
Abstract
Wheat accounts for about 20% to over 50% of the total calorie intake of food in regions where it is grown. However, there is a clear perception that disorders related to the consumption wheat are increasing, particularly in Western Europe, North America, and Australia. We consider here the evidence for this perception and discuss strategies and therapies that may be used to reduce the adverse impacts of wheat on the health of susceptible individuals. First, we will introduce the major groups of wheat grain proteins, focusing on those associated with adverse reactions, and discuss in detail the three major adverse reactions triggered by wheat consumption, namely gluten intolerance (celiac disease), wheat allergies, and non-celiac gluten/wheat sensitivity. Finally, will discuss other issues associated with the consumption of gluten-free foods focusing on gluten contamination of products purported to be gluten-free, gluten threshold or tolerance among celiac patients, and food labeling.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others



Keywords
Wheat accounts for about 20% to over 50% of the total calorie intake of food in regions where it is grown. However, there is a clear perception that disorders related to the consumption wheat are increasing, particularly in Western Europe, North America, and Australia. We consider here the evidence for this perception and discuss strategies and therapies that may be used to reduce the adverse impacts of wheat on the health of susceptible individuals. First, we will introduce the major groups of wheat grain proteins, focusing on those associated with adverse reactions, and discuss in detail the three major adverse reactions triggered by wheat consumption, namely celiac disease, wheat allergy, and non-celiac gluten/wheat sensitivity. Finally, will discuss other issues associated with the consumption of gluten-free foods focusing on gluten contamination of products purported to be gluten-free, gluten threshold or tolerance among celiac patients, and food labeling.
1 Introduction
Wheat provides one-quarter of the global annual demand for dietary plant proteins and has been consumed, if not cultivated, for at least 40,000 years, with breadmaking dating back at least 14,000 years (Arranz-Otaegui et al. 2018; Henry et al. 2011, 2014). However, wheat may also have adverse health effects on susceptible individuals, some of which are well documented and understood, such as celiac disease, while others have only recently been described and are still poorly understood. There is a perception that the range and prevalence of these conditions have increased in recent decades. We will, therefore, review the types, mechanisms, and prevalence of adverse responses to wheat consumption and discuss strategies to alleviate the effects in susceptible individuals. Most of these conditions are thought to relate to the protein components of the grain, particularly gluten but also other grain proteins. We will, therefore, begin by summarizing the properties of wheat grain proteins.
2 Wheat Grain Proteins
Wheat grain proteins were among the earliest proteins to be studied, so a vast literature has been amassed dating back almost 300 years. However, until recently, the focus was almost exclusively on the gluten proteins because of their unique role in determining the food processing quality of grain. However, this interest has widened in recent years to include other proteins which may affect grain processing or induce adverse reactions to wheat consumption. Although the total number of proteins present in the mature grain may never be determined, using proteomics almost 500 proteins have been separated from flour (Dupont et al. 2011) and over 1100 from mature whole grain (Skylas et al. 2000). Most of these proteins fall into just a few groups. We will, therefore, focus on these major components, particularly those that have effects, whether positive or negative, on human health. It is only possible to provide a broad overview here, and more detailed accounts are available in other review articles (for example, Shewry et al. 2009; Wrigley et al. 2006).
2.1 Gluten Proteins
Gluten can be defined as the cohesive viscoelastic mass that remains after dough made from wheat flour is washed to remove the starch and other particulate and soluble materials. Gluten consists mainly of proteins, with most of the remainder being lipids and residual starch. Isolated gluten contains small amounts of other proteins, either because they associate with the gluten proteins or are entrapped in the gluten network formed in dough. Based on the traditional fractionation method of Osborne (1924), gluten proteins are classified into two groups, the gliadins (classified as prolamins) and glutenins (classified as glutelins). This classification is based on their sequential extraction in a series of solvents, with prolamins being soluble in aqueous ethanol and glutelins in dilute acid or alkali. However, the amounts and proportions of these fractions vary widely with the precise solvents and extraction conditions, and it is now known that they comprise related proteins which differ in being present as monomers (gliadins) or as components of polymeric complexes which may exceed 1 × 106 Da in molecular mass (glutenins). Hence, it is more usual to consider them as a single protein family. The proportions of gliadins and glutenins in grain vary depending on the genotype and the environment, but the ratio of 30% gliadins to 49% glutenins is typical of 17 European cultivars, as reported by Seilmeier et al. (1991). However, high gluten protein content is generally associated with a higher proportion of monomeric gliadins (Godfrey et al. 2010). The gliadins and glutenins form the major storage proteins of wheat and are deposited in the starchy endosperm cells of the developing grain.
2.1.1 Gluten Protein Types
Gluten proteins are conventionally separated by electrophoresis, as shown in Fig. 1. Comparisons of amino acid sequences show that all of these components are related, but they can be classified into three groups and subgroups thereof. The gliadins are most effectively separated by electrophoresis at low pH (Fig. 1) giving four groups of bands which are called, in order of increasing mobility, α-gliadins, β-gliadins (now considered as one group with α-gliadins), γ-gliadins, and ω-gliadins. Reduction of the disulfide bonds which stabilize glutenin polymers allows the subunits to be separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) into two groups of bands, called high molecular weight (HMW) and low molecular weight (LMW) subunits. Comparison of sequences shows that the ω-gliadins and HMW glutenin subunits form distinct groups, which have been called sulfur-poor (to reflect their lack of cysteine residues) and HMW prolamins, respectively (Shewry et al. 1986). The α-gliadins, γ-gliadins and the major group of LMW subunits (called B-type) are more closely related to each other than to the ω-gliadins and HMW subunits and have been defined as sulfur-rich prolamins. In addition, two minor groups of LMW subunits, termed C-type and D-type, are most closely related to α- and γ-gliadins and to ω-gliadins, respectively, differing in the presence of unpaired cysteine residues which allow them to be incorporated into gluten in polymers by interchain disulfide bonds.

The classification and nomenclature of wheat gluten proteins separated by SDS-PAGE (right) and electrophoresis at low pH (left). The D-type of LMW subunits are only minor components and are not clearly resolved in the separation shown. Taken from Shewry et al. (1999) with permission
Although no gluten proteins have been sequenced completely at the protein level, a vast number of sequences derived from the sequences of cDNA and genomic DNA are available in databases. For example, using gluten protein query sequences, Bromilow et al. (2017) retrieved over 24,000 sequences from the UniProt database in May 2015. After removal of redundant, partial, and mis-assigned sequences, a curated database of 630 sequences was assembled. These sequences contain a vast amount of information, and it is only possible to provide a brief summary here, focusing on features which may relate to the role of gluten proteins in health and disease.
Firstly, the comparisons validate the broad classification into the S-rich, S-poor and HMW prolamin groups and into major subgroups of gliadins and LMW glutenin subunits. Secondly, all of the proteins have clearly defined domain structures with a central domain comprising repeated sequences based on one of two short repeated peptide motifs flanked by non-repetitive domains. However, the relative sizes of these domains vary widely between, and to a lesser extent within, the protein types. Notably, whereas the non-repetitive domains are reduced to a few amino acids in the ω-gliadins, the S-rich prolamins contain extensive non-repetitive N-terminal domains.
The sequences of the non-repetitive domains of the three protein groups are clearly related, and it has been suggested that they have been derived from a common ancestral protein by insertion of blocks of repeated peptide sequences. The non-repetitive domains contain most, and often all, of the cysteine residues which form intra-chain disulfide bonds in the monomeric α-gliadins and γ-gliadins, and both interchain and intrachain disulfide bonds in the polymeric HMW and B-type LMW subunits. Most C-type and D-type LMW subunits have single cysteine residues which form inter-chain bonds.
In terms of health impacts, the repeated sequences are of prime interest, as they contain the sequences known to trigger celiac disease (which are often called celiac epitopes). The short peptide motifs range in length from three to about ten amino acid residues and may be repeated in tandem or interspersed with a second motif. The repetitive sequences of some prolamins are highly conserved with clear consensus motifs, for example, PQQPFPQ, PQQPFPQQ, and PQGPQQ+GYYPTSPQQ+GQQ (letters are single letter abbreviations for amino acids: G, glycine; F, phenylalanine; P, proline; Q, glutamine; S, serine; Y, tyrosine; T, threonine). However, in other cases such as the LMW subunits, the sequences are degenerate and consensus motifs are difficult to define. Alignments of the repetitive domains of a range of gluten proteins are presented by Shewry et al. (2009).
The presence of repetitive sequence motifs determines the solubility of the prolamin monomers and skews their amino acid compositions. Unusually glutamine accounts for between 30% and 50% of the total amino acid residues. The numbers of glutamine residues and their contexts within the sequence also define the ability of gluten proteins to elicit a response in celiac disease.
2.2 Other Storage Proteins
Prolamins are only found in the grains of cereals and other grasses, where they usually form the major storage protein fraction. Other types of storage protein may also be present, such as globulins related to the 7S and 11S globulins which are the dominant storage proteins in the seeds of most other plants. In wheat, 7S globulins are the major storage protein in the aleurone layer and embryo, and small amounts of 11S globulins (called triticins) are present in the starchy endosperm. A third globulin type, called α-globulin, is low molecular weight member of the prolamin superfamily (discussed below). These proteins are discussed in detail in Shewry et al. (2009). Dupont et al. (2011) reported that total globulins accounted for about 0.4% of total proteins in white flour, although 7S globulins are depleted from this fraction.
2.3 The Prolamin Superfamily
Wheat gluten proteins are defined as prolamins based on the characteristic solubility and amino acid composition conferred by their repetitive domains. However, wheat grain also contains several types of proteins whose sequences are related to the non-repetitive domains of prolamins, particularly in the conservation of cysteine residues. They are therefore classified in the prolamin superfamily and together account for most components with molecular masses between about 15,000 and 25,000 Da in SDS-PAGE, although few individual components have been quantified. They include two types of proteins which are considered to be modified forms of prolamins with storage functions. These have been called avenin-like proteins or farinins and LMW gliadins or purinins (Kasarda et al. 2013) and together account for less than 2% of total flour proteins (Dupont et al. 2011). However, three groups of proteins, puroindolines, amylase/trypsin inhibitors (ATIs), and lipid transfer proteins (LTPs) have been shown to have significant impacts on grain functionality and health.
2.4 Puroindolines
The wheat grain contains two puroindoline (Pin) proteins, each with a mass of about 15,000 Da and comprising about 120 amino acid residues including six conserved cysteines. However, they differ from the rest of the prolamin superfamily in having a short domain which contains three or five tryptophan residues, which is thought to form a loop region in the folded protein. Pins are encoded by genes at the Hardness (Ha) locus, and allelic variation in their expression and/or sequences accounts for 60–80% of the variation in endosperm texture (hardness) in bread wheat (Turner et al. 2004). Their biological function is not known.
There is a massive literature on Pins, particularly on the relationship between protein sequences and variation in grain texture, which has been reviewed by Shewry et al. (2009) and Morris (2002). However, Pins have no reported impact on human health and will not be discussed further in this context.
2.5 Lipid Transfer Proteins
Unlike most members of the prolamin superfamily, lipid transfer proteins (LTPs) are not restricted to seeds and have been characterized from a range of plant species and tissues. Two forms occur, LTP1 with a molecular mass of about 9,000 Da and LTP2 with a molecular mass of 7000 Da, both having eight cysteine residues which form four intra-chain disulfide bonds. They were initially defined by their ability to transfer phospholipids from liposomes to mitochondria in vitro, but this function lacks specificity, and its relevance to their role in vivo is uncertain. LTPs are now more widely considered to be involved in plant defense, as they are concentrated in epidermal tissue and can inhibit fungal growth in vitro. LTPs are also a major group of food allergens, as discussed by Marion et al. (2004) and Jenkins et al. (2005). Wheat LTP has also been reviewed by Shewry et al. (2009).
2.6 Amylase/Trypsin Inhibitors (ATIs)
Water-soluble inhibitors of α-amylase and/or trypsin (ATIs) have been known for over 70 years (Kneen and Sandstedt 1946) and are estimated to account for up to 80% of the total albumins and 1% of the total proteins in wheat flour (Cordain 1999). They are readily observed as a group of bands migrating faster than the gluten proteins in SDS-PAGE separations. Early work led to some confusion due to the use of different numbering systems, based either on their electrophoretic mobility at alkaline pH (0.19, 0.28, 0.53) or their selective extraction in chloroform-methanol mixtures (CM1, CM2, etc.). However, with further characterization and the availability of full sequences, eleven subunits which form monomeric, dimeric and tetrameric structures were identified, and their genes to be assigned to chromosomes (as summarized in Table 1). ATIs are described in detail by Carbonero and Garcia-Olmedo (1999), and reviewed by Shewry et al. (2009).
Although all ATIs are inhibitory to human α-amylases, their relative activity varies, being very low for WTAI, higher for WMAI, and highest for WDAI (Salcedo et al. 2004). Similarly, although all three forms are also active against α-amylases from insects, WMAI and WTAI inhibit α-amylase from the coleopteran pest Tenebrio molitor more strongly than WDAI, which is more active against α-amylase from larvae of the lepidopteran Ephestia kuehniella (Salcedo et al. 2004). The CMx proteins are encoded at a single locus on group 4 chromosomes and are proposed to be trypsin inhibitors based on their similarity with the barley trypsin inhibitor BTI-CMe (Sanchez de la Hoz et al. 1994).
Further information on the diversity and relative abundances of ATIs come from two recent proteomic studies (Altenbach et al. 2011; Dupont et al. 2011). Coding sequences for two forms of WMAI, four forms of WDAI, six forms of WTAI and three forms of CMx were identified, and the relative abundances of the inhibitor types (summarized in Table 1) and of the individual subunit forms were estimated (Dupont et al. 2011; Altenbach et al. 2011).
2.7 Other Bioactive Proteins
Although ATIs form the major albumin fraction in wheat grain and flour, a number of other proteins are present in very small amounts that could nevertheless contaminate protein preparations used in dietary interventions and other studies. They include proteins related to the well-characterized chymotrypsin inhibitor and amylase/subtilisin inhibitor of barley, serpin-type protease inhibitors, xylanase inhibitors, ribosome-inactivating protein (tritin) related to the well-characterized and highly bioactive ricin of castor bean, pathogenesis-related proteins (proteins produced in response to infection, predation or damage), thionins and, in the embryo only, wheat germ agglutinin (WGA). It is not possible to describe all of these proteins here, but details can be found in review articles (Wrigley and Bietz 1988; Shewry et al. 2009).
3 Adverse Reactions to Wheat or Wheat Components
The contribution of wheat to health and disease in developed countries has been widely debated, including the suggestion that wheat consumption results in obesity and associated diseases. Although these suggestions are widely promoted in the popular press and social media, they are not borne out by detailed scientific studies (Brouns et al. 2013). However, there is clear scientific evidence that three types of the disorder may present in susceptible individuals when exposed to wheat or wheat components, most notably the proteins. These are celiac disease (CD), wheat allergy (WA), and non-celiac wheat sensitivity (NCWS). These disorders are often misclassified and referred to as allergies, which is incorrect. CD, WA, and NCWS differ in the timeframe in which they develop, the mechanisms involved, symptomatology, and options for diagnosis. Many studies have addressed the effects of gluten, and gluten is the assumed cause of a wide range of adverse reactions and symptoms that disappear after the elimination of gluten from the diet. However, it is important to be aware that avoidance of wheat gluten also results in the avoidance of other proteins and components of wheat. Even isolated wheat gluten, which is often used as an ingredient in the food industry, is known to contain many components. A short overview of the most relevant aspects of each of these disorders is given below.
3.1 Celiac Disease
CD is defined by Ludvigsson et al. (2013) as ‘a chronic small intestinal immune-mediated enteropathy precipitated by exposure to dietary gluten in genetically predisposed individuals.’ CD is triggered by the ingestion of gluten, and over time results in a variable degree of intestinal damage. In most patients with CD, the gut pathology will reverse on the transition to a gluten-free (GF) diet. CD may present in various degrees of ‘visibility,’ for which different terms are used, described in detail by Ludvigsson et al. (2013).
3.1.1 CD Etiology and Prevalence
CD results from a combination of a specific genetic predisposition and environmental factors. Approximately, 25–40% of the population has a genetic predisposition as shown by the presence of HLA-DQ2 or HLA-DQ8 receptor genes. However, it is estimated that only 4% of these DQ2/8 positive individuals actually develop CD, resulting in a mean CD prevalence of around 1% of the global population but there is a large variation (range 0.3–5.6%) between countries (Stein and Schuppan 2014; Catassi et al. 2015; Lionetti et al. 2015). The disease appears to be more common in women than in men, with a ratio of 2-3 to 1. Factors other than genetic predisposition may trigger the disease such as age (Pinto-Sánchez et al. 2016), the dose of initial gluten exposure (Koning 2012), disease or drug/alcohol-related changes in intestinal permeability, or exposure to viral infections (Lebwohl et al. 2018). Rubio-Tapia et al. (2012) documented an increase in the prevalence of CD over recent decades, which is often suggested to be the result of an increase in the consumption of wheat gluten.
Other factors contributing to the recent increase in the prevalence of gluten-induced disorders are major changes in overall lifestyle, diet, hygiene, and gut microbiota, the use of antibiotics and vaccines, and improvements in diagnosis and medical registrations leading to the recognition of formerly undiagnosed, misdiagnosed and unreported cases (Olivares et al. 2018). A well-known case of misdiagnosis comes from the Punjab province of India, where gluten sensitivity and intolerance were misdiagnosed for a very long time as ‘summer diarrhea’, which is quite prevalent in the region but is only reported in summer when wheat flour is used for making ‘chapatti’ flatbreads, instead of maize flour predominantly used in winter (Cataldo and Montalto 2007).
The initial development of CD may be related to the food that a child receives early in life. The influence of breastfeeding and the time of weaning on the disease are not clear (Ludvigsson and Fasano 2012). However, it has been suggested that intake of small quantities of gluten should start gradually before the age of 6 months, preferably simultaneously with breastfeeding (Ivarsson et al. 2013). The reason for this is that the immune-modulatory properties of breastfeeding and the intestinal flora should contribute to the prevention of auto-immune diseases (Agostini et al. 2008). A number of studies indicated a role of gut microbial diversity and related gut-associated immune competence in the etiology of CD (Nadal et al. 2007; Nistal et al. 2012, 2016; Olivares et al. 2018). Whether this link is truly causal or a consequence of altered dietary patterns in individuals that suffer from CD needs further study. It is often suggested that vital wheat gluten, increasingly used as a functional ingredient in food production, also plays a role. However, consumption data over the years are difficult to obtain. Kasarda (2013) calculated that the intake of vital gluten has tripled since 1977, from 136 to 408 g per year, or 0.37–1.12 g per day per capita of the population. However, the impact of this increase is a matter of debate given the many times higher intake of gluten from wheat flour in bread, 5–5.5 kg per year, or 13.7–15.1 g per day. Furthermore, the protein content of wheat has tended to decline as yield has increased (Shewry et al. 2016), and much of the vital gluten is used in breadmaking to compensate for this decrease. Ever since humans began to consume wheat, the ingestion of gluten contained in bread has been many times higher than the amount of gluten that causes CD. Barley and rye contain proteins related to wheat gluten, and hence, consumption of their grains contributes to daily ‘gluten’ exposure. The high contents of proline and glutamine in gluten proteins may contribute to them resisting digestion in the human gastrointestinal tract. In individuals with altered gut permeability, these undigested gluten fragments (peptides) may enter the lamina propria of the small intestine via transcellular or paracellular routes, leading to a cascade of reactions causing inflammation and immune responses. These mechanisms are described in detail by Fasano (2012) and Lebwohl et al. (2018).
When undigested gluten fragments enter the intestinal wall, the enzyme tissue transglutaminase (tTG or TG2) converts the glutamine in the peptides into glutamic acid, which then binds more strongly with the HLA-DQ2/8 receptor and with T-cells of the immune system, activating an inflammatory response that finally leads to damage to the intestinal villi. These inflammatory reactions allow substances to pass through the intestinal wall and cause problems elsewhere in the body such as dermatitis herpetiformis, which responds to a GF diet (Ludvigsson et al. 2013; Ciacci et al. 2015). In addition, neurological manifestations and ataxia have been described (Ludvigsson et al. 2013). Gluten ataxia is defined as ‘idiopathic sporadic ataxia and positive serum antigliadin antibodies even in the absence of duodenal enteropathy’. Wheat gluten consumption has also been suggested to be involved in the etiology of some mental/psychological disorders (Hadjivassiliou et al. 2010).
In the long term, CD-related inflammation and immune reactions will impact on the intestinal wall surface leading to the disappearance of the surface area increasing villi flattening of the gut mucosa (Fig. 2) and resulting in impairment of nutrient uptake, weight loss and, in children, growth retardation (Lebwohl et al. 2018). However, it should be noted that this is not the only manifestation of CD. Some individuals may have the HLA-DQ2 or DQ8 genes and CD-related antibodies in their blood without verifiable damage to the intestinal mucosa but may have one or more general complaints such as chronic fatigue, poor sleep or headaches. Often the symptoms of these people are not recognized as possibly representing CD and, as a result, such individuals are not tested or diagnosed for CD: a phenomenon which is called the subclinical CD, or potential CD. It is estimated that only 1 out of 8 people with CD (10,000–27,000 people) have been diagnosed based on symptomatology, which means that many individuals remain undiagnosed because of unclear or unrecognized symptoms, referred to as ‘the celiac iceberg’ (Fig. 3).

A schematic illustration of progressive tissue changes in the small intestine leading to flattening of the villi and more intraepithelial lymphocytes (black dots) in the gut mucosa lining, (source Cukrowska et al. 2017)

The celiac iceberg. (source Fasano 2007)
3.1.2 CD Causing Substances
Wheat gluten proteins all have high contents of proline and glutamine, which are concentrated in their repetitive domains. As a consequence, these domains may only be partially digestible by the enzymes present in the human intestine, producing indigestible gluten fragments which contain short sequences of amino acids known as epitopes. The identification of the fragments which cause immunological reactions have led to rapid progress in CD research (Pastorello et al. 2007; Tatham and Shewry 2008; Mamone et al. 2011; Ludvigsson et al. 2013). It includes the evaluation of epitopes with the greatest ability to stimulate T-cells (immunodominance) (Anderson et al. 2000, 2005; Shan et al. 2002, 2005; Tye-Din et al. 2010) and the agreement of an internationally accepted list of all epitopes known to play a role in CD (Sollid et al. 2012).
It has also been shown that there are large differences between individuals in the physical responses to single or combinations of gluten fragments. It is also clear that there are broad differences in the celiac activity of different groups of gluten proteins. In particular, the α-gliadins, and to a lesser extent the γ-gliadins, yield large numbers of immunogenic peptides in the form of indigestible protein fragments. People with CD also react to related proteins from rye (secalins) and barley (hordeins), although neither species contains proteins closely related to the α-gliadins. A minority of CD patients react to glutenins.
The number, type, and distribution of toxic epitopes within a wheat genotype are clearly of interest (Shewry and Tatham 2016). Salentijn and coworkers noted the presence of significantly fewer toxic epitopes in several tetraploid wheat species (Salentijn et al. 2009, 2013), while several other reports indicate that ‘modern’ hexaploid wheat types may induce more immune and inflammatory reactivity than ‘ancient’ tetraploid and diploid species, and hence result in more gastrointestinal problems in wheat-sensitive individuals (Molberg et al. 2005; Spaenij-Dekking et al. 2005; Pizzuti et al. 2006; Vincentini et al. 2007, 2009; Sofi et al. 2010, 2014; Carnevali et al. 2014; Gelinas and McKinnon 2016). However, other studies have shown that all types of wheat, whether ‘ancient’ species or modern cultivars, induce some degree of immune reactivity and thus should be avoided by CD patients. For example, when the contents of immunogenic α-gliadin in two old tetraploid wheats (Graziella Ra and Kamut) and four modern tetraploid durum wheat varieties (Capelli, Flaminio, Grazia, and Svevo) were compared (Gregorini et al. 2009; Colomba and Gregorini 2012), the older varieties had significantly more, and so are considered unsuitable for individuals with CD. More recent studies have confirmed these findings (Šuligoj et al. 2013; Escarnot et al. 2018).
By contrast, van den Broeck et al. (2010b) used immunoblotting with antibodies to the Glia-α9 and Glia-α20 epitopes to classify modern bread wheat varieties (1986–1998) and older types (1863–1982) as having low, medium or high reactivity. The Glia-α9 antibody reacts with one of the most harmful epitopes, present in a 33-mer peptide from α-gliadin, while the epitope recognized by Glia-α20 is harmful to a more restricted group of people. Based on this, the authors suggested that the relative reactions of wheat lines with these two antibodies could be used to classify them as more or less toxic for CD. The results showed that only 1 of the 36 modern wheat varieties studied had low levels of the 33-mer peptide, compared with 15 of the 50 older types. The number of lines with high reactivity to Glia-α20 was similar in both groups. However, no systematic differences were found between the modern bread wheat (T. aestivum subsp. aestivum) and older types of hexaploid wheat (T. aestivum subsp. spelta and compactum). The authors suggested that scientific breeding has contributed to an increased content of highly CD reactive epitopes in modern wheat varieties and accordingly to increased CD prevalence (van den Broeck et al. 2010b). However, more recent studies do not support this suggestion. Kasarda (2013) reviewed the arguments as to whether wheat breeding programs aimed at increasing gluten content may have contributed to the increase in CD in the USA during the latter half of the 20th century, but the available data do not demonstrate an increase in gluten due to breeding.
Ribeiro et al. (2016) compared modern varieties of bread wheat (Triticum aestivum subsp. aestivum), spelt (T. aestivum subsp. spelta) and durum wheat (Triticum turgidum subsp. durum), including modern cultivars and old landraces from different countries. The lines were grown in the same year and location and under identical environmental growing conditions and the amounts of potential CD immune-stimulatory epitopes measured with the R5 monoclonal antibody assay that recognizes several toxic peptides (QQPFP, QQQFP, LQPFP, QLPFP, QLPYP) that occur repeatedly in gluten proteins. Different gliadin types were quantified by reversed-phase high-performance liquid chromatography (RP-HPLC) and acid-polyacrylamide gel electrophoresis (A-PAGE) in order to correlate toxic epitopes with gliadin content. In line with earlier observations (Spaenij-Dekking et al. 2005; van den Broeck et al. 2010a, b), this study confirmed that there is significant heterogeneity in the level of the T-cell–stimulatory epitopes in wheat varieties. For example, the modern T. aestivum variety ‘Pernel’ presented more than 11-fold fewer toxic epitopes than the variety ‘Alejo’. In addition, high heterogeneity in the content of toxic epitopes in tetraploid lines was observed with values ranging from 26.40 ± 1.65 g per kg in ‘Basto Duro’ to 223.14 ± 30.04 g per kg in ‘Valenciano’. Spelt varieties proved to have more toxic epitopes compared to other types of hexaploid wheats, with a mean of approximately 311.15 g per kg. The authors concluded that intensive wheat breeding has not resulted in increases in toxic epitopes in modern wheat varieties, so cannot be the reason for increases in CD incidence.
It can, therefore, be concluded that there is no evidence to support the conjecture, particularly in social media, that ancient tetraploid grains and spelt are more tolerated by individuals suffering from CD and WA. In fact, peptides from both spelt and durum result in immune responses and should certainly not be recommended as alternatives to conventional modern wheat for celiac patients (Ribeiro et al. 2016; Vincentini et al. 2007, 2009; Gregorini et al. 2009; Šuligoj et al. 2013; Escarnot et al. 2018). Indeed, a recent comprehensive review of peptides from gluten digestion comparing old and modern wheat varieties concluded that the old varieties may actually contain more immunogenic and toxic sequences than modern varieties (Prandi et al. 2017). The relative proportions of CD-immunogenic and CD-toxic sequences in gluten proteins also vary depending on environmental factors such as climate, soil, fertilization and agricultural practices, and their interactions with the genotype (Ashraf 2014; Hajas et al. 2018).
Gianfrani et al. (2015) provided evidence that extensive in vitro gastrointestinal hydrolysis drastically reduced the immune stimulatory properties of Triticum monococcum gliadin. The MS-based analysis showed that several T. monococcum peptides, including known T-cell epitopes, were degraded during the gastrointestinal treatment, whereas many of T. aestivum gliadins survived the gastrointestinal digestion. However, these findings need confirmation in vivo, but a crucial observation is that not all peptides were degraded; thus, T. monococcum is still not safe for CD patients. More recently, Perez-Gregorio et al. (2018) showed that the composition of gluten hydrolysates depended on the digestion time and structural characteristics of the protein. On the other hand, the main T-cell stimulating epitopes formed during hydrolysis depend on the identity of the precursor protein. Glutenin oligopeptides were degraded faster than gliadins, particularly α-gliadin oligopeptides.
In addition to gluten, it has recently been recognized that non-gluten proteins, notably serpins, purinins, globulins, ATIs, and farinins, may also elicit antibody responses (Huebener et al. 2015). The growth conditions of the plant (shade and plant height) and grain storage conditions appeared to influence the ATI content of three different types of durum wheat grown at three locations in Italy (Prandi et al. 2013), with the effect of the environment being greater than that of genotype.
Recent research has indicated that ATIs may play a role in the development of both CD and non-celiac-related wheat hypersensitivity via the production of inflammatory factors (cytokines) in the intestine (Junker et al. 2012; Zevallos et al. 2017). These studies were carried out in animal models (mice) and in vitro assays on human cell lines, using isolated protein fractions enriched in ATIs. These studies indicate a clear need to carry out human trials with oral exposure to ATIs that have also been exposed to heat treatment and simulated gastrointestinal digestion. Zevallos et al. (2017) suggest that both the types and amounts of isoforms affect the degree of bioreactivity of ATIs. ATIs are heat resistant, and active forms may be present in wheat cooked for 5 min at 100°C and may cause an allergic reaction on ingestion (Pastorello et al. 2007). Rye and barley also contain a range of related ATI isoforms (Carbonero and García-Olmedo 1999).
3.1.3 CD Diagnosis and Solutions
CD can be diagnosed based on the pattern of symptoms, blood serology, genetic predisposition for CD and, finally, by the histological screening of small intestinal tissue obtained by biopsy. General guidelines for CD diagnosis are given by Bai and Ciacci (2017) and Lebwohl et al. (2018). A comparative overview of the characteristics of CD, WA, and NCWS is given in Table 2.
A completely GF diet appears to be the only remedy for people with CD. Even traces of wheat should be avoided in foods or food ingredients produced in a ‘wheat presence environment’ or other ‘gluten’-containing grains (barley, rye) present in GF food through cross-contamination that might occur during cultivation, harvest, transport, production (see below for further details). By completely avoiding gluten, the intestinal wall can recover, and symptoms disappear.
3.2 Wheat Allergy (WA)
An allergen is defined as a substance that causes an immediate immune reaction upon exposure by ingestion, inhalation, or skin contact. Depending on the route of allergen exposure and the underlying immunological mechanisms, WA may be classified as (1) an immediate food allergy, (2) wheat-dependent exercise-induced anaphylaxis, (3) a respiratory allergy, or (4) contact urticaria. IgE antibodies play a central role in the pathogenesis of these diseases.
3.2.1 WA Etiology and Prevalence
In WA, the body reacts to the protein as if it was a pathogen that needs to be dealt with. This misrecognition leads to inflammatory reactions and symptoms such as swollen membranes of the mouth and throat, difficulty in swallowing, shortness of breath, diarrhea, vomiting, abdominal pain, asthmatic reactions, and rashes. In some cases, a whole-body reaction may cause a sudden severe drop in blood pressure leading to anaphylactic shock or even death (Hadjivassiliou et al. 2015). The most widely occurring allergy to wheat is bakers’ asthma, a respiratory allergy resulting from the inhalation of wheat flour or starch.
For a schematic representation of allergy mechanisms see: https://upload.wikimedia.org/wikipedia/commons/e/e7/The_Allergy_Pathway.jpg. A number of detailed reviews of WA have been published (Tatham and Shewry 2008; Inomata 2009; Sapone et al. 2012; Gilissen et al. 2014; Cianferoni 2016; Pasha et al. 2016).
WA prevalence amongst children varies from <0.1% to 1%, depending on age and country (Hischenhuber et al. 2006; Kotaniemi-Syrjänen et al. 2010; Sapone et al. 2012). A large meta-analysis has shown that the general prevalence is at most approximately 0.2% (Zuidmeer et al. 2008). About one-half of children ‘outgrow their food allergy,’ and there are reports that more than 80% of children with WA have outgrown it by their 8th year, and 96% before their 16th year (Kotaniemi-Syrjänen et al. 2010). Accordingly, the number of adults with WA is much lower than the number of people with CD. It has also been shown that the prevalence of food allergies in women is generally higher than in men (Afify and Pali-Schöll 2017). Interestingly, the prevalence of NCWS is also higher in women than in men.
3.2.2 WA Causing Substances
Wheat proteins that cause allergic reactions are very diverse and differ between individuals (Mamone et al. 2011; Cianferoni 2016). The two main wheat protein groups causing food allergy are ω5-gliadins and ATIs, and equivalent proteins from other grains. To a lesser extent, reactions to LMW-glutenin subunits, WGA and LTPs have been reported (Gilissen et al. 2014; Baccioglu et al. 2017; Brans et al. 2012; Scherf et al. 2016). Children with wheat allergies react most commonly to α/β- and γ-gliadins (Pastorello et al. 2007; Tatham and Shewry 2008). One-fifth of people with WA are also allergic to barley and rye (Sicherer 2001). It has long been known that ATIs play a role in bakers’ asthma (flour dust allergy) and food allergy to wheat (Pastorello et al. 2007; Tatham and Shewry 2008). Recently, Zevallos et al. (2018) and Bellinghausen et al. (2018) showed that ATIs exacerbate allergic reactions in mice. Gélinas and Gagnon (2018) studied the effects of heat treatment on α-amylase inhibition by ATIs and noted that well-heated cereal foods lose the α-amylase activity, compared to those that are only mildly heated. In this respect, it should be noted that enzyme activity may be lost due to heat-induced changes in protein structure but that the protein itself is not degraded. In other words, a change of α-amylase activity does not necessarily mean that there is also a loss of inflammatory and/or immune reactivity in susceptible individuals.
3.2.3 WA Diagnosis and Solutions
WA can be tentatively diagnosed by the combination of a blood test and a skin test. The blood test will demonstrate the presence of specific immunoglobulin E (IgE) antibodies to allergens (in this case wheat proteins) present in the blood, while the skin test will show how the skin reacts via the IgE antibodies after exposure to a very small quantity of subcutaneously injected wheat protein. However, the presence of IgE antibodies against wheat in the blood certainly does not always mean that there is an active (food) allergy nor does the skin test yield a conclusive diagnosis (Sapone et al. 2012). A conclusive diagnosis is only obtained by a food challenge test, executed in a double-blind, placebo-controlled setup. Such tests have shown that WA is a relatively uncommon event, but that many wheat proteins are immunogenic (stimulating the production of IgEs) but have not been found to lead to clinical symptoms (Zuidmeer et al. 2008). It is, therefore, necessary for people with WA to completely avoid products containing wheat (and possibly other grains to which they react). Excellent reviews of WA are available (Battais et al. 2008; Gilissen et al. 2014; Cianferoni 2016).
3.3 Non-celiac Gluten or Wheat Sensitivity
In recent years, the third group of people has been identified who experience symptoms after eating wheat products, but who are not diagnosed to be suffering from either WA or CD. These individuals may have gastrointestinal symptoms which are similar to those for irritable bowel syndrome (IBS) and which improve on the transition to a GF diet. This group is referred to as being non-celiac gluten sensitive (NCGS) or more recently non-celiac wheat sensitive (NCWS). Ludvigsson et al. (2013) have defined NCWS as “one or more of a variety of immunological, morphological, or symptomatic manifestations that are precipitated by the ingestion of gluten in individuals in whom CD has been excluded”.
Despite the word ‘gluten’ in the current definition of NCGS/NCWS, it is far from certain that gluten is the main cause of symptoms in this group of people or indeed whether wheat is the direct cause.
3.3.1 NCWS Etiology and Prevalence
NCWS is a condition in which wheat ingestion leads to morphological or symptomatic manifestations despite the absence of CD. NCWS sufferers may show signs of an activated innate immune response, elevations in anti-tTG antibodies, endomysial antibodies, deamidated gliadin peptide antibodies, and increased mucosal permeability, all of which are characteristic of CD but without the associated enteropathy. Although up to 35% of the population may state that they feel better when avoiding gluten and/or wheat, the percentage of individuals suffering from NCWS is probably much lower, but this depends on the region of observation (DiGiacomo et al. 2013; Rubio-Tapia et al. 2012). At present, it is impossible to reliably estimate the number of people suffering from NCWS/NCGS. Although it is expected that this will be higher than the number of people with CD, precise figures are scarce and range from 0.5% to 10% of the population, but the current most likely estimate is 2–5% (Ludvigsson et al. 2013).
3.3.2 NCWS Causing Substances
Gluten and fermentable oligo-, di-, monosaccharides, and polyols (FODMAPs) are most widely discussed as causative agents of NCWS (Barrett and Gibson 2012; Skodje et al. 2018). Several studies with wheat products, not only with gluten, have been reported in which people were also exposed to other wheat components such as LTPs and ATIs (Biesiekierski et al. 2013; Vazquez–Roque et al. 2013). To date, no studies have been carried out to determine how patients diagnosed with NCWS react to individual components or their combinations. Although Kamut, an old type of tetraploid wheat, caused fewer symptoms in IBS patients compared to modern wheat (Sofi et al. 2014), a recent critical review concluded that further studies using well-defined grain samples grown under the same conditions are required to confirm this relationship (Shewry 2018).
FODMAPs are present in all wheats but also in fruits and vegetables, including legumes (Biesiekierski et al. 2011). Considering the large overlap between symptoms associated with IBS and NCWS, wheat-based products are increasingly being listed as foods to avoid since they contain fermentable carbohydrates. Biesiekierski et al. (2011) observed gluten to cause symptoms in one study, but showed that FODMAPs were the cause in a subsequent study (Biesiekierski et al. 2013). Similar results were reported by Skodje et al. (2018). However, the amount of fructans, the major FODMAPs in wheat, in wheat-based foods is low and far below the levels that would cause abdominal distress in normal healthy individuals (Brouns et al. 2017).
3.3.3 NCWS Diagnosis and Solutions
The diagnosis of NCWS/NCGS is difficult because people report symptoms that may indicate CD as well as symptoms that are known to occur with WA. Often these symptoms are self-diagnosed and also overlap with a cluster of symptoms defined as IBS (see Fig. 4). Caio et al. (2014) and Uhde et al. (2016) showed that people who suffer from NCWS symptoms improve significantly on a wheat/gluten free diet. However, well-defined biomarkers and a clinically validated diagnostic test are still lacking for NCWS/NCGS. Excellent reviews comparing aspects of CD and NCWS/NCGS are available (Volta et al. 2013; Schuppan et al. 2015; Scherf et al. 2016; Leonard et al. 2017; Catassi et al. 2017).

Significant overlap in symptoms induced by wheat grain components, gluten, amylase/trypsin inhibitors (ATIs), fermentable oligo-, di-, monosaccharides and polyols (FODMAPS), and food additives like vital gluten, wheat starch, etc. This figure is modified from Volta et al. (2013)
4 Hidden Gluten or Gluten Contamination
As discussed above, the only known therapy for gluten-induced disorders is adherence to an abstinent diet. With increased awareness and the growing number of self or medically diagnosed cases of celiac disease, gluten intolerance/sensitivity and wheat allergy, the demand for GF products is also increasing. In fact, the U.S. market for GF products is projected to reach USD 7.59 billion by 2020. Since gluten is widely used in food processing, it is a major challenge to identify truly GF products, as gluten can be found in unexpected products and even in products purported to be GF.
Products containing ‘hidden’ gluten include sausages, fish fingers, cheese spreads, soups, sauces, mixed seasonings, mincemeats, and even some medications and food supplements such as vitamin preparations. The major causes of contamination are either the use of common machinery during harvest, transportation, and processing or the use of common storage space. Successive cultivation of gluten-containing and GF crops may also lead to contamination of GF grains. Contamination is unavoidable if the same milling equipment is used for GF and gluten-containing grains. Interestingly there is no legislature in place regarding the maximum permitted levels of foreign grains in inherently GF cereals. However, in practice, 2% of other grains would be considered as the upper limit.
The importance of the issue of gluten contamination for celiac patients and the threat to consumers has been acknowledged for some time, and several studies using different detection procedures, such as PCR, quantitative polymerase chain reaction, enzyme-linked immunosorbent assay (ELISA) and mass spectrometry, have determined the levels of contamination in many commodities (Fritschy et al. 1985; Janssen et al. 1991; Van Eckert et al. 1992; Olexová et al. 2006; Hernando et al. 2008; Thompson et al. 2010; Lee et al. 2014; Farage et al. 2017, and references cited in Miranda and Simón 2017 and Do et al. 2018). For example, gluten contamination as high as 3000 mg per kg was found in buckwheat flour claimed to be GF. A literature review revealed that higher than recommended levels of gluten in purportedly GF commodities is a worldwide problem. Although the studies mainly focused on samples collected from industrialized countries where the regulations are much tighter, it is reasonable to extrapolate the results of these studies to the wider world (for detailed reviews, see Miranda and Simón 2017; Do et al. 2018). In a meta-analysis, Bustamante et al. (2017) charted the rise and fall of gluten content in cereal-based GF food products by using ELISA to analyze 3141 samples from 1998 to 2016. Gluten was detected in a total of 11.8% (371) samples, with breakfast cereals and cereal bars being the most highly contaminated commodities. Additionally, the study suggested that the number of contaminated samples containing low levels of gluten had declined whereas those containing over 100 ppm of gluten had increased during the period 2013–2016 (Bustamante et al. 2017). Although the prevalence of foods with detectable gluten has decreased over time in line with the evolving regulations relating to information on food composition and gluten content claims, the problem persists (Miranda and Simón 2017; Do et al. 2018).
5 Gluten Threshold or Tolerance Level
A GF diet does not necessarily mean ‘zero gluten’ because low levels of gluten may be tolerated by patients. Establishing a patient’s threshold for gluten intake is, therefore pertinent. Although a number of studies have determined the effect of low gluten intake in patients with CD, a more detailed study is required to reach a consensus. Initially, Ciclitira et al. (1984) studied the toxicity of gliadin doses and response time in a single patient and concluded that 10 and 100 mg of gliadins induce no or slight changes in the small intestine morphology, whereas 500 mg and 1 g of gliadin caused moderate to extensive damage. Later, a similar daily gluten intake of 10–100 mg was reported to be safe for consumption by CD patients (Hischenhuber et al. 2006). In a separate study with an observation period of 1–6 weeks, 2.4–4.8 mg gluten dose per day was found to cause no damage to jejunal morphometry (Ciclitira et al. 1985). Similarly, Ejderhamn et al. (1988) demonstrated that a daily intake of 4–14 mg of gliadin did not cause morphological changes in the mucosa of the small intestine of CD patients on the abstinent diet. Two groups in Finland also reported similar observations but with slightly higher daily gluten doses of 20–36 mg (Kaukinen et al. 1999; Peräaho et al. 2003). Later, Catassi et al. (1993) demonstrated that 100 mg of gliadins per day caused deterioration of the small intestine architecture, the effects being more pronounced with the higher dose of 500 mg per day. However, despite these studies, a consensus on the critical limit or threshold for gluten intake has not been reached. This uncertainty is to be attributed to the large variability among celiac patients, as demonstrated by an extensive double-blind placebo-controlled multicenter study of the effects of 10–50 mg of gluten per day on 40 CD patients. The patients were administered daily capsules containing 0 mg, 10 mg or 50 mg of gluten for 90 days and studied for clinical, serological, and histological changes in their small intestine. The study reported wide variation among patients in terms of gluten sensitivity. Some patients showed intestinal symptoms after ingesting only 10 mg of gluten daily while other patients showed no histological symptoms even after three months of a daily challenge with 50 mg gluten. In a similar study, it was reported that a daily intake of 50 mg of gluten for three months is sufficient to cause significant damage to the intestinal morphology of CD patients (Catassi et al. 2007a, b).
Because of the variation in the results of such studies, different countries allow different gluten levels in products for consumption by celiac patients. For example, North European countries (Denmark, Estonia, the Faroe Islands, Finland, Iceland, Latvia, Lithuania, Norway, and Sweden) permit up to 200 ppm of gluten in food for celiac patients. By contrast, North American and South European countries (Italy, Spain, Greece, Portugal, Andorra, Vatican City, San Marino, Gibraltar, Corsica, and Malta) adopted a more conservative limit of 20 ppm gluten in GF products (Do et al. 2018). Based on clinical data, an intermediate limit of 100 ppm was adopted by Finland.
Decisions on gluten thresholds depend mainly on two factors, (i) the minimum toxic dose, and (ii) the amount of GF product(s) consumed. The results of a food challenge study indicated that 200 ppm is not a safe threshold as a gluten intake limit of 50 ppm could be reached with the consumption of 250 g of so-called GF product(s) (Catassi and Fasano 2008). A 100 ppm limit that allows 10 mg gluten in 100 g of food is also impractical as the consumption of GF products in European countries could be as high as 500 g per day (Gibert et al. 2006; Catassi and Fasano 2008). However, a threshold of 20 ppm restricts the intake of gluten from GF food well below 50 mg, thus allowing a safety margin for variation in the gluten sensitivities and dietary habits of different patients. This knowledge has led the Codex Alimentarius Commission to define ‘any product having <20 ppm gluten could be treated as gluten-free because the gluten levels below this limit, in general, do not invoke a disease response.’
6 Food Labeling
As gluten sensitive individuals rely mostly on product labels to make dietary decisions, it is important to label all food ingredients and specifically those in pre-packed foods. The food ingredients that cause intolerance and/or allergenicity are documented in the ‘list of hypersensitivity’ assembled by the Codex Commission and include gluten-containing cereals, Crustacea, eggs, fish, peanuts, milk, tree nuts, and product derivatives of the above-listed items, often dubbed the ‘big eight’. These labeling regulations were established in various countries to help consumers, specifically people with various dietary allergies, to avoid the consumption of packaged food products that contain the major food allergens. In the USA, the Food Allergen Labeling and Consumer Protection Act (FALCPA), which was implemented in 2004, requires the specific declaration of the ‘big eight’ when added as ingredients to labeled food products. Similar legislation also exists in other countries, such as Canada, Australia, New Zealand, Japan, and the European Union (Do et al. 2018). More specifically, whereas gluten is not a permitted food additive in the European Union and the United States, other wheat ingredients such as soluble wheat proteins and starches are permitted and therefore can be used without declaration in food products. Therefore, the Codex Commission concluded that gluten ingredients from all wheat, barley and rye species, including spelt, Khorasan, Kamut and durum wheats (all forms of tetraploid T. turgidum), einkorn (T. monococcum), triticale, tritordeum, and their hybrid varieties are immunogenic, and should be declared. The major concern at present is the misbranding of single/multiple ingredient food products as GF without proper testing, particularly if they are derived from inherently GF grains like millet, flax, buckwheat, and sorghum. Thus, in order to brand them GF, it is important to test to ensure that the gluten level is below the prescribed limit of 20 mg per kg.
7 Potential Therapies
So far, the only reliable therapy available for gluten-related disorders is strict lifelong adherence to a gluten exclusion diet, which is difficult to follow, and is not free of side-effects (see below). The major constraint to the development of new therapies for gluten-induced disorders is the large number of cereal proteins (gluten and non-gluten) that present epitopes (see above), and the fact that different celiac patients differ in the magnitude of reactivity to different gluten epitopes (Koning 2012). Despite extensive efforts to map immunogenic peptides to the prolamins responsible (Juhász et al. 2018; Tye-Din et al. 2010), the repertoire of epitopes is still incomplete (Osorio et al. 2012; Sollid et al. 2012; Comino et al. 2013; Juhász et al. 2015). In general, it would be precautionary to say that gluten peptides more than nine amino acids in length, especially those derived from the repetitive domains or the non-repetitive C-terminal ends of most of the major prolamins, can potentially elicit an immune reaction in susceptible individuals (Osorio et al. 2012; Juhász et al. 2018). Since no extant wheat genotypes either in cultivation or wild are entirely free of gluten, no wheat types can be considered ‘celiac-safe’ (Mitea et al. 2010; Goryunova et al. 2012; Brouns et al. 2013; Shewry 2018).
7.1 Reduced-Gluten Cytogenetic Stocks and Mutations in Triticeae Cereals
An alternative approach to seeking ‘celiac-safe’ types of wheat is to exploit genetic stocks and mutants in which specific groups of gluten proteins are reduced or absent. Early studies compared sets of wheat aneuploid lines in which specific pairs of chromosomes are deleted (and the corresponding chromosomes of other genomes duplicated to improve genetic stability). Thus, some nulli-tetrasomic or deletion lines of Chinese Spring wheat showed low toxicity because certain gliadin loci had been eliminated due to the chromosomal defect (Ciclitira et al. 1980a, b; Frisoni et al. 1995; van den Broeck et al. 2009, 2011). Likewise, wheat mutants lacking α/β-, γ- and/or ω-gliadins and/or showing reduced accumulation or complete elimination of specific gliadins and/or LMW glutenins have been characterized (Moehs et al. 2018; Camerlengo et al. 2017; Waga and Skoczowski 2014; Pogna et al. 1998; Redaelli et al. 1994; Metakovsky et al. 1993). However, these genotypes can be best described as low-toxicity lines, as they still contain gliadins and glutenins. Similar barley mutants with reduced hordein (gluten) content were also identified (Moehs et al. 2018; Cook et al. 2018; Munck 1992; Tallberg 1981a, b, 1982).
Collectively, different celiac patients have different degrees of sensitivity to different prolamin epitopes, so it would be ideal to combine low toxicity alleles of gliadin and glutenin genes in a single genotype (Gil-Humanes et al. 2014). However, given the large number and complexity of the gliadin genes, and their presence and inheritance in tightly linked multigene blocks, the possibility of pyramiding all low toxicity gliadin genes in a single wheat variety is a formidable task through conventional breeding (Koning 2012).
Therefore, it can be concluded that it is not realistic to identify celiac-safe wheat genotypes by screening the extant wheat germplasm or to develop them via conventional breeding. Furthermore, there are logistical issues associated with the release of the reduced-gluten or low-toxicity wheat lines, such as whether they are suitable for individual celiac patients who may differ in their reactions to different gluten proteins, and how derivative food products should be labeled to reflect this.
7.2 Alternative Cereals and Pseudocereals
Another way to deal with this problem is to use inherently GF cereals, such as oats, maize, rice, sorghum, fonio, tef, millet, and Job’s tears or pseudocereals such as amaranth, quinoa, and buckwheat. However, none of these cereals and pseudocereals has similar technological and organoleptic properties to wheat. Furthermore, some individuals are sensitive to oat gluten proteins (avenins), and in rare cases even to maize proteins (Comino et al. 2013; Rosella et al. 2014; Ortiz-Sánchez et al. 2013). So far, the two major cereals that have not raised cause for concern are rice and sorghum (Rosella et al. 2014; Pontieri et al. 2013). However, rice grains have low protein and fiber content and are highly enriched in easily digestible carbohydrates that may contribute to less favorable high glycemic responses. Some rice grain storage proteins (other than prolamins and glutelins) are known to trigger a variety of allergenic reactions in susceptible individuals such as asthma, atopic dermatitis, diarrhea, and anaphylaxis (Matsuda et al. 2006; Nambu 2006; Trcka et al. 2012; Gilissen et al. 2014). Similarly, the use of pseudocereals is also controversial, due to the immunotoxicity of some quinoa (Zevallos et al. 2012, 2014) and buckwheat varieties (Panda et al. 2010; Stember 2006).
Other major issues associated with the use of GF commodities are as follows. (i) Unintended contamination of supposedly GF products that are on the market. Accidental contamination of intrinsically GF products could take place at any level from field to fork due to the ubiquitous nature of gluten or gluten-containing grains. Contamination and misbranding of products make it a real challenge to follow a GF diet. (ii) Strict adherence to a diet totally devoid of gluten-containing grains, or based on foods manufactured for celiac patients, may result in poor gut health of the consumer due to a negative effect on the gut microbiota. It has also been shown that this type of diet increases the risk of colon cancer in consumers, owing to the reduced content of dietary fiber and bioactive compounds (De Palma et al. 2009; Gil-Humanes et al. 2014 and references cited therein). (iii) Adaption to a GF diet may initially improve a patient’s condition, but long-term adherence results in multiple deficiencies and changes in body mass index (BMI), which increase vulnerability to other disorders (Theethira et al. 2014). As most GF foods are made with starches or refined flours with low fiber content, celiac patients consume more energy in the form of fat than in the form of carbohydrate (Martin et al. 2013). Furthermore, it has been shown in a number of studies that when they are diagnosed, individuals with CD have lower BMI than the regional population, but their BMI increases on the transition to the GF diet, especially in those that adhere closely to it (Kabbani et al. 2012; Sonti and Green 2012).
7.3 Genetically-Engineered Reduced-Gluten or ‘Celiac-Safe’ Wheat
Recently, several research groups have taken a genetic engineering approach to develop ‘celiac-safe’ wheat genotypes by either eliminating or detoxifying gluten proteins. In an elimination approach, Becker and co-workers produced a series of transgenic lines where α-gliadin genes were down-regulated using RNA interference (RNAi). In these lines α-gliadins were reduced by over 60% compared to the control cultivar. Compensatory increases in albumins, globulins, other gliadins, and LMW subunits were also reported (Becker et al. 2006, 2012; Becker and Folck 2006; Wieser et al. 2006). Using a similar approach, silencing of the ω5-gliadins was later achieved by Altenbach and co-workers (Altenbach and Allen 2011; Altenbach et al. 2014). More extensive studies were reported by Barro and co-workers in a series of papers which have been reviewed by others (Rosella et al. 2014; Gilissen et al. 2014; Shewry and Tatham 2016; Ribeiro et al. 2018; Jouanin et al. 2018). To summarize, two series of lines were generated downregulating γ-gliadins by between 65% and 97% (Gil-Humanes et al. 2008; Piston et al. 2011) or downregulating all gliadins (α/β, γ and ω) by 60–88% and LMW subunits (Gil-Humanes et al. 2010, 2011). Testing these genotypes with intestinal T-cell clones derived from biopsy samples of CD patients showed almost complete suppression of disease-related T-cell epitopes (Gil-Humanes et al. 2010). More recently, the same researchers used the CRISPR/Cas9 gene editing technology to target conserved regions adjacent to the coding sequence of the 33-mer peptide in the α2-gliadin genes (Sánchez-León et al. 2018). Two single-guide RNAs, dubbed sgAlpha-1 and sgAlpha-2, induced mutations in specific gliadin genes, leading to an 85% reduction in immunoreactivity of mutant lines. This study showed that CRISPR/Cas9 technology can be used to precisely and efficiently reduce the amount of celiac-causing epitopes.
Taking a slightly different approach, the Rustgi and von Wettstein group aimed to silence the wheat DEMETER (DME) genes, which are master regulators of gluten protein accumulation (excluding HMW subunits). To suppress DME expression, two series of transgenic lines were produced, one with DME-specific hairpin RNA and the other with DME-specific artificial micro RNA (amiRNA). The former series of transgenic lines expressed a 938-nucleotide hairpin with a 185-nucleotide stem and a 568-nucleotide loop. Using this construct, a total of 118 candidate transformants were obtained, seven of which exhibited 45–76% reductions in the amount of immunogenic gluten proteins (Wen et al. 2012; Rustgi et al. 2014). In the latter series of transgenic lines, three different amiRNAs were expressed to avoid off-target editing. Two amiRNAs were designed to target the active site sequence and one to target the N-terminal sequence of the wheat DME gene. The selected amiRNA sequences were subsequently assembled on a rice osaMIR528 template using overlapping primers and cloned under the control of the wheat HMWg 1Dy promoter. A total of 215 candidate transformants were obtained, 12 of which showed 54–88% reductions in their respective gluten protein contents (Brew-Appiah 2014; Rustgi et al. 2014). More recently, the same group used site-directed insertional mutagenesis to maximize the level of DME suppression and gluten elimination. The DME-specific transcription activator-like effector (TALE) repressor was introgressed into the wheat Dre2 (Derepressed for ribosomal protein S14 expression) gene using the CRISPR/Cas9 technology. Dre2 is a protein that facilitates deposition of iron-sulfur (Fe-S) clusters in the DME apozyme. Double-stranded breaks introduced into the wheat Dre2 homoeologues are repaired with the help of a donor construct carrying the DME-specific TALE repressor. This approach of simultaneously silencing the DME and Dre2 genes is expected to disrupt DME activity at two-time points during transcription and post-translation respectively, to limit the accumulation of immunogenic gluten proteins (Rustgi, unpublished data).
In a gluten detoxification approach, the Rustgi group expressed ‘glutenases’ in wheat endosperm. This research was inspired by earlier research by the Khosla group, who demonstrated that glutamine-specific proteases from germinating barley grains in combination with bacterial prolyl endopeptidases (PE-Peps) from Flavobacterium meningosepticum, Sphingomonas capsulata or Myxococcus xanthus can completely detoxify the proteolytically resistant proline-glutamine rich peptides (Bethune and Khosla 2012). Later PE-Peps with similar cleavage characteristics (but diverse physicochemical properties) were identified from a number of archeal, fungal and eukaryotic species and their applicability to gluten detoxification was demonstrated (Scherf et al. 2018). Based on these studies, and parameters such as target specificity, substrate length, optimum pH, and site of action, a PE-Pep from Flavobacterium meningosepticum and a glutamine specific endoprotease from barley (EP-B2) were selected for expression in wheat endosperm (Osorio et al. 2012, 2019). Wheat transformants expressing these glutenases were obtained, and several exhibited significant reductions in the amounts of indigestible gluten peptides separated by SDS-PAGE gels and RP-HPLC (Fig. 5). The gluten detoxification approach has specific advantages. Firstly, some CD patients are sensitive to the HMW subunit peptides (Dewar et al. 2006). Therefore, wheat transformants which lack specific gliadins and/or LMW subunits are still unsuitable for such patients. Secondly, the combination of enzymes used does not digest the gluten proteins within the grain and therefore, does not affect the end-use quality. The glutamine-specific endoprotease used in this study is encoded as a proenzyme, where the propeptide serves as both an inhibitor and chaperone to facilitate spatiotemporal regulation of the proteolytic activity and proper folding of the proteases, respectively (Bethune et al. 2006; Cappetta et al. 2002; Schilling et al. 2009; Cambra et al. 2012). Both of these properties are of immense importance as they prevent degradation of the prolamins in the protein bodies within the grain or during processing (Osorio et al. 2019). In addition, because the PE-Pep has a strict preference for peptide substrates of ≤33 amino acids in size, it can only degrade peptides generated by the glutamine-specific endoprotease (Gass and Khosla 2007), therefore, permitting both of the enzymes to accumulate within the protein bodies without degrading the gluten proteins. Thirdly, this approach does not require consumers to take dietary supplements, which are under different stages of development, so it is expected to be more acceptable to the public.

Differences in the degradation patterns of gluten proteins derived from a transgenic wheat line expressing Flavobacterium meningosepticum prolyl endopeptidase (FME108-10) and its untransformed sibling (FME108-12) under simulated gastro-intestinal conditions with endoprotease B isoform 2 (EP-B2). (a) Gradient tricine gel and densitometric analyses of the digested gliadin fraction derived from FME108-12 with no EP-B2 (left lane and trace), FME108-12 with EP-B2 (middle lane and trace), and FME108-10 with EP-B2 (right lane and trace). (b) Reverse-phase high-performance liquid chromatography of gluten proteins derived from the T2 grains of FME108-12 with EP-B2 (blue line) and FME108-10 with EP-B2 (red line) on a C18 column (modified after Osorio et al. 2019)
7.4 Non-dietary Therapies
In parallel to the efforts to develop dietary therapies for the CD, extensive research has been dedicated to developing non-dietary therapies. These therapies can be largely classified as: (1) luminal therapies based on the detoxification of gluten proteins like enzyme therapy, probiotic therapy, flour/dough pretreatment, and gluten inactivation by polymeric binding; (2) intestinal barrier enhancing therapies, which focus on reducing the permeability of the intestinal epithelial barrier; and (3) immune targeted therapies, which target either CD specific pathways or inflammatory mediators common in gastrointestinal inflammation. These therapies can be subdivided into tTG2 blockers, human leukocyte antigen (HLA) blockers, T-cell targeted therapies, alteration of inflammatory mediators, and vaccine therapy. An up-to-date list of these non-dietary therapies to treat CD which are in various stages of development, is presented in Table 3 and they have been extensively reviewed by Schuppan et al. (2009), Sollid and Khosla (2011), Osorio et al. (2012), Rashtak and Murray (2012), McCarville et al. (2015) and Ribeiro et al. (2018).
8 Conclusion
The recent availability of the reference wheat genome sequence provides knowledge of the complete gene complement of bread wheat including cis-regulatory elements, which will facilitate analysis of the complex transcriptional regulation of the equally complex gene families encoding allergenic and antigenic proteins (International Wheat Genome Sequencing Consortium 2018; Ramírez-González et al. 2018). This knowledge is also expected to facilitate the identification of previously uncharacterized epitopes (Juhász et al. 2018) and the development of novel approaches to produce wheat genotypes safe for all consumers without compromising the organoleptic properties and the end-use quality. The major feat of developing allergen-free and antigen-free celiac-safe wheat could be achieved by a combination of technologies, including genome-editing, genotype and tissue-culture independent genetic transformation procedures, advances in biochemical and immunological detection procedures, and more sensitive and more accurate non-invasive phenotyping methods. All of these approaches are currently being developed and deployed. The challenge will be to bring them together.
References
Afify SM, Pali-Schöll I (2017) Adverse reactions to food: the female dominance – A secondary publication and update. World Allergy Organ J 10:43.
Agostini C, Decsi T, Fewtrell M, Goulet O, Kolacek S, Koletzko B, Fleischer Michaelsen K, Moreno L, Puntis J, Rigo J, Shamir R, Szajewska H, Turck D, van Goudoevero J (2008) Medical Position Paper. Complementary Feeding: A Commentary by the ESPGHAN. Journal of Pediatric Gastroenterology and Nutrition 46:99-110.
Al-toma A, Visser OJ, van Roessel HM, von Blomberg BM, Verbeek WH, Scholten PE, Ossenkoppele GJ, Huijgens PC, Mulder CJ (2007) Autologous hematopoietic stem cell transplantation in refractory celiac disease with aberrant T cells. Blood 109:2243–2249.
Altenbach SB, Allen PV (2011) Transformation of the US bread wheat “Butte 86” and silencing of omega-5 gliadin genes. GM Crops 2:66-73.
Altenbach SB, Tanaka CK, Seabourn BW (2014) Silencing of omega-5 gliadins in transgenic wheat eliminates a major source of environmental variability and improves dough mixing properties of flour. BMC Plant Biol 14:1.
Altenbach SB, Vensel WH, Dupont FM (2011) The spectrum of low molecular weight alpha-amylase/protease inhibitor genes expressed in the US bread wheat Butte 86. BMC Research Notes 4:242.
Anderson RP, Degano P, Godkin AJ, Jewell DP, Hill AV (2000) In vivo antigen challenge in celiac disease identifies a single transglutaminase modified peptide as the dominant A-gliadin T-cell epitope. Nat Med 6:337–342.
Anderson RP, van Heel DA, Tye-Din JA, Barnardo M, Salio M, Jewell DP, Hill AVS (2005) T cells in peripheral blood after gluten challenge in coeliac disease. Gut 54:1217–1223.
Arranz-Otaegui A, Carretero LG, Ramsey MN, Fuller DQ, Richter T (2018) Archaebotanical evidence reveals the origins of bread 14.400 years ago in northeastern Jordan. Proc Natl Acad Sci USA 115:7925-7930.
Ashraf M (2014) Stress-induced changes in wheat grain composition and quality. Crit Rev Food Sci Nutr 54:1576–1583.
Baccioglu A, Kalpaklıoglu F, Altan G (2017) Review of wheat dependent exercise induced anaphylaxis with two cases, and a new co-factor – myorelaxant. J Immunol Clin Res 4:1045.
Bai JC, Ciacci C (2017) World gastroenterology organisation global guidelines: celiac disease February 2017. J Clin Gastroenterol 51:755–768.
Barrett JS, Gibson PR (2012) Fermentable oligosaccharides, disaccharides, monosaccharides and polyols (FODMAPs) and nonallergic food intolerance: FODMAPs or food chemicals. Therap Adv Gastroenterol 5:261-268.
Battais F, Richard C, Jacquenet S, Denery-Papini S, Moneret-Vautrin DA (2008) Wheat grain allergies: an update on wheat allergens. Eur Ann Allergy Clin Immunol 40:67-76.
Becker D, Folck A (2006) Inhibierung der α-Gliadingeneexpression in hexaploiden Brotweizen. Getreidetechnologie 30:153-156.
Becker D, Folck A, Knies P, Lörz H, Wieser H (2006) Silencing the a-gliadins in hexaploid bread wheat. In: Lookhart LG, Ng WPK (eds), Gluten Proteins. AACC International, St Paul, MN, pp. 86-89.
Becker D, Wieser H, Koehler P, Folck A, Mühling KH, Zörb C (2012) Protein composition and techno-functional properties of transgenic wheat with reduced α-gliadin content obtained by RNA interference. J Appl Bot Food Qual 85:23-33.
Bellinghausen I, Weigmann B, Zevallos V, Maxeiner J, Reißig S, Waisman A, Schuppan D, Saloga J (2018) Wheat amylase-trypsin inhibitors exacerbate intestinal and airway allergic immune responses in humanized mice. J Allergy Clin Immunol https://doi.org/10.1016/j.jaci.2018.02.041
Berlin C, Berg EL, Briskin MJ, Andrew DP, Kilshaw PJ, Holzmann B, Weissman IL, Hamann A, Butcher EC (1993) Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 74:185–195.
Bethune MT, Khosla C (2012) Oral enzyme therapy for celiac sprue. Methods Enzymol 502:241-271.
Bethune MT, Strop P, Tang Y, Sollid LM, Khosla C (2006) Heterologous expression, purification, refolding, and structural-functional characterization of EP-B2, a self-activating barley cysteine endoprotease. Chem Biol 13:637–647.
Biesiekierski JR, Muir JG, Gibson PR (2013) Is gluten a cause of gastrointestinal symptoms in people without celiac disease? Curr Allergy Asthma Rep 13:631-638.
Biesiekierski JR, Rosella O, Rose R, Liels K, Barrett JS, Shepherd SJ, Gibson PR, Muir JG (2011). Quantification of fructans, galacto-oligosacharides and other short-chain carbohydrates in processed grains and cereals. J Hum Nutr Diet 24:154-176.
Brans R, Sauer I, Czaja K, Pfützner W, Merk HF (2012) Microarray-based detection of specific IgE against recombinant ω-5-gliadin in suspected wheat-dependent exercise-induced anaphylaxis. Eur J Dermatol 22:358-362.
Brar P, Lee S, Lewis S, Egbuna I, Bhagat G, Green PH (2007) Budesonide in the treatment of refractory coeliac disease. Am J Gastroenterol 102:2265–2269.
Brew-Appiah RAT (2014) Epigenetic and post-transcriptional elimination of celiac-causing wheat storage proteins. Ph.D. Dissertation, Washington State University, Pullman, pp. 187.
Bromilow S, Gethings LA, Buckley M, Bromley M, Shewry PR, Langridge JI, Clare Mills EN (2017) A curated gluten protein sequence database to support development of proteomics methods for determination of gluten in gluten-free foods. J Proteomics 163:67-75.
Brouns F, Delzenne N, Gibson G (2017) The dietary fibers – FODMAPs controversy. Cereal Foods World 62:98-103.
Brouns FJPH, van Buul VJ, Shewry PR (2013) Does wheat make us fat and sick? J Cereal Sci 58:209-215.
Bustamante MÁ, Fernández-Gil MP, Churruca I, Miranda J, Lasa A, Navarro V, Simón E (2017) Evolution of Gluten Content in Cereal-Based Gluten-Free Products: An Overview from 1998 to 2016. Nutrients 9:21.
Caio G, Volta U, Tovoli F, De Giorgio R (2014) Effect of gluten free diet on immune response to gliadin in patients with non-celiac gluten sensitivity. BMC Gastroenterol 14:26.
Cambra I, Hernández D, Diaz I, Martinez M (2012) Structural basis for specificity of propeptide-enzyme interaction in barley C1A cysteine peptidases. PLoS One 7:e37234.
Camerlengo F, Sestili F, Silvestri M, Colaprico G, Margiotta B, Ruggeri R, Lupi R, Masci S, Lafiandra D (2017) Production and molecular characterization of bread wheat lines with reduced amount of α-type gliadins. BMC Plant Biol 17:248.
Cappetta M, Roth I, Díaz A, Tort J, Roche L (2002) Role of the prosegment of Fasciola hepatica cathepsin L1 in folding of the catalytic domain. Biol Chem 383:1215-1221.
Carbonero P, Garcia-Olmedo F (1999) A multigene family of trypsin/α-amylase inhibitors from cereals. In: Shewry PR, Casey R (eds) Seed proteins. Kluwer Publishing, Surrey, U.K., pp. 617–634.
Carnevali A, Gianotti A, Benedett S, Tagliamonte MC, Primiterra M, Laghi L, Danesi F, Valli V, Ndaghijimana M, Capozzi F, Canestrari F, Bordoni A (2014) Role of Kamut® brand khorasan wheat in the counteraction of non-celiac wheat sensitivity and oxidative damage. Food Res Int 63:218–226.
Cataldo F, Montalto G (2007) Celiac disease in the developing countries: a new and challenging public health problem. World J Gastroenterol 13:2153-2159.
Catassi C (2015) Gluten Sensitivity. Ann Nutr Metab 67(suppl 2):16–26.
Catassi C, Alaedini A, Bojarski C, Bonaz B, Bouma G, Carroccio A, Castillejo G, De Magistris L, Dieterich W, Di Liberto D, Elli L, Fasano A, Hadjivassiliou M, Kurien M, Lionetti E, Mulder CJ, Rostami K, Sapone A, Scherf K, Schuppan D, Trott N, Volta U, Zevallos V, Zopf Y, Sanders DS (2017) The Overlapping Area of Non-Celiac Gluten Sensitivity (NCGS) and Wheat-Sensitive Irritable Bowel Syndrome (IBS): An Update. Nutrients 9:1268.
Catassi C, Fabiani E, Iacono G, D’Agate C, Francavilla R, Biagi F, Volta U, Accomando S, Picarelli A, De Vitis I, Pianelli G, Gesuita R, Carle F, Mandolesi A, Bearzi I, Fasano A (2007a) A prospective, double-blind, placebo-controlled trial to establish a safe gluten threshold for patients with celiac disease. Am J Clin Nutr 85:160-166.
Catassi C, Fasano A (2008) Celiac disease. In: Arendt EK, Dal Bello F (eds) Gluten-free cereal products and beverages. Elsevier Inc., pp. 1–26.
Catassi C, Gatti S, Lionetti E (2015) World perspective and celiac disease. Epidemiology Dig Dis 33:141–146.
Catassi C, Kryszak D, Louis-Jacques O, Duerksen DR, Hill I, Crowe SE, Brown AR, Procaccini NJ, Wonderly BA, Hartley P, Moreci J, Bennett N, Horvath K, Burk M, Fasano A (2007b) Detection of Celiac disease in primary care: a multicenter case-finding study in North America. Am J Gastroenterol 102:1454-1460.
Catassi C, Rossini M, Rätsch IM, Bearzi I, Santinelli A, Castagnani R, Pisani E, Coppa GV, Giorgi PL (1993) Dose dependent effects of protracted ingestion of small amounts of gliadin in coeliac disease children: a clinical and jejunal morphometric study. Gut 34:1515-1519.
Ciacci C, Ciclitira P, Hadjivassiliou M, Kaukinen K, Ludvigsson JF, McGough N, Sanders DS, Woodward J, Leonard JN, Swift GL (2015) The gluten-free diet and its current application in coeliac disease and dermatitis herpetiformis. United European Gastroenterol J 3:121–135.
Ciacci C, Maiuri L, Russo I, Tortora R, Bucci C, Cappello C, Santonicola A, Luciani A, Passananti V, Iovino P (2009) Efficacy of budesonide therapy in the early phase of treatment of adult coeliac disease patients with malabsorption: an in vivo / in vitro pilot study. Clin Exp Pharmacol Physiol 36:1170–1176.
Cianferoni A (2016) Wheat allergy: diagnosis and management. J Asthma Allergy 9:13-25.
Ciclitira PJ, Cerio R, Ellis HJ, Maxton D, Nelufer JM, Macartney JM (1985) Evaluation of a gliadin-containing gluten-free product in coeliac patients. Hum Nutr Clin Nutr 39:303-308.
Ciclitira PJ, Evans DJ, Fagg NL, Lennox ES, Dowling RH (1984) Clinical testing of gliadin fractions in coeliac patients. Clin Sci (Lond) 66:357-364.
Ciclitira PJ, Hunter JO, Lennox ES (1980a) Clinical testing of bread made from nullisomic-6A wheats in celiac patients. Lancet 2:234-236.
Ciclitira PJ, Hunter JO, Lennox ES (1980b) Clinical testing in celiac patients of bread made from wheats deficient in some α-gliadins. Clinical Sci 59:25.
Colomba MS, Gregorini A (2012) Are ancient durum wheats less toxic to celiac patients? A study of α-gliadin from Graziella Ra and Kamut. ScientificWorldJournal 2012:837416.
Comino I, Moreno M, Real A, Rodríguez-Herrera A, Barro F, Sousa C (2013) The gluten-free diet: Testing alternative cereals tolerated by celiac patients. Nutrients 5:4250-4268.
Cook F, Hughes N, Nibau C, Orman-Ligeza B, Schatlowski N, Uauy C, Trafford K (2018) Barley lys3 mutants are unique amongst shrunken-endosperm mutants in having abnormally large embryos. J Cereal Sci 82:16-24.
Cordain L (1999) Cereal grains: humanity’s double-edged sword. World Rev Nutr Diet 84:19–73.
Croese J, O’Neil J, Masson J, Cooke S, Melrose W, Pritchard D, Speare R (2006) A proof of concept study establishing Necator americanus in Crohn’s patients and reservoir donors. Gut 55:136–137.
Cukrowska B, Sowińska A, Bierła JB, Czarnowska E, Rybak A, Grzybowska-Chlebowczyk U (2017) Intestinal epithelium, intraepithelial lymphocytes and the gut microbiota - Key players in the pathogenesis of celiac disease. World J Gastroenterol 23:7505–7518.
De Palma G, Nadal I, Carmen Collado M, Sanz Y, Collado M (2009) Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br J Nutr 102:1154–1160.
Dewar DH, Amato M, Ellis HJ, Pollock EL, Gonzalez-Cinca N, Wieser H, Ciclitira PJ (2006) The toxicity of high molecular weight glutenin subunits of wheat to patients with coeliac disease. Eur J Gastroenterol Hepatol 18:483-491.
Di Sabatino A, Rovedatti L, Rosado MM, Carsetti R, Corazza GR, MacDonald TT (2009) Increased expression of mucosal addressin cell adhesion molecule 1 in the duodenum of patients with active celiac disease is associated with depletion of integrin alpha4beta7-positive T cells in blood. Hum Pathol 40:699–704.
DiGiacomo DV, Tennyson CA, Green PH, Demmer RT (2013) Prevalence of gluten-free diet adherence among individuals without celiac disease in the USA: results from the Continuous National Health and Nutrition Examination Survey 2009–2010. Scand J Gastroenterol 48:921-925.
Do AB, Khuda SE, Sharma GM (2018) Undeclared food allergens and gluten in commercial food products analyzed by ELISA. J AOAC Int 101:23-35.
Dupont FM, Vensel WH, Tanaka CK, Hurkman WJ, Altenbach SB (2011) Deciphering the complexities of the wheat flour proteome using quantitative two-dimensional electrophoresis, three proteases and tandem mass spectrometry Proteome Sci 9:10.
Edwards JC, Szczepanski L, Szechinski J, Filipowicz-Sosnowska A, Emery P, Close DR, Stevens RM, Shaw T (2004) Efficacy of B-celltargeted therapy with rituximab in patients with rheumatoid arthritis. N Engl J Med 350:2572–2581.
Ehren J, Moron B, Martin E, Bethune MT, Gray GM, Khosla C (2009) A food-grade enzyme preparation with modest gluten detoxification properties. PLoS ONE 4:e6313.
Ejderhamn J, Veesess B, Strandvik B (1988) The long term effect of continual ingestion of wheat starch-containing gluten-free products in celiac patients. In: Kumar PJ (ed) Coeliac disease: one hundred years. Leeds, United Kingdom, Leeds University Press, pp. 294–297.
Engstrom N, Saenz-Mendez P, Scheers J, Scheers N (2017) Towards Celiac-safe foods: Decreasing the affinity of transglutaminase 2 for gliadin by addition of ascorbyl palmitate and ZnCl2 as detoxifiers. Sci Rep 7:77.
Escarnot E, Gofflot S, Sinnaeve G, Dubois B, Bertin P, Mingeot D (2018) Reactivity of gluten proteins from spelt and bread wheat accessions towards A1 and G12 antibodies in the framework of celiac disease. Food Chem 268:522–532.
Farage P, de Medeiros Nóbrega YK, Pratesi R, Gandolfi L, Assunção P, Zandonadi RP (2017) Gluten contamination in gluten-free bakery products: a risk for coeliac disease patients. Public Health Nutr 20:413-416.
Fasano A (2007) https://www.slideshare.net/saturni/gliadin-intestinal-permeability-and-celiac-disease-from-innate-immunity-to-autoimmunity (retried August 2018).
Fasano A (2012) Leaky gut and autoimmune diseases. Clinic Rev Allerg Immunol 42:71-78.
Frisoni M, Corazza GR, Lafiandra D, De Ambroggio E, Filipponi C, Bonvicini F, Borasio E, Porcheddu E, Gasbarrini G (1995) Wheat deficient in gliadins: promising tool for coeliac disease. Gut 36:375-378.
Fritschy F, Windemann H, Baumgarten E (1985) Bestimmung von Weizen gliadinen in Lebensmitteln mittels ELISA [Determination of wheat gliadins in foods by ELISA]. Z Lebensm Unters Forsch 181: 379-385.
Gass J, Bethune MT, Siegel M, Spencer A, Khosla C (2007) Combination enzyme therapy for gastric digestion of dietary gluten in patients with celiac sprue. Gastroenterology 133:472-480
Gass J, Khosla C (2007) Prolyl endopeptidases. Cell Mol Life Sci 64:345-355.
Gélinas P, Gagnon F (2018) Inhibitory activity towards human α-amylase in wheat flour and gluten. Food Sci Technol 53:467-474.
Gelinas P, McKinnon C (2016) Gluten weight in ancient and modern wheat and the reactivity of epitopes towards R5 and G12 monoclonal antibodies. Int J Food Sci Technol 51:1801–1810.
Gessendorfer B, Hartmann G, Wieser H, Koehler P (2011) Determination of celiac disease-specific peptidase activity of germinated cereals. Eu Food Res Technol 232:205–209.
Ghosh S, Goldin E, Gordon FH, Malchow HA, Rask-Madsen J, Rutgeerts P, Vyhnálek P, Zádorová Z, Palmer T, Donoghue S; Natalizumab Pan-European Study Group (2003) Natalizumab for active Crohn’s disease. N Engl J Med 348:24–32.
Gianfrani C, Camarca A, Mazzarella G, Di Stasio L, Giardullo N, Ferranti P, Picariello G, Rotondi Aufiero V, Picascia S, Troncone R, Pogna N, Auricchio S, Mamone G (2015) Extensive in vitro gastrointestinal digestion markedly reduces the immune-toxicity of Triticum monococcum wheat: Implication for celiac disease. Mol Nutr Food Res 59:1844-1854.
Gibert A, Espadaler M, Angel Canela M, Sánchez A, Vaqué C, Rafecas M (2006) Consumption of gluten-free products: should the threshold value for trace amounts of gluten be at 20, 100 or 200 p.p.m.? Eur J Gastroenterol Hepatol 18:1187-1195.
Gil-Humanes J, Piston F, Altamirano-Fortoul R, Real A, Comino I, Sousa A, Rosell CM, Barro F (2014) Reduced-gliadin wheat bread: an alternative to the gluten-free diet for consumers suffering gluten-related pathologies. PLoS One 9:e90898.
Gil-Humanes J, Piston F, Hernando A, Alvarez JB, Shewry PR, Barro F (2008) Silencing of γ-gliadins by RNA interference (RNAi) in bread wheat. J Cereal Sci 48:565-568.
Gil-Humanes J, Pistón F, Shewry PR, Tosi P, Barro F (2011) Suppression of gliadins results in altered protein body morphology in wheat. J Exp Bot 62:4203-4213.
Gil-Humanes J, Piston F, Tollefsen S, Sollid LM, Barro F (2010) Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference. Proc Natl Acad Sci USA 107:17023-17028.
Gilissen LJWJ, van der Meer IM, Smulders MJM (2014) Reducing the incidence of allergy and intolerance to cereals. J Cereal Sci 59:337-353.
Gillett HR, Arnott ID, McIntyre M, Campbell S, Dahele A, Priest M, Jackson R, Ghosh S (2002) Successful infliximab treatment for steroid-refractory celiac disease: a case report. Gastroenterology 122:800–805.
Godfrey D, Hawkesford MJ, Powers SJ, Millar S, Shewry PR (2010) Effects of crop nutrition on wheat grain composition and end use quality. J Agric Food Chem 58:3012–3021.
Goryunova SV, Salentijn EM, Chikida NN, Kochieva EZ, van der Meer IM, Gilissen LJ, Smulders MJ (2012) Expansion of the gamma-gliadin family in Aegilops and Triticum. BMC Evol Biol 12:215.
Gregorini A, Colomba M, Ellis HJ, Ciclitira PJ (2009) Immunogenicity characterization of two ancient wheat α-gliadin peptides related to coeliac disease. Nutrients 1:276-290.
Hadjivassiliou M, Sanders D, Grünewald RA, Woodroofe N, Boscolo S, Aeschlimann D (2010) Gluten sensitivity: from gut to brain. Lancet Neurol 9:318–330.
Hadjivassiliou M, Sanders DD, Aeschlimann DP (2015) Gluten-Related Disorders: Gluten Ataxia. Dig Dis 33:264–268.
Hajas L, Scherf KA, Török K, Bugyi Z, Schall E, Poms RE, Koehler P, Tömösközi S (2018) Variation in protein composition among wheat (Triticum aestivum L.) cultivars to identify cultivars suitable as reference material for wheat gluten analysis. Food Chem 267:387–394.
Hauser SL, Waubant E, Arnold DL, Vollmer T, Antel J, Fox RJ, Bar-Or A, Panzara M, Sarkar N, Agarwal S, Langer-Gould A, Smith CH; HERMES Trial Group (2008) B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N Engl J Med 358:676–688.
Henry AG, Brooks AS, Piperno DR (2011) Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium). Proc Natl Acad Sci USA 108:486–491.
Henry AG, Brooks AS, Piperno DR (2014) Plant foods and the dietary ecology of Neanderthals and early modern humans. J Human Evolution 69:44-54.
Hernando A, Mujico JR, Mena MC, Lombardía M, Méndez E (2008) Measurement of wheat gluten and barley hordeins in contaminated oats from Europe, the United States and Canada by Sandwich R5 ELISA. Eur J Gastroenterol Hepatol 20:545-554.
Herold KC, Gitelman S, Greenbaum C, Puck J, Hagopian W, Gottlieb P, Sayre P, Bianchine P, Wong E, Seyfert-Margolis V, Bourcier K, Bluestone JA; Immune Tolerance Network ITN007AI Study Group. (2009) Treatment of patients with new onset Type 1 diabetes with a single course of anti-CD3 mAb Teplizumab preserves insulin production for up to 5 years. Clin Immunol 132:166–173.
Hischenhuber C, Crevel R, Jarry B, Mäki M, Moneret-Vautrin DA, Romano A, Troncone R, Ward R (2006) Review article: safe amounts of gluten for patients with wheat allergy or coeliac disease. Aliment Pharmacol Ther 23:559-575.
Huebener S, Tanaka CK, Uhde M, Zone JJ, Vensel WH, Kasarda DD, Beams L, Briani C, Green PH, Altenbach SB, Alaedini A (2015) Specific nongluten proteins of wheat are novel target antigens in celiac disease humoral response. J Proteome Res14:503−511.
Huibregtse IL, Marietta EV, Rashtak S, Koning F, Rottiers P, David CS, van Deventer SJ, Murray JA (2009) Induction of antigen specific tolerance by oral administration of Lactococcus lactis delivered immunodominant DQ8-restricted gliadin peptide in sensitized nonobese diabetic Abo DQ8 transgenic mice. J Immunol 183:2390–2396.
Inomata N (2009) Wheat allergy. Curr Opin Allergy Clin Immunol 9:238–243.
International Wheat Genome Sequencing Consortium (2018) Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 361:eaar7191.
Ivarsson A1, Myléus A, Norström F, van der Pals M, Rosén A, Högberg L, Danielsson L, Halvarsson B, Hammarroth S, Hernell O, Karlsson E, Stenhammar L, Webb C, Sandström O, Carlsson A (2013) Prevalence of childhood celiac disease and changes in infant feeding. Pediatrics 131:e687-e694.
Janssen FW, Hägele GH, de Baaij JA (1991) Gluten free products, the Dutch experience. In: Mearin ML, Mulder CJJ (eds) Coeliac Disease. Dordrech: Kluwer Academic, pp. 95–100.
Jenkins JA, Griffiths-Jones S, Shewry PR, Breiteneder H, Mills ENC (2005) Structural relatedness of plant food allergens with specific reference to cross-reactive allergens: an in silico analysis. J Allergy Clin Immunol 115:163–170.
Jouanin A, Gilissen LJWJ, Boyd LA, Cockram J, Leigh FJ, Wallington EJ, van den Broeck HC, van der Meer IM, Schaart JG, Visser RGF, Smulders MJM (2018) Food processing and breeding strategies for coeliac-safe and healthy wheat products. Food Res Int1 10:11-21.
Juhász A, Belova T, Florides CG, Maulis C, Fischer I, Gell G, Birinyi Z, Ong J, Keeble-Gagnère G, Maharajan A, Ma W, Gibson P, Jia J, Lang D, Mayer KFX, Spannagl M, International Wheat Genome Sequencing Consortium, Tye-Din JA, Appels R, Olsen O-A (2018) Genome mapping of seed-borne allergens and immunoresponsive proteins in wheat. Sci Adv 4:eaar8602.
Juhász A, Haraszi R, Maulis C (2015) ProPepper: a curated database for identification and analysis of peptide and immune-responsive epitope composition of cereal grain protein families. Database 2015:1–16.
Junker Y, Zeissig S, Kim SJ, Barisani D, Wieser H, Leffler DA, Zevallos V, Libermann TA, Dillon S, Freitag TL, Kelly CP, Schuppan D (2012) Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J Exp Med 209:2395-2408.
Kabbani TA, Goldberg A, Kelly CP, Pallav K, Tariq S, Peer A, Hansen J, Dennis M, Leffler DA (2012) Body mass index and the risk of obesity in coeliac disease treated with the gluten-free diet. Aliment Pharmacol Ther 35:723-729.
Kapoerchan VV, Wiesner M, Overhand M, van der Marel GA, Koning F, Overkleeft HS (2008) Design of azidoproline containing gluten peptides to suppress CD4+ T-cell responses associated with coeliac disease. Bioorg Med Chem 16:2053–2062.
Kasarda DD (2013) Can an increase in celiac disease be attributed to an increase in the gluten content of wheat as a consequence of wheat breeding? J Agric Food Chem 61:1155-1159.
Kasarda DD, Adalsteins E, Lew EJ-L, Lazo GR, Altenbach SB (2013) Farinin:characterisation of a novel wheat endosperm protein belonging to the prolamin superfamily J Agric Food Chem 61:2407-2417.
Kaukinen K, Collin P, Holm K, Rantala I, Vuolteenaho N, Reunala T, Mäki M (1999) Wheat starch-containing gluten-free flour products in the treatment of coeliac disease and dermatitis herpetiformis. A long-term follow-up study. Scand J Gastroenterol 34:163-169.
Keech CL, Dromey J, Chen Z, Anderson RP, McCluskey J (2009) Immune tolerance induced by peptide immunotherapy in an HLA-DQ2-dependent mouse model of gluten immunity. Gastroenterology 136:A355.
Kneen E, Sandstedt RM (1946) Distribution and general properties of an amylase inhibitor in cereals. Arch Biochem 9:235–249.
Koning F (2012) Celiac disease: quantity matters. Semin Immunopathol 34:541–549.
Kotaniemi-Syrjänen A, Palosuo K, Jartti T, Kuitunen M, Pelkonen AS, Mäkelä MJ (2010) The prognosis of wheat hypersensitivity in children. Pediatr Allergy Immunol 21(2 Pt 2):e421-e428.
Lähdeaho ML, Kaukinen K, Laurila K, Vuotikka P, Koivurova OP, Kärjä-Lahdensuu T, Marcantonio A, Adelman DC, Mäki M (2014) Glutenase ALV003 attenuates gluten-induced mucosal injury in patients with celiac disease. Gastroenterology 146:1649-1658.
Lebwohl B, Sanders DS, Green PHR (2018) Coeliac disease. Lancet 391:70-81.
Lee HJ, Anderson Z, Ryu D (2014) Gluten Contamination in Foods Labeled as ‘Gluten Free’ in the United States. J Food Prot 77:1830–1833.
Lee L, Zhang Y, Ozar B, Sensen CW, Schriemer DC (2016) Carnivorous Nutrition in Pitcher Plants (Nepenthes spp.) via an Unusual Complement of Endogenous Enzymes. J Proteome Res 15:3108-3117.
Leonard MM, Sapone A, Catassi C, Fasano A (2017) Celiac disease and nonceliac gluten sensitivity: A review. JAMA 318:647-656.
Lionetti E, Gatti S, Pulvirenti A, Catassi C (2015) Celiac disease from a global perspective. Best Pract Res Clin Gastroenterol 29:365-379.
Ludvigsson JF, Fasano A (2012) Timing of introduction of gluten and celiac disease risk. Ann Nutr Metab 60 Suppl 2:22-29.
Ludvigsson JF, Leffler DA, Bai JC, Biagi F, Fasano A, Green PH, Hadjivassiliou M, Kaukinen K, Kelly CP, Leonard JN, Lundin KE, Murray JA, Sanders DS, Walker MM, Zingone F, Ciacci C (2013) The Oslo definitions for coeliac disease and related terms. Gut 62:43-52.
Mamone G, Picariello G, Addeo F, Ferranti P (2011) Proteomic analysis in allergy and intolerance to wheat products. Expert Rev Proteomics 8:95-115.
Marion D, Douliez J-P, Gautier M-F, Elmorjani K (2004) Plant lipid transfer proteins: relationships between allergenicity and structural, biological and technological properties. In: Mills ENC, Shewry PR (eds), Plant Food Allergens, Oxford: Blackwell Publishing, pp. 57-69.
Martin J, Geisel T, Maresch C, Krieger K, Stein J (2013) Inadequate nutrient ixntake in patients with celiac disease: results from a German dietary survey. Digestion 87:240-246.
Matsuda T, Nakase M, Alvarez AM, Izumi H, Kato T, Tada Y (2006) Rice-seed allergenic proteins and hypoallergenic rice. In: Mine Y, Shahidi F (eds) Nutraceutical proteins and peptides in health and disease, CRC Press, pp. 493–511.
McCarville JL, Caminero A, Verdu EF (2015) Celiac treatments, adjuvant therapies and alternatives to the gluten-free diet. In: Arranz E, Fernández-Bañares F, Rosell CM, Rodrigo L, Peña AS (eds) Advances in the understanding of gluten related pathology and the Evolution of Gluten-Free Foods. OmniaScience, Barcelona, Spain, pp. 223-253.
Mei HE, Frölich D, Giesecke C, Loddenkemper C, Reiter K, Schmidt S, Feist E, Daridon C, Tony HP, Radbruch A, Dörner T (2010) Steady state generation of mucosal IgA+ plasmablasts is not abrogated by B cell depletion therapy with rituximab. Blood 116:5181–5190.
Metakovsky EV, Davidov SD, Chernakov VM, Upelniek VP (1993) Gliadin allele identification in common wheat. III. Frequency of occurrence and appearance of spontaneous mutations at the gliadin-coding loci. J Genet Breed 47:221-236.
Miranda J, Simón E (2017) Gluten Content Change Over the Two Last Decades. In: E. Simón et al. (eds) Nutritional and Analytical Approaches of Gluten-Free Diet in Celiac Disease, SpringerBriefs in Food, Health, and Nutrition, Springer, pp. 47-57.
Mitea C, Havenaar R, Drijfhout JW, Edens L, Dekking L, Koning F (2008) Efficient degradation of gluten by a prolyl endoprotease in a gastrointestinal model: Implications for coeliac disease. Gut 57:25–32.
Mitea C, Salentijn EM, van Veelen P, Goryunova SV, van der Meer IM, van den Broeck HC, Mujico JR, Montserrat V, Gilissen LJ, Drijfhout JW, Dekking L, Koning F, Smulders MJ (2010) A universal approach to eliminate antigenic properties of alpha-gliadin peptides in celiac disease. PloS One 5:e15637.
Moehs CP, Austil WJ, Holm A, Large TAG, Loeffler D, Mullenberg J, Schnable PS, Skinner W, van Boxtel J, Wu L, McGuire C (2018) Development of reduced gluten wheat enabled by determination of the genetic basis of the lys3a low hordein barley mutant. bioRxiv https://doi.org/10.1101/354548
Molberg Ø, Uhlen AK, Jensen T, Flæte NS, Fleckenstein B, Arentz-Hansen H, Raki M, Lundin KEA, Sollid LM (2005) Mapping of gluten T-cell epitopes in the bread wheat ancestors: implications for celiac disease. Gasteroenterology 128:393-401.
Morris CF (2002) Puroindolines: The molecular genetic basis of wheat grain hardness. Plant Mol Biol 48:633-647.
Munck L (1992) The case of high-lysine barley breeding. In: Shewry PR (ed), Barley: Genetics, Biochemistry, Molecular Biology and Biotechnology. CAB International, Wallingford Oxon, pp. 573-601.
Nadal I, Donat E, Ribes-Koninckx C, Calabuig M, Sanz Y (2007) Imbalance in the composition of the duodenal microbiota of children with coeliac disease. J Med Microbiol 56:1669–1674.
Nambu M (2006) Rice Allergy. Pediatrics 117:2331.
Nistal E, Caminero A, Herrán AR, Pérez-Andres J, Vivas S, Ruiz de Morales JM, Sáenz de Miera LE, Casqueiro J (2016) Study of duodenal bacterial communities by 16S rRNA gene analysis in adults with active celiac disease vs non-celiac disease controls. J Appl Microbiol 120:1691–1700.
Nistal E, Caminero A, Vivas S, Ruiz de Morales JM, Sáenz de Miera LE, Rodríguez-Aparicio LB, Casqueiro J (2012) Differences in faecal bacteria populations and faecal bacteria metabolism in healthy adults and celiac disease patients. Biochimie 94:1724–1729.
Olexová L, Dovičovičová L, Švec M, Siekel P, Kuchta T (2006) Detection of gluten-containing cereals in flours and “gluten-free” bakery products by polymerase chain reaction. Food Control 17:234-237.
Olivares M, Walker AW, Capilla A, Benítez-Páez A, Palau F, Parkhill J, Castillejo G, Sanz Y (2018) Gut microbiota trajectory in early life may predict development of celiac disease. Microbiome 6:36.
Ortiz-Sánchez JP, Cabrera-Chávez F, de la Barca AM (2013) Maize prolamins could induce a gluten-like cellular immune response in some celiac disease patients. Nutrients 5:4174-4183.
Osborne TB (1924) The vegetable proteins, 2nd edition. Longmans Green & Co, London, UK.
Osorio C, Wen N, Gemini R, Zemetra R, von Wettstein D, Rustgi S (2012) Targeted modification of wheat grain protein to reduce the content of celiac causing epitopes. Funct Integr Genomics 12:417-438.
Osorio CE, Wen N, Mejias JH, Liu B, Reinbothe S, von Wettstein D, Rustgi S (2019) Development of wheat genotypes expressing a glutamine-specific endoprotease from barley and a prolyl endopeptidase from Flavobacterium meningosepticum or Pyrococcus furiosus as a potential remedy to celiac disease. Funct Integr Genomics 19:123–136.
Panda R, Taylor SL, Goodman RE (2010) Development of a Sandwich Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Buckwheat Residues in Food. J Food Sci 75:T110-T117.
Pasha I, Saeed F, Tauseef Sultan M, Batool R, Aziz M, Ahmed W (2016) Wheat allergy and intolerance; Recent updates and perspectives. Crit Rev Food Sci Nutr 56:13–24.
Pastorello EA, Farioli L, Conti A, Pravettoni V, Bonomi S, Iametti S, Fortunato D, Scibilia J, Bindslev-Jensen C, Ballmer-Weber B, Robino AM, Ortolani C (2007) Wheat IgE-mediated food allergy in European patients: alpha-amylase inhibitors, lipid transfer proteins and low-molecular-weight glutenins. Allergenic molecules recognized by double-blind, placebo-controlled food challenge. International archives of allergy and immunology 144:10–22.
Paterson BM, Lammers KM, Arrieta MC, Fasano A, Meddings JB (2007) The safety, tolerance, pharmacokinetic and pharmacodynamic effects of single doses of AT-1001 in coeliac disease subjects: a proof of concept study. Aliment Pharmacol Ther 26:757–766.
Peräaho M, Kaukinen K, Paasikivi K, Sievänen H, Lohiniemi S, Mäki M, Collin P (2003) Wheat-starch-based gluten-free products in the treatment of newly detected coeliac disease: prospective and randomized study. Aliment Pharmacol Ther 17:587–594.
Perez-Gregorio MR, Días R, Mateus N, de Freitas V (2018) Identification and characterization of proteolytically resistant gluten-derived peptides Food Funct 9:1726–1735.
Pinier M, Verdu EF, Nasser-Eddine M, David CS, Vézina A, Rivard N, Leroux JC (2009) Polymeric binders suppress gliadin-induced toxicity in the intestinal epithelium. Gastroenterology 136:288–298.
Pinto-Sánchez MI, Verdu EF, Liu E, Bercik P, Green PH, Murray JA, Guandalini S, Moayyedi P (2016) Gluten introduction to infant feeding and risk of celiac disease: Systematic review and meta-analysis. J Pediatr 168:132–43.
Piston F, Gil-Humanes J, Rodríguez-Quijano M, Barro F (2011) Down-regulating γ-gliadins in bread wheat leads to non-specific increases in other gluten proteins and has No major effect on dough gluten strength. PLoS One 6:e24754.
Pizzuti D, Buda A, D’Odorico A, D’Incà R, Chiarelli S, Curioni A, Martines D (2006) Lack of intestinal mucosal toxicity of Triticum monococcum in coeliac disease patients. Scand J Gastroenterol 41:1305–1311.
Pogna NE, Monari AM, Cacciatori P, Redaelli R, Ng PKW (1998) Development and characterization of bread wheat lines lacking chromosome 1B-, 1D-, 6A- and 6D-encoded prolamins. In: Proc. IXth Intern. Wheat Genetics Symposium, Saskatoon, Saskatchewan, Canada, pp. 265-268.
Pontieri P, Mamone G, De Caro S, Tuinstra MR, Roemer E, Okot J, De Vita P, Ficco DB, Alifano P, Pignone D, Massardo DR, Del Giudice L (2013) Sorghum, a healthy and gluten-free food for celiac patients as demonstrated by genome, biochemical, and immunochemical analyses. J Agric Food Chem 61:2565-2571.
Prandi B, Faccini A, Tedeschi T, Galaverna G, Sforza S (2013) LC/MS analysis of proteolytic peptides in wheat extracts for determining the content of the allergen amylase/trypsin inhibitor CM3: Influence of growing area and variety. Food Chem140:141-146.
Prandi B, Tedeschi T, Folloni S, Galaverna G, Sforza S (2017) Peptides from gluten digestion: A comparison between old and modern wheat varieties. Food Res Int 91:92–102.
Ramírez-González RH, Borrill P, Lang D, Harrington SA, Brinton J, Venturini L, Davey M, Jacobs J, van Ex F, Pasha A, Khedikar Y, Robinson SJ, Cory AT, Florio T, Concia L, Juery C, Schoonbeek H, Steuernagel B, Xiang D, Ridout CJ, Chalhoub B, Mayer KFX, Benhamed M, Latrasse D, Bendahmane A; International Wheat Genome Sequencing Consortium, Wulff BBH, Appels R, Tiwari V, Datla R, Choulet F, Pozniak CJ, Provart NJ, Sharpe AG, Paux E, Spannagl M, Bräutigam A, Uauy C (2018) The transcriptional landscape of polyploid wheat. Science 361:eaar6089.
Rashtak S, Murray JA (2012) Review article: coeliac disease, new approaches to therapy. Aliment Pharmacol Ther 35:768-781.
Redaelli R, Metakovsky EV, Davydov SD, Pogna NE (1994) Two-dimensional mapping of gliadins using biotypes and null mutants of common wheat cultivar Saratovskaya 29. Hereditas 121:131-137.
Reinisch W, de Villiers W, Bene L, Simon L, Rácz I, Katz S, Altorjay I, Feagan B, Riff D, Bernstein CN, Hommes D, Rutgeerts P, Cortot A, Gaspari M, Cheng M, Pearce T, Sands BE (2010) Fontolizumab inmoderate to severe Crohn’s disease: a phase 2, randomized, double-blind, placebo-controlled, multiple-dose study. Inflamm Bowel Dis 16:233–242.
Ribeiro M, Nunes FM, Rodriguez-Quijano M, Carrillo JM, Branlard G, Igrejas G (2018) Next-generation therapies for celiac disease: The gluten-targeted approaches. Trends Food Sci Technol 75:56-71.
Ribeiro M, Rodriguez-Quijano M, Nunes FM, Carrillo JM, Branlard G, Igrejas G (2016) New insights into wheat toxicity: Breeding did not seem to contribute to a prevalence of potential celiac disease’s immunostimulatory epitopes. Food Chem 213:8–18.
Rosella CM, Barro F, Sousa C, Mena MC (2014) Cereals for developing gluten-free products and analytical tools for gluten detection. J Cereal Sci 59:354-364.
Rubio-Tapia A, Ludvigsson JF, Brantner TL, Murray JA, Everhart JE (2012) The prevalence of celiac disease in the United States. Am J Gastroenterol 107:1538-1544.
Rustgi S, Wen N, Osorio C, Brew-Appiah RAT, Wen S, Gemini R, Mejias JH, Ankrah N, Moehs CP, von Wettstein D (2014) Natural dietary therapies for the ‘gluten syndrome’, Scientia Danica, Series B, Biologica 3:1-87.
Salcedo G, Sanchez-Monge R, Garcia-Casado G, Armentia A, Gomez L, Barber D (2004) The cereal α-amylase/trypsin inhibitor family associated with bakers’ asthma and food allergy. In Mills ENC, Shewry PR (eds) Plant food allergens. Blackwell Science, Oxford U.K., pp. 70-86.
Salentijn EM, Esselink DG, Goryunova SV, van der Meer IM, Gilissen LJ, Smulders MJ (2013) Quantitative and qualitative differences in celiac disease epitopes among durum wheat varieties identified through deep RNA-amplicon sequencing. BMC Genomics 14:905.
Salentijn EM, Goryunova SV, Bas N, van der Meer IM, van den Broeck HC, Bastien T, Gilissen LJ, Smulders MJ (2009) Tetraploid and hexaploid wheat varieties reveal large differences in expression of alpha-gliadins from homoeologous Gli-2 loci. BMC Genomics 10:48.
Salmi M, Jalkanen S (1999) Molecules controlling lymphocyte migration to the gut. Gut 45:148–153.
Sanchez de la Hoz P, Castagnaro A, Carbonero P (1994) Sharp divergence between wheat and barley at loci encoding novel members of the trypsin/alpha-amylase inhibitors family. Plant Mol Biol 26:1231-1236.
Sánchez-León S, Gil-Humanes J, Ozuna CV, Giménez MJ, Sousa C, Voytas DF, Barro F (2018) Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol J 16:902-910.
Sandborn WJ, Colombel JF, Frankel M, Hommes D, Lowder JN, Mayer L, Plevy S, Stokkers P, Travis S, Van Assche G, Baumgart DC, Targan SR (2010) Anti-CD3 antibody visilizumab is not effective in patients with intravenous corticosteroid-refractory ulcerative colitis. Gut 59:1485–1492.
Sapone A, Bai JC, Ciacci C, Dolinsek J, Green PH, Hadjivassiliou M, Kaukinen K, Rostami K, Sanders DS, Schumann M, Ullrich R, Villalta D, Volta U, Catassi C, Fasano A (2012) Spectrum of gluten-related disorders: consensus on new nomenclature and classification. BMC Med 10:13.
Scherf KA, Brockow K, Biedermann T, Koehler P, Wieser H (2016) Wheat-dependent exercise-induced anaphylaxis. Clin Exp Allergy 46:10-20.
Scherf KA, Wieser H, Koehler P (2018) Novel approaches for enzymatic gluten degradation to create high-quality gluten-free products. Food Res Int 110:62-72.
Schilling K, Körner A, Sehmisch S, Kreusch A, Kleint R, Benedix Y, Schlabrakowski A, Wiederanders B (2009) Selectivity of propeptide-enzyme interaction in cathepsin L-like cysteine proteases. Biol Chem 390:167-174.
Schuppan D, Junker Y, Barisani D (2009) Celiac disease: from pathogenesis to novel therapies. Gastroenterology 137:1912-1933.
Schuppan D, Pickert G, Ashfaq-Khan M, Zevallos V (2015) Non-celiac wheat sensitivity: Differential diagnosis, triggers and implications. Best Pract Res Clin Gastroenterol 29:469-476.
Seilmeier W, Belitz H-D, Wieser H (1991) Separation and quantitative determination of high-molecular-weight subunits of glutenin from different wheat varieties and genetic variants of the variety Sicco. Z Lebensm Unters Forsch 192:124-129.
Shan L, Molberg Ø, Parrot I, Hausch F, Filiz F, Gray GM, Sollid LM, Khosla C (2002) Structural basis for gluten intolerance in celiac sprue. Science 297:2275–2279.
Shan L, Qiao SW, Arentz-Hansen H, Molberg Ø, Gray GM, Sollid LM, Khosla C (2005) Identification and analysis of multivalent proteolytically resistant peptides from gluten: Implications for celiac sprue. J Proteome Res 4:1732–1741.
Shewry PR (2018) Do ancient types of wheat have health benefits compared with modern bread wheat? J Cereal Sci 79:469-476.
Shewry PR, D’Ovidio R, Lafiandra D, Jenkins JA, Mills ENC, Bekes F (2009) Wheat grain proteins. In: Khan K, Shewry PR (eds)Wheat Chemistry and technology. AACC International Inc., St. Paul, MN, pp. 223-298.
Shewry PR, Pellny TK, Lovegrove A (2016) Is modern wheat bad for health? Nat Plant 2:1-3.
Shewry PR, Tatham AS (2016) Improving wheat to remove coeliac epitopes but retain functionality. J Cereal Sci 67:12-21.
Shewry PR, Tatham AS, Forde J, Kreis M, Miflin BJ (1986) The classification and nomenclature of wheat gluten proteins: A reassessment. J Cereal Sci 4:97-106.
Shewry PR, Tatham AS, Halford NG (1999) The prolamins of the Triticeae. In: Shewry PR, Casey R (eds) Seed Proteins, Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 37–84.
Sicherer SH (2001) Clinical implications of cross-reactive food allergens. J Allergy Clin Immunol 108:881-890.
Siegel M, Bethune MT, Gass J, Ehren J, Xia J, Johannsen A, Stuge TB, Gray GM, Lee PP, Khosla C (2006) Rational design of combination enzyme therapy for coeliac sprue. Chem Biol 13:649–658.
Siegel M, Garber ME, Spencer AG, Botwick W, Kumar P, Williams RN, Kozuka K, Shreeniwas R, Pratha V, Adelman DC (2012) Safety, tolerability, and activity of ALV003: results from two phase I single, escalating-dose clinical trials. Dig Dis Sci 57:440-450.
Skodje GI, Sarna VK, Minelle IH, Rolfsen KL, Muir JG, Gibson PR, Veierød MB, Henriksen C, Lundin KEA (2018) Fructan, rather than gluten, induces symptoms in patients with self-reported non-celiac gluten sensitivity. Gastroenterology 154:529-539.
Skylas DJ, Mackintosh JA, Cordwell SJ, Basseal DJ, Walsh BJ, Harry J, Blumenthal C, Copeland L, Wrigley CW, Rathmell W (2000) Proteome approach to the characterisation of protein composition in the developing and mature wheat-grain endosperm. J Cereal Sci 32:169–188.
Sofi F, Ghiselli L, Cesari F, Gori AM, Mannini L, Casini A, Vazzana C, Vecchio V, Gensini GF, Abbate R, Benedettelli S (2010) Effects of short-term consumption of bread obtained by an old Italian grain variety on lipid, inflammatory, and haemorheological variables: an intervention study. J Med Food 13: 1–6.
Sofi F, Whittaker A, Gori AM, Cesari F, Surrenti E, Abbate R, Gensini GF, Benedettelli S, Casini A (2014) Effect of Triticum turgidum subsp. turanicum wheat on irritable bowel syndrome: a double-blinded randomised dietary intervention trial. Br J Nutr 111:1992–1999.
Sollid LM, Khosla C (2011) Novel therapies for coeliac disease. J Int Med 269:604-613.
Sollid LM, Qiao SW, Anderson RP, Gianfrani C, Koning F (2012). Nomenclature and listing of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics 64:455-460.
Sonti R, Green PH (2012) Celiac disease: obesity in celiac disease. Nat Rev Gastroenterol Hepatol 9:247-248.
Spaenij-Dekking L1, Kooy-Winkelaar Y, van Veelen P, Drijfhout JW, Jonker H, van Soest L, Smulders MJ, Bosch D, Gilissen LJ, Koning F (2005) Natural variation in toxicity of wheat: potential for selection of nontoxic varieties for celiac disease patients. Gastroenterology 129:797–806.
Stein J, Schuppan D (2014) Coeliac disease-new pathophysiological findings and their implications for therapy. Viszeralmedizin 30:156-165.
Stember RH (2006) Buckwheat allergy. Allergy Asthma Proc 27:393-395.
Stepniak D, Spaenij-Dekking L, Mitea C, Moester M, de Ru A, Baak-Pablo R, van Veelen P, Edens L, Koning F (2006) Highly efficient gluten degradation with a newly identified prolyl endoprotease: Implications for celiac disease. Am J Physiol Gastrointest Liver Physiol 291:G621–G629.
Šuligoj T, Gregorini A, Colomba M, Ellis HJ, Ciclitira PJ (2013) Evaluation of the safety of ancient strains of wheat in coeliac disease reveals heterogeneous small intestinal T cell responses suggestive of coeliac toxicity. Clin Nutr 32:1043-1049.
Tallberg A (1981a) Protein and lysine content in high-lysine double-recessives of barley. I. Combinations between mutant 1508 and a Hiproly back-cross. Hereditas 94:253-260.
Tallberg A (1981b) Protein and lysine content in high-lysine double-recessives of barley. II. Combinations between mutant 7 and a Hiproly back-cross. Hereditas 94:261-268.
Tallberg A (1982) Characterization of high-lysine barley genotypes. Hereditas 96:229-245
Tatham AS, Shewry PR (2008) Allergens to wheat and related cereals. Clin Exp Allergy 38:1712-1726.
Theethira TG, Dennis M, Leffler DA (2014) Nutritional consequences of celiac disease and the gluten-free diet. Expert Rev Gastroenterol Hepatol 8:123-129.
Thompson T, Lee AR, Grace T (2010) Gluten contamination of grains, seeds, and flours in the United States: a pilot study. J Am Diet Assoc 110:937-940.
Trcka J, Schäd SG, Scheurer S, Conti A, Vieths S, Gross G, Trautmann A (2012) Rice-induced anaphylaxis: IgE-mediated allergy against a 56-kDa glycoprotein. Int Arch Allergy Immunol 158:9-17.
Turner AS, Bradburne RP, Fish L, Snape JW (2004) New quantitative trait loci influencing grain texture and protein content in bread wheat. J Cereal Sci 40:51-60.
Tye-Din JA, Stewart JA, Dromey JA, Beissbarth T, van Heel DA, Tatham A, Henderson K, Mannering SI, Gianfrani C, Jewell DP, Hill AV, McCluskey J, Rossjohn J, Anderson RP (2010) Comprehensive, quantitative mapping of T cell epitopes in gluten in celiac disease. Sci Transl Med 2:41–51.
Uhde M, Ajamian M, Caio G, De Giorgio R, Indart A, Green PH, Verna EC, Volta U, Alaedini A (2016) Intestinal cell damage and systemic immune activation in individuals reporting sensitivity to wheat in the absence of coeliac disease. Gut 65:1930-1937.
van den Broeck H, Hongbing C, Lacaze X, Dusautoir J-C, Gilissen L, Smulders M, van der Meer I (2010a) In search of tetraploid wheat accessions reduced in celiac disease-related gluten epitopes. Mol Biosyst 6:2206-2213.
van den Broeck HC, de Jong HC, Salentijn EMJ, Dekking L, Bosch D, Hamer RJ, Gilissen LJWJ, van der Meer IM, Smulders MJM (2010b) Presence of celiac disease epitopes in modern and old hexaploid wheat varieties: wheat breeding may have contributed to increased prevalence of celiac disease. Theor Appl Genet 121:1527-1539.
van den Broeck HC, Gilissen LJWJ, Smulders MJM., van der Meer IM, Hamer RJ (2011) Dough quality of bread wheat lacking alpha-gliadins with celiac disease epitopes and addition of celiac-safe avenins to improve dough quality. J Cereal Sci 53:206–216.
van den Broeck HC, van Herpen TW, Schuit C, Salentijn EM, Dekking L, Bosch D, Hamer RJ, Smulders MJ, Gilissen LJ, van der Meer IM (2009) Removing celiac disease-related gluten proteins from bread wheat while retaining technological properties: a study with Chinese Spring deletion lines. BMC Plant Biol 9:41.
Van Eckert R, Pfannhauser W, Riedl O (1992) Vienna Food Research Institute, Vienna, Austria. Contribution to quality assessment during production of gluten-free food. Ernährung/Nutrition 16:511-512.
Vazquez-Roque MI, Camilleri M, Smyrk T, Murray JA, Marietta E, O’Neill J, Carlson P, Lamsam J, Janzow D, Eckert D, Burton D, Zinsmeister AR (2013) A controlled trial of gluten-free diet in patients with irritable bowel syndrome-diarrhea: effects on bowel frequency and intestinal function. Gastroenterology 144:903-911.
Vincentini O, Borrelli O, Silano M, Gazza L, Pogna N, Luchetti R, De Vincenzi M (2009) T-cell response to different cultivars of farro wheat, Triticum turgidum ssp. dicoccum, in coeliac disease patients. Clin Nutr 28:272–277.
Vincentini O, Maialetti F, Gazza L, Silano M, Dessi M, De Vincenzi M, Pogna NE (2007) Environmental factors of celiac disease: Cytotoxicity of hulled wheat species Triticum monococcum, T. turgidum ssp. dicoccum and T. aestivum ssp. spelta. J Gastroenterol Hepatol 22:1816–1822.
Vivas S, Ruiz de Morales JM, Ramos F, Suárez-Vilela D (2006) Alemtuzumab for refractory celiac disease in a patient at risk for enteropathy-associated T-cell lymphoma. N Engl J Med 354:2514–2515.
Volta U, Caio G, Tovoli F, De Giorgio R (2013) Non-celiac gluten sensitivity: questions still to be answered despite increasing awareness. Cell Mol Immunol 10:383–392.
Waga J, Skoczowski A (2014) Development and characteristics of ω-gliadin-free wheat genotypes. Euphytica 195:105–116.
Waldmann TA, Conlon KC, Stewart DM, Worthy TA, Janik JE, Fleisher TA, Albert PS, Figg WD, Spencer SD, Raffeld M, Decker JR, Goldman CK, Bryant BR, Petrus MN, Creekmore SP, Morris JC (2013) Phase 1 trial of IL-15 trans presentation blockade using humanized Mikbeta1 mAb in patients with T-cell large granular lymphocytic leukemia. Blood 121:476-484.
Walters MJ, Wang Y, Lai N, Baumgart T, Zhao BN, Dairaghi DJ, Bekker P, Ertl LS, Penfold ME, Jaen JC, Keshav S, Wendt E, Pennell A, Ungashe S, Wei Z, Wright JJ, Schall TJ (2010) Characterization of CCX282-B, an orally bioavailable antagonist of the CCR9 chemokine receptor, for treatment of inflammatory bowel disease. J Pharmacol Exp Ther 335:61–69.
Watts RE, Siegel M, Khosla C (2006) Structure-activity relationship analysis of the selective inhibition of transglutaminase 2 by dihydroisoxazoles. J Med Chem 49:7493–7501.
Wen S, Wen N, Pang J, Langen G, Brew-Appiah RAT, Mejias JH, Osorio C, Yang M, Gemini R, Moehs CP, Zemetra RS, Kogel K-H, Liu B, Wang X, von Wettstein D, Rustgi S (2012) Structural genes of wheat and barley 5-methylcytosine DNA glycosylases and their potential applications for human health. Proc Natl Acad Sci USA 109:20543-20548.
West K (2009) CP-690550, a JAK3 inhibitor as an immunosuppressant for the treatment of rheumatoid arthritis, transplant rejection, psoriasis and other immune-mediated disorders. Curr Opin Investig Drugs 10:491–504.
Wieser H, Koehler P, Folck A, Becker D (2006) Characterization of wheat with strongly reduced α-gliadin content. In: 9th Gluten Workshop, pp. 13-16.
Wolf C, Siegel JB, Tinberg C, Camarca A, Gianfrani C, Paski S, Guan R, Montelione G, Baker D, Pultz IS (2015) Engineering of Kuma030: A Gliadin Peptidase That Rapidly Degrades Immunogenic Gliadin Peptides in Gastric Conditions. J Am Chem Soc 137:13106-13113.
Wrigley CW, Bekes F, Bushuk W (2006) Gluten: A balance of gliadin and glutenin. In Wrigley CW, Bekes F, Bushuk W (eds) Gliadin and glutenin: The unique balance of wheat quality. Am Assoc Cereal Chem, St. Paul, MN, pp. 1-28
Wrigley CW, Bietz JA (1988) Proteins and Amino Acids, In: Pomeranz Y (ed) Wheat Chemistry and Technology, Am Assoc Cereal Chem, St. Paul, MN, pp. 159-275.
Xia J, Bergseng E, Fleckenstein B, Siegel M, Kim CY, Khosla C, Sollid LM (2007) Cyclic and dimeric gluten peptide analogues inhibiting DQ2-mediated antigen presentation in coeliac disease. Bioorg Med Chem 15:6565–6573.
Yokoyama S, Perera PY, Waldmann TA, Hiroi T, Perera LP (2013) Tofacitinib, a janus kinase inhibitor demonstrates efficacy in an IL-15 transgenic mouse model that recapitulates pathologic manifestations of celiac disease. J Clin Immunol 33:586-594.
Yokoyama S, Watanabe N, Sato N, Perera PY, Filkoski L, Tanaka T, Miyasaka M, Waldmann TA, Hiroi T, Perera LP (2009) Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes. Proc Natl Acad Sci USA 106:15849-15854.
Zevallos VF, Ellis HJ, Suligoj T, Herencia LI, Ciclitira PJ (2012) Variable activation of immune response by quinoa (Chenopodium quinoa Willd.) prolamins in celiac disease. Am J Clin Nutr 96:337-344.
Zevallos VF, Herencia LI, Chang F, Donnelly S, Ellis HJ, Ciclitira PJ (2014) Gastrointestinal Effects of Eating Quinoa (Chenopodium quinoa Willd.) in Celiac Patients. Am J Gastroenterol 109:270-278.
Zevallos VF, Raker V, Tenzer S, Jimenez-Calvente C, Ashfaq-Khan M, Rüssel N, Pickert G, Schild H, Steinbrink K, Schuppan D (2017) Nutritional wheat amylase-trypsin inhibitors promote intestinal inflammation via activation of myeloid cells. Gastroenterology 152:1100-1113.
Zevallos VF, Raker VK, Maxeiner J, Scholtes P, Steinbrink K, Schuppan D (2018) Dietary wheat amylase trypsin inhibitors exacerbate murine allergic airway inflammation. Eur J Nutr https://doi.org/10.1007/s00394-018-1681-6.
Zhao J, de Vera J, Narushima S, Beck EX, Palencia S, Shinkawa P, Kim KA, Liu Y, Levy MD, Berg DJ, Abo A, Funk WD (2007) R-spondin1, a novel intestinotrophic ameliorates experimental colitis in mice. Gastroentrology 132:1331–1343.
Zuidmeer L, Goldhahn K, Rona RJ, Gislason D, Madsen C, Summers C, Sodergren E, Dahlstrom J, Lindner T, Sigurdardottir ST, McBride D, Keil T (2008) The prevalence of plant food allergies: a systematic review. J Allergy Clin Immunol 121:1210-1218.
Acknowledgements
Rothamsted Research receives grant-aided support from the Biotechnology and Biological Sciences Research Council (BBSRC) of the UK and the work forms part of the Designing Future Wheat strategic programme (BBS/E/C/000I0250). Financial support by NIH grants 1R01 GM080749-01A2, Life Sciences Discovery Fund Grant 3143956-01 and Clemson Faculty Succeeds Grant 15-202-EQUIP-5701-430-1502211 to SR is also gratefully acknowledged. Maastricht University NUTRIM receives grants from the Dutch Topsector AgriFood (TKI 1601P01) and from public private partnerships for the ‘Well on Wheat?’ research consortium addressing the health aspects of wheat consumption and wheat and gluten avoidance (http://www.um-eatwell.nl/wow/).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

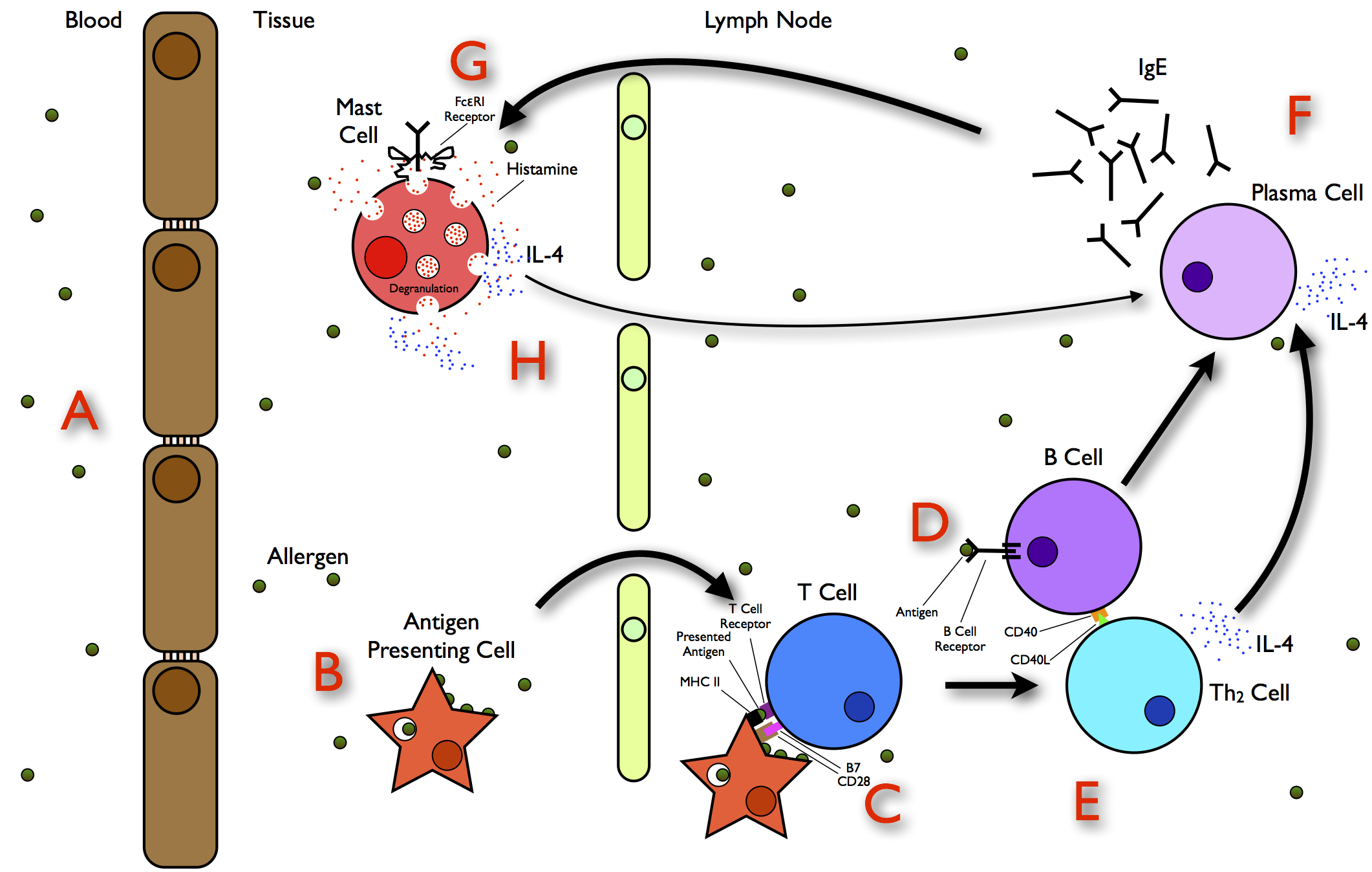{kind=link}
Cite this chapter
Rustgi, S., Shewry, P., Brouns, F. (2020). Health Hazards Associated with Wheat and Gluten Consumption in Susceptible Individuals and Status of Research on Dietary Therapies. In: Igrejas, G., Ikeda, T., Guzmán, C. (eds) Wheat Quality For Improving Processing And Human Health. Springer, Cham. https://doi.org/10.1007/978-3-030-34163-3_20
Download citation
DOI: https://doi.org/10.1007/978-3-030-34163-3_20
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-34162-6
Online ISBN: 978-3-030-34163-3
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)