
Abstract
Many birds and mammals give alarm calls when they detect predators or other threats, and these calls have been used as classic models for understanding signal design. Here we consider signal design and usage, and how individuals acquire and use information from the alarm calls of other species. Alarm calls often encode detailed information on danger, such as the type of predator, its current behavior, size, or proximity. Alarm calls are sometimes very similar among species or can share generic acoustic features, and both help to explain recognition of heterospecific alarms. However, alarm calls can vary greatly among species, and taxonomically widespread eavesdropping also requires learning the association between calls and danger. Once heterospecifics eavesdrop on alarm calls, there is potentially selection on callers to modify their alarm calls or usage. If callers benefit from eavesdroppers’ responses to their alarm calls, they may be selected to enhance signal efficacy, leading to interspecific communication and mutual benefit. Alternatively, callers can be selected to manipulate eavesdroppers, using deceptive signaling, including mimicry, causing the eavesdropper to suffer a cost. If callers suffer a cost from eavesdroppers’ responses, their signaling can be modified to make eavesdropping harder, leading to cue denial. Overall, alarm signals provide an insight into the evolution of signal design, and the complex flow of information within and among species in natural communities.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
12.1 Alarm Calls, Heterospecific Eavesdropping, and Communication
12.1.1 Alarm Calls and Eavesdropping
Many birds and mammals give alarm calls when they detect predators or other threats. These calls have evolved to warn conspecifics of danger, or to deter predators by harassing them or informing them that they have lost the element of surprise (Klump and Shalter 1984; Caro 2005; Zuberbühler 2009). Alarm calls are of widespread ecological importance because most species are vulnerable to predators, and these calls have been used as classic models for understanding signal design and the evolution of communication (Marler 1955, 1957; Maynard Smith 1965; Sherman 1977). Furthermore, given the broad relevance of information on danger, it is not surprising that many individuals respond to the alarm calls of other species that face similar threats (Seppänen et al. 2007; Goodale et al. 2010; Schmidt et al. 2010; Magrath et al. 2015a). Here we consider signal design and usage, and how individuals acquire and use information from the alarm calls of heterospecifics.
12.1.2 Definitions and Scope of Chapter
We consider responses to heterospecific alarm calls regardless of whether they involve communication, eavesdropping, or deception. Communication entails the use of signals that have evolved because the sender benefits from the receiver’s response, so that signals have an “intended” audience (Bradbury and Vehrencamp 2011). Furthermore, receivers of signals are selected to respond only if they also benefit, so that communication requires evolution of both signalers and receivers (Maynard Smith and Harper 2003). Unlike communication, eavesdropping entails reception of signals intended for others. Here we use eavesdropping to mean “interceptive eavesdropping” (Peake 2005), in which individuals respond to signals themselves, and not just communication interactions among others (“social eavesdropping”; McGregor and Dabelsteen 1996; Peake 2005; Searcy and Nowicki 2005). Eavesdroppers, like receivers in communication, will be selected to respond if they gain relevant information. In contrast to both communication and eavesdropping, senders may use signals to deceive an intended audience. Deception differs from communication because, despite the receiver being the intended recipient, it does not benefit from its response to the signal, so that there is a clear conflict of interest between sender and receiver (Searcy and Nowicki 2005). We include communication, eavesdropping, and deception because it can be difficult to know which is in play during receiver responses to alarm calls, and because there is likely to be an evolutionary continuum from eavesdropping to communication and deception (Kostan 2002; Radford et al. 2011).
In this chapter, we first consider what information is conveyed during alarm communication within species, and how it is encoded. This provides the backdrop for understanding what information could be acquired by heterospecifics. We then consider the benefits of eavesdropping on other species’ calls, and how individuals are able to recognize the alarm calls of other species. We follow by considering how eavesdropping can lead to communication, deception, or suppression of information. We finish by focusing on vocal mimicry, which unites the themes of call structure, eavesdropping, communication, and deception.
12.2 Alarm Communication Within Species
12.2.1 Function of Alarm Calls
We define alarm calls broadly to include any calls given specifically when a predator is nearby, and which prompt antipredator behavior in conspecifics (Klump and Shalter 1984; Zuberbühler 2009; Magrath et al. 2015a). Calls intended to communicate with predators are sometimes considered signals of unprofitability or deterrence, rather than alarm calls (Caro 2005; Bradbury and Vehrencamp 2011), but conspecifics are also likely to respond to these calls. Furthermore, signals could evolve because of their effects on both predators and conspecifics, so that a broad definition is important for understanding alarm call evolution.
Alarm calls can be named and classified in many ways, such as according to the context of production or their acoustic structure, but many species have three main types, which are associated with different antipredator responses (Klump and Shalter 1984). (1) Flee alarm calls are given in response to immediate danger, and prompt listeners to flee or become cryptic. When given to flying predators, such as hawks, these are often called “aerial” or “hawk” alarm calls. (2) Mobbing alarm calls are given to predators not posing immediate danger, and prompt others to approach, monitor, and sometimes harass the predator. (3) Distress alarm calls are given when individuals are attacked, which might startle the predator, recruit others to help the caller, or attract secondary predators.
Alarm calls often communicate about the type of threat, the degree of danger posed, or both, with varying degrees of specificity. “Functionally referential” alarm calls communicate about the type of threat (Gill and Bierema 2013; Townsend and Manser 2013; Suzuki 2016). Vervet monkeys, Chlorocebus pygerythrus, for a classic example, produce acoustically distinct alarm calls to leopards, eagles, and snakes, and respond appropriately to playback of those calls, such as by looking down to “snake” alarms and up to “eagle” alarms (Seyfarth et al. 1980a, b). Similarly, Japanese great tits, Parus minor, have different alarm calls for different predators, and nestlings respond by fleeing the nest when warned of snakes but crouching down when warned of crows (Suzuki 2011). These responses are appropriate given that snakes can enter nests but crows cannot. Some birds even have alarm calls given specifically to brood parasites (Gill and Sealy 1996; Feeney and Langmore 2013; Wheatcroft and Price 2015). Other species have alarm calls that communicate about the predator’s current behavior. For example, Siberian jays, Perisoreus infaustus, and noisy miners, Manorina melanocephala, both have acoustically different calls to perched and airborne raptors (Griesser 2008; Cunningham and Magrath 2017; Farrow et al. 2017). Alarm calls can also communicate about the degree of danger, related to predator proximity or type, such as in white-browed scrubwrens, Sericornis frontalis (Leavesley and Magrath 2005), black-capped chickadees, Poecile atricapillus (Templeton et al. 2005) and New Holland honeyeaters, Phylidonyris novaehollandiae (McLachlan and Magrath 2020). Alarm calls may also communicate simultaneously about both the type of danger and degree of threat, as shown for meerkats, Suricata suricatta (Manser 2001; Manser et al. 2001, 2002). Not all alarm calls convey specific information, as some are given in a broad range of contexts, such as “general” or “variable-use” calls given to a variety of threats and in social interactions (Marler 1957; Fichtel and Kappeler 2002). In these cases, context and call variability can nonetheless guide listeners’ responses (Wheatcroft 2015). Overall, different alarm calls allow listeners to take appropriate action according to the type or urgency of the threat.
12.2.2 Coding: Marler’s Insight on Structure and Function
In a classic paper on signal design, Peter Marler proposed that the acoustic structure of alarm calls should reflect their function (Marler 1955). Passerine alarm calls given to flying hawks should be difficult to locate, to reduce the risk of attack, so Marler predicted that they should be high-pitched, pure tones, with a gradual onset and ending. By contrast, mobbing calls given to perched raptors should be easily locatable so that others can join in mobbing. They should therefore be abrupt, repetitive, and more broadband than calls given to flying hawks. Consistent with these predictions, many European passerines have “seeet” aerial alarm calls (Fig. 12.1a) and mobbing calls with exactly these acoustic features (Marler 1955, 1957).

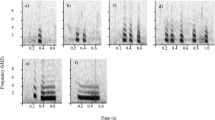
Similarities and differences among alarm calls given by passerines to flying hawks. (a) Seeet alarm calls given by some European passerine species, redrawn from Marler (1957): (a) reed bunting, Emberiza schoeniclus; (b) blackbird, Turdus merula; (c) great tit, Parus major; (d) blue tit, Cyanistes caeruleus; and (e) chaffinch, Fringilla coelebs. (b) Aerial alarm calls given by Australian members of the superfamily Meliphagoidea. Only single elements are shown, but calls often have multiple similar elements. (a) Western thornbill, Acanthiza inornata; (b) white-winged fairy-wren, Malurus leucopterus; (c) superb fairy-wren, M. cyaneus; (d) splendid fairy-wren, M. splendens; (e) variegated fairy-wren, M. lamberti; (f) southern whiteface, Aphelocephala leucopsis; (g) chestnut-rumped thornbill, Acanthiza uropygialis; (h) white-browed scrubwren, Sericornis frontalis; (i) brown gerygone, Gerygone mouki; (j) New Holland honeyeater, Phylidonyris novaehollandiae; (k) white-plumed honeyeater, Ptilotula penicillata; (l) noisy miner, Manorina melanocephala; recordings by Robert Magrath and Janet Gardner. Note the different time and frequency scales in (a) and (b)
Subsequent research supports Marler’s main conclusions about seeet aerial alarm calls versus mobbing calls. Playback experiments show that seeet calls are harder for predators to locate than mobbing calls (Klump and Shalter 1984; Jones and Hill 2001). Seeet calls are also likely to give minimal distance cues. The long duration and gradual onset and termination will reduce the ability to detect reverberations, while the narrow frequency range stops assessment of the relative attenuation of different frequencies; both these features can be used to judge distance (Naguib and Wiley 2001). In addition to being difficult to locate, seeet calls are difficult to hear, especially for hawks. The high frequency of these calls (7–8 kHz) means they are unlikely to be heard at a great distance because of relatively rapid attenuation (Wiley and Richard 1982) and because avian hearing often becomes less sensitive above 4–5 kHz (Dooling 2004). More subtly, differences in hearing sensitivities are exploited by great tits, whose seeet call is at a frequency to which it is much more sensitive than the sparrowhawk, Accipiter nisus, its major predator (Klump et al. 1986). This call frequency therefore minimizes the risk of eavesdropping by hawks.
12.2.3 Generic Acoustic Features
Alarm call structure reflects function more broadly, and potentially for additional reasons than those suggested by Marler (1955). In a diversity of birds and mammals, flee alarm calls are often more narrow-band than mobbing or distress alarms given by the same species, which is likely to affect audibility or localizability (Marler 1957; Aubin 1989; Jurisevic and Sanderson 1994a; Bradbury and Vehrencamp 2011). These broad differences might also reflect constraints of motivational-structural rules, because aggressive calls are often of greater bandwidth, less tonal, and of lower frequency than fearful calls (Morton 1977, 2017). Related to this idea is that some alarm calls may be selected to capture the attention of listeners, by taking advantage of acoustic features that are difficult to ignore or easy to hear over a long distance (Aubin 1989; Jurisevic and Sanderson 1998; Fitch et al. 2002; Neudorf and Sealy 2002; Blumstein and Récapet 2009; Rendall et al. 2009; Branch and Freeberg 2012). In addition, the contrasting function of flee and mobbing calls may select for contrasting call structures, following the principle of antithesis (Darwin 1872; Hurd et al. 1995). In this case call structure could be arbitrary but selected to contrast with calls of different meaning. In a recent examination of this idea, however, superb fairy-wrens were able to discriminate between their flee and mobbing alarm calls despite their surprisingly subtle acoustic differences (Tegtman and Magrath in press). Overall, alarm call structure is not completely arbitrary, and it is likely to enhance or diminish audibility or locatability depending on the call type. In addition, alarm calls can be a reflection of the caller’s internal state or the listener’s perception, or enhance the ability to discriminate between calls of contrasting meaning.
12.2.4 Interspecific Alarm Call Diversity
Despite the broad association of call structure and meaning, there is huge diversity among species’ alarm calls. While c. 7 kHz seeet flee alarm calls are given to flying hawks by many species in Europe and North America, comparable aerial alarm calls given by other species can vary dramatically in frequency and structure. For example, Australian honeyeater (Family Meliphagidae) aerial alarm calls are typically composed of short, rapidly repeated elements, often with frequency sweeps, and with mean frequencies of 2–4 kHz (Rooke and Knight 1977; Jurisevic and Sanderson 1994b). Furthermore, in the superfamily Meliphagoidea (including Meliphagidae, Maluridae, and Acanthizidae), aerial alarm calls vary in frequency, duration, bandwidth, and possession of rapid frequency modulation, and very few have a seeet-like structure (Jurisevic and Sanderson 1998; Fallow et al. 2011, 2013; Cunningham and Magrath 2017; Fig. 12.1b). Similarly, birds in Sri Lankan mixed flocks have a diversity of alarm calls given to raptors, many of which are of relatively low frequency and high bandwidth (Goodale and Kotagama 2005). In neither case is it clear why there is so much variation. There is also great diversity in mobbing call structure. A study of 52 species of American passerines from 12 families revealed substantial diversity among mobbing calls; only about 50% had mobbing calls with abrupt elements of broad frequency range (Ficken and Popp 1996). Similarly, calls used for mobbing vary “drastically” among 22 species of Himalayan passerines, perhaps in part because they are also used in social contexts within species (Wheatcroft and Price 2013).
12.2.5 Alarm Call Usage
In addition to encoding information within call element structure and frequency, call usage and composition can affect meaning in multiple ways. First, the number of elements included in a call can convey graded information on the degree of danger. For example, superb fairy-wrens, Malurus cyaneus, New Holland honeyeaters and white-browed scrubwrens include more elements in their aerial alarm calls when a flying predator is closer or more dangerous (Leavesley and Magrath 2005; Fallow and Magrath 2010; McLachlan and Magrath 2020), and black-capped chickadees include more D notes in their chick-a-dee calls when mobbing a more dangerous raptor (Templeton et al. 2005). Second, the overall rate of call delivery can change call meaning. For example, a suite of passerines from India, Japan, and Sweden have a higher call repetition rate when calling to predators than during social interactions, and playbacks show that listeners respond with antipredator behavior when call repetition rate is higher (Wheatcroft 2015). In this case, call usage changes the meaning of the same variable-use calls. Third, the relative use of different calls can convey graded and possibly categorical information about danger. For example, Arabian babblers, Turdoides squamiceps, include relatively more high-pitched than low-pitched calls when mobbing a live cat than an owl mount, implying that the combination of calls conveys information about the type or magnitude of danger (Naguib et al. 1999). Similarly, white-handed gibbon, Hylobates lar, songs contain relatively more “hoo” notes in songs prompted by predators than in duet songs with their mates (Clarke et al. 2006). Finally, call syntax can affect meaning. For example, Japanese great tits give D notes alone in the absence of predators but together with other note types when predators are near, and the response to compound calls depends on whether the D notes are before or after the other notes (Suzuki et al. 2016). In this case the order of notes within a call produces a compound meaning, rather than merely the relative abundance of different notes.
12.3 Information from Heterospecific Alarms
12.3.1 Diversity of Eavesdropping
Heterospecific alarm calls are a potentially rich source of information about danger for species facing similar threats, and over 70 species have been shown through experimental playback to respond to other species’ alarm calls (Magrath et al. 2015a). Individuals usually respond to playback of heterospecific alarm calls with typical antipredator behavior, such as by becoming vigilant, fleeing to cover or freezing after flee alarm calls, or approaching and calling in response to mobbing or distress calls. Examples span a diversity of taxa, with both mammals and birds responding to alarm calls of other species of mammals and birds. There are even a few examples of lizards responding to bird alarm calls (Vitousek et al. 2007; Ito and Mori 2010; Ito et al. 2013), showing that individuals do not need to have their own alarm calls, or even to produce any vocalizations, to be responsive to the calls of other species.
12.3.2 Information Gained
Do individuals gain similarly detailed information on danger from heterospecfic and conspecific alarm calls? There has been no broad study, but in some cases they do, whereas in other cases they do not. Individuals can gain information on predator type from other species’ referential alarm calls. For example, ring-tailed lemurs, Lemur catta, respond to Verreaux’ sifaka’s, Propithecus verreauxi, aerial alarm calls by looking up and to their terrestrial alarm calls by running to trees (Oda and Masataka 1996). Comparable patterns are found in other primates (Zuberbühler 2000b; Kirchhof and Hammerschmidt 2006). Similarly, Carolina chickadees, Poecile carolinensis, respond appropriately to the aerial and mobbing calls of tufted titmice, Baeolophus bicolor (Hetrick and Sieving 2012), Australian magpies, Cracticus tibicen, orient appropriately to aerial and mobbing alarm calls of noisy miners (Dawson Pell et al. 2018), and yellow-casqued hornbills, Ceratogymna elata, respond to the relevant “eagle” but not irrelevant “leopard” alarm calls given by Diana monkeys, Cercopithecus diana (Rainey et al. 2004). Individuals can also respond to graded heterospecific alarm calls. Superb fairy-wrens, white-browed scrubwrens, and red-breasted nuthatches, Sitta canadensis, all respond to graded heterospecific alarm calls indicating the degree of danger (Templeton and Greene 2007; Fallow and Magrath 2010). In other cases, eavesdroppers gain only partial information. For example, although banded mongooses, Mungos mungo, increased vigilance after playback of lapwing, Vanellus spp., alarm calls, they did not respond to urgency information in the alarm calls of the only species tested, the crowned plover, V. coronatus (Müller and Manser 2008). Overall, individuals can use at least some of the detailed information encoded in other species’ alarms, but we know little about potential constraints on decoding heterospecific alarms (Murray and Magrath 2015).
12.3.3 Benefits
The ability to eavesdrop on other species’ alarm calls is likely to bring many benefits, ultimately increasing survival and reproductive success. Being able to eavesdrop on heterospecifics means that there are more eyes looking for danger, but there could also be specific benefits gained from heterospecific alarm calls over conspecifics ones. For example, species on the ground or with a foraging niche that restricts vigilance may rely on alarm calls of other species that are highly vigilant, such a flycatching species that rely on vigilance to detect prey (Goodale and Kotagama 2008; Martínez and Zenil 2012). In a within-species test of this idea, New Holland honeyeaters were slower to spot gliding predator models, and more likely to flee to alarm calls, when feeding on nectar in flowers than when hawking for insects from exposed perches (McLachlan et al. 2019). Similarly, some species lack alarm calls or rarely give them, and yet respond to those of other species (Sullivan 1985; Vitousek et al. 2007; Sridhar et al. 2009; Srinivasan et al. 2010). Immediate responses to alarm calls presumably reduce the probability of detection or attack by predators, but there will also be indirect benefits. First, being attentive to heterospecific alarms can increase foraging success. Scimitarbills, Rhinopomastus cyanomelas, eavesdrop on alarm calls of pied babblers, Turdoides bicolor, and when in mixed groups they use babblers for information on danger and so reduce personal vigilance and increase foraging success (Ridley et al. 2014). Second, eavesdropping on alarm calls can also affect the choice of feeding location or habitat (Parejo et al. 2012). Third, heterospecific alarms potentially provide information on danger without resource competition, which could diminish the net benefit of comparable information from conspecifics (Seppänen and Forsman 2007). Finally, eavesdropping may facilitate associative learning about previously unrecognized predators (Curio et al. 1978; Vieth et al. 1980) or alarm calls (Potvin et al. 2018).
12.4 Mechanisms of Recognition of Heterospecific Alarm Calls
12.4.1 Learned and Unlearned Recognition
The great diversity of alarm calls within and among species raises the puzzle of how individuals recognize the alarm calls of other species. The answer is complex; animals can recognize some alarm calls on first exposure, whereas other alarm calls require learning, and both mechanisms have potential costs and benefits. Reacting to acoustic features alone allows for response to novel alarm calls on first exposure, but restricts the response to calls with a narrow range of characteristics (Hollén and Radford 2009). By contrast, learning allows individuals to gain information from alarm calls with a broad range of acoustic features, but leaves individuals more vulnerable to predators until they have learned to recognize the calls (Griffin 2004). For most species, recognition of heterospecific alarm calls probably involves both innate responses and learning, which helps explain the widespread occurrence of eavesdropping across taxa (Magrath et al. 2015a, b; Potvin et al. 2018).
12.4.2 Acoustic Similarity to Conspecific Calls
Individuals often respond to novel calls that are acoustically similar to conspecific calls. Recognition of conspecific alarm calls appears to rely on one or a few key acoustic features, with generalization of response to unfamiliar calls which share those features (Wiley and Richard 1982; Ghirlanda and Enquist 2003; ten Cate and Rowe 2007; Fallow et al. 2013; Magrath et al. 2015a; Meise et al. 2018). Indeed, the mutual response by many European songbirds to one another’s high-pitched seeet flee alarms is probably due to acoustic similarity (Marler 1957; Sect. 12.2.2). More recent work has revealed some key acoustic features that are generalized to heterospecific alarms, with secondary features modifying the response (Aubin and Brémond 1989; Aubin 1991; Johnson et al. 2003; Fallow et al. 2011, 2013; Dutour et al. 2017). For example, superb fairy-wrens flee to cover to unfamiliar natural and synthetic aerial alarm calls when their peak frequencies are similar to conspecific calls (Fig. 12.2), and adjust how much time they spend in cover based on similarity to the rate of frequency modulation of conspecific calls (Fallow et al. 2011, 2013). Similarly, several European species of birds respond to synthetic distress calls that share the key property of an increasing frequency sweep, but their response is also likely to be affected by fine-scale acoustic differences (Aubin 1991). Acoustic similarities among species’ alarms need not be exact to prompt response, probably because of natural variability in alarm calls and because responding to alarm calls is critical for survival. Conspecific alarm calls vary among individuals and are affected by environmental degradation during transmission, creating selection for a broad response range that is likely to mean generalization to a greater range of heterospecific alarm calls (Blumstein and Munos 2005; Leavesley and Magrath 2005; Fallow et al. 2011, 2013; Magrath et al. 2015a). In addition, the costs of not responding to an alarm call when there is a predator will select for a low threshold for signal detection (Wiley 2015), and therefore response to a greater range of both conspecific and heterospecific alarm calls.

Fairy-wrens, Malurus cyaneus, respond to unfamiliar aerial alarm calls that are acoustically similar in peak frequency to their own alarms. (a) Response of fairy-wrens (inset) to playback of conspecific and unfamiliar heterospecific aerial alarm calls from members of the Acanthizidae and other Maluridae: (a) western thornbill, Acanthiza inornata; (b) white-winged fairy-wren, Malurus leucopterus; (c) superb fairy-wren, M. cyaneus; (d) splendid fairy-wren, M. splendens; (e) variegated fairy-wren, M. lamberti; (f) southern whiteface, Aphelocephala leucopsis; (g) chestnut-rumped thornbill, Acanthiza uropygialis; (h) brown gerygone, Gerygone mouki. Results from Fallow et al. (2011). (b) Response by fairy-wrens (inset) to playbacks of computer-synthesized calls based on variation in the Maluridae, Acanthizidae and Meliphagidae. The red shading highlights sounds to which fairy-wrens usually fled, with a darker shade representing a higher percentage. The 4 kHz unmodulated call is similar to that of a familiar local species, the New Holland honeyeater, Phylidonyris novaehollandiae. Figure (b) modified from Fallow et al. 2013; photograph in (a) and (b) by Jessica McLachlan
12.4.3 Generic Acoustic Features
Although acoustic similarity to conspecific calls can allow recognition of novel alarm calls, it is not the only potential mechanism of unlearned response. The alarm calls of many species contain acoustic features that are inherently attention-grabbing or frightening, and this may also prompt immediate response to unfamiliar calls (Hirth and McCullough 1977; Morton 1977; Fitch et al. 2002; Rendall et al. 2009). Many mobbing, distress, “panic,” or general alarm calls, for example, contain elements that are harsh, abrupt, broadband, or nonlinear. These acoustic features tend to be honest signals of caller distress, increase listener attention and response, and prevent habituation (Manser 2001; Blumstein and Récapet 2009; Townsend and Manser 2011; Blumstein and Chi 2012; Blesdoe and Blumstein 2014; Karp et al. 2014). For example, great-tailed grackles, Quiscalus mexicanus, increase vigilance after playback of novel synthetic calls that include nonlinear features such as abrupt transitions and noise (Slaughter et al. 2013), and western gray kangaroos, Macropus fuliginosus, do not habituate to novel sounds with nonlinear features (Biedenweg et al. 2011).
12.4.4 Learning
Although some acoustic structures prompt unlearned responses, alarm calls vary greatly among species (Sect. 12.2.4), so that learning is essential to allow taxonomically widespread eavesdropping. In addition to variation in alarm calls within and among species, eavesdroppers face the problem that the species present in a community can vary geographically and temporally, increasing the need for learning (Griffin 2004). Learning allows individuals to tailor their responses to the most relevant alarm calls in the current community (Nocera and Ratcliffe 2010), as well as to ignore non-alarm calls and the alarm calls of species that do not share similar predators (Magrath et al. 2009a). Studies of both geographic and temporal patterns of response to heterospecific alarm calls provide indirect but compelling evidence for learning (Hauser 1988; Ramakrishnan and Coss 2000; Zuberbühler 2000a; Magrath et al. 2009b; Haff and Magrath 2012; Magrath and Bennett 2012; Haff and Magrath 2013; Wheatcroft and Price 2013). For example, bonnet macaques, Macaca radiata, respond to playback of alarm calls of only the locally common of two species of langur at any one site (Ramakrishnan and Coss 2000; Fig. 12.3); Himalayan bird communities respond more strongly to familiar than to unfamiliar heterospecific alarm calls, regardless of acoustic similarity (Wheatcroft and Price 2013); and fledgling white-browed scrubwrens, respond to aerial alarm calls of New Holland honeyeaters at a younger age where their territories overlap (Haff and Magrath 2013).

Response of bonnet macaques, Macaca radiata, to playback of the alarm calls of Nilgiri langurs, Semnopithecus johnii, and Hanuman langurs, S. entellus, in two separate geographic areas. Nilgiri langurs were only common at Mundanthurai, while Hanuman langurs were only common at Mudumalai. Macaques usually only fled to playback of the familiar species’ alarm calls. Redrawn from Ramakrishnan and Coss (2000)
Experimental evidence demonstrates that individuals can learn to respond to unfamiliar heterospecific alarm calls (Shriner 1999; Magrath et al. 2015b; Potvin et al. 2018; Dutour et al. 2019). For example, wild superb fairy-wrens learned over a 2-day period to respond to novel sounds, as if they were alarm calls, when those sounds were repeatedly presented with gliding hawk models (Magrath et al. 2015b; Fig. 12.4). Individual birds were trained to one of two novel sounds, with the other acting as a control, and fled only to the trained sound. This shows that they learned to recognize specific sounds, and were not merely responding because they became scared of playbacks in general. Further work on this system has revealed that fairy-wrens can learn to recognize novel alarm calls through association with a chorus of known alarm calls (Potvin et al. 2018), demonstrating that learning can take place without visual pairing of predators with alarm calls or fearful responses.

Superb fairy-wrens, Malurus cyaneus, were trained to respond to unfamiliar alarm calls. (a) Unfamiliar calls included the aerial alarm call of the allopatric chestnut-rumped thornbill (TB), Acanthiza uropygialis, and a sound synthesized on computer (SYN), and both were different to conspecific calls (FW) and local heterospecific calls. (b) Unfamiliar calls were presented to fairy-wrens along with gliding model predators of pied currawongs (CW), Strepera graculina, and collared sparrowhawks (SH), Accipiter cirrocephalus. Each focal bird was exposed to only one of the two novel calls during training with model predators, whereas the other was unexposed and acted as a control sound. (c) After training, focal fairy-wrens fled only to the exposed sound and not to the unexposed novel sound. Figure reproduced from Magrath et al. (2015b)
Learning and acoustic attributes could simultaneously be important in alarm call recognition if some features of alarm calls facilitate learning. For example, unfamiliar calls that have “attention grabbing” features (Sect. 12.2.3) may be easier to learn about than calls that lack such features (Ghirlanda and Enquist 2003). One possible example concerns greater responses to “variable use” calls that are given at higher rates when predators are near (Sect. 12.2.5). Playback experiments show that both conspecifics and heterospecifics are more responsive to higher rate calls from their own and other species (Randler and Förschler 2011; Wheatcroft 2015), and it is plausible that high call rates promote learning about call structure. Call structure itself can also affect learning. For example, it is harder to train nestling great tits to beg to parental alarm calls than to conspecific or heterospecific songs (Rydén 1978, 1982). Similarly, young Belding’s ground squirrel, Spermophilus beldingi, can discriminate between acoustically different parental alarm calls before emergence from their burrows, but do not treat them as alarm calls until after they have experience outside the burrow (Mateo 1996).
12.5 Eavesdropping, Communication, and Deception
12.5.1 Eavesdropping Facilitates Evolution
Eavesdropping on heterospecific alarm calls, regardless of the mechanism of recognition, sets the stage for further evolution in the structure or use of signals, and potentially reception and use by listeners (Marler 1957; Kostan 2002; Dabelsteen 2005; Searcy and Nowicki 2005). As soon as members of other species respond to alarm calls, there is potentially selection to enhance or suppress their responses by modifying alarm calls or their usage (Kostan 2002; Dabelsteen 2005). If callers are unaffected by eavesdropper responses, their signaling will be unaffected, but if callers benefit, they will be selected to enhance that benefit. By contrast, callers will be selected to make calls more difficult to overhear if eavesdropper responses are costly to callers, which has recently been dubbed “cue denial” (Searcy and Yasukawa 2017). Listeners will be selected to enhance detection and response to alarm calls if they bring benefits, but to ignore such calls if they are deceptive. Here, we consider these scenarios.
12.5.2 Communication
Eavesdroppers and senders could both benefit, potentially leading to interspecific communication, not merely eavesdropping. If mobbing or distress calls prompt members of others species to approach and harass predators, there should be selection to call specifically when heterospecifics are nearby, or to modify calls to make them more recognizable and audible to heterospecifics. The caller gains help in mobbing, the eavesdropper gains information on danger, and collective mobbing is likely to be more effective at deterring predators (Krams and Krama 2002; Krams et al. 2006). Similarly, if heterospecifics flee or become cryptic after hearing aerial alarm calls, the caller could benefit if the predator does not detect any prey and departs the area (Maynard Smith 1965). Alternatively, the caller might reduce risk to itself by prompting all individuals to flee, thereby confusing an attacking predator, diluting the risk of attack, or reducing the risk of being singled out by fleeing alone and becoming the most conspicuous individual (Sherman 1985; Cresswell 1994; Caro 2005; De Vos and O’Riain 2010). Again, both parties are likely to benefit. In these cases, alarm calls might evolve features that make them easy to recognize or learn, which could select for similarities among species’ alarm calls (Marler 1957), or acoustic attributes that prompt attention or facilitate learning (Sects. 12.2.3, 12.4.3, 12.4.4). Finally, individuals may give alarm calls specifically when other species are present, again suggesting active communication. For example, fork-tailed drongos, Dicrurus adsimilis, rarely give alarms to terrestrial predators when alone, but do so when near pied babblers, which are vulnerable to terrestrial threats (Ridley et al. 2007). Drongos benefit in the longer term because they can also steal food from babblers (Sect. 12.5.4).
12.5.3 Cue Denial
If callers suffer a cost from eavesdropping, signal structure and use are likely to evolve to make eavesdropping more difficult. We are not aware of examples involving eavesdropping by prey species, but seeet flee alarm calls have attributes that make them difficult for predators to locate or overhear (Sects. 12.2.2, 12.2.3). Similarly, Diana monkeys suppress calling after detecting chimpanzees, Pan troglodytes, because unlike other predators that abandon hunting when detected, chimpanzees can pursue monkeys through the treetops (Zuberbühler et al. 1997; Zuberbühler 2000a). These cases are the opposite of communication. In communication, signaling evolves from cues to enhance the reception by listeners (Bradbury and Vehrencamp 2011), whereas cue denial hinders reception.
12.5.4 Deception
Turning the tables, eavesdroppers are vulnerable to deception if callers can exploit their responses. Indeed, several species use alarm calls deceptively, when predators are absent, to scare members of other species and steal food (Munn 1986; Møller 1988; Goodale and Kotagama 2005; Flower 2011). In Amazonian mixed-species flocks, white-winged tanager-shrikes, Lanio versicolor, and bluish-slate antshrikes, Thamnomanes schistogyns, act as sentinels in warning of danger. But they also give alarm calls deceptively when competing for a flying insect with another species, which then usually flees to cover (Munn 1986). Similarly, in addition to giving alarm calls to warn pied babblers of danger (Sect. 12.5.2), fork-tailed drongos, Dicrurus adsimilis, use deceptive alarm calls to steal food from both babblers and meerkats (Ridley et al. 2007; Flower 2011; Flower and Gribble 2012). There is therefore a fine balance between cooperation and deception (Ridley et al. 2007; Radford et al. 2011). Indeed, such deception is likely to be effective only if it is not too common. We consider deception and call structure further in the next section on vocal mimicry.
12.6 Vocal Mimicry
12.6.1 Avian Mimicry of Alarm Calls
About 15–20% of songbird species mimic the vocalizations of other species, commonly including their alarm calls (Marshall 1950; Baylis 1982; Kelley et al. 2008; Dalziell et al. 2015). Several species from at least 15 passerine families mimic heterospecific alarm calls (Table 12.1), and they can be given alone or in combination with nonmimetic vocalizations (Goodale and Kotagama 2006; Flower 2011; Wheatcroft and Price 2013; Igic and Magrath 2014; Dalziell and Welbergen 2016). Furthermore, heterospecific alarm calls and predator vocalizations are often mimicked together and may serve similar functions (Chu 2001b; Ratnayake et al. 2010; Kelley and Healy 2011; Goodale et al. 2014b). Mimicry can include a single type of alarm call, such as an aerial or mobbing alarm call (Flower 2011; Wheatcroft and Price 2013), or several types given in the same calling bout (Chu 2001b; Kelley and Healy 2011).
12.6.2 Mimetic Accuracy
The similarity of mimetic alarm calls to the species being copied—termed mimetic accuracy—varies among species, probably for multiple reasons. First, there are likely to be constraints on a mimic’s vocal abilities. For example, superb lyrebirds, Menura novaehollandiae, and northern mockingbirds, Mimus polyglottos, are able to produce almost perfect copies of some heterospecific vocalizations but not of others, probably reflecting species-specific motor abilities (Zollinger and Suthers 2004; Zann and Dunstan 2008; Dalziell and Magrath 2012). Likewise, mimicry of alarm calls varies in accuracy (Flower 2011; Fallow et al. 2013; Igic and Magrath 2013), but it is unknown if this is related to vocal constraints. Second, mimics may only be selected to copy the key features of alarm calls that prompt heterospecific response, whereas other acoustic features might not be copied accurately. For example, brown thornbills, Acanthiza pusilla, produce imperfect copies of heterospecific alarm calls, yet retain the key acoustic features important for provoking alarm response (Fallow et al. 2013; Igic and Magrath 2013). Third, selection for acoustic mimetic accuracy may be frequency dependent, as it is in other mimetic systems (Lindström et al. 1997). This implies that accuracy of specific mimetic alarms will need to be higher when they are relatively more common compared to the heterospecific’s alarm calls. Finally, because not responding to alarm calls could be lethal, listeners are likely to have low thresholds for signal recognition, and so respond to imperfect copies (Sect. 12.4.2).
12.6.3 Function
The function of mimetic alarm calls depends on their target audience, the type of alarm calls mimicked, and the contexts in which they are used (Dalziell et al. 2015). The intended audience of mimetic alarm calls can be conspecifics, other prey species, or predators (Flower 2011; Goodale et al. 2014a; Igic et al. 2015). Mimicry of different types of alarms calls affects function, with aerial alarms scaring or distracting listeners, and mobbing calls attracting them (Flower 2011; Goodale et al. 2014b). Furthermore, mimetic alarm calls can be used in the same context as their respective nonmimetic versions (Morton 1976; Goodale and Kotagama 2006; Wheatcroft and Price 2013), or deceptively in the absence of predators (Flower 2011).
12.6.3.1 Mobbing Predators
One proposed function of mimetic alarm calls is the attraction of heterospecifics to help mob predators. An individual mobbing a predator may benefit from attracting heterospecifics if a larger mobbing group dilutes predation risk to itself (Ficken 1989), or is more effective at causing the predator to depart (Pettifor 1990; Flasskamp 1994). That being the case, mimicry may by-pass the need for heterospecifics to learn to recognize a mimic’s nonmimetic alarm calls, and so ensure response regardless of familiarity (Wheatcroft and Price 2013). Furthermore, individuals sometimes respond less intensely even to familiar heterospecific alarm calls than to conspecific alarm calls (Wheatcroft and Price 2013; Ridley et al. 2014). In such instances, alarm mimicry may act to deceive heterospecifics about the identity of the caller and so increase their mobbing. For example, orange-billed babblers, Turdoides rufescens, and ashy-headed laughing-thrushes, Garrulax cinereifrons, are more likely to mob after hearing mimicry of their own mobbing alarm calls by greater racket-tailed drongos, Dicurus paradiseus, than after hearing the drongo’s own mobbing alarm calls (Goodale et al. 2014b). Mimicry might also be more effective in attracting heterospecific mobbers when a mixture of nonmimetic and mimetic calls evokes stronger mobbing than nonmimetic calls alone (Chu 2001a). More generally, it is possible that mimicry of a multispecies mobbing chorus is more evocative than a single-species mobbing choruses.
12.6.3.2 Deceiving Predators
Mimetic alarm calls can be used deceptively to trick predators. For example, brown thornbills scare a common nest predator by using their own aerial alarm calls in combination with mimicry of heterospecific aerial alarm calls when their nestlings are attacked (Igic and Magrath 2014; Igic et al. 2015; Fig. 12.5). Pied currawongs, Strepera graculina, their major nest predator, respond to playback of these calls as if they themselves are under threat from a flying predator, either scanning the sky for danger or fleeing to a safe location, which could provide the thornbill’s nestlings with an opportunity to escape (Igic et al. 2015). The inclusion of mimetic calls increases the effectiveness of deception, as currawongs respond more strongly to this mixture of calls than to the thornbill’s own alarms. As another example in the same context, female superb lyrebirds mimic both predator calls and alarm calls of local heterospecifics during nest defense (Dalziell and Welbergen 2016). Outside the context of nest defense, mimicry of alarm calls when captured by a predator might startle predators into releasing the mimic (Chu 2001b; Igic and Magrath 2013), a potential function of distress calls (Wise et al. 1999; Neudorf and Sealy 2002).

Brown thornbills, Acanthiza pusilla, use vocal mimicry of alarm calls to deceive predators. (a) (a) New Holland honeyeaters, Phylidonyris novaehollandiae, produce (b) aerial and (c) mobbing calls, and (d) brown thornbills, mimic both the honeyeater’s (e) aerial and (f) mobbing alarm calls. Photographs by Jessica McLachlan. (b) Playback of thornbill alarm call mimicry at artificial thornbill nests scares their major nest predator, the pied currawong, Strepera graculina. Columns show the mean ± standard error delay to resume feeding after playback, and pie charts show the immediate response of currawongs to playback. Modified from Igic et al. (2015)
12.6.3.3 Deception and Kleptoparasitism
Mimetic alarm calls can also be used deceptively to increase the success of stealing food from heterospecifics (see also Sect. 12.5.4). In the best-studied example, fork-tailed drongos deceptively use both their own and mimetic alarm calls to steal food from meerkats and pied babblers (Flower 2011). When a target individual is handling food, drongos produce nonmimetic alarm calls, mimetic alarm calls, or a mixture of the two, despite the absence of a predator (Flower 2011). The victim often drops the food and flees to cover, enabling the drongo to steal it (Flower 2011). Mimicry increases the success of deception by allowing the drongo to switch to a different alarm call when a particular alarm call becomes ineffective at deceiving the target (Flower et al. 2014). Alarm mimicry therefore helps the drongo to avoid frequency-dependent constraints on deception. As noted in Sect. 12.5.4, drongos also increase the efficiency of their deception by acting as sentinels for their targets and producing “true” alarm calls in the presence of predators.
12.6.3.4 Intraspecific Communication and Teaching
Mimicking heterospecific alarm calls and predator vocalizations may help mimics communicate with conspecifics, or teach young about sounds indicating danger. Alarm calls encode general information about predators, such as their type, size, location, and behavior, thereby enabling listeners to choose appropriate responses (Sect. 12.3.2). Mimicry of predator calls could potentially convey even more detail, including the species of predator (Hailman 2009; Billings et al. 2017), which might help listeners locate predators more quickly and respond more effectively. Furthermore, mimicry might help young learn to recognize heterospecific alarm calls or the calls of predators. During nest defense, the parents of some species produce mimetic alarm calls and predator calls in combination with nonmimetic alarm calls (Dalziell et al. 2015), and this may promote learning by offspring to recognize heterospecific calls of danger (Oatley 1969; Goodale et al. 2014a; Potvin et al. 2018). Young could associate these “dangerous” sounds with nonmimetic alarm calls, which young often recognize innately (Magrath et al. 2010).
12.7 Conclusions and Opportunities
Communication is often envisaged as an interaction between two individuals, usually members of the same species. One individual produces a signal that is directed at the other individual and both benefit. The signal has evolved specifically because the sender benefits from the receiver’s response, and the receiver benefits from the information gained from the signal and its subsequent response (Maynard Smith and Harper 2003). But life is more complex, as exemplified by the production and use of alarm calls in natural communities. Alarm calls can be directed at multiple individuals, including conspecifics and heterospecifics; listeners can be the intended recipients or eavesdroppers that listen in on signals intended for others; callers usually benefit from listeners’ responses but may be indifferent or suffer a cost; and listeners usually benefit from their responses to alarm calls, but may be deceived into costly responses.
In this chapter we have summarized research on the information encoded in alarm calls and the responses by heterospecifics. The alarm calls of both conspecifics and heterospecifics are a rich source of information on danger, providing immediate and longer term benefits. Alarm calls are sometimes very similar among species and can share generic acoustic features, and both help to explain recognition by heterospecifics. However, alarm calls can also vary greatly among species, and are often not recognized without learning their association with danger. Individuals are likely to treat heterospecific alarm calls like any other cue of danger but, once they start to respond, there is potentially selection on callers to modify their alarm calls or usage. If callers benefit from eavesdroppers’ responses, they may be selected to enhance signal efficacy, leading to interspecific communication and mutual benefit. Alternatively, callers can be selected to manipulate eavesdroppers, using deceptive signaling and even mimicry, causing the eavesdropper to suffer a cost. If callers suffer a cost from eavesdropper’s responses, their signaling can be modified to make eavesdropping harder, leading to cue denial.
We see many opportunities for research. Despite Marler’s (1955) observations on acoustic form and function in alarm calls, we know little about the evolutionary history of alarm call structure, or the combined importance of acoustic structure and learning in the development of responses to heterospecific alarm calls. Again, despite the celebrated early study of vervet monkey alarm communication and subsequent work on other species (Sect. 12.3.2), we know little about the detail of information gained by heterospecifics. Furthermore, we are only beginning to understand the roles that eavesdropping, communication, and deception play during interspecific interactions. The study of avian vocal mimicry has already provided insights into signal design, and the balance between communication and deception, and will repay further work. Overall, alarm calls provide an excellent model to study the complex web of interspecific interactions.
References
Adkisson CS, Conner RN (1978) Interspecific vocal imitation in white-eyed vireos. Auk 95:602–606
Aubin T (1989) The role of frequency modulation in the process of distress call recognition by the starling (Sturnus vulgaris). Behaviour 108:57–72
Aubin T (1991) Why do distress calls evoke interspecific responses? An experimental study applied to some species of birds. Behav Process 23:103–111
Aubin T, Brémond JC (1989) Parameters used for recognition of distress calls in two species: Larus argentatus and Sturnus vulgaris. Bioacoustics 2:22–33
Baylis JR (1982) Avian vocal mimicry: its function and evolution. In: Kroodsma DE, Miller EH, Ouellet H (eds) Acoustic communication in birds. Academic, New York, pp 51–83
Biedenweg TA, Parsons MH, Fleming PA, Blumstein DT (2011) Sounds scary? Lack of habituation following the presentation of novel sounds. PLoS One 6:e14549
Billings AC, Greene E, MacArthur-Waltz D (2017) Steller’s jays assess and communicate about predator risk using detection cues and identity. Behav Ecol 28:776–783
Blesdoe EK, Blumstein DT (2014) What is the sound of fear? Behavioral responses of white-crowned sparrows Zonotrichia leucophrys to synthesized nonlinear acoustic phenomena. Curr Zool 60:534–541
Blumstein DT, Chi YY (2012) Scared and less noisy: glucocorticoids are associated with alarm call entropy. Biol Lett 8:189–192
Blumstein DT, Munos O (2005) Individual, age and sex-specific information is contained in yellow-bellied marmot alarm calls. Anim Behav 69:353–361
Blumstein DT, Récapet C (2009) The sound of arousal: the addition of novel non-linearities increases responsiveness in marmot alarm calls. Ethology 115:1074–1081
Bradbury JW, Vehrencamp SL (2011) Principles of animal communication, 2nd edn. Sinaur, Sunderland, MA
Branch CL, Freeberg TM (2012) Distress calls in tufted titmice (Baeolophus bicolor): are conspecifics or predators the target? Behav Ecol 23:854–862
Caro TM (2005) Antipredator defenses in birds and mammals. University of Chicago Press, Chicago
Chu M (2001a) Heterospecific responses to scream calls and vocal mimicry by Phainopeplas (Phainopepla nitens) in distress. Behaviour 138:775–787
Chu M (2001b) Vocal mimicry in distress calls of Phainopeplas. Condor 103:389–395
Clarke E, Reichard UH, Zuberbühler K (2006) The syntax and meaning of wild gibbon songs. PLoS One 1:e73
Cresswell W (1994) The function of alarm calls in redshanks, Tringa totanus. Anim Behav 47:736–738
Cunningham S, Magrath RD (2017) Functionally referential alarm calls in noisy miners communicate about predator behaviour. Anim Behav 129:171–179
Curio E, Ernst U, Vieth W (1978) Adaptive significance of avian mobbing II. Cultural transmission of enemy recognition in blackbirds-effectiveness and some constraints. Z Tierpsychol 48:184–202
Dabelsteen T (2005) Public, private or anonymous? Facilitating and countering eavesdropping. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Melbourne, pp 38–62
Dalziell AH, Magrath RD (2012) Fooling the experts: accurate vocal mimicry in the song of the superb lyrebird, Menura novaehollandiae. Anim Behav 83:1401–1410
Dalziell AH, Welbergen JA (2016) Elaborate mimetic vocal displays by female Superb lyrebirds. Front Ecol Evol 4:34
Dalziell AH, Welbergen JA, Igic B, Magrath RD (2015) Avian vocal mimicry: a unified conceptual framework. Biol Rev 90:643–658
Darwin C (1872) The expression of emotions in man and animals, University of Chicago Press, 1965 edn. Murray, London
Dawson Pell FSE, Potvin DA, Ratnayake CP, Fernández-Juricic E, Magrath RD, Radford AN (2018) Birds orient their heads appropriately in response to functionally referential alarm calls of heterospecifics. Anim Behav 140:109–118
De Vos A, O’Riain J (2010) Sharks shape the geometry of a selfish seal herd: experimental evidence from seal decoys. Biol Lett 6:48–50
Dooling R (2004) Audition: can birds hear everything they sing? In: Marler P, Slabbekoorn H (eds) Nature’s music: the science of birdsong. Elsevier, San Diego, pp 206–225
Dutour M, Léna J-P, Lengagne T (2017) Mobbing calls: a signal transcending species boundaries. Anim Behav 131:3–11
Dutour M, Léna J-P, Dumet A, Gardette V, Mondy N, Lengagne T (2019) The role of associative learning process on the response of fledgling great tits (Parus major) to mobbing calls. Anim Cogn 22:1095–1103
Fallow PM, Magrath RD (2010) Eavesdropping on other species: mutual interspecific understanding of urgency information in avian alarm calls. Anim Behav 79:411–417
Fallow PM, Gardner JL, Magrath RD (2011) Sound familiar? Acoustic similarity provokes responses to unfamiliar heterospecific alarm calls. Behav Ecol 22:401–410
Fallow PM, Pitcher BJ, Magrath RD (2013) Alarming features: birds use specific acoustic properties to identify heterospecific alarm calls. Proc R Soc B 280:20122539
Farrow LF, Doohan SJ, McDonald PG (2017) Alarm calls of a cooperative bird are referential and elicit context-specific antipredator behavior. Behav Ecol 28:724–731
Feeney WE, Langmore NE (2013) Social learning of a brood parasite by its host. Biol Lett 9:20130443
Fichtel C, Kappeler PM (2002) Anti-predator behavior of group-living Malagasy primates: mixed evidence for a referential alarm call system. Behav Ecol Sociobiol 51:262–275
Ficken MS (1989) Are mobbing calls of Steller’s jays a “confusion chorus”? J Field Ornithol 60:52–55
Ficken MS, Popp J (1996) A comparative analysis of passerine mobbing calls. Auk 113:370–380
Fitch WT, Neubauer J, Herzel H (2002) Calls out of chaos: the adaptive significance of nonlinear phenomena in mammalian vocal production. Anim Behav 63:407–418
Flasskamp A (1994) The adaptive significance of avian mobbing V. An experimental test of the ‘move on’ hypothesis. Ethology 96:322–333
Flower TP (2011) Fork-tailed drongos use deceptive mimicked alarm calls to steal food. Proc R Soc B 278:1548–1555
Flower TP, Gribble M (2012) Kleptoparasitism by attacks versus false alarm calls in fork-tailed drongos. Anim Behav 83:403–410
Flower TP, Gribble M, Ridley AR (2014) Deception by flexible alarm mimicry in an African bird. Science 344:513–516
Gammon DE, Altizer CE (2011) Northern mockingbirds produce syntactical patterns of vocal mimicry that reflect taxonomy of imitated species. J Field Ornithol 82:158–164
Ghirlanda S, Enquist M (2003) A century of generalization. Anim Behav 66:15–36
Gill SA, Bierema AKM (2013) On the meaning of alarm calls: a review of functional reference in avian alarm calling. Ethology 119:449–461
Gill SA, Sealy SG (1996) Nest defence by yellow warblers: recognition of a brood parasite and an avian nest predator. Behaviour 133:263–282
Goodale E, Kotagama SW (2005) Alarm calling in Sri Lankan mixed-species bird flocks. Auk 122:108–120
Goodale E, Kotagama SW (2006) Context-dependent vocal mimicry in a passerine bird. Proc R Soc B 273:875–880
Goodale E, Kotagama SW (2008) Response to conspecific and heterospecific alarm calls in mixed-species bird flocks of a Sri Lankan rainforest. Behav Ecol 19:887–894
Goodale E, Beauchamp G, Magrath RD, Nieh JC, Ruxton GD (2010) Interspecific information transfer influences animal community structure. Trends Ecol Evol 25:354–361
Goodale E, Ratnayake CP, Kotagama SW (2014a) The frequency of vocal mimicry associated with danger varies due to proximity to nest and nesting stage in a passerine bird. Behaviour 151:73–88
Goodale E, Ratnayake CP, Kotagama SW (2014b) Vocal mimicry of alarm-associated sounds by a drongo elicits flee and mobbing responses from other species that participate in mixed-species bird flocks. Ethology 120:266–274
Greenlaw JS, Shackelford CE, Brown RE (1998) Call mimicry by eastern towhees and its significance in relation to auditory learning. Wilson Bull 110:431–434
Griesser M (2008) Referential calls signal predator behavior in a group-living bird species. Curr Biol 18:69–73
Griffin AS (2004) Social learning about predators: a review and prospectus. Learn Behav 32:131–140
Haff TM, Magrath RD (2012) Learning to listen? Nestling response to heterospecific alarm calls. Anim Behav 84:1401–1410
Haff TM, Magrath RD (2013) Eavesdropping on the neighbours: fledgling response to heterospecific alarm calls. Anim Behav 85:411–418
Hailman JP (2009) Context of blue jay (Cyanocitta cristata) mimicking Cooper’s Hawk (Accipiter cooperii) cackle. Fla Field Nat 37:94–94
Hauser MD (1988) How infant Vervet monkeys learn to recognize starling alarm calls: the role of experience. Behaviour 105:187–201
Hetrick SA, Sieving KE (2012) Antipredator calls of tufted titmice and interspecific transfer of encoded threat information. Behav Ecol 23:83–92
Hindmarsh AM (1984) Vocal mimicry in starlings. Behaviour 90:302–324
Hirth DH, McCullough DR (1977) The evolution of alarm signals in ungulates with special reference to the white-tailed deer. Am Nat 111:31–42
Hollén LI, Radford AN (2009) The development of alarm call behaviour in mammals and birds. Anim Behav 78:791–800
Hurd PL, Wachtmeister C-A, Enquist M (1995) Darwin’s principle of antithesis revisted: a role for perceptual biases in the evolution of intraspecific signals. Proc R Soc B 259:201–205
Igic B, Magrath RD (2013) Fidelity of vocal mimicry: identification and accuracy of mimicry of heterospecific alarm calls by the brown thornbill. Anim Behav 85:593–603
Igic B, Magrath RD (2014) A songbird mimics different heterospecific alarm calls in response to different types of threat. Behav Ecol 25:538–548
Igic B, McLachlam J, Lehtinen I, Magrath RD (2015) Crying wolf to a predator: deceptive vocal mimicry by a bird protecting young. Proc R Soc B 282:20150798
Ito R, Mori A (2010) Vigilance against predators induced by eavesdropping on heterospecific alarm calls in a non-vocal lizard Oplurus cuvieri cuvieri (Reptilia: Iguania). Proc R Soc B 277:1275–1280
Ito R, Ikeuchi I, Mori A (2013) A day gecko darkens its body color in response to avian alarm calls. Curr Herpetol 32:26–33
Johnson FR, McNaughton EJ, Shelly CD, Blumstein DT (2003) Mechanisms of heterospecific recognition in avian mobbing calls. Aust J Zool 51:577–585
Jones KJ, Hill WL (2001) Auditory perception of hawks and owls for passerine alarm calls. Ethology 107:717–726
Jurisevic MA, Sanderson KJ (1994a) Alarm vocalisations in Australian birds: convergent characteristics and phylogenetic differences. Emu 94:69–77
Jurisevic MA, Sanderson KJ (1994b) The vocal repertoires of six honeyeater (Meliphagidae) species from Adelaide, South Australia. Emu 94:141–148
Jurisevic MA, Sanderson KJ (1998) A comparative analysis of distress call structure in Australian passerine and non-passerine species: influence of size and phylogeny. J Avian Biol 29:61–71
Karp D, Manser M, Wiley EM, Townsend SW (2014) Nonlinearities in meerkat alarm calls prevent receivers from habituating. Ethology 120:189–196
Kelley LA, Healy SD (2011) The mimetic repertoire of the spotted bowerbird Ptilonorhynchus maculatus. Naturwissenschaften 98:501–507
Kelley LA, Coe RL, Madden JR, Healy SD (2008) Vocal mimicry in songbirds. Anim Behav 76:521–528
Kirchhof J, Hammerschmidt K (2006) Functionally referential alarm calls in tamarins (Saguinus fuscicollis and Saguinus mystax) – evidence from playback experiments. Ethology 112:346–354
Klump GM, Shalter MD (1984) Acoustic behavior of birds and mammals in the predator context. 1. Factors affecting the structure of alarm signals. 2. The functional-significance and evolution of alarm signals. Z Tierpsychol 66:189–226
Klump GM, Kretzschmar E, Curio E (1986) The hearing of an avian predator and its avian prey. Behav Ecol Sociobiol 18:317–324
Kostan KM (2002) The evolution of mutualistic interspecific communication: assessment and management across species. J Comp Psychol 116:206–209
Krams I, Krama T (2002) Interspecific reciprocity explains mobbing behaviour of the breeding chaffinches, Fringilla coelebs. Proc R Soc B 269:2345–2350
Krams I, Krama T, Igaune K (2006) Mobbing behaviour: reciprocity-based co-operation in breeding pied flycatchers Ficedula hypoleuca. Ibis 148:50–54
Leavesley AJ, Magrath RD (2005) Communicating about danger: urgency alarm calling in a bird. Anim Behav 70:365–373
Lindström L, Alatalo RV, Mappes J (1997) Imperfect Batesian mimicry—the effects of the frequency and the distastefulness of the model. Proc R Soc B 264:149–153
Magrath RD, Bennett T (2012) A micro-geography of fear: learning to eavesdrop on alarm calls of neighbouring heterospecifics. Proc R Soc B 279:902–909
Magrath RD, Pitcher BJ, Gardner JL (2009a) An avian eavesdropping network: alarm signal reliability and heterospecific response. Behav Ecol 20:745–752
Magrath RD, Pitcher BJ, Gardner JL (2009b) Recognition of other species’ aerial alarm calls: speaking the same language or learning another? Proc R Soc B 276:769–774
Magrath RD, Haff TM, Horn A, Leonard ML (2010) Calling in the face of danger: predation risk and acoustic communication by parent birds and their offspring. Adv Study Behav 41:187–253
Magrath RD, Haff TM, Fallow PM, Radford AN (2015a) Eavesdropping on heterospecific alarm calls: from mechanisms to consequences. Biol Rev 90:560–586
Magrath RD, Haff TM, McLachlan JR, Igic B (2015b) Wild birds learn to eavesdrop on heterospecific alarm calls. Curr Biol 25:1–4
Manser MB (2001) The acoustic structure of suricates’ alarm calls varies with predator type and the level of response urgency. Proc R Soc B 268:2315–2324
Manser MB, Bell MB, Fletcher LB (2001) The information that receivers extract from alarm calls in suricates. Proc R Soc B 268:2485–2491
Manser MB, Seyfarth RM, Cheney DL (2002) Suricate alarm calls signal predator class and urgency. Trends Cogn Sci 6:55–57
Marler P (1955) Characteristics of some animal calls. Nature 176:6–8
Marler P (1957) Specific distinctiveness in the communication signals of birds. Behaviour 11:13–37
Marshall AJ (1950) The function of vocal mimicry in birds. Emu 50:5–16
Martínez AE, Zenil RT (2012) Foraging guild influences dependence on heterospecific alarm calls in Amazonian bird flocks. Behav Ecol 23:544–550
Mateo JM (1996) Early auditory experience and the ontogeny of alarm-call discrimination in Belding’s ground squirrels (Spermophilus beldingi). J Comp Psychol 110:115–124
Maynard Smith J (1965) The evolution of alarm calls. Am Nat 99:59–63
Maynard Smith J, Harper D (2003) Animal signals. Oxford University Press, Oxford
McGregor PK, Dabelsteen T (1996) Communication networks. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University press, Ithaca, pp 409–425
McLachlan JR, Ratnayake CP, Magrath RD (2019) Personal information about danger trumps social information from avian alarm calls. Proc R Soc B 286:20182945
McLachlan JR, Magrath RD (2020) Speedy revelations: how alarm calls can convey rapid, reliable information about urgent danger. Proc R Soc B. (in press)
Meise K, Franks DW, Bro-Jørgensen J (2018) Multiple adaptive and non-adaptive processes determine responsiveness to heterospecific alarm calls in African savannah herbivores. Proc R Soc B 285:20172676
Møller AP (1988) False alarm calls as a means of resource usurpation in the great tit Parus major. Ethology 79:25–30
Morton ES (1976) Vocal mimicry in the thick-billed Euphonia. Wilson Bull 88:485–486
Morton ES (1977) On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. Am Nat 111:855–869
Morton ES (2017) Animal vocal communication: assessment and management roles. Cambridge University Press, Cambridge
Müller CA, Manser MB (2008) The information banded mongooses extract from heterospecific alarms. Anim Behav 75:897–904
Munn CA (1986) Birds that ‘cry wolf’. Nature 319:143–145
Murray TG, Magrath RD (2015) Does signal deterioration compromise eavesdropping on other species’ alarm calls? Anim Behav 108:33–41
Naguib M, Wiley RH (2001) Estimating the distance to a source of sound: mechanisms and adaptations for long-range communication. Anim Behav 62:825–837
Naguib M, Mundry R, Ostreiher R, Hultsch H, Schrader L, Todt D (1999) Cooperatively breeding Arabian babblers call differently when mobbing in different predator-induced situations. Behav Ecol 10:636–640
Neudorf DL, Sealy SG (2002) Distress calls of birds in a neotropical cloud forest. Biotropica 34:118–126
Nocera JJ, Ratcliffe LM (2010) Migrant and resident birds adjust antipredator behavior in response to social information accuracy. Behav Ecol 21:121–128
Oatley T (1969) The functions of vocal imitation by African Cossyphas. Ostrich 40:85–89
Oda R, Masataka N (1996) Interspecific responses of ringtailed lemurs to playback of antipredator alarm calls given by Verreaux’s sifakas. Ethology 102:441–453
Parejo D, Avilés JM, Rodríguez J (2012) Alarm calls modulate the spatial structure of a breeding owl community. Proc R Soc B 279:2135–2141
Peake TM (2005) Eavesdropping in communication networks. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 13–37
Pettifor RA (1990) The effects of avian mobbing on a potential predator, the European kestrel, Falco tinnunculus. Anim Behav 39:821–827
Potvin DA, Ratnayake CP, Radford AN, Magrath RD (2018) Birds learn socially to recognize heterospecific alarm calls by acoustic association. Curr Biol 28:2632–2637
Radford AN, Bell MBV, Hollen LI, Ridley AR (2011) Singing for your supper: sentinel calling by kleptoparasites can mitigate the cost to victims. Evolution 65:900–906
Rainey HJ, Züberbuhler K, Slater PJB (2004) Hornbills can distinguish between primate alarm calls. Proc R Soc B 271:755–759
Ramakrishnan U, Coss RG (2000) Recognition of heterospecific alarm vocalizations by bonnet macaques (Macaca radiata). J Comp Psychol 114:3–12
Randler C, Förschler MI (2011) Heterospecifics do not respond to subtle differences in chaffinch mobbing calls: message is encoded in number of elements. Anim Behav 82:725–730
Ratnayake CP, Goodale E, Kotagama SW (2010) Two sympatric species of passerine birds imitate the same raptor calls in alarm contexts. Naturwissenchaften 97:103–108
Rendall D, Owren MJ, Ryan MJ (2009) What do animal signals mean? Anim Behav 78:233–240
Ridley AR, Child MF, Bell MBV (2007) Interspecific audience effects on the alarm-calling behaviour of a kleptoparasitic bird. Biol Lett 3:589–591
Ridley AR, Wiley EM, Thompson AM (2014) The ecological benefits of interceptive eavesdropping. Funct Ecol 28:197–205
Riegert J, Jůzlová Z (2018) Vocal mimicry in the song of Icterine warblers (Hippolais icterina): possible functions and sources of variability. Ethol Ecol Evol 30:430–446
Rooke IJ, Knight TA (1977) Alarm calls of honeyeaters with reference to locating sources of sound. Emu 77:193–198
Rydén O (1978) The significance of antecedent auditory experiences on later reactions to the ‘seeet’ alarm-call in great tit nestlings Parus major. Z Tierpsychol 47:396–409
Rydén O (1982) Selective resistance to approach: a precursor to fear responses to an alarm call in great tit nestlings Parus major. Dev Psychobiol 15:113–120
Schmidt KA, Dall SRX, van Gils JA (2010) The ecology of information: an overview on the ecological significance of making informed decisions. Oikos 119:304–316
Searcy WA, Nowicki S (2005) The evolution of animal communication: reliability and deception in signalling systems. Princeton University Press, Princeton, NJ
Searcy WA, Yasukawa K (2017) Eavesdropping and cue denial in avian acoustic signals. Anim Behav 124:273–282
Seppänen A, Forsman JT (2007) Interspecific social learning: novel preference can be acquired from a competing species. Curr Biol 17:1248–1252
Seppänen JT, Forsman JT, Monkkonen M, Thomson RL (2007) Social information use is a process across time, space, and ecology, reaching heterospecifics. Ecology 88:1622–1633
Seyfarth RM, Cheney DL, Marler P (1980a) Monkey responses to three different alarm calls: evidence of predator classification and semantic communication. Science 210:801–803
Seyfarth RM, Cheney DL, Marler P (1980b) Vervet monkey alarm calls: semantic communication in a free-ranging primate. Anim Behav 28:1070–1094
Sherman PW (1977) Nepotism and the evolution of alarm calls. Science 197:1246–1253
Sherman PW (1985) Alarm calls of Belding’s ground squirrels to aerial predators: nepotism or self-preservation? Behav Ecol Sociobiol 17:313–323
Shriner WM (1999) Antipredator responses to a previously neutral sound by free-living adult golden-mantled ground squirrels, Spermophilus lateralis (Sciuridae). Ethology 105:747–757
Slaughter EI, Berlin ER, Bower JT, Blumstein DT (2013) A test of the nonlinearity hypothesis in great-tailed grackles (Quiscalus mexicanus). Ethology 119:309–315
Snow BK (1974) Vocal mimicry in violaceous Euphonia, Euphonia violacea. Wilson Bull 86:179–180
Sridhar H, Beauchamp G, Shanker K (2009) Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim Behav 78:337–347
Srinivasan U, Raza RH, Quadar S (2010) The nuclear question: rethinking species importance in multi-species animal groups. J Anim Ecol 79:948–954
Sullivan KA (1985) Selective alarm calling by downy woodpeckers in mixed-species flocks. Auk 102:184–187
Suzuki TN (2011) Parental alarm calls warn nestlings about different predatory threats. Curr Biol 21:R15–R16
Suzuki TN (2016) Semantic communication in birds: evidence from field research over the past two decades. Ecol Res 31:307–319
Suzuki TN, Wheatcroft D, Griesser M (2016) Experimental evidence for compositional syntax in bird calls. Nat Commun 7:10986
Tegtman N, Magrath RD (in press) Discriminating between similar alarm calls of contrasting function. Philos Trans R Soc B.
Templeton CN, Greene E (2007) Nuthatches eavesdrop on variations in heterospecific chickadee mobbing alarm calls. Proc Natl Acad Sci USA 104:5479–5482
Templeton CN, Greene E, Davis K (2005) Allometry of alarm calls: black-capped chickadees encode information about predator size. Science 308:1934–1937
ten Cate C, Rowe C (2007) Biases in signal evolution: learning makes a difference. Trends Ecol Evol 22:380–387
Townsend SW, Manser M (2011) The function of nonlinear phenomena in meerkat alarm calls. Biol Lett 7:47–49
Townsend SW, Manser MB (2013) Functionally referential communication in mammals: the past, present and the future. Ethology 119:1–11
Vernon CJ (1973) Vocal imitation by South African birds. Ostrich 44:23–30
Vieth W, Curio E, Ernst U (1980) The adaptive significance of avian mobbing III. Cultural transmission of enemy recognition in blackbirds - cross-species tutoring and properties of learning. Anim Behav 28:1217–1229
Vitousek MN, Adelman JS, Gregory NC, St Clair JJH (2007) Heterospecific alarm call recognition in a non-vocal reptile. Biol Lett 3:632–634
Wheatcroft D (2015) Repetition rate of calls used in multiple contexts communicates presence of predators to nestlings and adult bird. Anim Behav 103:35–44
Wheatcroft D, Price T (2013) Learning and signal copying facilitate communication among bird species. Proc R Soc B 280:20123070
Wheatcroft D, Price TD (2015) Rates of signal evolution are associated with the nature of interspecific communication. Behav Ecol 26:83–90
Wiley RH (2015) Noise matters: the evolution of communication. Harvard, Cambridge, MA
Wiley RH, Richard DG (1982) Adaptations for acoustic communication in birds: sound transmission and signal detection. In: Kroodsma DE, Miller EH, Ouellet H (eds) Acoustic communication in birds, Production, perception, and design features of sound, vol 1. Academic, New York, pp 131–181
Wise KK, Conover MR, Knowlton FF (1999) Response of coyotes to avian distress calls: testing the startle-predator and predator-attraction hypotheses. Behaviour 136:935–949
Zann R, Dunstan E (2008) Mimetic song in superb lyrebirds: species mimicked and mimetic accuracy in different populations and age classes. Anim Behav 76:1043–1054
Zollinger SA, Suthers RA (2004) Motor mechanisms of a vocal mimic: implications for birdsong production. Proc R Soc B 271:483–491
Zuberbühler K (2000a) Causal knowledge of predators’ behaviour in wild Diana monkeys. Anim Behav 59:209–220
Zuberbühler K (2000b) Interspecies semantic communication in two forest primates. Proc R Soc B 267:713–718
Zuberbühler K (2009) Survivor signals: the biology and psychology of animal alarm calling. Adv Study Behav 40:277–322
Zuberbühler K, Noë R, Seyfarth RM (1997) Diana monkey long-distance calls: messages for conspecifics and predators. Anim Behav 53:589–604
Acknowledgments
We thank all our collaborators in our work on communication and eavesdropping in Australian birds. In recent years these have included Lauren Ascah, Tom Bennett, Andrew Cockburn, Sean Cunningham, Francesca Dawson Pell, Anastasia Dalziell, Pam Fallow, Janet Gardner, Esteban Fernández Juricic, Jessica McLachlan, Trevor Murray, Helen Osmond, Dominique Potvin, Andy Radford, Chaminda Ratnayake, Tom Rowell, Natalie Tegtman, You Zhou, and others who have helped in various ways. We are also grateful for financial support from the Australian Research Council and the Research School of Biology, ANU, and to the Australian Bird and Bat Banding Scheme, Australian National Botanic Gardens, and ANU Ethics Committee permission to carry out the work.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Magrath, R.D., Haff, T.M., Igic, B. (2020). Interspecific Communication: Gaining Information from Heterospecific Alarm Calls. In: Aubin, T., Mathevon, N. (eds) Coding Strategies in Vertebrate Acoustic Communication. Animal Signals and Communication, vol 7. Springer, Cham. https://doi.org/10.1007/978-3-030-39200-0_12
Download citation
DOI: https://doi.org/10.1007/978-3-030-39200-0_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-39199-7
Online ISBN: 978-3-030-39200-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)