
Abstract
In the past decade, the facilitated and unlimited access to caloric dense food, in parallel with sedentary lifestyle habits, has contributed to an alarming increase in the incidence and prevalence of non-alcoholic fatty liver disease (NAFLD). NAFLD is currently the most prevalent liver disease worldwide, affecting ~25% of the general population. NAFLD is commonly associated with obesity and the metabolic syndrome, and includes a wide spectrum of liver lesions, from simple steatosis to non-alcoholic steatohepatitis (NASH), eventually progressing to liver cirrhosis, with increased risk for development of hepatocellular carcinoma (HCC). Still, NAFLD pathogenesis remains unsolved. In parallel, disease diagnosis and stratification of patients remains challenging. In this regard, extracellular vesicles (EVs), which consist of submicron membrane-bound structures secreted from different cell types with important functions in cell-to-cell communication and found in a wide variety of body fluids, are emerging as key players in NAFLD pathogenesis and as promising disease biomarkers. In this chapter, we will focus on the most recent findings concerning the role of EVs as new structures mediating autocrine and paracrine intercellular communication in the liver, as well as new diagnostic tools for NAFLD.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Non-alcoholic fatty liver disease (NAFLD) represents a common condition characterized by the pathologic accumulation of fat within more than 5% of hepatocytes, in the absence of other forms of liver disease, such as viral infections or excessive alcohol intake, among others [1]. NAFLD encompasses a spectrum of liver lesions ranging from simple steatosis to non-alcoholic steatohepatitis (NASH). Hepatic or simple steatosis is usually benign and considered one of the earliest and less severe stages of NAFLD, exhibiting a relatively favorable clinical course. In about 5–10% of patients, hepatic steatosis may progress to NASH, a more malignant state that is prone to further progress to cirrhosis and hepatocellular carcinoma (HCC), substantially contributing to liver-related morbidity and mortality [1].
NAFLD is currently the most prevalent liver disease worldwide, affecting around one-fourth of the general population [2]. In fact, several NAFLD epidemiological studies have been conducted in both the United States and Europe, indicating that over than 64 million people in the United States are believed to possess NAFLD [3,4,5]. In Europe, the prevalence of NAFLD varies between countries. In Spain, a 2010 multicenter cross-sectional population study revealed that the overall prevalence of NAFLD was 25.8% [6]. In Romania, the estimated prevalence was 20% [7] while a Greek study stated that 31.3 and 39.8% of liver biopsies from 498 individuals showed hepatic steatosis or NASH, respectively [8]. In this regard, it was predicted that over 52 million individuals would be affected by NAFLD in Germany, France, Italy and the United Kingdom [3]. Although NAFLD incidence is increasing worldwide, it is most noticeable in specific population groups, particularly those with metabolic comorbidities. NAFLD prevalence is increased in patients with type 2 diabetes, who also display an increased risk for development of NASH [9, 10]. Two major European studies reported a NAFLD prevalence rate between 42.6 and 69.5% in patients with type 2 diabetes [11, 12]. Obese individuals are also more likely to develop NAFLD. In particular, 65.7% of obese patients undergoing bariatric surgery have NAFLD, with 33.6% also evidencing NASH [13]. Overall, patients with metabolic syndrome, pathological conditions characterized by abdominal obesity, hypertension, dyslipidemia and glucose intolerance are at increased risk of developing NAFLD [14].
NAFLD is thought to increase cardiovascular risk, as most NAFLD patients die from cardiovascular-related problems [15,16,17,18,19]. Indeed, the number of NAFLD patients with advanced disease (i.e., cirrhosis, end-stage liver disease, HCC), a major cause of liver disease-related morbidity and mortality, is alarmingly increasing [20, 21]. Both liver-specific and overall mortality among NAFLD patients is estimated at 0.77 and 11.77 per 1000 person-year, respectively, further increasing in patients with NASH (15.44 and 25.56 per 1000 person-year, respectively) [2]. Of note, NASH is currently the second indication for liver transplantation among all chronic liver diseases and is expected to become the leading indication for transplantation in the next decades [22, 23]. In parallel with NAFLD pathogenesis remaining incompletely understood, one of the major clinical challenges is the difficulty of obtaining differential diagnosis between the different disease severity stages, and also the identification of patients that might be at higher risk for disease progression. As such, there is a substantial unmet need for novel non-invasive and accurate tools that might allow the diagnosis and risk stratification of NAFLD patients. In this regard, extracellular vesicles (EVs) are emerging as promising molecular targets and biomarkers, being involved in disease pathogenesis and harboring diagnostic and prognostic potential. In this chapter, we will review the most recent findings concerning the role of EVs in NAFLD pathogenesis and in diagnosis/prognosis.
NASH Diagnosis and Monitoring: Current Approaches
Liver biopsy is still the gold standard procedure to undoubtedly identify and stage NAFLD. Kleiner and Blunt proposed a scoring system, named the NAFLD Activity Score (NAS), which is calculated by the unweighted sum of the scores for steatosis (0–3), lobular inflammation (0–3) and hepatocyte ballooning (0–2) [24]. Consequently, NAS ranges from 0 to 8 and a total score of 0–2 is considered a non-NASH diagnosis while scores greater or equal to 5 are diagnosed as NASH. Of note, fibrosis was not included as a component of the activity score due to its irreversible nature and because it was thought to only result from disease progression. Still, fibrosis is often present in NASH; to that matter, the Steatosis Activity Fibrosis (SAF) score, which does not sub-classifies NAFLD based solely in NASH, was proposed. This score evaluates three variables: steatosis, on a scale of 0–3 (S0: <5%, S1: 5–33%, S2: 34–66%, S3: >67%); ballooning and lobular inflammation, each graded between 0 and 2 and then summed, representing activity (A0–A4), and fibrosis, scored between 0 and 4 (F0: none, F1: perisinusoidal or periportal, F2: perisinusoidal and periportal, F3: bridging or F4: cirrhosis) [25]. This new score was shown to decrease intra-observer variation among pathologists and includes the fibrosis component in the final decision which, although not required for the diagnosis of NASH, is now considered to represent the best predictor of advanced liver disease and mortality [26]. Nonetheless, the SAF score still requires a liver biopsy to be performed, carrying several intrinsic limitations, including invasiveness, poor acceptability, variability and cost [27]. Therefore, alternative non-invasive strategies have been proposed in order to achieve a more accurate diagnosis and to assist in the management of NAFLD patients [27, 28]. For instance, the Hepamet Fibrosis Score (HFS), mainly based on serum markers, was recently developed an validated in a cohort of 2453 patients with NAFLD, presenting area under the receiver operating characteristic (AUC) values greater than other fibrosis score systems (the NAFLD fibrosis score [NFS] and FIB-4) [29]. Currently, 2 different complementary approaches are being used as non-invasive methods in the diagnosis and management of NAFLD: a “biological” approach, which mainly relies on the quantification of serum biomarkers, and a “physical” approach, related with the measurement of the intrinsic physical properties of the liver parenchyma (liver stiffness) by different imaging techniques [27].
Although the vast majority of NAFLD patients are clinically asymptomatic, approximately 20% display elevated liver enzymes [30]. Serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) may present normal to moderate elevations—about 1.5–2 times the upper limit—although these are considered poor markers of fatty liver disease [31, 32]. Several predictive models (SteatoTest [33], Fatty Liver Index [34], Hepatic Steatosis Index [35], lipid accumulation product [36], the Index of NASH [37] or the NAFLD Liver Fat Score [38], based on the differential combination of serum transaminases, triglyceride levels and clinical information have been developed (reviewed in [39]) to help in the diagnosis of steatosis. It should be noted that diagnostic performances of each test are difficult to compare since they are based against different standards, namely liver biopsy or imaging techniques. Still, in a cohort of 324 patients with suspected NAFLD and liver biopsy, the Fatty Liver Index, NAFLD Fat Score and Hepatic Steatosis Index were retrospectively evaluated, providing similar AUC values (0.83, 0.80 and 0.81, respectively) [40]. However, these scores are still not widely used as they fail to provide substantial additional information when compared with current clinical, laboratory and imaging studies, and await further validation studies. Cytokeratin-18 (CK18) fragments have been extensively studied as biomarkers for NASH diagnosis. CK18 fragments are released from apoptotic hepatocytes and the caspase-cleaved fragment M30 is detected by enzyme-linked immunosorbent assay (ELISA) in the serum of patients with NASH, with a AUC value of 0.83 [41]. In two subsequent meta-analyses, the pooled AUC for CK18 to predict NASH was 0.82 (median sensitivity and specificity of 66–78% and 82–87%, respectively) [42, 43]. Nonetheless, there are still several drawbacks with regards to using CK18 as a diagnosis biomarker for NASH, restraining its translation into clinical practice. In particular, there is significant variability in the suggested cutoffs and diagnostic accuracy among studies [44], no commercially available clinical tests [45] and a rather limited sensitivity when used alone [46]. On this note, the diagnostic accuracy of CK18 is increased when used in combination with sFas levels [47], uric acid [48], adiponectin and resistin [49, 50], ALT or the presence of metabolic syndrome [51], among others. Some models have been proposed but most of them were studied in small and highly selected populations (morbidly obese patients) and, as such, need to be further validated. Serum metabolomics was also shown to be of great value in the diagnosis of NAFLD/NASH and recent studies have reported specific serum metabolomic signatures that allowed the specific diagnosis of NAFLD (OWL Liver Care) and the differential diagnosis of steatosis and NASH (OWL Liver Test) [52, 53] as well as between NASH subtypes [54], that are currently commercialized by OWL Metabolomics.
Imaging techniques, either ultrasound- or magnetic resonance-based, are widely used in NAFLD diagnosis. Conventional ultrasonography is currently the most commonly used imaging technique for the diagnosis of steatosis due to its general availability [27]. With ordinal ultrasonography scores, steatosis may be subjectively categorized as mild, moderate and severe [55, 56] Importantly, in a large meta-analysis, including 34 studies and 2815 patients with suspected/diagnosed liver diseases, pooled sensitivities and specificities of ultrasonography to differentiate moderate/severe fatty liver and absence of steatosis, were 85% and 93%, respectively [57]. Still, in daily practice, ultrasonography is only used to provide a diagnosis of presence or absence of steatosis, only being capable of detecting liver fat amounts greater than 2.5–20% [58] and displaying lower accuracy in obese patients with concomitant renal disease [59, 60]. According to the European Guidelines for the management of NAFLD [20], ultrasonography constitutes the first choice imaging technique to monitor steatosis in adults at risk for disease development. Controlled Attenuation Parameter (CAP) is a new non-invasive transient elastography (TE)-based imaging technique for the staging of steatosis. Good inter-observer reproducibility was reported (concordance rates between observers of 0.82–0.84) [55, 56], and in a cohort of patients with chronic liver diseases (15% with NAFLD), steatosis was accurately detected by CAP. Still, this technique was not precise enough to discern between the different stages of steatosis [61]. Furthermore, CAP determinations might be influenced by the presence of covariates, such as the body mass index or diabetes [27]. Finally, magnetic resonance-based imaging techniques have been reported as the most accurate available imaging techniques to quantify liver fat and fibrosis. Magnetic resonance imaging proton density fat fraction (MRI-PDFF) is highly accurate (AUC: 0.950) [62], and reproducible, fast, and allows for evaluation of the entire liver, quantification of fat content, and stratification of steatosis (with excellent sensitivity for detecting mild steatosis) [63]. This technique has been validated in several studies [64, 65] and is emerging as the gold standard for the quantification of liver fat. However, it is not widely available, it is rather expensive and measurements are presumed to be affected by food intake [66]. Novel methodologies, including point shear wave elastography (pSWE) and 2-dimensional shear wave elastography (2D-SWE) are now being evaluated [67,68,69].
Of note, the differential diagnosis of simple steatosis and NASH by imaging techniques remains challenging, as MRI-PDFF cannot effectively detect liver inflammation and ballooning, nor NASH resolution or fibrosis improvements [70]. The same holds true for magnetic resonance elastography (MRE) [71,72,73,74,75] and TE [72, 76,77,78].
Overall, there is still no biomarker or imaging method capable of accurately diagnosing, staging and performing the follow-up of NAFLD (including fibrosis) for which new approaches are eagerly awaited. In this regard, EVs are emerging as novel potential NAFLD biomarkers, while also participating in disease pathogenesis.
Extracellular Vesicles
EVs are a heterogeneous population of membrane vesicles ranging from 30 nm to 2 μM in diameter, secreted by diverse cell types and containing distinct biomolecules, including proteins, nucleic acids and lipids [79,80,81,82]. The first reports on EVs considered them membrane debris with no biological significance, and a way to eliminate needless compounds by the cells [83]. However, new evidence demonstrating their potential to stimulate adaptative immune responses [84, 85], opened their role in intercellular communication. In the last decade, this is an emerging field which is exponentially increasing, with special interest in their capacity to exchange components between cells and acting as signaling vehicles (Fig. 9.1).

Timeline (1955–2019) of articles referring to extracellular vesicles, microvesicles and exosomes in PubMed. (https://www.ncbi.nlm.nih.gov/pubmed/). Accession date: December 2019 Abbreviations: EV extracellular vesicle; MV microvesicle
Based on the current knowledge of their biogenesis, EVs are classified as exosomes, microvesicles (MV) and apoptotic bodies (Fig. 9.2) [79,80,81,82, 85, 86]. Exosomes (30–200 nm) are formed as intraluminal vesicles (ILV) by the inward budding of the endosomal membrane during maturation of multivesicular endosomes (MVE). They are secreted from the lumen of late endosomes, also called multivesicular bodies (MVBs), to the extracellular space by fusion of MVBs with the plasma membrane. Microvesicles (50–1000 nm) are generated by the outward budding and fission of the plasma membrane and the subsequent release of vesicles into the extracellular space. Finally, apoptotic bodies (800–5000 nm) are released by cells when plasma membrane blebbing occurs during programmed cell death. All EVs may contain cytoplasmic proteins, lipid raft-interacting proteins, membrane proteins, lipids, metabolites, DNA and different types of RNA, including mRNAs, microRNAs (miRNAs) and other non-coding RNAs (ncRNAs) [85, 87]. Their molecular cargo varies widely among cell types and conditions (e.g., physiology/pathology), directly affecting the fate and function of these membrane vesicles [80]. EVs are found in all biological fluids, including serum, plasma, urine, saliva, and bile, among others, as well as in culture supernatants [81, 82]. However, despite their different biogenesis, once they reach the extracellular zone, exosomes and microvesicles display a similar appearance, overlapping in size, and often presenting a common biomolecular composition [80]. Therefore, it is difficult to ascertain the origin of EVs when they are isolated from the extracellular medium or from diverse biological fluids.

Biogenesis, overall composition and release of EVs. MVs are produced by the outward budding and fission of the plasma membrane, whereas exosomes are formed by the inward budding of the multivesicular body and released upon fusion of multi-vesicular bodies with the plasma membrane. EVs are lipid-bilayer membrane vesicles which contain cytoplasmic proteins, lipid raft-interacting proteins, membrane proteins, lipids, metabolites, DNA and different types of RNA. Abbreviations: ER endoplasmic reticulum; MVB multivesicular body
A wide variety of methods have been proposed for the isolation of EVs from extracellular fluids: differential centrifugation/ultracentrifugation, flotation on density gradients, separation by size exclusion chromatography, precipitation with different polymers, filtration and antibody-based purification (immuno-affinity) [79, 88]. These methods allow the separation of EVs from non-vesicular entities, such as protein aggregates, lipoparticles, viruses and cell debris, with different rates of success [80]. Given the heterogeneous EV population, each purification method will result in enrichment of specific EV subpopulations, with distinct recovery/specificity rates [88]. Nonetheless, combination of multiple isolation procedures is capable of specifically separating subpopulations of vesicles based on their size, density, surface proteins, sugar, lipid composition or other biophysical properties, such as surface charge. Even so, considering that a single optimal separation method or a gold standard is not yet defined, the isolation method should be chosen based on the downstream application and on the scientific question that is being addressed [79, 88]. Once EVs are obtained, they should be properly characterized using multiple, complementary techniques, in order to obtain reproducible results. In this regard, the International Society for Extracellular Vesicles (ISEV) regularly publishes minimal requirement guidelines for the study of EVs, focusing on adequate and standardized characterization [79, 88]. In parallel, a consortium called EV-TRACK (transparent reporting and centralizing knowledge in extracellular vesicle research) has been gathered to build a crowdsourcing knowledgebase (http://evtrack.org/) that centralizes EV biology and methodology with the goal of stimulating authors, reviewers, editors and funders to put experimental guidelines into practice [89].
The characterization of EVs should include determination of its morphology, size and concentration, as well as reporting of the components typically associated with EVs, particularly membrane proteins [79, 88]. Scanning electron microscopy (SEM), transmission electron microscopy (TEM), cryo-electon microscopy (cryo-EM) and atomic force microscopy (AFM) images enable the analysis of EV morphology and size, while also providing information on the heterogeneity of the EV preparation. Particle number and size can be measured quantitatively by analyzing large numbers of single EVs with light scattering technologies, such as nanoparticle tracking analysis (NTA) and dynamic light scattering (DLS), as well as with high resolution flow cytometry (hrFC) or tunable resistive pulse sensing (TRPS). The analysis of EV-associated proteins is typically performed through immunoblotting, flow cytometry and/or mass spectrometry. It has been described that transmembrane or glycosylphosphatidylinositol (GPI)-anchored proteins localized in plasma membrane and/or endosomes, such as tetraspanins (CD63, CD81, others) or integrins, may be considered as markers of any type of EVs, as their presence demonstrates the existence of the lipid-bilayer structure characteristic of EVs. Additionally, cytosolic proteins with membrane-binding ability, such as ESCRT-I/II/III (e.g., TSG101), heat shock proteins, ALIX and ARF6 are also commonly found in EVs, given the nature of its biogenesis. Besides proteins, phospholipids found in lipid bilayers are also potential positive controls for identifying EVs. Still, these might be non-specific, as other particles such as lipoproteins can also contain phospholipids. It remains important to clarify the ratios of cholesterol, sphingomyelin, ceramide, and phosphatidylcholine/ethanolamine/inositol found in EVs, and how these differ from the ratios found in lipoproteins. In fact, there are still no markers capable of accurately distinguishing between every different EV subtype [80, 85, 88].
The mechanisms involved in EV-mediated cell-to-cell communication are vast and still incompletely understood. When interacting with target cells, EVs may modulate cellular signaling pathways in a pleiotropic manner, including the direct activation of cell surface receptors via protein and bioactive lipid ligands, or by merging their membrane contents into the recipient cell [80]. This type of communication is believed to occur in both physiological conditions as well as in pathological states [85]. In physiological conditions, EVs participate in the maintenance of stemness [90], tissue repair [91], blood coagulation [92], immune surveillance [93], neuronal plasticity [94] and several other physiological functions [85]. In turn, EVs may contribute to tumorigenesis by inducing abnormal cell proliferation [95], stimulating tumor growth [96], promoting extracellular matrix remodeling [97], and facilitating tumor metastasis [98] and immune escape [93]. Beyond cancer, EVs appear to also play a role in the spread of different pathogens, in the local propagation of neurodegenerative diseases, and in several liver diseases, including NAFLD (reviewed in [99, 100]).
Role of Extracellular Vesicles in NAFLD Pathogenesis
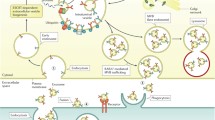
As central mediators of cell-to-cell communication, EVs have recently arisen as novel players in NAFLD pathogenesis and progression (Fig. 9.3; Table 9.1). Data from different diet-induced animal models of NASH have shown that EV concentration increases with disease progression, in a time-dependent manner [101,102,103]. This may result from accumulation of lipotoxic lipids and their downstream mediators in the liver, already shown to increase the capacity of hepatocytes to form and release different types of EVs [102,103,104,105]. EVs can then be internalized by macrophages, neutrophils and monocytes, leading to their activation and recruitment to the liver, promoting and exacerbating the inflammatory responses observed in NASH. In fact, palmitic acid (PA) and lysophosphatidylcholine were shown to increase the release of microvesicles carrying TNF-related apoptosis inducing ligand (TRAIL) ligand from both mouse and human hepatocytes [105]. In mice, this promotes the expression of pro-inflammatory cytokines IL-1β and IL-6 in bone marrow-derived macrophages, in a rho-associated, coiled-coil-containing protein kinase 1 (ROCK1)-dependent manner [105]. Of note, administration of fasudil, a ROCK1 inhibitor, to mice with NASH decreases the amount of EVs in the serum as well as liver injury, inflammation and fibrosis. PA-stimulated hepatocytes have also been shown to release EVs enriched in ceramide, which increase macrophage recruitment to the liver via sphingosine-1-phosphate (S1P). Indeed, increased levels of ceramide- and S1P-enriched EVs were reported in plasma from both mice and patients with NASH [102]. Importantly, in mice fed a high-fructose, -saturated fat and -cholesterol diet, blocking of S1P was shown to improve liver histology, namely reducing hepatocyte ballooning and inflammatory foci, in parallel with reduced hepatomegaly, serum transaminases and accumulation of hepatic macrophages [106]. Finally, EVs carrying C-X-C motif ligand 10 (CXCL10), a potent chemokine that is released by lipotoxic hepatocytes in a Mixed Lineage Kinase 3 (MLK3)-dependent manner, have also been shown to associate with macrophage recruitment [104]. In addition, the majority of mitochondrial DNA (mtDNA) released from hepatocytes circulate within microparticles and promote activation of neutrophils and kupffer cells through toll-like receptor 9 (TLR9) [107]. More recently, it was shown that the NLRP3 inflammasome is activated by microvesicles released from fat-laden cells undergoing lipotoxicity, either in hepatocytes and macrophages, further reinforcing the role of EVs in disease progression from simple steatosis to NASH [108]. On that note, the transplant of circulating EVs from high fat-fed mice to chow-fed mice was shown to induce accumulation and consequent activation of myeloid cells in the liver, thus promoting liver inflammation and injury [109].

Role of EVs in NAFLD pathogenesis. EVs are released by lipotoxic hepatocytes, thus contributing to the recruitment and activation of macrophages, Kupffer and stellate cell activation, as well as angiogenesis, through targeting of endothelial cells. Hepatic stellate cells and cholangiocytes might also secrete EVs, thus also contributing for disease progression. In addition, adipose tissue- and gut-derived EVs are also known to target the liver and contribute to NAFLD. Finally, increased blood concentration of EVs might help in NAFLD diagnosis. Abbreviations: EVs extracellular vesicles
Injured hepatocytes are able to communicate with other liver cell types, such as hepatic stellate cells (HSCs), thus triggering the expression of fibrosis-related genes [110, 111]. In fact, EVs released from PA-stimulated hepatocytes have been shown to enhance the expression of fibrosis markers in HSCs. Among its cargo, these EVs were shown to carry several miRNAs, including miR-192, already associated with NAFLD progression and liver fibrosis [110]. Interestingly, incubation of HSC cultures with plasma EVs from mice fed a high-fat diet, triggered their activation, again underscoring EVs as key cell-to-cell communication mediators. In addition, in a dietary murine model of NASH, mice treated with thiazolidinediones, a group of insulin sensitizers that activate peroxisome proliferator-activated receptors (PPARs), exhibited reduced secretion of EVs from hepatocytes, thereby impairing HSCs activation [111]. This emphasizes the crucial role of PPARs inhibition, especially PPAR-γ, in the phenotypical switch of HSCs from quiescent to their active form. In agreement, hepatocyte-derived EVs released during lipotoxicity are enriched with miR-128-3p, a miRNA that direct targets PPAR-γ and promote HSCs migration, proliferation and activation, therefore contributing to fibrosis and NAFLD progression [112].
Hepatocytes are not the only source of EVs in the liver that actively contribute to this type of paracrine communication. In fact, both mouse and human HSCs release EVs that target hepatocytes and HSCs themselves [113,114,115]. For instance, it was shown that under fibrotic conditions, HSCs release EVs carrying lower levels of miR-214, when compared with physiological situations, which directly target connective tissue growth factor 2 (CCN2), a pivotal activator of HSCs, thereby promoting fibrosis [113]. Curiously, CCN2 itself can be present within HSCs-derived EVs, thus enhancing expression of several pro-fibrotic genes in quiescent HSCs, contributing to their activation [114]. On the other hand, EVs secreted by quiescent HSCs were shown to carry high levels of Twist1, a transcription factor that binds to the miR-214 promoter, increasing its expression. In turn, miR-214 expression is stimulated in receptor cells, thus suppressing the expression CCN2 and its downstream effectors [115].
Current evidence indicates that accumulation of fat in the adipose tissue actively contributes to hepatic steatosis, mainly through the release of adipokines and free fatty acids (FFAs) into circulation, which will lastly end up in the liver [116]. EVs, more specifically exosomes, have already been implicated in this process, representing an important component of the adipose tissue-liver axis. Visceral adipose tissue-derived exosomes obtained from obese patients were shown to dysregulate pro-fibrogenic transforming growth factor-β (TGF-β)-related pathways in both hepatocytes and HSCs [117]. Furthermore, exosomal miRNAs isolated from human and mice adipose tissue constitute the majority of circulating exosomal miRNAs [118]. Of note, under physiologic conditions, exosomal miR-99b targets the hepatic fibroblast growth factor-21 (FGF-21) and represses its expression. Interestingly, only exosomal, but not free miR-99b, is able to regulate FGF-21 in the liver [118]. Finally, transfer of exosomes from adipose-derived stem cells (ADSC) to obese mice improved insulin resistance and reduced obesity, in parallel with reduction of hepatic steatosis. Furthermore, the authors reported that ADSC-derived exosomes induced a M2 anti-inflammatory phenotype when transferred into macrophages, a process that was dependent on the activation of arginase-1 by STAT3-carrying exosomes [119]. In parallel with the adipose tissue-liver axis, several studies support the notion of a gut-liver axis in the pathogenesis of several liver diseases [120]. In fact, high-fat diet-induced dysbiosis in mice leads to the release of gut-derived exosomes carrying injury-related high mobility group box 1 (HMGB1) protein, thus contributing to the development and progression of hepatic steatosis [121]. Last but not least, EVs have also been reported to have pro-angiogenic properties in the liver through targeting of endothelial cells [103, 122]. In particular, hepatocytes undergoing lipoapoptosis release microvesicles exhibiting Vanin-1 on its surface, thus allowing interaction with lipid raft domains of endothelial cells, resulting in cell migration and tube formation [103]. Interestingly, treating primary rat endothelial cells with plasma-derived exosomes from high-fat diet-fed rats markedly increased oxidative stress and the expression of vascular cell adhesion molecule 1 (VCAM1), thus contributing to a pro-inflammatory and pro-angiogenic environment [123]. In addition, exosomes carrying Sonic Hedgehog (Shh) and Indian Hedgehog ligands (Ihh), released by HSCs and cholangiocytes, engage a pro-angiogenic switch in endothelial cells during cirrhosis [122]. However, the role of these EVs in angiogenesis in the NAFLD context needs further exploration.
Extracellular Vesicles as Non-invasive Biomarkers for NAFLD
It is now widely established that circulating EVs are remarkably stable, thus constituting promising non-invasive biomarkers [100]. A pioneer study on this field compared the EV blood profile from patients with simple steatosis (NAFL) or NASH, to patients with chronic hepatitis C (CHC) or healthy controls. In order to ascertain the possible cells of origin, the authors measured the presence of EVs with leuko-endothelial surface markers by flow cytometry [124]. Serum EVs derived from CD4+ and CD8+ T cells were found increased in patients with NAFL/NASH and CHC, compared to healthy individuals, but they were unable to specifically differentiate between these disease conditions (AUC: 0.57 and 0.65, respectively). On the other hand, patients with NAFL/NASH displayed marked increases in serum EVs containing surface markers from invariant natural killer T cells (iNKT) and monocytes/macrophages (CD14+), as well as lower levels of neutrophil- (CD15+) and endothelial cell- (CD41+) derived EVs. Noteworthy, CD14+- and iNKT-derived EVs positively correlated with serum ALT levels and NAS score, and allowed the differential diagnosis of NAFL/NASH and CHC (AUC:0.999 and 0.97, respectively). Importantly, these two type of EV populations were reported to be key players in liver fibrosis during NAFLD pathogenesis [125, 126]. Further, the release of EVs from immune cells might be involved in liver inflammation and, consequently, in the progression of NAFL to NASH. The levels of adipose tissue-derived EVs in obese patients also correlate with the levels of liver transaminases and were shown to contribute to insulin resistance, interfering with the insulin signaling pathway in hepatocytes [127]. C16:0 ceramide- and S1P-enriched EVs might also embody promising diagnostic biomarkers for NALFD/NASH, since they were shown to progressively increase in the plasma of obese patients with simple steatosis, and further in NASH patients with early fibrosis (F1), when compared with control obese patients [102]. However, these findings were obtained from a small cohort of patients (n = 43) and their diagnostic accuracy remains incompletely explored. As such, further studies, including larger cohorts of patients, should ideally be performed. Specifically concerning fibrosis, a previous study found that CD14+ and CD16+ EVs count could predict fibrosis severity, being inversely associated with NAFLD-related liver fibrosis, while also increasing the diagnostic capability of the enhanced liver fibrosis score (LFS) in patients with NAFLD (AUC: 0.948 and 0.967 for CD14+ and CD16+ EVs, respectively, vs. 0.915 for LFS alone) [128].
Increased concentration of serum EVs has been described in dietary murine models of NASH. In mice fed a choline-deficient L-amino acid defined (CDAA) diet, EV levels were shown to increase early in disease progression, further increasing with time and correlating with hepatocyte cell death, fibrosis and neo-angiogenesis [101]. Furthermore, proteomic analysis of blood EVs from CDAA-fed mice revealed a distinct protein cargo, when compared with EVs isolated from control mice, with most of the identified proteins being already described as players in NASH pathogenesis, namely affecting cell death and inflammatory pathways, among others. Although no AUC values were reported, the authors stated that this proteomic signature allowed for the discrimination of diseased mice compared to controls. In addition, miR-122, a liver-specific miRNA, was found enriched in blood EVs from mice fed a CDAA diet compared to controls, while its hepatic levels were reduced, pinpointing for a potential diagnostic capability of miR-122-EVs for NAFLD [101]. However, future validation studies should be performed, in order to clearly assess its accuracy for NAFLD. Similarly, increased amounts of hepatocyte-derived EVs were detected in blood from diet-induced NASH mice, and found to correlate with disease severity [103]. In a similar model, endothelial-derived EVs (CD144+) were found increased, an effect that could be reverted by treatment with atorvastatin [129]. EV-derived hepatocyte mtDNA is also increased in the plasma of both mice and human patients with NASH, contributing to the activation of the TLR9 pathway, and the activation of sterile inflammation [107]. Nevertheless, the mechanisms by which mtDNA is targeted in EVs, as well as the accuracy of these particles to diagnose NAFLD remains to be clarified.
Conclusions and Future Directions
In the past decade, the number of reports addressing the role of EVs in human disease has grown exponentially. Particularly for liver diseases, including NAFLD, hundreds of papers have already been published, showcasing promising results that might translate into the clinics in a near future. Nonetheless, several aspects still need to be addressed before this jump can be made, particularly those related with methodological aspects. For instance, many studies use the terms “exosome” and “microvesicles” indiscriminately, without proper characterization of the isolated EV fraction. Further, considering that EV biogenesis is not entirely understood, and purification protocols are not homogeneous, many inconsistencies are still found in the literature [79, 80, 85]. In order to bypass these problems, and aid in the rapid translation of EVs into the clinics, standardized, large-scale and cost-effective protocols are urgently needed. In this regard, the EV-TRACK knowledgebase constitutes a key resource that researchers should consult in order to standardize research in this filed and increase reproducibility of EV-related reports [80].
NAFLD is a complex metabolic multisystem and multicellular disease, involving extra-hepatic organs and several cell types in liver, which encompasses different degrees of autocrine, paracrine and endocrine communication. Although the investigation on this filed is still scarce and inconclusive, there is still much to discover, it is now clear that inter-cell and inter-organ communication in NAFLD might be of pivotal important and is mediated in part, by circulating EVs. Mostly due to the accumulation of toxic lipid species within hepatocytes, these liver cells are currently considered as one of the major sources of EVs in NAFLD, consubstantiating a key mechanism for disease progression into more nefarious stages. Although the role of EVs in NAFLD pathogenesis is unquestionable, there are still unsolved questions that should be addressed in the future: what are the major EV contents that directly contributes for disease progression? And how can we specifically target these EVs? Furthermore, deeply studying the molecular mechanisms underlying EV biogenesis will contribute with key concepts that will hopefully allow the manipulation of EV generation in patients, thus opening a new window for therapeutic interventions. Still, this idea should be approached with caution, as manipulation of the machinery involved in EV biogenesis might hold potential secondary effects on healthy tissues [80]. In this regard, the role of EVs as biomarkers for NAFLD is probably more close to make this translational jump. In fact, many reports have already illustrated the potential of these vesicles to act as either diagnostic or prognostic biomarkers. In order to advance the field, future studies should assess the diagnostic value of EVs in larger cohorts of patients, including properly characterized individuals (biopsy-proven). Further, stratifying patients according to their metabolic status (presence/absence of diabetes, obesity, hypercholesterolemia, among others), as well as to the presence/absence of fibrosis, might reveal interesting and could provide decisive results. Lastly, it is widely known that patients with advanced NASH are at higher risk of progressing to HCC. It will also be important to query whether EVs might help in in the prediction of patients who might be at risk for experiencing disease progression to HCC. In this regard, we have recently described a specific proteomic profile in serum EVs that allowed the specific diagnosis of HCC, when compared with healthy controls or patients with intrahepatic cholangiocarcinoma [130]. It will now be imperative to conduct studies in order to address the diagnostic capacity of these EVs, in this context.
In the next decade, several EV-related studies are envisioned in the NAFLD field, which might contribute with new concepts that will help in deciphering disease pathogenesis and possibly provide new diagnostic and prognostic tools to be applied into daily clinics [81, 85, 131]. The wide-range cellular and biological functions of EVs, as well as their ability of encapsulating and protecting biological and artificial therapeutic compounds, support the idea that EVs and their components may also be used as novel therapeutic targets, therapeutic agents and/or drug delivery vehicles to treat NAFLD.
Abbreviations
- 2D-SWE:
-
2-dimensional shear wave elastography
- ADSCs:
-
Adipose-derived stem cells
- AFM:
-
Atomic force microscopy
- ALT:
-
Alanine aminotransferase
- AST:
-
Aspartate aminotransferase
- AUC:
-
Area under the receiver operating curve
- cryo-EM:
-
Cryo-electron microscopy
- CAP:
-
Controlled attenuation parameter
- CCN2:
-
Connective tissue growth factor 2
- CDAA:
-
Choline-deficient L-amino acid defined
- CHC:
-
Chronic hepatitis C
- CK18:
-
Cytokeratin 18
- CXCL10:
-
C-X-C motif ligand 10
- DLS:
-
Dynamic light scattering
- ELISA:
-
Enzyme-linked immunosorbent assay
- EVs:
-
Extracellular vesicles
- FFAs:
-
Free fatty acids
- FGF21:
-
Fibroblast growth factor 21
- GPI:
-
Glycosylphosphatidylinositol
- hrFC:
-
High-resolution flow cytometer
- HCC:
-
Hepatocellular carcinoma
- HFS:
-
Hepamet fibrosis score
- HMGB1:
-
High mobility group box 1
- HSCs:
-
Hepatic stellate cells
- iNKT:
-
Invariant natural killer T cells
- Ihh:
-
Indian hedgehog
- ILVs:
-
Intraluminal vesicles
- ISEV:
-
International society for extracellular vesicles
- LFS:
-
Liver fibrosis score
- miRNAs:
-
microRNAs
- mtDNA:
-
Mitochondrial DNA
- MLK3:
-
Mixed lineage kinase 3
- MRE:
-
Magnetic resonance elastography
- MRI-PDFF:
-
Magnetic resonance imaging-derived proton density fat fraction
- MVBs:
-
Multivesicular bodies
- MVEs:
-
Multivesicular endosomes
- MVs:
-
Microvesicles
- ncRNAs:
-
Non-coding RNAs
- NAFL:
-
Non-alcoholic fatty liver
- NAFLD:
-
Non-alcoholic fatty liver disease
- NAS:
-
NAFLD activity score
- NASH:
-
Non-alcoholic steatohepatitis
- NFS:
-
NAFLD fibrosis score
- NTA:
-
Nanoparticle tracking analysis
- pSWE:
-
Point shear wave elastography
- PA:
-
Palmitic acid
- PPARs:
-
Peroxisome proliferator-associated receptors
- ROCK1:
-
Rho-associated, coiled-coil-containing protein kinase 1
- S1P:
-
Spingosine-1-phosphate
- SAF:
-
Steatosis activity fibrosis
- SEM:
-
Scanning electron microscopy
- Shh:
-
Sonic hedgehog
- TE:
-
Transient elastography
- TEM:
-
Transmission electron microscopy
- TGFβ:
-
Transforming growth factor β
- TLR9:
-
Toll-like receptor 9
- TRAIL:
-
TNF-related apoptosis inducing ligand
- TRPS:
-
Tunable resistive pulse sensing
- VCAM1:
-
Vascular cell adhesion molecule 1
References
Cohen JC, Horton JD, Hobbs HH. Human fatty liver disease: old questions and new insights. Science. 2011;332:1519–23.
Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M. Global epidemiology of nonalcoholic fatty liver disease—meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology. 2016;64:73–84.
Younossi ZM, Blissett D, Blissett R, Henry L, Stepanova M, Younossi Y, et al. The economic and clinical burden of nonalcoholic fatty liver disease in the United States and Europe. Hepatology. 2016;64:1577–86.
Younossi ZM. Non-alcoholic fatty liver disease—a global public health perspective. J Hepatol. 2019;70:531–44.
Younossi ZM, Tampi R, Priyadarshini M, Nader F, Younossi IM, Racila A. Burden of illness and economic model for patients with nonalcoholic steatohepatitis in the United States. Hepatology. 2019;69:564–72.
Caballeria L, Pera G, Auladell MA, Toran P, Munoz L, Miranda D, et al. Prevalence and factors associated with the presence of nonalcoholic fatty liver disease in an adult population in Spain. Eur J Gastroenterol Hepatol. 2010;22:24–32.
Radu C, Grigorescu M, Crisan D, Lupsor M, Constantin D, Dina L. Prevalence and associated risk factors of non-alcoholic fatty liver disease in hospitalized patients. J Gastrointestin Liver Dis. 2008;17:255–60.
Zois CD, Baltayiannis GH, Bekiari A, Goussia A, Karayiannis P, Doukas M, et al. Steatosis and steatohepatitis in postmortem material from Northwestern Greece. World J Gastroenterol. 2010;16:3944–9.
Mishra A, Younossi ZM. Epidemiology and natural history of non-alcoholic fatty liver disease. J Clin Exp Hepatol. 2012;2:135–44.
Leite NC, Villela-Nogueira CA, Pannain VL, Bottino AC, Rezende GF, Cardoso CR, et al. Histopathological stages of nonalcoholic fatty liver disease in type 2 diabetes: prevalences and correlated factors. Liver Int. 2011;31:700–6.
Williamson RM, Price JF, Glancy S, Perry E, Nee LD, Hayes PC, et al. Prevalence of and risk factors for hepatic steatosis and nonalcoholic Fatty liver disease in people with type 2 diabetes: the Edinburgh type 2 diabetes study. Diabetes Care. 2011;34:1139–44.
Targher G, Bertolini L, Padovani R, Rodella S, Tessari R, Zenari L, et al. Prevalence of nonalcoholic fatty liver disease and its association with cardiovascular disease among type 2 diabetic patients. Diabetes Care. 2007;30:1212–8.
Flegal KM, Carroll MD, Kit BK, Ogden CL. Prevalence of obesity and trends in the distribution of body mass index among US adults, 1999-2010. JAMA. 2012;307:491–7.
Machado MV, Cortez-Pinto H. Management of fatty liver disease with the metabolic syndrome. Expert Rev Gastroenterol Hepatol. 2014;8:487–500.
Adams LA, Lymp JF, St Sauver J, Sanderson SO, Lindor KD, Feldstein A, et al. The natural history of nonalcoholic fatty liver disease: a population-based cohort study. Gastroenterology. 2005;129:113–21.
Anstee QM, Targher G, Day CP. Progression of NAFLD to diabetes mellitus, cardiovascular disease or cirrhosis. Nat Rev Gastroenterol Hepatol. 2013;10:330–44.
Lonardo A, Sookoian S, Chonchol M, Loria P, Targher G. Cardiovascular and systemic risk in nonalcoholic fatty liver disease—atherosclerosis as a major player in the natural course of NAFLD. Curr Pharm Des. 2013;19:5177–92.
Stahl EP, Dhindsa DS, Lee SK, Sandesara PB, Chalasani NP, Sperling LS. Nonalcoholic Fatty liver disease and the heart: JACC state-of-the-art review. J Am Coll Cardiol. 2019;73:948–63.
Lonardo A, Nascimbeni F, Mantovani A, Targher G. Hypertension, diabetes, atherosclerosis and NASH: cause or consequence? J Hepatol. 2018;68:335–52.
European Association for the Study of the L, European Association for the Study of D, European Association for the Study of O. EASL-EASD-EASO clinical practice guidelines for the management of non-alcoholic fatty liver disease. J Hepatol. 2016;64:1388–402.
Chalasani N, Younossi Z, Lavine JE, Charlton M, Cusi K, Rinella M, et al. The diagnosis and management of nonalcoholic fatty liver disease: practice guidance from the American Association for the Study of Liver Diseases. Hepatology. 2018;67:328–57.
Wong RJ, Aguilar M, Cheung R, Perumpail RB, Harrison SA, Younossi ZM, et al. Nonalcoholic steatohepatitis is the second leading etiology of liver disease among adults awaiting liver transplantation in the United States. Gastroenterology. 2015;148:547–55.
Goldberg D, Ditah IC, Saeian K, Lalehzari M, Aronsohn A, Gorospe EC, et al. Changes in the prevalence of hepatitis C virus infection, nonalcoholic steatohepatitis, and alcoholic liver disease among patients with cirrhosis or liver failure on the waitlist for liver transplantation. Gastroenterology. 2017;152:1090–9. e1091.
Kleiner DE, Brunt EM, Van Natta M, Behling C, Contos MJ, Cummings OW, et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology. 2005;41:1313–21.
Bedossa P, Consortium FP. Utility and appropriateness of the fatty liver inhibition of progression (FLIP) algorithm and steatosis, activity, and fibrosis (SAF) score in the evaluation of biopsies of nonalcoholic fatty liver disease. Hepatology. 2014;60:565–75.
Angulo P, Kleiner DE, Dam-Larsen S, Adams LA, Bjornsson ES, Charatcharoenwitthaya P, et al. Liver fibrosis, but no other histologic features, is associated with long-term outcomes of patients with nonalcoholic fatty liver disease. Gastroenterology. 2015;149:389–97. e310.
Castera L, Friedrich-Rust M, Loomba R. Noninvasive assessment of liver disease in patients with nonalcoholic fatty liver disease. Gastroenterology. 2019;156:1264–81. e1264.
European Association for Study of L, Asociacion Latinoamericana para el Estudio del H. EASL-ALEH Clinical Practice Guidelines. Non-invasive tests for evaluation of liver disease severity and prognosis. J Hepatol. 2015;63:237–64.
Ampuero J, Pais R, Aller R, Gallego-Duran R, Crespo J, Garcia-Monzon C, et al. Development and validation of hepamet fibrosis scoring system—a simple, non-invasive test to identify patients with nonalcoholic fatty liver disease with advanced fibrosis. Clin Gastroenterol Hepatol. 2019. https://doi.org/10.1016/j.cgh.2019.05.051. [Epub ahead of print]
Gao X, Fan JG, Study Group of L, Metabolism CSoE. Diagnosis and management of non-alcoholic fatty liver disease and related metabolic disorders: consensus statement from the Study Group of Liver and Metabolism, Chinese Society of Endocrinology. J Diabetes. 2013;5:406–15.
Attar BM, Van Thiel DH. Current concepts and management approaches in nonalcoholic fatty liver disease. ScientificWorldJournal. 2013;2013:481893.
Kotronen A, Westerbacka J, Bergholm R, Pietilainen KH, Yki-Jarvinen H. Liver fat in the metabolic syndrome. J Clin Endocrinol Metab. 2007;92:3490–7.
Poynard T, Ratziu V, Naveau S, Thabut D, Charlotte F, Messous D, et al. The diagnostic value of biomarkers (SteatoTest) for the prediction of liver steatosis. Comp Hepatol. 2005;4:10.
Bedogni G, Bellentani S, Miglioli L, Masutti F, Passalacqua M, Castiglione A, et al. The Fatty Liver Index: a simple and accurate predictor of hepatic steatosis in the general population. BMC Gastroenterol. 2006;6:33.
Lee JH, Kim D, Kim HJ, Lee CH, Yang JI, Kim W, et al. Hepatic steatosis index: a simple screening tool reflecting nonalcoholic fatty liver disease. Dig Liver Dis. 2010;42:503–8.
Bedogni G, Kahn HS, Bellentani S, Tiribelli C. A simple index of lipid overaccumulation is a good marker of liver steatosis. BMC Gastroenterol. 2010;10:98.
Otgonsuren M, Estep MJ, Hossain N, Younossi E, Frost S, Henry L, et al. Single non-invasive model to diagnose non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). J Gastroenterol Hepatol. 2014;29:2006–13.
Kotronen A, Peltonen M, Hakkarainen A, Sevastianova K, Bergholm R, Johansson LM, et al. Prediction of non-alcoholic fatty liver disease and liver fat using metabolic and genetic factors. Gastroenterology. 2009;137:865–72.
Stern C, Castera L. Non-invasive diagnosis of hepatic steatosis. Hepatol Int. 2017;11:70–8.
Fedchuk L, Nascimbeni F, Pais R, Charlotte F, Housset C, Ratziu V, et al. Performance and limitations of steatosis biomarkers in patients with nonalcoholic fatty liver disease. Aliment Pharmacol Ther. 2014;40:1209–22.
Feldstein AE, Wieckowska A, Lopez AR, Liu YC, Zein NN, McCullough AJ. Cytokeratin-18 fragment levels as noninvasive biomarkers for nonalcoholic steatohepatitis: a multicenter validation study. Hepatology. 2009;50:1072–8.
Kwok R, Tse YK, Wong GL, Ha Y, Lee AU, Ngu MC, et al. Systematic review with meta-analysis: non-invasive assessment of non-alcoholic fatty liver disease—the role of transient elastography and plasma cytokeratin-18 fragments. Aliment Pharmacol Ther. 2014;39:254–69.
Musso G, Gambino R, Cassader M, Pagano G. Meta-analysis: natural history of non-alcoholic fatty liver disease (NAFLD) and diagnostic accuracy of non-invasive tests for liver disease severity. Ann Med. 2011;43:617–49.
Vilar-Gomez E, Chalasani N. Non-invasive assessment of non-alcoholic fatty liver disease: clinical prediction rules and blood-based biomarkers. J Hepatol. 2018;68:305–15.
Younossi ZM, Loomba R, Anstee QM, Rinella ME, Bugianesi E, Marchesini G, et al. Diagnostic modalities for nonalcoholic fatty liver disease, nonalcoholic steatohepatitis, and associated fibrosis. Hepatology. 2018;68:349–60.
Cusi K, Chang Z, Harrison S, Lomonaco R, Bril F, Orsak B, et al. Limited value of plasma cytokeratin-18 as a biomarker for NASH and fibrosis in patients with non-alcoholic fatty liver disease. J Hepatol. 2014;60:167–74.
Tamimi TI, Elgouhari HM, Alkhouri N, Yerian LM, Berk MP, Lopez R, et al. An apoptosis panel for nonalcoholic steatohepatitis diagnosis. J Hepatol. 2011;54:1224–9.
Huang JF, Yeh ML, Huang CF, Huang CI, Tsai PC, Tai CM, et al. Cytokeratin-18 and uric acid predicts disease severity in Taiwanese nonalcoholic steatohepatitis patients. PLoS One. 2017;12:e0174394.
Younossi ZM, Jarrar M, Nugent C, Randhawa M, Afendy M, Stepanova M, et al. A novel diagnostic biomarker panel for obesity-related nonalcoholic steatohepatitis (NASH). Obes Surg. 2008;18:1430–7.
Younossi ZM, Page S, Rafiq N, Birerdinc A, Stepanova M, Hossain N, et al. A biomarker panel for non-alcoholic steatohepatitis (NASH) and NASH-related fibrosis. Obes Surg. 2011;21:431–9.
Anty R, Iannelli A, Patouraux S, Bonnafous S, Lavallard VJ, Senni-Buratti M, et al. A new composite model including metabolic syndrome, alanine aminotransferase and cytokeratin-18 for the diagnosis of non-alcoholic steatohepatitis in morbidly obese patients. Aliment Pharmacol Ther. 2010;32:1315–22.
Mayo R, Crespo J, Martinez-Arranz I, Banales JM, Arias M, Minchole I, et al. Metabolomic-based noninvasive serum test to diagnose nonalcoholic steatohepatitis: results from discovery and validation cohorts. Hepatol Commun. 2018;2:807–20.
Barr J, Vazquez-Chantada M, Alonso C, Perez-Cormenzana M, Mayo R, Galan A, et al. Liquid chromatography-mass spectrometry-based parallel metabolic profiling of human and mouse model serum reveals putative biomarkers associated with the progression of nonalcoholic fatty liver disease. J Proteome Res. 2010;9:4501–12.
Alonso C, Fernandez-Ramos D, Varela-Rey M, Martinez-Arranz I, Navasa N, Van Liempd SM, et al. Metabolomic identification of subtypes of nonalcoholic steatohepatitis. Gastroenterology. 2017;152:1449–61. e1447.
Ballestri S, Lonardo A, Romagnoli D, Carulli L, Losi L, Day CP, et al. Ultrasonographic fatty liver indicator, a novel score which rules out NASH and is correlated with metabolic parameters in NAFLD. Liver Int. 2012;32:1242–52.
Hamaguchi M, Kojima T, Itoh Y, Harano Y, Fujii K, Nakajima T, et al. The severity of ultrasonographic findings in nonalcoholic fatty liver disease reflects the metabolic syndrome and visceral fat accumulation. Am J Gastroenterol. 2007;102:2708–15.
Hernaez R, Lazo M, Bonekamp S, Kamel I, Brancati FL, Guallar E, et al. Diagnostic accuracy and reliability of ultrasonography for the detection of fatty liver: a meta-analysis. Hepatology. 2011;54:1082–90.
Bril F, Ortiz-Lopez C, Lomonaco R, Orsak B, Freckleton M, Chintapalli K, et al. Clinical value of liver ultrasound for the diagnosis of nonalcoholic fatty liver disease in overweight and obese patients. Liver Int. 2015;35:2139–46.
de Moura Almeida A, Cotrim HP, Barbosa DB, de Athayde LG, Santos AS, Bitencourt AG, et al. Fatty liver disease in severe obese patients: diagnostic value of abdominal ultrasound. World J Gastroenterol. 2008;14:1415–8.
Mottin CC, Moretto M, Padoin AV, Swarowsky AM, Toneto MG, Glock L, et al. The role of ultrasound in the diagnosis of hepatic steatosis in morbidly obese patients. Obes Surg. 2004;14:635–7.
Sasso M, Beaugrand M, de Ledinghen V, Douvin C, Marcellin P, Poupon R, et al. Controlled attenuation parameter (CAP): a novel VCTE guided ultrasonic attenuation measurement for the evaluation of hepatic steatosis: preliminary study and validation in a cohort of patients with chronic liver disease from various causes. Ultrasound Med Biol. 2010;36:1825–35.
Idilman IS, Aniktar H, Idilman R, Kabacam G, Savas B, Elhan A, et al. Hepatic steatosis: quantification by proton density fat fraction with MR imaging versus liver biopsy. Radiology. 2013;267:767–75.
Chan WK, Nik Mustapha NR, Wong GL, Wong VW, Mahadeva S. Controlled attenuation parameter using the FibroScan(R) XL probe for quantification of hepatic steatosis for non-alcoholic fatty liver disease in an Asian population. United European Gastroenterol J. 2017;5:76–85.
Romeo S, Kozlitina J, Xing C, Pertsemlidis A, Cox D, Pennacchio LA, et al. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat Genet. 2008;40:1461–5.
Caussy C, Soni M, Cui J, Bettencourt R, Schork N, Chen CH, et al. Nonalcoholic fatty liver disease with cirrhosis increases familial risk for advanced fibrosis. J Clin Invest. 2017;127:2697–704.
Liu K, Wong VW, Lau K, Liu SD, Tse YK, Yip TC, et al. Prognostic value of controlled attenuation parameter by transient elastography. Am J Gastroenterol. 2017;112:1812–23.
Dietrich CF, Bamber J, Berzigotti A, Bota S, Cantisani V, Castera L, et al. EFSUMB guidelines and recommendations on the clinical use of liver ultrasound elastography, update 2017 (long version). Ultraschall Med. 2017;38:e48.
Friedrich-Rust M, Poynard T, Castera L. Critical comparison of elastography methods to assess chronic liver disease. Nat Rev Gastroenterol Hepatol. 2016;13:402–11.
Ferraioli G, Wong VW, Castera L, Berzigotti A, Sporea I, Dietrich CF, et al. Liver ultrasound elastography: an update to the world federation for ultrasound in medicine and biology guidelines and recommendations. Ultrasound Med Biol. 2018;44:2419–40.
Caussy C, Reeder SB, Sirlin CB, Loomba R. Noninvasive, quantitative assessment of liver fat by MRI-PDFF as an endpoint in NASH trials. Hepatology. 2018;68:763–72.
Sporea I, Bota S, Peck-Radosavljevic M, Sirli R, Tanaka H, Iijima H, et al. Acoustic radiation force impulse elastography for fibrosis evaluation in patients with chronic hepatitis C: an international multicenter study. Eur J Radiol. 2012;81:4112–8.
Park CC, Nguyen P, Hernandez C, Bettencourt R, Ramirez K, Fortney L, et al. Magnetic resonance elastography vs transient elastography in detection of fibrosis and noninvasive measurement of steatosis in patients with biopsy-proven nonalcoholic fatty liver disease. Gastroenterology. 2017;152:598–607. e592.
Chen J, Talwalkar JA, Yin M, Glaser KJ, Sanderson SO, Ehman RL. Early detection of nonalcoholic steatohepatitis in patients with nonalcoholic fatty liver disease by using MR elastography. Radiology. 2011;259:749–56.
Loomba R, Wolfson T, Ang B, Hooker J, Behling C, Peterson M, et al. Magnetic resonance elastography predicts advanced fibrosis in patients with nonalcoholic fatty liver disease: a prospective study. Hepatology. 2014;60:1920–8.
Loomba R, Cui J, Wolfson T, Haufe W, Hooker J, Szeverenyi N, et al. Novel 3D magnetic resonance elastography for the noninvasive diagnosis of advanced fibrosis in NAFLD: a prospective study. Am J Gastroenterol. 2016;111:986–94.
Siddiqui MS, Vuppalanchi R, Van Natta ML, Hallinan E, Kowdley KV, Abdelmalek M, et al. Vibration-controlled transient elastography to assess fibrosis and steatosis in patients with nonalcoholic fatty liver disease. Clin Gastroenterol Hepatol. 2019;17:156–63. e152.
Lee CM, Jeong WK, Lim S, Kim Y, Kim J, Kim TY, et al. Diagnosis of clinically significant portal hypertension in patients with cirrhosis: splenic arterial resistive index versus liver stiffness measurement. Ultrasound Med Biol. 2016;42:1312–20.
Imajo K, Kessoku T, Honda Y, Tomeno W, Ogawa Y, Mawatari H, et al. Magnetic resonance imaging more accurately classifies steatosis and fibrosis in patients with nonalcoholic fatty liver disease than transient elastography. Gastroenterology. 2016;150:626–37. e627.
Lotvall J, Hill AF, Hochberg F, Buzas EI, Di Vizio D, Gardiner C, et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International Society for Extracellular Vesicles. J Extracell Vesicles. 2014;3:26913.
van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19:213–28.
Lapitz A, Arbelaiz A, Olaizola P, Aranburu A, Bujanda L, Perugorria MJ, et al. Extracellular vesicles in hepatobiliary malignancies. Front Immunol. 2018;9:2270.
Yanez-Mo M, Siljander PR, Andreu Z, Zavec AB, Borras FE, Buzas EI, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066.
Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33:967–78.
Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV, Melief CJ, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183:1161–72.
S ELA, Mager I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12:347–57.
Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200:373–83.
Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–89.
Thery C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7:1535750.
Consortium E-T, Van Deun J, Mestdagh P, Agostinis P, Akay O, Anand S, et al. EV-TRACK: transparent reporting and centralizing knowledge in extracellular vesicle research. Nat Methods. 2017;14:228–32.
Ratajczak J, Miekus K, Kucia M, Zhang J, Reca R, Dvorak P, et al. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia. 2006;20:847–56.
Bjorge IM, Kim SY, Mano JF, Kalionis B, Chrzanowski W. Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine—a new paradigm for tissue repair. Biomater Sci. 2017;6:60–78.
Del Conde I, Shrimpton CN, Thiagarajan P, Lopez JA. Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood. 2005;106:1604–11.
Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol. 2014;14:195–208.
Budnik V, Ruiz-Canada C, Wendler F. Extracellular vesicles round off communication in the nervous system. Nat Rev Neurosci. 2016;17:160–72.
Takasugi M, Okada R, Takahashi A, Virya Chen D, Watanabe S, Hara E. Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nat Commun. 2017;8:15729.
Skog J, Wurdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, et al. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10:1470–6.
Mu W, Rana S, Zoller M. Host matrix modulation by tumor exosomes promotes motility and invasiveness. Neoplasia. 2013;15:875–87.
Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell. 2016;30:836–48.
Zhang X, Ji X, Wang Q, Li JZ. New insight into inter-organ crosstalk contributing to the pathogenesis of non-alcoholic fatty liver disease (NAFLD). Protein Cell. 2018;9:164–77.
Eguchi A, Feldstein AE. Extracellular vesicles in non-alcoholic and alcoholic fatty liver diseases. Liver Res. 2018;2:30–4.
Povero D, Eguchi A, Li H, Johnson CD, Papouchado BG, Wree A, et al. Circulating extracellular vesicles with specific proteome and liver microRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS One. 2014;9:e113651.
Kakazu E, Mauer AS, Yin M, Malhi H. Hepatocytes release ceramide-enriched pro-inflammatory extracellular vesicles in an IRE1alpha-dependent manner. J Lipid Res. 2016;57:233–45.
Povero D, Eguchi A, Niesman IR, Andronikou N, de Mollerat du Jeu X, Mulya A, et al. Lipid-induced toxicity stimulates hepatocytes to release angiogenic microparticles that require Vanin-1 for uptake by endothelial cells. Sci Signal. 2013;6:ra88.
Ibrahim SH, Hirsova P, Tomita K, Bronk SF, Werneburg NW, Harrison SA, et al. Mixed lineage kinase 3 mediates release of C-X-C motif ligand 10-bearing chemotactic extracellular vesicles from lipotoxic hepatocytes. Hepatology. 2016;63:731–44.
Hirsova P, Ibrahim SH, Krishnan A, Verma VK, Bronk SF, Werneburg NW, et al. Lipid-induced signaling causes release of inflammatory extracellular vesicles from hepatocytes. Gastroenterology. 2016;150:956–67.
Mauer AS, Hirsova P, Maiers JL, Shah VH, Malhi H. Inhibition of sphingosine 1-phosphate signaling ameliorates murine nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver Physiol. 2017;312:G300–13.
Garcia-Martinez I, Santoro N, Chen Y, Hoque R, Ouyang X, Caprio S, et al. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J Clin Invest. 2016;126:859–64.
Cannito S, Morello E, Bocca C, Foglia B, Benetti E, Novo E, et al. Microvesicles released from fat-laden cells promote activation of hepatocellular NLRP3 inflammasome: a pro-inflammatory link between lipotoxicity and non-alcoholic steatohepatitis. PLoS One. 2017;12:e0172575.
Deng ZB, Liu Y, Liu C, Xiang X, Wang J, Cheng Z, et al. Immature myeloid cells induced by a high-fat diet contribute to liver inflammation. Hepatology. 2009;50:1412–20.
Lee YS, Kim SY, Ko E, Lee JH, Yi HS, Yoo YJ, et al. Exosomes derived from palmitic acid-treated hepatocytes induce fibrotic activation of hepatic stellate cells. Sci Rep. 2017;7:3710.
McCommis KS, Hodges WT, Brunt EM, Nalbantoglu I, McDonald WG, Holley C, et al. Targeting the mitochondrial pyruvate carrier attenuates fibrosis in a mouse model of nonalcoholic steatohepatitis. Hepatology. 2017;65:1543–56.
Povero D, Panera N, Eguchi A, Johnson CD, Papouchado BG, de Araujo Horcel L, et al. Lipid-induced hepatocyte-derived extracellular vesicles regulate hepatic stellate cell via microRNAs targeting PPAR-gamma. Cell Mol Gastroenterol Hepatol. 2015;1:646–63. e644.
Chen L, Charrier A, Zhou Y, Chen R, Yu B, Agarwal K, et al. Epigenetic regulation of connective tissue growth factor by MicroRNA-214 delivery in exosomes from mouse or human hepatic stellate cells. Hepatology. 2014;59:1118–29.
Charrier A, Chen R, Chen L, Kemper S, Hattori T, Takigawa M, et al. Exosomes mediate intercellular transfer of pro-fibrogenic connective tissue growth factor (CCN2) between hepatic stellate cells, the principal fibrotic cells in the liver. Surgery. 2014;156:548–55.
Chen L, Chen R, Kemper S, Charrier A, Brigstock DR. Suppression of fibrogenic signaling in hepatic stellate cells by Twist1-dependent microRNA-214 expression: role of exosomes in horizontal transfer of Twist1. Am J Physiol Gastrointest Liver Physiol. 2015;309:G491–9.
Liu W, Baker RD, Bhatia T, Zhu L, Baker SS. Pathogenesis of nonalcoholic steatohepatitis. Cell Mol Life Sci. 2016;73:1969–87.
Koeck ES, Iordanskaia T, Sevilla S, Ferrante SC, Hubal MJ, Freishtat RJ, et al. Adipocyte exosomes induce transforming growth factor beta pathway dysregulation in hepatocytes: a novel paradigm for obesity-related liver disease. J Surg Res. 2014;192:268–75.
Thomou T, Mori MA, Dreyfuss JM, Konishi M, Sakaguchi M, Wolfrum C, et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature. 2017;542:450–5.
Zhao H, Shang Q, Pan Z, Bai Y, Li Z, Zhang H, et al. Exosomes from adipose-derived stem cells attenuate adipose inflammation and obesity through polarizing M2 macrophages and beiging in white adipose tissue. Diabetes. 2018;67:235–47.
Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology. 2010;52:1836–46.
Chen Y, Sun H, Bai Y, Zhi F. Gut dysbiosis-derived exosomes trigger hepatic steatosis by transiting HMGB1 from intestinal to liver in mice. Biochem Biophys Res Commun. 2019;509:767–72.
Witek RP, Yang L, Liu R, Jung Y, Omenetti A, Syn WK, et al. Liver cell-derived microparticles activate hedgehog signaling and alter gene expression in hepatic endothelial cells. Gastroenterology. 2009;136:320–30. e322.
Heinrich LF, Andersen DK, Cleasby ME, Lawson C. Long-term high fat feeding of rats results in increased numbers of circulating microvesicles with pro-inflammatory effects on endothelial cells. Br J Nutr. 2015;113:1704–11.
Kornek M, Lynch M, Mehta SH, Lai M, Exley M, Afdhal NH, et al. Circulating microparticles as disease-specific biomarkers of severity of inflammation in patients with hepatitis C or nonalcoholic steatohepatitis. Gastroenterology. 2012;143:448–58.
Karlmark KR, Weiskirchen R, Zimmermann HW, Gassler N, Ginhoux F, Weber C, et al. Hepatic recruitment of the inflammatory Gr1+ monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology. 2009;50:261–74.
Syn WK, Oo YH, Pereira TA, Karaca GF, Jung Y, Omenetti A, et al. Accumulation of natural killer T cells in progressive nonalcoholic fatty liver disease. Hepatology. 2010;51:1998–2007.
Kranendonk ME, Visseren FL, van Herwaarden JA, Nolte-‘t Hoen EN, de Jager W, Wauben MH, et al. Effect of extracellular vesicles of human adipose tissue on insulin signaling in liver and muscle cells. Obesity (Silver Spring). 2014;22:2216–23.
Welsh JA, Scorletti E, Clough GF, Englyst NA, Byrne CD. Leukocyte extracellular vesicle concentration is inversely associated with liver fibrosis severity in NAFLD. J Leukoc Biol. 2018;104:631–9.
Ajamieh H, Farrell GC, McCuskey RS, Yu J, Chu E, Wong HJ, et al. Acute atorvastatin is hepatoprotective against ischaemia-reperfusion injury in mice by modulating eNOS and microparticle formation. Liver Int. 2015;35:2174–86.
Arbelaiz A, Azkargorta M, Krawczyk M, Santos-Laso A, Lapitz A, Perugorria MJ, et al. Serum extracellular vesicles contain protein biomarkers for primary sclerosing cholangitis and cholangiocarcinoma. Hepatology. 2017;66:1125–43.
Lener T, Gimona M, Aigner L, Borger V, Buzas E, Camussi G, et al. Applying extracellular vesicles based therapeutics in clinical trials—an ISEV position paper. J Extracell Vesicles. 2015;4:30087.
Acknowledgements
Disclosures: Authors disclose no conflicts.
Grant Support: Spanish Ministry of Economy and Competitiveness [J.M. Banales (FIS PI12/00380, FIS PI15/01132, FIS PI18/01075 and Miguel Servet Programme CON14/00129), M.J. Perugorria (FIS PI14/00399, PI17/00022 and “Ramon y Cajal” Programme RYC-2015-17755), cofinanced by “Fondo Europeo de Desarrollo Regional” (FEDER)]; ISCIII CIBERehd: J.M. Banales and M.J. Perugorria; “Diputación Foral de Gipuzkoa” (J.M. Banales: DFG15/010, DFG16/004), BIOEF (Basque Foundation for Innovation and Health Research: EiTB Maratoia BIO15/CA/016/BD to J.M. Banales), Department of Health of the Basque Country (M.J. Perugorria: 2015111100; J.M. Banales: 2017111010), “Fundación Científica de la Asociación Española Contra el Cancer” (AECC Scientific Foundation, to J.M. Banales) and PTDC/MED-PAT/31882/2017 (Portuguese Foundation of Science, Portugal, to R.E. Castro) and Centro Internacional sobre el Envejecimiento (OLD-HEPAMARKER, 0348_CIE_6_E to R.E. Castro). A. Lapitz was funded by the Basque Government (PRE_2017_1_0345). A.L. Simão was funded by the Portuguese Foundation of Science (FCT; SFRH/BD/104160/2014). MA was partially supported by the Chilean government through the Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT 1119145 to M.A.) and the Comisión Nacional de Investigación Científica y Tecnológica (grant CONICYT PIA/Basal PFB12, Basal Centre for Excellence in Science and Technology to M.A.).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Rodrigues, P.M. et al. (2020). Extracellular Vesicles in Non-alcoholic Fatty Liver Disease: Key Players in Disease Pathogenesis and Promising Biomarker Tools. In: Romero-Gomez, M. (eds) NAFLD and NASH. Springer, Cham. https://doi.org/10.1007/978-3-030-37173-9_9
Download citation
DOI: https://doi.org/10.1007/978-3-030-37173-9_9
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-37172-2
Online ISBN: 978-3-030-37173-9
eBook Packages: MedicineMedicine (R0)