (Oda et al. 2011; Xu et al. 2014). Above studies demonstrate that ABC transporters play a central role in detoxification and accumulation of toxic metals in hyperaccumulator/tolerant plants .
9 P1B-Type Metal ATPases (HMAs)
P1B-type metal ATPases are a subfamily of P-type ATPases and are responsible for the transport of a number of important and potentially toxic metals (Cu2+, Zn2+, Cd2+, Pb2+) along the cell membranes (Argüello 2003; Axelsen and Palmgren 2001; Østerberg and Palmgren 2018; Rensing et al. 1999). These ion pumps use active ATP energy to pump various charged substrates along biological membranes and share a common enzymatic mechanism in which ATP hydrolysis is carried out for the transport of ions from the membrane (Axelsen and Palmgren 2001). These ATPases can be classified into several subfamilies according to both DNA sequence and functional protein similarities: for example, H+-ATPases (type 3A) in plants and fungi, Na+/K+-ATPases in animals, Ca2+-ATPases (type 2C/D) and heavy metal ATPases (type 1B) in bacteria, plants, and mammalian system (Axelsen and Palmgren 2001; Rosenzweig and Argüello 2012). P1B-ATPases , in addition to the conserved regions in the P-type ATPases, such as DKTGT, GDGxNDxP, PxxK, and S/TGE, possess six to eight transmembrane domains (TMs), a HP locus, and a CPx/SPC motif (Williams and Mills 2005) necessary for metal transport, and putative metal-binding sites in the N and/ or C terminus (Williams and Mills 2005). These metal ATPases based on their substrate specificity are dived into two subgroups, Cu/Ag (Cu+-ATPases) and Zn/Cd/Co/Pb transporters (Zn2+-ATPases) (Axelsen and Palmgren 2001). Arabidopsis thaliana and rice genomes possess eight and nine P1B-ATPase genes, respectively, which have been classified into six groups (Williams and Mills 2005). Among these ATPases, HMA2, HMA3, and HMA4 are closely related in sequence and are specific to Zn/Cd transporters. HMA2 and HMA4 are located in the plasma membrane of pericycle and control root to shoot long-distance transport of Zn and Cd, while HMA3 is involved in vacuolar transport of Zn and Cd (Hanikenne et al. 2008; Hussain et al. 2004; Liu et al. 2017; Morel et al. 2009; Wong and Cobbett 2009). Lekeux et al. (2018) have reported that C terminus of HMA4 plays an important role in Zn binding, and this region of HMA4 substantially diverges between A. thaliana (nonaccumulator) and A. halleri (accumulator). Their data further showed that di-Cys motifs in C-terminal region of HMA4 contribute to high-affinity zinc binding in plants. In B. juncea, BjHMMA4 was upregulated by Zn and Cd in the roots, stems, and leaves (Wang et al. 2019). A detailed study with BjHMA4 protein showed that a repeat region named BjHMA4R in the C-terminal region of HMA4 is not far from the last transmembrane domain and is localized in the cytosol. This C-terminal region substantially binds Cd2+ and improves Cd tolerance and accumulation in B. juncea. AtHMA1 is located in chloroplast membrane and is involved in the translocation of Cu and Zn into and out of chloroplast, respectively (Boutigny et al. 2014; Kim et al. 2009; Moreno et al. 2008; Seigneurin-Berny et al. 2006). Recently, it has been shown that HMA1 in the leaves of Sedum plumbizincicola plays an important role in the detoxification of Cd in chloroplast by exporting Cd out of the chloroplast (Zhao et al. 2018). SpHMA1RNA interference transgenic plants and CRISPR/Cas9-induced HMA1 mutant lines showed significant increase in Cd accumulation in the chloroplasts compared with wild-type plants. Their data showed that SpHMA1 in S. plumbizincicola leaves is a chloroplast Cd exporter and protects photosynthesis by inhibiting the Cd accumulation in the chloroplast. The AtHMA5 to AtHMA8 ATPases are involved in Cu transport in the cell. AtHMA5 contributes in the detoxification of excess Cu in roots by increasing Cu translocation from roots to shoots (Kobayashi et al. 2008). AtHMA6 (PAA1) and AtHMA8 (PAA2) are localized in chloroplast envelope and thylakoids and deliver Cu into chloroplast (Abdel-Ghany et al. 2005; Shikanai et al. 2003). Recently, 20 HMA genes have been found in soybean and are annotated as GmHMA1 to GmHMA20 (Fang et al. 2016). Phylogenetically, these 20 HMAs were divided into six clusters. Based on the Arabidopsis and rice HMA function, six GmHMAs (5, 19,13,16,14 and 18) were classified as Zn2+-ATPases, while the remaining HMAs were classified as Cu+-ATPases (Fang et al. 2016). The expression pattern of these 20 genes in both wild and cultivated soybean lines suggests that they may be functionally conserved and divergent and possibly involved in Cd detoxification (Fang et al. 2016). Li et al. (2015) performed genome-wide of the Populus trichocarpa HMA gene family and identified 17 HMA genes, which were showed to be differentially regulated by excessive metal stress. Tissue-specific expression of HMA genes showed that HMA1 and HMA4 were highly expressed in the leaves of populous, whereas HMA 5-HMA8 genes were upregulated in roots when plants were subjected to high metal concentration (Li et al. 2015). Given the main functions of HMA proteins in metal transport in A. thaliana and other plants in Brassicaceae , we performed phylogeny analysis, multiple sequence alignments, 3D structure prediction, and validation, as well as subcellular localization prediction of these proteins, with the goal of investigating the interacting proteins present in different plant species in Brassicaceae. The main approach of generating interactome protein analysis is to check for the specificity in structure and function between B. rapa (genome is recently published) and A. halleri (a metal accumulator plant) and then compare to genomic data of A. thaliana (a model plant). Study on protein–protein interactions is important in order to understand the complexity of the function of HMA proteins. Detailed bioinformatics and comparative structural studies with HMA1–HMA4 in Arabidopsis thaliana, A. halleri, B. rapa, and B. juncea have shown the similarities and dissimilarities in the structural component of these transporters and their interaction with other proteins. The differences have been seen in the domain analysis and subcellular localization of these proteins among plant species (Jusovic and Memon 2015, Unpublished data, see Table 1 and Fig. 3).

Protein–protein interaction prediction, where the interaction of other proteins with HMA1, HMA2, HMA3, and HMA4 in Arabidopsis thaliana is shown. The interactome is generated by STRING software. Stronger protein associations are represented by thicker lines
(String v9.1 software was used for interaction analysis; Franceschini et al. 2013) (Maida and Memon, Unpublished data)
9.1 Interactome Analysis Between HMAs and Other Metal Transporters
The interactome analysis revealed the strong interactions of HMA4 with HMA1, HMA2, and HMA3 and also with other metal transporters like different ZIP (Guerinot 2000) and ZAT (van der Zaal et al. 1999), whereas all HMA proteins showed strong interactions with ATCCS proteins (copper chaperone for superoxide dismutase) (Chu et al. 2005). The interaction of HMAs with different metal transporters and also with other several heavy metal transporter domain-containing proteins (Fig. 1) suggests that the metal tolerance mechanism in the plants is a function of the expression of many transporter genes and the subcellular localization of the different transporter proteins in accumulator plants. The detailed description of these transporters is shown in Table 1.
All previous bioinformatics analyses have given some information about their regulation, but not much work has been done on their structural analysis and their interactome. We have here clearly shown the HMA interaction with other proteins, and some structural differences have also been identified in our bioinformatics analysis (data not shown). These differences are seen in the domain analysis and subcellular localizations of these proteins. Experimental determination of 3D structures is important to better understand the function of these proteins, which is crucial for proper functioning of all cellular processes in plants. Especially, docking sites and domains need to be experimentally researched further in order to understand the function and role of these proteins in accumulator plants.
A significant advance in metal-induced gene expression and the role of different metal transporters in metal uptake transport, accumulation, and detoxification have been carried out in metal accumulator and nonaccumulator plant species. Genomic analysis of well-known metal accumulator species (e.g., Arabidopsis halleri, Noccaea caerulescens, Brassica juncea, B. napus, B. nigra) has shown the role of several metal transporters, including metal ATPases in metal accumulation and tolerances in plants (Cevher-Keskin et al. 2019; Dalyan et al. 2017; Memon 2016).
Above data show that P1B-ATPase family plays an important role in heavy metal transport in plants. HMA genes have not only been identified in model plants like Arabidopsis thaliana and rice, but these HMAs have also been identified in B. napus, B. rapa, B. juncea, Glycine max, and Populus trichocarpa. The HMA1 to HMA4 in the model plants have been thoroughly characterized at both gene and protein levels. The role of HMA1 to HMA4 in Cu, Zn, and Cd transport in plat cell has been well documented, and especially, the importance of C terminus of HMA4 in metal binding and its role in metal tolerance and accumulation in plants have been suggested. Functional studies of these transporters have been carried out in yeast, and extremely useful information related to the transporter proteins and metal interaction has been obtained (Fang et al. 2016; Wang et al. 2019).
Furthermore, the plant genomic sequencing and bioinformatics analysis with Arabidopsis, rice, Brassica napus, and B. rapa have shown us that a large number of transporter gene families exist in the genomes of these plants. In the last decades, a range of plant transporter genes involved in metal uptake and translocation have been cloned and characterized, but there are many other transporter genes which are still not identified and await for identification and functional analysis. The future work should be focused on the expression and function of these transporter genes at cellular and subcellular levels in order to find out their specific roles in metal localization and compartmentalization in the cell. More studies with the structural analysis of the transporter proteins will reveal the basic mechanism of the substrates selection and transport activity.
From the information obtained from functional studies of these transporters together with their structural analyses, it is possible to assign their role in metal transport and accumulation at specific site and location of the cell and tissues. Two different kinds of strategies can be carried out with these transporters and their expression in plants. For accumulator plants, the transporters related to metal uptake and translocation can be overexpressed to optimize the translocation of toxic metals to aerial parts, which would be the target for phytoremediation. Another strategy could be used for edible crop plants where the low uptake transporters could be engineered to minimize the transport of toxic cation in edible crops.
10 Recent Advances and New Possibilities in Phytoremediation
microRNAs (miRNAs) are universal regulator for gene expression in both plants and mammalian system when encountered to biotic or abiotic stress. Recently, a high-throughput sequence analysis with metal accumulator and tolerant plants has shown the upregulation and downregulation of several miRNAs when plants were exposed to metals (e.g., Zn, Cd, Cu) (Memon 2016). Several studies have demonstrated that heavy metal-induced gene expression in the accumulator plants can be regulated by different miRNAs (Gielen et al. 2012; Memon 2016; Zhou et al. 2012). Genome wide identification of miRNAs and corresponding target genes that are induced in response to metal stress in A. thaliana, rice, tobacco, radish, B. napus, and B. rapa have been documented to some extent (Liu et al. 2015; Yuan et al. 2011). However, these studies are limited and need further attention in terms of miRNA-regulated heavy metal-related gene expression in hyperaccumulator plants and their impact on metal accumulation and tolerance in the cell.
Recently, several gene editing technologies have been developed such as customized homing nuclease (meganuclease), zinc-finger nuclease (ZNFs), and transcription activator-like effector nuclease (TALENs) (Zhu et al. 2017). There are several advantages and disadvantages associated with these techniques. Recently, CRISPR-Cas 9 technology (clustered regularly interspaced short palindromic repeats(CRISPR)-associated protein 9) has been successfully used for genome editing in bacteria, plants, mammalian, and human system (Barrangou and Doudna 2016; Doudna and Charpentier 2014; Park et al. 2017). This new technology has great potential to help molecular biologists and crop breeders to edit or modify the genome or genes of crop plants that are able to produce high yields under conditions of biotic/abiotic stress (Kumlehn et al. 2018; Scheben et al. 2017; Schindele et al. 2018; Wolter and Puchta 2018). CRISPR/Cas9 technique could be used to introduce point mutations in the sequences of the metal transporter genes in order to enhance their activity or modify their selectivity toward metal of interest. The increase in transporter activities (overexpression) will enhance the ability of the plant to uptake and translocate the metals in shoots efficiently, and this could be paramount importance for phytoremediation. In other case, the transporter activities could be reduced or knocked out by deleting the sequences of specific transporter gene/genes. In this case, the crop plants could be generated, which could be used in contaminated soils for human consumption. For example, OsNramp5, which is major transporter responsible for Mn and Cd uptake in rice (Sasaki et al. 2014), was successfully knockout in the rice by using CRISPR/Cas9 system. These OsNramp5 mutants showed a remarkable reduction in Cd content in rice grain while maintaining all important agronomical traits such as growth, yield, and test (Tang et al. 2017). Overall, this CRISPR/Cas9-mediated genome editing has a great potential for exploiting plant genomes to enhance the metal accumulation and metal tolerance capacities in plants which could be effectively used for phytoremediation.
11 Conclusion
In this review, I have summarized the recent advances in the field of phytoremediation. The obligate and facultative hyperaccumulator species have been identified, and their role in metal accumulation has been described. The metal distribution and localization in the leaf cells of hyperaccumulator plants have been analyzed by X-ray microprobe analyzer, and their distribution pattern at subcellular level has been examined. The global hyperaccumulator database (www.hyperaccumulators.org) has been recently established by the Center for Mined Land Rehabilitation of The University of Queensland, Australia. The database is freely available and contains information about taxonomy, worldwide distribution, ecology, analytical data, and other relevant information of all known metal and metalloid hyperaccumulator plant species, which could be useful for biologists, environmentalists, and molecular biologists for their research. Recent developments in the genome analysis of B. rapa and B. napus have shown the expression of many metal transporters in these plant species when subjected to high metal concentration in the soil. These data have been compared with the data of well-known dicot plant species Arabidopsis thaliana and monocot species Oryza sativa. Recent development in functional analysis of metal transporters including ZIPs, ZIPs, MTPs, NRAMPs, ABC transporters, and HMAs in accumulator plants has been summarized. Interactome analyses of the different HMAs and their interaction with other metal transporters indicate that the hyperaccumulation is fascinating phenomenon controlled by multigene-regulated process. Furthermore, the role of the transporters in metal accumulation and detoxification in hyperaccumulator plants and their implication in phytoremediation have been discussed.
References
Abdel-Ghany SE, Müller-Moulé P, Niyogi KK, Pilon M, Shikanai T (2005) Two P-type ATPases are required for copper delivery in Arabidopsis thaliana chloroplasts. Plant Cell 17:1233–1251
Agorio A et al (2017) Phosphatidylinositol 3-phosphate–binding protein AtPH1 controls the localization of the metal transporter NRAMP1 in Arabidopsis. PNAS 114:E3354–E3363
Ali H, Khan E, Sajad MA (2013) Phytoremediation of heavy metals—concepts and applications. Chemosphere 91:869–881
Alkorta I, Garbisu C (2001) Phytoremediation of organic contaminants in soils. Bioresour Technol 79:273–276
Anjum NA, Ahmad I, Pereira ME, Duarte AC, Umar S, Khan NA (2012) The plant family Brassicaceae: contribution towards phytoremediation, vol 21. Springer, Dordrecht
Anjum NA, Gill SS, Duarte AC, Pereira E, Ahmad I (2013) Silver nanoparticles in soil–plant systems. J Nanopart Res 15:1896
Argüello JM (2003) Identification of ion-selectivity determinants in heavy-metal transport P 1B-type ATPases. J Membr Biol 195:93–108
Axelsen KB, Palmgren MG (2001) Inventory of the superfamily of P-type ion pumps in Arabidopsis. Plant Physiol 126:696–706
Babst-Kostecka AA, Waldmann P, Frérot H, Vollenweider P (2016) Plant adaptation to metal polluted environments—physiological, morphological, and evolutionary insights from Biscutella laevigata. Environ Exp Bot 127:1–13
Bairoch A (1993) The PROSITE dictionary of sites and patterns in proteins, its current status. Nucleic Acids Res 21:3097
Baker AJM, McGrawth SP, Reeves RD, Smith JAC (2000). Metal hyperaccumulator plants: a review of the ecology and physiology of a biological resources for phytoremediation of metal-polluted soils. In: Phytoremediation of contaminated soil and water (Terry, N, Banuelos, G, eds) CRC press, Lewis Publishers, USA. ISBN-13:978-1566704502, pp. 85–107.
Baker A, Brooks RR (1989) Terrestrial higher plants which hyperaccumulate metallic elements. A review of their distribution, ecology and phytochemistry. Biorecovery 1:81–126
Barrangou R, Doudna JA (2016) Applications of CRISPR technologies in research and beyond. Nat Biotechnol 34:933–941. https://doi.org/10.1038/nbt.3659
Basic N, Salamin N, Keller C, Galland N, Besnard G (2006) Cadmium hyperaccumulation and genetic differentiation of Thlaspi caerulescens populations. Biochem Syst Ecol 34:667–677
Belouchi A, Kwan T, Gros P (1997) Cloning and characterization of the OsNramp family from Oryza sativa, a new family of membrane proteins possibly implicated in the transport of metal ions. Plant Mol Biol 33:1085–1092
Bernal M et al (2012) Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis. Plant Cell 24:738–761
Bhargava A, Carmona FF, Bhargava M, Srivastava S (2012) Approaches for enhanced phytoextraction of heavy metals. J Environ Manag 105:103–120
Bhati KK et al (2015) Genome-wide identification and expression characterization of ABCC-MRP transporters in hexaploid wheat. Front Plant Sci 6:488. https://doi.org/10.3389/fpls.2015.00488
Boutigny S et al (2014) HMA1 and PAA1, two chloroplast-envelope PIB-ATPases, play distinct roles in chloroplast copper homeostasis. J Exp Bot 65:1529–1540. https://doi.org/10.1093/jxb/eru020
Boyd J, Banzhaf S (2007) What are ecosystem services? The need for standardized environmental accounting units. Ecol Econ 63:616–626
Cailliatte R, Schikora A, Briat J-F, Mari S, Curie C (2010) High-affinity manganese uptake by the metal transporter NRAMP1 is essential for Arabidopsis growth in low manganese conditions. Plant Cell 22:904–917
Cakir B, Kiliçkaya O, Olcay AC (2013) Genome-wide analysis of Aux/IAA genes in Vitis vinifera: cloning and expression profiling of a grape Aux/IAA gene in response to phytohormone and abiotic stresses. Acta Physiol Plant 35:365–377
Campbell P, Stokes P, Galloway J (1983) Effects of atmospheric deposition on the geochemical cycling and biological availability of metals. Heavy Metals Environ 2:760–763
Cellier MF (2012) Nramp: from sequence to structure and mechanism of divalent metal import. In: Current topics in membranes, vol 69. Elsevier, pp 249–293
Cellier M, Prive G, Belouchi A, Kwan T, Rodrigues V, Chia W, Gros P (1995) Nramp defines a family of membrane proteins. Proc Nat Acad Sci U S A 92:10089–10093
Cevher-Keskin B, Yıldızhan Y, Yüksel B, Dalyan E, Memon AR (2019) Characterization of differentially expressed genes to Cu stress in Brassica nigra by Arabidopsis genome arrays. Environ Sci Pollut Res 26:299–311
Chaney RL, Baklanov IA (2017) Phytoremediation and phytomining: status and promise. In: Advances in botanical research, vol 83. Elsevier, pp 189–221
Chaney RL, Baker AJ, Morel JL (2018) The long road to developing agromining/phytomining. In: Agromining: farming for metals. Springer, Cham, pp 1–17
Chen J-Q, Meng X-P, Zhang Y, Xia M, Wang X-P (2008) Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol Lett 30:2191–2198
Chu C-C, Lee W-C, Guo W-Y, Pan S-M, Chen L-J, Li H-m, Jinn T-L (2005) A copper chaperone for superoxide dismutase that confers three types of copper/zinc superoxide dismutase activity in Arabidopsis. Plant Physiol 139:425–436
Clarkson DT, Lüttge U (1989) Mineral nutrition: divalent cations, transport and compartmentation. In: Progress in botany. Springer, Berlin, pp 93–112
Clemens S, Palmgren MG, Krämer U (2002) A long way ahead: understanding and engineering plant metal accumulation. Trends Plant Sci 7:309–315
Cohen CK, Garvin DF, Kochian LV (2004) Kinetic properties of a micronutrient transporter from Pisum sativum indicate a primary function in Fe uptake from the soil. Planta 218:784–792
Colangelo EP, Guerinot ML (2006) Put the metal to the petal: metal uptake and transport throughout plants. Curr Opin Plant Biol 9:322–330
Dalyan E et al (2017) The identification of genes associated with Pb and Cd response mechanism in Brassica juncea L. by using Arabidopsis expression array. Environ Exp Bot 139:105–115
Davidson AL, Dassa E, Orelle C, Chen J (2008) Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol Mol Biol Rev 72:317–364. , table of contents. https://doi.org/10.1128/MMBR.00031-07
Davies T, Coleman J (2000) The Arabidopsis thaliana ATP-binding cassette proteins: an emerging superfamily. Plant Cell Environ 23:431–443
De Vos C, Schat H (1991) Free radicals and heavy metal tolerance. In: Ecological responses to environmental stresses. Springer, Dordrecht, pp 22–31
Dean M, Allikmets R (2001) Complete characterization of the human ABC gene family. J Bioenerg Biomembr 33:475–479
Diwan H, Khan I, Ahmad A, Iqbal M (2010) Induction of phytochelatins and antioxidant defence system in Brassica juncea and Vigna radiata in response to chromium treatments. Plant Growth Regul 61:97–107
Doudna JA, Charpentier E (2014) The new frontier of genome engineering with CRISPR-Cas9. Science 346:1258096
Eide D, Broderius M, Fett J, Guerinot ML (1996) A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc Nat Acad Sci U S A 93:5624–5628
Fang X et al (2016) Genome-wide characterization of soybean P 1B-ATPases gene family provides functional implications in cadmium responses. BMC Genomics 17:376
Franceschini A et al (2013) STRING v9.1: protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res 41:D808–D815. https://doi.org/10.1093/nar/gks1094
Galeas ML, Klamper EM, Bennett LE, Freeman JL, Kondratieff BC, Quinn CF, Pilon-Smits EA (2008) Selenium hyperaccumulation reduces plant arthropod loads in the field. New Phytol 177:715–724
Gao H et al (2018) NRAMP2, a trans-Golgi network-localized manganese transporter, is required for Arabidopsis root growth under manganese deficiency. New Phytol 217:179–193
Gielen H, Remans T, Vangronsveld J, Cuypers A (2012) MicroRNAs in metal stress: specific roles or secondary responses? Int J Mol Sci 13:15826–15847. https://doi.org/10.3390/ijms131215826
Gleba D et al (1999) Use of plant roots for phytoremediation and molecular farming. Proc Nat Acad Sci U S A 96:5973–5977
Grotz N, Fox T, Connolly E, Park W, Guerinot ML, Eide D (1998) Identification of a family of zinc transporter genes from Arabidopsis that respond to zinc deficiency. Proc Nat Acad Sci U S A 95:7220–7224
Gruenheid S, Cellier M, Vidal S, Gros P (1995) Identification and characterization of a second mouse Nramp gene. Genomics 25:514–525
Guerinot ML (2000) The ZIP family of metal transporters. Biochim Biophys Acta Biomembr 1465:190–198
Guerinot ML, Salt DE (2001) Fortified foods and phytoremediation. Two sides of the same coin. Plant Physiol 125:164–167
Hall J, Williams LE (2003) Transition metal transporters in plants. J Exp Bot 54:2601–2613
Hanikenne M, Nouet C (2011) Metal hyperaccumulation and hypertolerance: a model for plant evolutionary genomics. Curr Opin Plant Biol 14:252–259
Hanikenne M et al (2008) Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature 453:391
Henikoff S, Greene EA, Pietrokovski S, Bork P, Attwood TK, Hood L (1997) Gene families: the taxonomy of protein paralogs and chimeras. Science 278:609–614
Huitson SB, Macnair MR (2003) Does zinc protect the zinc hyperaccumulator Arabidopsis halleri from herbivory by snails? New Phytol 159:453–459
Hussain D et al (2004) P-type ATPase heavy metal transporters with roles in essential zinc homeostasis in Arabidopsis. Plant Cell 16:1327–1339. https://doi.org/10.1105/tpc.020487
Hwang JU et al (2016) Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol Plant 9:338–355. https://doi.org/10.1016/j.molp.2016.02.003
Isaure M-P et al (2015) Evidence of various mechanisms of Cd sequestration in the hyperaccumulator Arabidopsis halleri, the non-accumulator Arabidopsis lyrata, and their progenies by combined synchrotron-based techniques. J Exp Bot 66:3201–3214
Ishimaru Y et al (2012) Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci Rep 2:286. https://doi.org/10.1038/srep00286
Ivanov R, Bauer P (2017) Sequence and coexpression analysis of iron-regulated ZIP transporter genes reveals crossing points between iron acquisition strategies in green algae and land plants. Plant Soil 418:61–73
Kang J, Park J, Choi H, Burla B, Kretzschmar T, Lee Y, Martinoia E (2011a) Plant ABC transporters vol 9. Arabidopsis Book, 2012/02/04 edn. https://doi.org/10.1199/tab.0153
Kang J, Park J, Choi H, Burla B, Kretzschmar T, Lee Y, Martinoia E (2011b) Plant ABC transporters. The Arabidopsis Book/Am Soc Plant Biol 9:e0153
Kim YY, Choi H, Segami S, Cho HT, Martinoia E, Maeshima M, Lee Y (2009) AtHMA1 contributes to the detoxification of excess Zn(II) in Arabidopsis. Plant J 58:737–753. https://doi.org/10.1111/j.1365-313X.2009.03818.x
Klaumann S, Nickolaus SD, Fürst SH, Starck S, Schneider S, Ekkehard Neuhaus H, Trentmann O (2011) The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana. New Phytol 192:393–404
Kobayashi Y et al (2008) Amino acid polymorphisms in strictly conserved domains of a P-type ATPase HMA5 are involved in the mechanism of copper tolerance variation in Arabidopsis. Plant Physiol 148:969–980. https://doi.org/10.1104/pp.108.119933
Kolaj-Robin O, Russell D, Hayes KA, Pembroke JT, Soulimane T (2015) Cation diffusion facilitator family: structure and function. FEBS Lett 589:1283–1295
Kumar PN, Dushenkov V, Motto H, Raskin I (1995) Phytoextraction: the use of plants to remove heavy metals from soils. Environ Sci Technol 29:1232–1238
Kumar V, Mahajan M, Yadav SK (2012) Toxic metals accumulation, tolerance and homeostasis in brassicaoilseed species: overview of physiological, biochemical and molecular mechanisms. In: The plant family Brassicaceae. Springer, Dordrecht, pp 171–211
Kumlehn J, Pietralla J, Hensel G, Pacher M, Puchta H (2018) The CRISPR/Cas revolution continues: from efficient gene editing for crop breeding to plant synthetic biology. J Integr Plant Biol 60:1127–1153. https://doi.org/10.1111/jipb.12734
Küpper H, Zhao FJ, McGrath SP (1999) Cellular compartmentation of zinc in leaves of the hyperaccumulator Thlaspi caerulescens. Plant Physiol 119:305–312
Küpper H, Lombi E, Zhao F-J, McGrath SP (2000) Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri. Planta 212:75–84
Lane TS et al (2016) Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology. BMC Biotechnol 16:47
Lanquar V, Ramos MS, Lelièvre F, Barbier-Brygoo H, Krieger-Liszkay A, Krämer U, Thomine S (2010) Export of vacuolar manganese by AtNRAMP3 and AtNRAMP4 is required for optimal photosynthesis and growth under manganese deficiency. Plant Physiol 152:1986–1999
Lasat MM (2000) The use of plants for the removal of toxic metals from contaminated soils. US Environmental Protection Agency, Washington, DC
Lasat MM (2002) Phytoextraction of toxic metals: a review of biological mechanisms. J Environ Qual 31:109–120
Lefèvre F, Baijot A, Boutry M (2015) Plant ABC transporters: time for biochemistry? Biochem Soc Trans 43:931–936
Lekeux G et al (2018) di-Cysteine motifs in the C-terminus of plant HMA4 proteins confer nanomolar affinity for zinc and are essential for HMA4 function in vivo. J Exp Bot 69:5547–5560
Li H et al (2013) Simultaneous saccharification and fermentation of broken rice: an enzymatic extrusion liquefaction pretreatment for Chinese rice wine production. Bioprocess Biosyst Eng 36:1141–1148
Li D et al (2015) Genome-wide analysis and heavy metal-induced expression profiling of the HMA gene family in Populus trichocarpa. Front Plant Sci 6:1149
Lin YF et al (2009) Arabidopsis IRT3 is a zinc-regulated and plasma membrane localized zinc/iron transporter. New Phytol:182, 392–404
Lin Y-F, Severing EI, te Lintel HB, Schijlen E, Aarts MG (2014) A comprehensive set of transcript sequences of the heavy metal hyperaccumulator Noccaea caerulescens. Front Plant Sci 5:261
Lin Y, Gritsenko D, Feng S, Teh YC, Lu X, Xu J (2016) Detection of heavy metal by paper-based microfluidics. Biosens Bioelectron 83:256–266
Liu W et al (2015) Transcriptome-wide analysis of chromium-stress responsive microRNAs to explore miRNA-mediated regulatory networks in radish (Raphanus sativus L.). Sci Rep 5:14024. https://doi.org/10.1038/srep14024
Liu H, Zhao H, Wu L, Liu A, Zhao FJ, Xu W (2017) Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol 215:687–698. https://doi.org/10.1111/nph.14622
Lone MI, He Z, Stoffella PJ, Yang X (2008) Phytoremediation of heavy metal polluted soils and water: progresses and perspectives. J Zhejiang Univ 9:210–220
Lysak MA, Koch MA, Pecinka A, Schubert I (2005) Chromosome triplication found across the tribe Brassicaceae. Genome Res 15:516–525
Ma L, Komar K, Tu C, Zhang W, Cai Y, Kennelley E (2001) Addendum: a fern that hyperaccumulates arsenic. Nature 411:438
Marschner H (2011) Marschner’s mineral nutrition of higher plants. Academic, Boston
Martens SN, Boyd RS (1994) The ecological significance of nickel hyperaccumulation: a plant chemical defense. Oecologia 98:379–384
Mäser P et al (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126:1646–1667
McGrath SP, Zhao F-J (2003) Phytoextraction of metals and metalloids from contaminated soils. Curr Opin Biotechnol 14:277–282
McGrath S, Zhao J, Lombi E (2002) Phytoremediation of metals, metalloids, and radionuclides. Adv Agron 75:1–56
Memon AR (2016) Metal hyperaccumulators: mechanisms of hyperaccumulation and metal tolerance. In: Phytoremediation. Springer, Berlin, pp 239–268
Memon AR, Schröder P (2009) Implications of metal accumulation mechanisms to phytoremediation. J Environ Sci Pollut Res 16:162–175
Memon AR, Yatazawa M (1982) Chemical nature of manganese in the leaves of manganese accumulator plants. J Soil Sci Plant Nutr 28:401–412
Memon AR, Yatazawa M (1984) Nature of manganese complexes in manganese accumulator plant-Acanthopanax sciadophylloides. J Plant Nutr 7:961–974
Memon AR, Zahirovic E (2014) Genomics and transcriptomics analysis of Cu accumulator plant Brassica nigra L. J Appl Biol Sci 8:1–8
Memon AR, Itô S, Yatazawa M (1979) Absorption and accumulation of iron, manganese and copper in plants in the temperate forest of Central Japan J. Soil Sci Plant Nutr 25:611–620
Memon AR, Aktoprakligil D, Özdemir A, Verti A (2001) Heavy metal accumulation and detoxification mechanisms in plants. Turk J Bot 25:111–121
Memon AR, Yildizhan Y, Kaplan E (2008) Metal accumulation in crops–human health issues. In: Trace elements as contaminants nutrients—consequences in ecosystems human health. Wiley, Hoboken, pp 81–98
Meng JG, Zhang XD, Tan SK, Zhao KX, Yang ZM (2017) Genome-wide identification of Cd-responsive NRAMP transporter genes and analyzing expression of NRAMP 1 mediated by miR167 in Brassica napus. Biometals 30:917–931. https://doi.org/10.1007/s10534-017-0057-3
Merlot S, de la Torre VSG, Hanikenne M (2018) Physiology and molecular biology of trace element hyperaccumulation. In: Agromining: farming for metals. Springer, Cham, pp 93–116
Milner MJ, Seamon J, Craft E, Kochian LV (2013) Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J Exp Bot 64:369–381
Morel M, Crouzet J, Gravot A, Auroy P, Leonhardt N, Vavasseur A, Richaud P (2009) AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol 149:894–904. https://doi.org/10.1104/pp.108.130294
Moreno I et al (2008) AtHMA1 is a thapsigargin-sensitive Ca2+/heavy metal pump. J Biol Chem 283:9633–9641
Nagaharu U (1935) Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn J Bot 7:389–452
Nevo Y, Nelson N (2006) The NRAMP family of metal-ion transporters. BBA – Mol Cell Res 1763:609–620
Noret N, Meerts P, Vanhaelen M, Dos Santos A, Escarré J (2007) Do metal-rich plants deter herbivores? A field test of the defence hypothesis. Oecologia 152:92–100
Oda K, Otani M, Uraguchi S, Akihiro T, Fujiwara T (2011) Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast. Biosci Biotechnol Biochem 75:1211–1213
Østerberg JT, Palmgren M (2018) Heavy metal pumps in plants: structure, function and origin. In: Advances in botanical research. Elsevier, pp 57–89
Ovečka M, Takáč T (2014) Managing heavy metal toxicity stress in plants: biological and biotechnological tools. Biotechnol Adv 32:73–86
Ozturk M, Memon AR, Gucel S, Sakcali MS (2012) Brassicas in Turkey and their potential role for degraded habitats’ remediation. In: The plant family Brassicaceae. Springer, Dordrecht, pp 265–287
Padmavathiamma PK, Li L (2007) Phytoremediation technology: hyper-accumulation metals in plants. Water Air Soil Pollut 184:105–126
Pang K, Li Y, Liu M, Meng Z, Yu Y (2013) Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L). Gene 526:411–428. https://doi.org/10.1016/j.gene.2013.05.051
Park J et al (2012) The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J 69:278–288. https://doi.org/10.1111/j.1365-313X.2011.04789.x
Park JY, Moon BY, Park JW, Thornton JA, Park YH, Seo KS (2017) Genetic engineering of a temperate phage-based delivery system for CRISPR/Cas9 antimicrobials against Staphylococcus aureus. Sci Rep 7:44929
Paunov M, Koleva L, Vassilev A, Vangronsveld J, Goltsev V (2018) Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in Durum Wheat. Int J Mol Sci 19:787
Pedas P, Ytting CK, Fuglsang AT, Jahn TP, Schjoerring JK, Husted S (2008) Manganese efficiency in barley: identification and characterization of the metal ion transporter HvIRT1. Plant Physiology 148:455–466
Peñarrubia L, Andrés-Colás N, Moreno J, Puig S (2010) Regulation of copper transport in Arabidopsis thaliana: a biochemical oscillator? J Biol Inorg Chem 15:29
Pence NS et al (2000) The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc Nat Acad Sci 97:4956–4960
Perea-García A, Garcia-Molina A, Andrés-Colás N, Vera-Sirera F, Pérez-Amador MA, Puig S, Peñarrubia L (2013) Arabidopsis copper transport protein COPT2 participates in the cross talk between iron deficiency responses and low-phosphate signaling. Plant Physiol 162:180–194
Peuke AD, Rennenberg H (2005) Phytoremediation: molecular biology, requirements for application, environmental protection, public attention and feasibility. EMBO Rep 6:497–501
Pollard AJ, Reeves RD, Baker AJ (2014) Facultative hyperaccumulation of heavy metals and metalloids. Plant Sci 217:8–17
Qin L et al (2017) Genome-wide identification and expression analysis of NRAMP family genes in soybean (Glycine Max L). Front Plant Sci 8:1436. https://doi.org/10.3389/fpls.2017.01436
Rascio N, Navari-Izzo F (2011) Heavy metal hyperaccumulating plants: how and why do they do it? And what makes them so interesting? Plant Sci 180:169–181
Reeves RD, Baker AJM (2000) Metal accumulating plants. In: Phytoremediation of Toxic Metals: Using Plants to Clean up the Environment (Raskin I, Finsley BD eds), Wiley, New York, 193–229.
Reeves RD, Schwartz C, Morel JL, Edmondson J (2001) Distribution and metal-accumulating behavior of Thlaspi caerulescens and associated metallophytes in France. Int J Phytoremediation 3:145–172
Reeves R, Baker A, Jaffré T, Erskine P, Echevarria G, van der Ent A (2017) A global database for hyperaccumulator plants of metal and metalloid trace elements. New Phytol:14907
Reeves RD, Baker AJ, Jaffré T, Erskine PD, Echevarria G, van der Ent A (2018) A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol 218:407–411
Rensing C, Ghosh M, Rosen BP (1999) Families of soft-metal-ion-transporting ATPases. J Bacteriol 181:5891–5897
Ricachenevsky FK, Sperotto RA (2016) Into the wild: Oryza species as sources for enhanced nutrient accumulation and metal tolerance in rice. Front Plant Sci 7:974
Ricachenevsky FK, Menguer PK, Sperotto RA, Williams LE, Fett JP (2013) Roles of plant metal tolerance proteins (MTP) in metal storage and potential use in biofortification strategies. Front Plant Sci 4:144
Rogers EE, Eide DJ, Guerinot ML (2000) Altered selectivity in an Arabidopsis metal transporter. Proc Nat Acad Sci 97:12356–12360
Rosenzweig AC, Argüello JM (2012) Toward a molecular understanding of metal transport by P1B-Type ATPases. In: Current topics in membranes, vol 69. Elsevier, pp 113–136
Salt DE, Blaylock M, Kumar NP, Dushenkov V, Ensley BD, Chet I, Raskin I (1995) Phytoremediation: a novel strategy for the removal of toxic metals from the environment using plants. Bio/Technol 13:468
Sancenón V, Puig S, Mira H, Thiele DJ, Peñarrubia L (2003) Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol Biol 51:577–587
Sancenón V, Puig S, Mateu-Andrés I, Dorcey E, Thiele DJ, Peñarrubia L (2004) The Arabidopsis copper transporter COPT1 functions in root elongation and pollen development. J Biol Chem 279:15348–15355
Sarma RK, Gowtham I, Bharadwaj R, Hema J, Sathishkumar R (2018) Recent advances in metal induced stress tolerance in plants: possibilities and challenges. In: Plants under metal and metalloid stress. Springer, Singapore, pp 1–28
Sasaki A, Yamaji N, Ma JF (2014) Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J Exp Bot 65:6013–6021. https://doi.org/10.1093/jxb/eru340
Scheben A, Wolter F, Batley J, Puchta H, Edwards D (2017) Towards CRISPR/Cas crops–bringing together genomics and genome editing. New Phytol 216:682–698
Schindele P, Wolter F, Puchta H (2018) Transforming plant biology and breeding with CRISPR/Cas9, Cas12 and Cas13. FEBS Lett 592:1954–1967. https://doi.org/10.1002/1873-3468.13073
Seigneurin-Berny D et al (2006) HMA1, a new Cu-ATPase of the chloroplast envelope, is essential for growth under adverse light conditions. J Biol Chem 281:2882–2892
Shikanai T, Müller-Moulé P, Munekage Y, Niyogi KK, Pilon M (2003) PAA1, a P-type ATPase of Arabidopsis, functions in copper transport in chloroplasts. Plant Cell 15:1333–1346
Stein RJ et al (2017) Relationships between soil and leaf mineral composition are element-specific, environment-dependent and geographically structured in the emerging model Arabidopsis halleri. New Phytol 213:1274–1286
Stephan UW, Schmidke I, Stephan VW, Scholz G (1996) The nicotianamine molecule is made-to-measure for complexation of metal micronutrients in plants. Biometals 9:84–90
Stephens BW, Cook DR, Grusak MA (2011) Characterization of zinc transport by divalent metal transporters of the ZIP family from the model legume Medicago truncatula. BioMetals 24:51–58
Szczygłowska M, Bodnar M, Namieśnik J, Konieczka P (2014) The use of vegetables in the biomonitoring of cadmium and lead pollution in the environment. Crit Rev Anal Chem 44:2–15
Tang X et al (2017) A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nature Plants 3:17018
Tangahu BV, Abdullah S, Rozaimah S, Basri H, Idris M, Anuar N, Mukhlisin M (2011) A review on heavy metals (As, Pb, and Hg) uptake by plants through phytoremediation. Int J Chem Eng 2011:1–31
Tiong J, McDonald G, Genc Y, Shirley N, Langridge P, Huang CY (2015) Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytologist 207:1097–1109
Vamerali T, Bandiera M, Mosca G (2010) Field crops for phytoremediation of metal-contaminated land. A review. Environ Chem Lett 8:1–17
van der Ent A, Echevarria G, Baker AJ, Morel JL (2018) Preface. In: Agromining: farming for metals extracting unconventional resources using plants. Springer, Cham, pp V–VI
van der Zaal BJ, Neuteboom LW, Pinas JE, Chardonnens AN, Schat H, Verkleij JA, Hooykaas PJ (1999) Overexpression of a novel Arabidopsis gene related to putative zinc-transporter genes from animals can lead to enhanced zinc resistance and accumulation. Plant Physiol 119:1047–1056
Vara Prasad MN, de Oliveira Freitas HM (2003) Metal hyperaccumulation in plants: biodiversity prospecting for phytoremediation technology. Electron J Biotechnol 6:285–321
Verbruggen N, Hermans C, Schat H (2009) Molecular mechanisms of metal hyperaccumulation in plants. New Phytol 181:759–776
Verbruggen N, Hanikenne M, Clemens S (2013) A more complete picture of metal hyperaccumulation through next-generation sequencing technologies. Front Plant Sci 4:388
Verrier PJ et al (2008) Plant ABC proteins–a unified nomenclature and updated inventory. Trends Plant Sci 13:151–159
von Wirén N et al (1999) Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants. Plant Physiol 119:1107–1114
Wang Z et al (2016) MoTe 2: a type-II Weyl topological metal. Physical Rev Lett 117:056805
Wang J, Liang S, Xiang W, Dai H, Duan Y, Kang F, Chai T (2019) A repeat region from the Brassica juncea HMA4 gene BjHMA4R is specifically involved in Cd2+ binding in the cytosol under low heavy metal concentrations. BMC Plant Biol 19:89. https://doi.org/10.1186/s12870-019-1674-5
Warwick SI, Black LD (1991) Molecular systematics of Brassica and allied genera (subtribe Brassicinae, Brassiceae)—chloroplast genome and cytodeme congruence. Theor Appl Genet 82:81–92
Weast RC, Astle MJ, Beyer WH (1988) CRC handbook of chemistry and physics vol 69. CRC Press, Boca Raton
Williams LE, Mills RF (2005) P1B-ATPases–an ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci 10:491–502
Wintz H et al (2003) Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J Biol Chem 278:47644–47653
Wolter F, Puchta H (2018) The CRISPR/Cas revolution reaches the RNA world: Cas13, a new Swiss Army knife for plant biologists. Plant J 94:767–775. https://doi.org/10.1111/tpj.13899
Wong CKE, Cobbett CS (2009) HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytologist 181:71–78. https://doi.org/10.1111/j.1469-8137.2008.02638.x
Xu X, Shi J, Chen X, Chen Y, Hu T (2009) Chemical forms of manganese in the leaves of manganese hyperaccumulator Phytolacca acinosa Roxb. (Phytolaccaceae). Plant Soil 318:197
Xu Y et al (2014) Os ABCB 14 functions in auxin transport and iron homeostasis in rice (Oryza sativa L.). Plant J 79:106–117
Yamasaki H, Hayashi M, Fukazawa M, Kobayashi Y, Shikanai T (2009) SQUAMOSA promoter binding protein–like7 is a central regulator for copper homeostasis in Arabidopsis. Plant Cell 21:347–361
Yan C, Duan W, Lyu S, Li Y, Hou X (2017) Genome-wide identification, evolution, and expression analysis of the ATP-binding cassette transporter gene family in Brassica rapa. Front Plant Sci 8:349
Yang J et al (2016) The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat Genet 48:1225
Yuan M, Li X, Xiao J, Wang S (2011) Molecular and functional analyses of COPT/Ctr-type copper transporter-like gene family in rice. BMC Plant Biol 11:69
Zhang W, Cai Y, Tu C, Ma LQ (2002) Arsenic speciation and distribution in an arsenic hyperaccumulating plant. Sci Total Environ 300:167–177
Zhao H, Wang L, Zhao FJ, Wu L, Liu A, Xu W (2018) SpHMA1 is a chloroplast cadmium exporter protecting photochemical reactions in the Cd hyperaccumulator Sedum plumbizincicola. Plant Cell Environ 42:1112–1124
Zheng X, Chen L, Li X (2018) Arabidopsis and rice showed a distinct pattern in ZIPs genes expression profile in response to Cd stress. Bot Stud 59:22
Zhou ZS, Song JB, Yang ZM (2012) Genome-wide identification of Brassica napus microRNAs and their targets in response to cadmium. J Exp Bot 63:4597–4613. https://doi.org/10.1093/jxb/ers136
Zhu C et al (2017) Characteristics of genome editing mutations in cereal crops. Trends Plant Sci 22:38–52. https://doi.org/10.1016/j.tplants.2016.08.009
Acknowledgments
The author thanks Ms. Fatma Kusur, Department of Molecular Biology and Genetics, Usak University, and Mr. Muhammet Memon, International Biomedical and Genomic Research Center, Dokuz Eylul University, for their help in reference arrangement and their valuable comments on some part of this review. This work is supported by BAP project F010.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Memon, A.R. (2020). Heavy Metal–Induced Gene Expression in Plants. In: Naeem, M., Ansari, A., Gill, S. (eds) Contaminants in Agriculture. Springer, Cham. https://doi.org/10.1007/978-3-030-41552-5_7
Download citation
DOI: https://doi.org/10.1007/978-3-030-41552-5_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-41551-8
Online ISBN: 978-3-030-41552-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)