
Abstract
Disturbances to forest ecosystems alter nutrient availability and loss, with potassium and nitrate typically exhibiting the most mobile behavior. However, the sources and ecosystem processing of potassium and nitrate differ in important ways that shape their response to large-scale forest disturbance. In this chapter we compare the mobilization and subsequent recovery of potassium and nitrate following natural and experimental disturbances at the Hubbard Brook Experimental Forest, New Hampshire. The response of soil and stream water chemistry was evaluated for both natural (i.e., ice storm) and anthropogenic (devegetation, whole-tree harvest, strip cut) experiments. The greatest losses of nutrients were evident in a devegetation experiment, followed by a whole-tree harvest, with similar losses occurring from an ice storm and a strip cut. There was remarkable consistency in the stoichiometric behavior in solutions across this wide range of disturbance types, with an approximately 0.1 molar ratio of potassium to nitrate. After large-scale forest harvest, nitrate concentrations increased and subsequently recovered more rapidly than potassium. We highlight the role of canopy and fresh plant litter leaching of potassium, root nitrogen uptake, microbial processing of nitrogen, and retention of potassium by cation exchange in controlling variation in the magnitude, timing, and duration of nutrient losses.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
1 Introduction
The Hubbard Brook Ecosystem Study (HBES) was initiated in the early 1960s with the goal of improving basic understanding of the ecosystem dynamics of forested landscapes . Bormann and Likens (1967) applied the small watershed budget approach, originally formulated to quantify the hydrologic cycle (Hewlett and Hibbert 1967) to biogeochemical cycles (Likens et al. 1967). By measuring the chemistry of hydrologic inputs (precipitation) and outputs (stream discharge) in a catchment with watertight bedrock, a mass balance for elements can be quantified precisely. After several years of pre-treatment monitoring , Likens et al. (1970) applied this approach to evaluate ecosystem responses to the deforestation of a small watershed (W2; 15.6 ha) at the Hubbard Brook Experimental Forest (HBEF) . Since that beginning continued monitoring and experimental treatments in the Hubbard Brook watersheds have further elaborated the mechanisms underlying long-term changes in the biogeochemistry of northern hardwood ecosystems and responses to natural and anthropogenic disturbances (Likens 2013). The present chapter is a further contribution to this long legacy of discovery in biogeochemistry from the HBES.
In forest ecosystems, disturbances can occur as relatively discrete events that result in a drastic change in environmental conditions and resource availability, usually associated with mortality of overstory trees . In northeastern forests the natural disturbance cycle most commonly reflects the death of one or a few trees in small patches caused by windthrow (Bormann and Likens 1979), but occasionally extensive disturbances occur as a result of severe windstorms (microbursts, tornadoes, and hurricanes), wildfire, ice storms , insects, and pathogen irruptions. In the HBEF three severe storms have impacted the experimental watersheds and adjacent forests during the past 80 years: the hurricane of 1938 (Merrens and Peart 1992), a severe ice storm in 1998 (Rhoads et al. 2002), and a large microburst windstorm in 2013 that knocked down about 25 ha of forest immediately west of the gauged watersheds (Battles et al. 2017). Biogeochemical responses to such disturbance events are distinct from those creating small canopy gaps (Houlton et al. 2003). Thus, the northeastern forest landscape was originally characterized as a shifting mosaic of forest stands of different sizes, shapes, and ages resulting from this combination of disturbances (Bormann and Likens 1979). However, with the exception of a few virgin tracts (Runkle 1982), the modern northern forest landscape has been pervasively changed by extensive forest harvest, beginning in the mid-nineteenth century. Although forestry practices have varied through time and space, forest harvests then and now comprise large-scale disturbances that in many ways are comparable to the natural disturbances of northern forests, especially from the mortality of overstory trees. Thus, a better understanding of both natural forest ecosystem dynamics and those of the fully managed forest can be gleaned by studying watershed-scale responses to experimental treatments.
The early watershed-scale experiments at Hubbard Brook, together with other seminal studies of the biogeochemistry of forest soils (e.g., McColl and Cole 1968; Johnson and Cole 1980), have provided key insights into the mechanisms whereby forest disturbances could result in increased leaching of nutrients from soil and transport to surface water. Mortality of canopy trees greatly decreases root uptake of mineral nutrients , while microbial mineralization of soil organic matter and plant detritus continues to generate soluble cations and anions. Cations (K+, Ca2+, Mg2+) can be retained by the soil cation exchange complex, whereas anions are typically more mobile and more readily transported through soil. Thus, the supply of mobile anions to soil solution can influence leaching of base cations (Johnson and Cole 1980). The relative mobility of nutrient cations differs, reflecting the lytropic or Hofmeister series, NH4 +~K+>Mg2+>Ca2+, so that cation exchange reactions can attenuate leaching responses. Moreover, decreases in pH associated with nitrification can decrease the mobility of the major anion, SO4 2−, due to enhanced retention associated with the protonation of soil surfaces (Nodvin et al. 1986a). Finally, the supply and mobility of NO3 − is altered by microbial processes, including N mineralization, immobilization, nitrification, and denitrification (Vitousek et al. 1997), and by atmospheric inputs (Galloway et al. 2004).
Largely because of their importance as nutrients , ionic properties, and high solubility, the most mobile solutes in forest soils following large-scale disturbance are usually K+ and NO3 −. Both are essential plant nutrients for which demand is greatly reduced by canopy tree mortality, and leaching of these nutrients might be expected to increase roughly in parallel after disturbance and to return to pre-disturbance values as vegetation recovers. In fact, Likens et al. (1978) observed that K+ and NO3 − exhibited by far the largest response of all nutrients measured to experimental deforestation of W2 at the HBEF, and their recovery roughly paralleled vegetation recovery. However, the sources and mobility of K+ and NO3 − differ in several important ways. The high mobility of K+ results in part from its nonstructural form in plant tissues, so that it is rapidly leached from detritus following the death of plant tissues (Attiwill 1968). The mobility of K+ in soil is consistent with its place in the lyotropic series, but unlike NO3 −, K+ is retained by cation exchange sites. Moreover, K+ input is primarily associated with weathering of primary minerals. In contrast, NO3 − is not present in significant amounts in plant tissues or minerals and is generated from decay of detritus only following mineralization of organic matter and subsequent microbial nitrification. Moreover, NO3 − is supplied to northeastern forests from the atmosphere over recent decades mostly by atmospheric deposition as a result of elevated emissions of nitrogen oxides (Aber et al. 1989; Driscoll et al. 2003). Finally, NO3 − can be immobilized by microbial uptake to a much greater extent than for K+, because of the relatively high requirement for N by microbial biomass . In sum, these contrasts between K+ and NO3 − likely shape differences in their behavior in response to large-scale forest disturbances .
The goal of this chapter is to contribute to a better understanding of the mechanisms of forest biogeochemical response to large-scale disturbances. Our objective is to compare and contrast the mobilization and subsequent recovery of K+ and NO3 − following natural and experimental disturbances of the Hubbard Brook watersheds. We hypothesize that the difference in sources and mobility of K+ and NO3 − should result in nonparallel behavior of these solutes. Moreover, long-term studies at the HBEF have documented unexpected and unexplained patterns of NO3 − loss from the experimental watersheds; in particular, decreasing NO3 − losses have occurred since the 1970s, despite maturing forest which assimilates less nitrogen and relatively small changes in atmospheric nitrogen inputs during 1970s–1990s (Yanai et al. 2013; Groffman et al. 2018). We hoped that coincident patterns of K+ flux in soils might provide new insights into the mechanism underlying these patterns.
2 Methods and Approaches
The HBEF is a 3160 ha forest experiment station located in the White Mountain National Forest in central New Hampshire (43° 56′N, 71°45′W). The climate is cool temperate, humid continental, with mean July and January temperatures of 19 and −9 °C, respectively (at 450 m elevation). Annual precipitation averages about 1400 mm and is distributed nearly evenly throughout the year. Approximately 30% of annual precipitation occurs as snow (Federer et al. 1989).
The HBEF encompasses the entire catchment of the 4th order stream Hubbard Brook. The bedrock geology underlying the gauged watersheds that are the subject of this chapter is in the Littleton Formation, a metamorphic schist from the Devonian period. Detailed studies indicated that this bedrock is relatively watertight so that deep seepage of water is minimal and essentially all outflow is captured by V-notch weirs at the base of the six small gauged watersheds (W1–W6). The watersheds are situated on a moderately steep, south-facing slope and are drained by first- and second-order streams. Soils are derived from glacial till of varying thickness and are mostly well-drained, acid Spodosols (Bailey et al. 2014). Recent studies have emphasized spatial variation in the soils within the HBEF, including some lateral development near bedrock outcrops and the stream channel and more vertical development away from these hydrologic constraints (Bailey et al. 2014; Gannon et al. 2014). On ridgetops and upper slopes soils are thin and often bedrock limited whereas deeper glacial till is found on lower slopes.
The forest vegetation of the HBEF is mostly northern hardwood forest dominated by sugar maple (Acer saccharum Marsh.), American beech (Fagus grandifolia Ehrh.), and yellow birch (Betula alleghaniensis Britt.) with increasing amounts of the conifers red spruce (Picea rubens Sarg.) and balsam fir (Abies balsamea (L.) Mill) at the highest elevations. Forest on the reference watersheds (W1, W6) is mostly about 100 years old, recovering from intensive harvest in the early twentieth century, as well as from some earlier cutting (1870s–1880s) and damage by the 1938 hurricane.
Among the six gauged watersheds on the south-facing slope of the HBEF, two are reserved as references (W3, hydrologic reference, and W6, biogeochemical reference). The earliest experiment at the HBEF was the devegetation of W2 in which all trees were cut and left in place in 1965 and regrowth was prevented for three years using herbicides (Likens et al. 1970). W4 was logged by clear-cutting of 25-m-wide, horizontally oriented strips. One-third of the strips were logged in each of three years (1970, 1972, and 1974) and a buffer strip was retained along the stream channel in the lower watershed; this experiment was successful in minimizing nutrient outflow in stream water during and after logging (Hornbeck et al. 1986). In winter 1983–1984 W5 was clear-cut as a whole-tree harvest, in which all aboveground biomass of trees >2.5 cm DBH was removed from the watershed (Dahlgren and Driscoll 1994).
A “natural” large-scale disturbance experiment was provided across all the gauged watershed by the severe 1998 ice storm (Irland 1998). The upper two-thirds of the watersheds were severely damaged by the 1.2–1.4 cm of ice that accumulated on the tree crowns. Detailed measurements of damage indicated 28–30% crown damage watershed wide in the mature forests on W1 and W6; the most severe damage was measured on the 24–28-year-old W4, whereas minimal canopy damage occurred in the 14-year-old W5 (Rhoads et al. 2002).
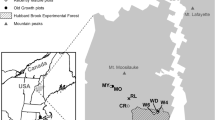
Stream discharge has been measured continuously on the gauged watersheds since the early 1960s using V-notch weirs. Precipitation is measured on a network of rain gauges, and the annual water balance is calculated on a water year basis from June 1 to May 31 (Likens 2013). Stream water samples for chemical analysis have been collected weekly since 1963 immediately above the weir in each watershed. In addition, beginning in the 1980s stream water samples for chemical analysis have been collected at a series of locations arranged longitudinally upstream from the weir in W1, W5, and W6 (Fuss et al. 2015). Soil solution samples have been collected monthly from three soil horizons (Oa, Bh, Bs) at three elevations, in the lower and higher elevations of the hardwood zone and within the spruce-fir zone, of W1, W5, and a reference watershed immediately west of W6 using zero-tension lysimeters (Fuss et al. 2015; Fig. 22.1).

Location of the experimental watersheds at the Hubbard Brook Experimental Forest , NH. Location of rain gauges, stream weirs, lysimeter, and stream chemistry sampling sites is shown
In this chapter we report on concentrations of K+ and NO3 − in the water samples described above. The analytical chemistry approaches are detailed in Buso et al. (2000). For some purposes in this chapter, we report volume-weighted mean concentrations. Appling a linear interpolation, we calculated daily concentration for days between sampling dates (Buso et al. 2000). The daily flux was estimated by multiplying the daily concentration by the daily streamflow. We calculated period (monthly and annual) volume-weighted mean concentrations by dividing the summation of daily fluxes to the summation of streamflows for the corresponding period. In describing overall disturbance effects on solute concentrations, we calculated average values for the interval during which concentrations were elevated above pre-disturbance background levels (Table 22.1). To test if post-disturbance concentrations of K+ and NO3 − in stream and soil solutions are significantly different from pre-disturbance concentrations, we used a two-sample t-test. In the t-test, the monthly observations during the elevated concentration interval (post-disturbance interval) were compared to the monthly observations during the pre-disturbance interval. The length of pre-disturbance period was determined based on availability of concentration measurements and to ensure a period that minimizes the seasonality in measurements. We also investigated long-term trends in concentrations of K+ and NO3 − in stream and soil solutions using the nonparametric seasonal Kendall’s tau test (Hirsch et al. 1982). Seasonal trend analysis was conducted on monthly data. The slope of trends was estimated using the Sen Estimator (Sen 1968).
3 Results
3.1 Reference Watershed: Patterns of NO3 − and K+
Concentrations of NO3 − in the stream draining the biogeochemical reference watershed (W6) have significantly declined since the 1960s and 1970s (−0.16 μmol L−1 year−1; Fig. 22.2; Yanai et al. 2013). This decline has preceded a marked decrease in atmospheric NO3 − deposition since the early 2000s (−1 μmol L−1 year−1) which was the result of controls on atmospheric nitrogen oxide emissions. The mechanisms driving this unexpected tightening of the nitrogen cycle, a so-called nitrogen oligotrophication (Groffman et al. 2018), remain unclear although the possible role of increased carbon supply associated with carbon dioxide fertilization to the forest, increases in the duration of the growing season, or increased mycorrhiza activity due to recovery from acid deposition has been posited. During most years since the early 1990s, stream NO3 − concentrations have been at or near analytical detection limits. However, occasional large excursions from this strong N retention have been detected in 1981, 1991, 2000–2001, and 2015 (Fig. 22.2). The episode in 2000–2001 was clearly tied to canopy damage from the severe ice storm in 1998 (Rhoads et al. 2002) and consequent decrease in tree N uptake (Houlton et al. 2003). Earlier NO3 − loss episodes were tentatively attributed to effects of severe soil frost (Mitchell et al. 1996) and consequent damage to root systems depressing plant uptake (Tierney et al. 2001; Cleavitt et al. 2008). However, neither canopy damage nor severe soil frost coincided with the 2015 peak. It is also notable that the latter peak spanned three years, 2014–2016, in the adjacent W1 (Fig. 22.2) which was treated with a calcium silicate (wollastonite) to increase pH and soil exchangeable Ca2+ to levels that prevailed prior to the peak acid deposition era (1950–2000; Peters et al. 2004).

Long-term annual volume-weighted concentrations of NO3 − and K+ in stream water in W6 (a) and W1 (b) at the HBEF the occurrence of the soil freezing and ice storm events are shown by vertical lines. Watershed 6 is the biogeochemical reference watershed. Watershed 1 was treated with wollastonite (CaSiO3) in 1999
Concentrations of K+ in stream water in the reference watershed (W6) have fluctuated to a similar degree over the years; however, the significant long-term temporal decline (−0.03 μmol L−1 year−1) is less marked than for NO3 − (−0.16 μmol L−1 year−1). Some indications of the coupling between K+ and NO3 − losses are apparent from coincident peaks in annual volume-weighted mean concentrations of both solutes in a few years (1971, 1981, 2000–2001 and 2015; Fig. 22.2), but in other years high fluxes are decoupled, so that linear correlations between these solutes are moderate (annual volume-weighted concentrations, slope 0.18 mol K+ (mol NO3 −)−1; r2 = 0.31). Thus, while some common mechanism (e.g., reduced plant uptake) may contribute to leaching losses of both solutes, clearly other driving factors are involved.
The latter conclusion is reinforced by considering the seasonal patterns of stream NO3 − and K+ concentrations in the reference watershed (W6; Fig. 22.3). The long-term weekly average NO3 − concentrations exhibit a clear increase beginning in early November and peaking in April prior to leaf out, followed by a marked decline to trace levels in early summer, presumably reflecting primarily high root uptake to supply canopy foliage expansion (Fig. 22.3). In contrast, K+ concentrations increase and peak several weeks earlier than NO3 − in fall, coinciding with leaching from senesced foliage, perhaps especially litter falling in and near stream channels. The decline in K+ concentrations associated with leaf emergence in spring is modest and much delayed in comparison with that of NO3 −, possibly due to interactions of K+ with the soil exchanger (Fig. 22.3). Finally, the magnitude of the difference in stream K+ concentration between the growing and dormant season is much less (ca. twofold) in comparison with that of NO3 − (ca. tenfold).

Seasonal variations in NO3 − and K+ concentrations in stream water in W6: (a) long-term mean values (1964–2016) and (b) for 2015. The error bars indicate standard errors. In plot (a), because of limited observations and substantial variability in NO3 − concentrations in the first week of observations, the related data point was not shown
During the unexplained flush of NO3 − and K+ from W6 in 2015 (Fig. 22.2a), the high NO3 − flux was confined to the late winter peak as concentrations remained at detection limits (0.3 μmol L−1; Buso et al. 2000) throughout the growing season and into early winter (Fig. 22.3). The increased K+ concentrations in the W6 stream reflected primarily higher concentrations in early summer and early autumn and was clearly decoupled from the NO3 − flush. The seasonal pattern of NO3 − loss in 2015 is similar to that following the 1980 soil freezing event (Mitchell et al. 1996) although in 1981 increases in stream NO3 − were evident in late fall (starting in November).
Concentrations of NO3 − and K+ in soil solutions (collected using zero-tension lysimeters; Fuss et al. 2015) decline in parallel from surface soil to deep soil horizons, with highest levels draining the forest floor organic horizons (Fig. 22.4). Presumably these depth patterns reflect the effects of differences between the supply of K+ (leaching from detritus, mineral weathering) and NO3 − (deposition, nitrification) and biological uptake (root, microbial) and, for K+, exchange on soil surfaces. In upper soil horizons (Oa and Bh) concentrations of K+ and NO3 − in soil solutions decrease with increasing elevations in the watershed (Fig. 22.4). In the case of K+, this may be due to the shallow mineral soil resulting in low K+ supply. Dittman et al. (2007) suggested that low NO3 − concentrations in the high elevation spruce-fir zone were due to N immobilization associated with high litter and soil C/N. One notable difference between the two solutes is that NO3 − concentrations in streams are consistently lower than in soil solutions (including subsoil), whereas K+ concentration is higher in streams than subsoil. The latter pattern is probably influenced by leaching from mineral soil or glacial till or from fresh detritus in streams, although shallow and rapid flow paths that short-circuit soil micropores also could contribute as detailed in the discussion. Decreases in NO3 − concentrations are likely associated with immobilization in the riparian zone between the soil and the stream channel (Dittman et al. 2007).

Long-term mean NO3 − and K+ concentrations in bulk deposition, throughfall, soil solutions by horizon and stream water for three elevation zones: spruce-fir (750 m), high elevation hardwood (730 m), and low elevation hardwood (600 m) in reference sites west of W6 (1984–2016). Throughfall concentrations are average of measurements during the growing seasons of 1989–1992 in five sites at different elevation zones (Lovett et al. 1996)
3.2 Disturbance Effects: General
The response of soil and stream water chemistry to large-scale disturbance has been evaluated in a series of experiments, both natural (i.e., 1998 ice storm ) and anthropogenic (harvesting) (W2, W4, W5). In Table 22.1 we compare and contrast the magnitude of response of NO3 − and K+ concentrations in stream water during the three-to-six year interval following the disturbances during which concentrations were elevated in comparison with pre-disturbance average values. Clearly, NO3 − is the most mobile solute in these northern hardwood forests following vegetation disturbance. Devegetation of W2 resulted in over a 50-fold increase in NO3 − concentration. The elimination of nearly all plant uptake for three years caused marked accumulation of NH4 + on soil exchange sites, the product of organic matter mineralization, and subsequent microbial nitrification. By comparison, K+ concentration increased 13-fold, partly from leaching of felled detritus, but especially by mobilization from cation exchange sites associated with acidity derived from nitrification (Van Miegroet and Cole 1984). The stoichiometric response of K+ to NO3 − associated with the W2 disturbance was 0.094 mol K+ (mol NO3 −)−1 (Table 22.1).
Under less extreme forest harvest disturbances (W4, strip cut; Hornbeck et al. 1986), NO3 − concentrations in streams increased by only two- to threefold as root uptake by both residual and recovering vegetation retained mineralized N. Similarly, K+ losses were markedly lower, only 60% higher than pre-treatment, but the stoichiometric response of K+ to NO3 − associated with the strip cut was 0.090 mol K+ (mol NO3 −)−1, similar to that of W2. The whole-tree harvest of W5 resulted in much higher concentrations of both NO3 − (21-fold) and K+ (nearly 3-fold) compared to the pre-treatment concentrations and intermediate between the W2 and W4 experiments. A slightly higher ratio of K+ to NO3 − leaching in the W5 experiment (0.109 mol K+ (mol NO3 −)−1; Table 22.1) was evident compared to other treated watersheds (W2 and W4) despite the removal of aboveground biomass in W5 and hence a much lower supply of K+ from leaching of dead organic matter (Likens et al. 1994).
The response of stream NO3 − concentrations to the severe ice storm of 1998 was more modest than for the forest harvest experiments (Table 22.1); nevertheless, NO3 − loss increased significantly (p < 0.001) for about three years after the ice storm in the mature forest watersheds: W1, W3, and W6 (Table 22.1; Houlton et al. 2003). The increase in NO3 − concentration was similar across all three watersheds, 3.5- to 4-fold. Ice storm damage to the canopy in these watersheds was similar averaging 29% canopy loss with greater damage in mid to upper elevations (Rhoads et al. 2002). The response of stream NO3 − to ice damage in the 24–28-year-old W4 was greater than for the mature forests, approaching values observed in response to forest harvest (Fig. 22.5a). This higher NO3 − leaching in W4 reflects greater canopy damage in that younger forest owing to high susceptibility of the initially regrowing pin cherry (Prunus pensylvanica Sarg.) at this age (Rhoads et al. 2002). In striking contrast, no NO3 − leaching pulse was observed in the adjacent, 14-year-old W5 (Fig. 22.5b) where canopy damage was minimal, despite similar ice loading, owing to flexible stems of younger trees.

Long-term annual volume-weighted concentrations of NO3 − and K+ in stream water in (a) W4 and (b) W5 at the HBEF the occurrence of the soil freezing and ice storm events are shown by vertical lines
The response of concentrations of K+ to the ice storm disturbance was more muted than of NO3 − in W1 and W6, but the stoichiometric response was similar to the harvesting treatments (W1 0.072 mol K+ (mol NO3 −)−1; W6 0.109 mol K+ (mol NO3 −)−1; Table 22.1). However, in W4 where the NO3 − pulse was greatest, ice storm damage did not cause a significant increase in stream K+ concentrations, further evidence of mechanistic decoupling of the mobilization of these two ions.
3.3 Seasonality of Response to Disturbances
Both NO3 − and K+ responded to the whole-tree harvest of W5 with a marked increase in stream water concentrations, with a distinct peak in 1986 two years after the 1983–1984 harvest. However, the seasonal pattern of these responses was distinctly different for NO3 − and K+ (Fig. 22.6) in this peak year of leaching loss. In particular, the NO3 − concentration increase was most apparent during the dormant season, January to April, while concentrations declined dramatically in May and June, coincident with leaf expansion. In contrast, the seasonal pattern of K+ concentration was muted, as the increase persisted through the year with only a minor decline in summer. Thus, in comparison with the long-term average behavior of W6 (Fig. 22.3) and the seasonal pattern during the 2015 pulse of loss, seasonal variation in K+ during the postharvest of W5 was more muted, whereas NO3 − leaching was similar.

Seasonal variation in concentrations of NO3 − and K+ in stream water in W5 for 1986 the peak year of solute loss following the whole-tree harvest
3.4 Annual Patterns Following Disturbance
The timing and duration of the response of stream water solute concentrations to the watershed disturbances differed both between solutes and among the disturbances. In general, after forest harvest NO3 − concentrations increased more quickly than those of K+ and the subsequent decline was also more rapid for NO3 −, as K+ concentrations remained elevated for over a decade after harvest whereas NO3 − concentrations returned to baseline levels within about 3 years (Fig. 22.5b). Not surprisingly the progressive strip harvest of W4 (1970–1974) resulted in a delayed and broader peak of stream concentrations than the more discrete whole-tree harvest (1983–1984) of W5 (Fig. 22.5a). Notably, the peak in K+ concentrations in W4 occurred six years following the completion of harvest on W4 (1980) compared with two years on W5. In contrast, the timing and duration of high NO3 − leaching was more similar between W4 and W5 than for K+. Finally, following ice storm damage , the timing and duration of the NO3 − response in the reference watersheds (Fig. 22.2) was quite similar to that following the harvest of W5 (Fig. 22.5b) and the peak of K+ while smaller followed a similar rapid recovery pattern as NO3 − (Fig. 22.2).
3.5 Response of Soils and Soil Solutions to Disturbance
In parallel with stream water chemistry , soil solution NO3 − and K+ concentrations increased markedly following large-scale disturbance (e.g., whole-tree harvest on W5; Fig. 22.7). Concentrations of NO3 − in forest floor Oa horizon samples increased rapidly during the spring and summer for two years after the harvest, probably as a result of decreased root uptake and increased nitrification in the absence of mature forest vegetation. Increased K+ concentration in Oa soil solution was roughly parallel with that of NO3 − though the magnitude of increase was less. Following this early peak, concentrations of both solutes in Oa solution gradually declined during the next three years, probably in part as a result of increasing root uptake by recovering vegetation (Mou et al. 1993); for K+, a decrease in the supply provided by decaying root systems certainly also contributed (Fahey et al. 1998). Thereafter, a marked increase in Oa solution K+ concentrations was observed from years 3–8 following the harvest, reaching values similar to the initial peak immediately following harvest (Fig. 22.7). This interval was also marked by substantial season variation, with peak concentrations occurring in late winter. Notably, NO3 − concentrations in Oa solutions during years 3–8 also were elevated compared to the baseline and followed similar seasonal cycling for K+ (Fig. 22.8).

Long-term concentrations of NO3 − (a) and K+ (b) in lysimeters at Oa and Bs depths in W5

Seasonal variation in concentrations of NO3 − and K+ in Oa lysimeters for 6 years (1988–1992) in W5. The error bars indicate standard errors
Soil exchangeable K+ was measured in the three elevation zones before the whole-tree harvest on W5 (1983) and at intervals during forest recovery (1986, 1991, 1998) (Romanowicz et al. 1996). Exchangeable K+ in mineral soil horizons (Bs) increased markedly by the second summer after harvest. Thereafter, exchangeable K+ in the Bs declined significantly. Although a small increase in exchangeable K+ also was observed in the forest floor horizons in year 2 in the highest elevation zone of W5, on a watershed-wide basis this pool did not change significantly over time.
4 Discussion
Observations of the response of concentrations of NO3 − and K+, two relatively soluble nutrients in temperate forest soils, in ecosystem solutions draining northern hardwood forest watersheds following large-scale disturbances indicate considerable variation in the magnitude, timing, and duration of nutrient losses. Some of this variation can be attributed to specific mechanisms that influence the supply and solubility of NO3 − and K+ and provide insights into factors that may be responsible for long-term patterns of nutrient losses from the HBEF watersheds. In particular, we highlight the likely roles of (1) NO3 − supply from atmospheric deposition and canopy leaching of K+, (2) rapid leaching of K+ from fresh plant detritus, (3) hydrologic flow paths and events, (4) root uptake by forest vegetation, (5) key microbial processes in the N cycle, and (6) soil cation exchange reactions. We evaluate the effect of each of these factors in accounting for differences in NO3 − and K+ behavior and their linkages and conclude with a synthesis some implications for biogeochemical dynamics in northern hardwood forest ecosystems and suggestions for future research.
4.1 Atmospheric Deposition and Canopy Leaching
A significant amount of NO3 − is supplied to the HBEF in the form of atmospheric deposition which remained elevated through most of the duration of the HBES (1963~2000) until recent declines have been evident due to nitrogen oxide emission controls (Lloret and Valiela 2016); in fact, NO3 − concentrations in precipitation are similar to levels in soil solutions in the mature forest (Fig. 22.4). However, tracer studies (e.g., Kendall et al. 1995; Campbell et al. 2007) indicate that most of the NO3 − in soil solutions and streams is the product of internal N transformations involving microbial activity, rather than derived directly from precipitation NO3 − which is rapidly immobilized in soil by biotic processes (see below). Thus, the direct effect of atmospheric NO3 − on spatial and temporal patterns of NO3 − concentrations in soil and stream water is likely minimal.
The input of K+ from precipitation (long-term mean 16.65 mol ha−1 year−1; 1.14 μmol L−1; Fig. 22.4) is relatively minor compared with supply by mineral weathering (Likens et al. 1994). However, K+ is readily supplied to the soil surface in large amounts by canopy leaching, owing to its soluble form in leaf cells (Parker 1983; Fig. 22.4). Thus, seasonal variation in K+ supply to soils by canopy leaching results from leaf shedding in the deciduous canopy (see Lovett et al. 1996). In winter and spring we might expect lower K+ inputs to soils associated solely with direct precipitation. On the other hand, however, fresh leaf litter is also subject to rapid leaching of K+.
4.2 Leaching from Fresh Detritus
One of the principal mechanisms expected to cause decoupling between K+ and NO3 − concentrations in ecosystem solutions is the leaching of nutrients from fresh detritus, a large source of K+ but not NO3 −. In the reference watershed this effect is evident most clearly during and immediately after leaf litter fall in October–December: K+ flux increases to a much greater degree than NO3 − flux (Fig. 22.3).
The supply of K+ from leaching of fresh detritus differed markedly among the watershed disturbances, which should be reflected in the relative increase of K+ concentration compared to NO3 − concentration in soil and streams. In particular, K+ input in detritus followed the order W2 > W4 > W5 > ice storm . Although severe canopy damage occurred in the ice storms, tree mortality above the background rate only doubled (Battles et al. 2014); hence, mortality of fine roots, a principal source of K+ in the harvested watersheds (Fahey et al. 1988), was minimal. This factor could help to explain the small increase in K+ losses in soil solutions following the ice storm disturbance; however, K+/NO3 − stream concentration ratio was comparable to harvested watersheds (0.109 and 0.072 mol K+ (mol NO3 −)−1 for W6 and W1, respectively; Table 22.1; see Synthesis below). At the other extreme, in W2 all aboveground biomass was left in place and as expected K+ losses were particularly high in the forest 2–3 years after harvest (Likens et al. 1970), and these high K+ losses were also undoubtedly facilitated by the elevated release of NO3 − which enhanced the transport of K+ due to its role as a mobile anion; however, again the K+ released to stream water relative to NO3 − was comparable to other disturbances (0.094 mol K+ (mol NO3 −)−1).
In the case of the whole-tree harvest of W5, belowground detritus was a substantial contributor to K+ losses (Fahey et al. 1988; Likens et al. 1994). The timing of the initial spike of K+ concentration in streams was roughly coincident with that of NO3 − (Fig. 22.5b) and probably resulted from rapid mortality of fine roots during the first summer postharvest (0.109 mol K+ (mol NO3 −)−1). Thereafter, gradual mortality and leaching from the remaining dead root system could have supplied prolonged loss of K+ for 3–4 years, contributing to the decoupling of K+ and NO3 − loss in 1987–1991 (Fig 22.5b). This mechanism also might help to account for some of the unexpected resurgence of K+ leaching from surface soil horizons in W5 in 1989–1991 (Fig. 22.7), although it is unclear why this pattern exhibited such a pronounced late winter peak (Fig. 22.8).
4.3 Hydrologic Effects
During low flow periods in summer, water generally percolates through the highly porous surface soil layers that predominate in the HBEF landscape, and stream discharge is mostly derived from subsurface flow; hence, stream water concentrations of K+ and NO3 − are similar to soil solutions in subsoil Bs horizon (Fig. 22.4). However, when evapotranspiration rates are low during leafless periods, water tables can migrate upward in the soil profile, especially in certain landscape positions (Bailey et al. 2014). At these times percolating water moves laterally in upper soil horizons, short-circuiting deep flow paths. Romanowicz et al. (1996) noted that during these high flow periods in W5, K+ concentrations in stream water were similar to those in surface soil horizons, and much higher than in subsoil solutions. In contrast, in summer surface soil solution K+ concentration was elevated (by leaching of detritus) and stream water levels were low. These observations are consistent with the hypothesized changing flow path mechanism outlined above.
A role of hydrologic factors in regulating soil solutions and stream water NO3 − concentrations also seems likely. Notably, the high annual NO3 − fluxes that occurred several times since the 1970s (Fig. 22.2) were associated primarily with high spring concentrations. Some of these unexplained NO3 − flux events in the reference watersheds coincide with rain-on-snow events in late winter. These events could cause flushing of soil micropores or rapid flow of NO3 −-rich surface drainage to streams.
4.4 Plant Uptake of NO3 − and K+
Both NO3 − and K+ are macronutrients and are absorbed in large amounts by plant roots in the mature forest. Plant uptake undoubtedly contributes to declining concentrations of both solutes with increasing depth in soil in the mature forests (Fig. 22.4) and to seasonal differences between the dormant and growing season (Fig. 22.3).
Although plant demand for N exceeds that for K+, because more N is resorbed from foliage than K+ (Ryan and Bormann 1982), the difference in root uptake is smaller. Moreover, much of N uptake is in the form of NH4 + (Campbell et al. 2014); however, any decrease in root uptake of NH4 + can translate to increased nitrification and NO3 − supply to leaching water. The large-scale disturbances of the HBEF watersheds reduced plant uptake, and the degree of reduction followed the order: W2 > W5 > W4 > ice storm . This differential reduction in root uptake played a dominant role in regulating the differences among the watersheds in NO3 − and K+ leaching (W2 > W5 > W4 > ice storms; Table 22.1). This reduction in root uptake would be expected to influence leaching losses during the growing season to a greater degree than during the dormant season; hence, we would expect the response to disturbance to be greater in the growing season. However, stream water concentrations of NO3 − actually increased to a greater degree during the dormant season than the growing season in the peak flush year in W5 (1986; Fig. 22.6). Perhaps demand by the residual vegetation, the growth of new understory shrubs and herbs, and microbial biomass following whole-tree harvest were sufficient to buffer losses.
The influence of root uptake in directly regulating seasonal variations in K+ losses is less clear. In W5 soil solutions, K+ concentrations were elevated in comparison with the reference watershed in soil solutions from all depths and in stream water throughout the year (Romanowicz et al. 1996); thus, assimilation by roots apparently does not greatly reduce K+ concentrations, at least in the mobile soil water collected by the zero-tension lysimeters. However, root uptake may be derived primarily from micropore water that is underrepresented in our soil solution collections. Moreover, as noted by Romanowicz et al. (1996), roots permeate near-stream saturated zones and may thereby directly influence stream water concentrations during the growing season.
4.5 Microbial Activity
The effect of microbial activity on NO3 − dynamics is much more complex than for K+ and certainly contributes to decoupling between the fluxes of these solutes. Although microbial decomposition of plant detritus contributes to K+ leaching , this effect only slightly accelerates the leaching process. In contrast, NO3 − dynamics are influenced by microbial immobilization, microbial mineralization of organic matter leading to ammonification and subsequent nitrification, and gaseous loss of N during microbial nitrification and denitrification. Researchers at the HBEF have long puzzled about the causes of long-term decrease in NO3 − losses from the reference watershed despite continuing N deposition and maturing of the forest (thereby reducing net accumulation in biomass ; Yanai et al. 2013). Some limited evidence suggests possible decreases in soil N mineralization and so-called N oligotrophication (Groffman et al. 2018) perhaps driven by increased C inputs and consequent microbial immobilization. Isotopic tracer studies indicate rapid microbial immobilization of precipitation NO3 − in organic matter-rich surface soil horizons, even during the spring snowmelt period (Campbell et al. 2007). Another possible explanation is increasing denitrification losses, as suggested by some evidence for periodically high rates of denitrification (Morse et al. 2015) even in the mostly well-drained soils of the HBEF. Finally, a recent hypothesis (N-bank; Lovett et al. 2018) suggests a long-term shift in the mineral soil as a N sink vs source depending upon changing forest demand.
The contribution of these microbially mediated processes to the decoupling between NO3 − and K+ leaching following large-scale disturbance is not clear. Certainly the relatively high NO3 − and K+ leaching from W2 immediately after forest cutting and herbicide treatment resulted from the large supply of NO3 − via nitrification owing to elimination of NH4 + uptake by plants and its accumulation in soils. In contrast, measurements of in situ nitrification following the ice storm disturbance did not detect a significant increase, and the direct role of reduced root uptake of NO3 − was suggested (Houlton et al. 2003).
Following forest harvest, changes in conditions would seem to favor increased denitrification, including increased soil wetness (due to lower evapotranspiration), increased supply of NO3 −, and increased dead organic matter. Unfortunately, measurements of denitrification response to forest cutting have been limited to observations of increases in nitrous oxide production after the whole-tree harvest of W5 which is suggestive of increases in rates of denitrification (Bowden and Bormann 1986).
4.6 Soil Cation Exchange
A key contrast between NO3 − and K+ is the ionic charge: K+ is retained (albeit at low selectivity) on the cation exchange complex whereas NO3 − is not. Although some soils with elevated pools of iron and aluminum exhibit anion adsorption (Johnson and Todd 1983), this is not the case for the Spodosols at the HBEF (Nodvin et al. 1986b). Moreover, NO3 − is not a strongly adsorbing anion (Matson et al. 1987). The mineral soil horizons at the HBEF are coarse textured (loamy sands, low in clay), and most of the cation exchange capacity is associated with soil organic matter, especially in surface horizons (Johnson 2002); thus, K+ is retained by cation exchange especially in the upper soil. This mechanism of K+ retention would be expected to buffer K+ losses in comparison to NO3 − and thereby contribute to decoupling following large-scale disturbance . This pattern is clearly illustrated by the lower peak and longer tail on the annual NO3 − compared to K+ concentrations in stream water in W5 and W4 (Fig. 22.5). However, this pattern also is influenced by the dynamics of detrital sources (see above). Qualitative separation of these complementary effects is indicated by changes in soil exchangeable K+ pools in W5 which peaked two years after harvest and declined to pre-treatment levels between 1991 and 1998. Notably, elevated K+ concentrations in stream water persisted thereafter (Fig. 22.5b) suggesting continued increased supply from detritus. Nevertheless, the decoupling between NO3 − and K+ concentrations during the first several years after disturbance (Fig. 22.5b) is certainly explained in large part by K+ buffering by the cation exchange complex (Romanowicz et al. 1996).
5 Synthesis
Although there is considerable variation in the patterns of loss and the source-sink processes of K+ and NO3 − (discussed above) with the greatest losses occurring in W2, followed by W5 and the ice storm and W4, at a coarse scale across disturbance type and time scale, there is remarkable consistency in their stoichiometric behavior in solutions with an approximately 0.1 molar ratio of K+/NO3 − (Fig. 22.9). At a first approximation, the processes driving NO3 − loss control K+ in an approximately stoichiometric manner. Stream sites show this behavior most consistently, particularly for those stream sites near the gauging stations lower in the watersheds. There is some deviation in steam sites at the highest elevations (732 and 701 m) draining shallow soils which emerge from bedrock in W5 (so called E podzols; Bailey et al. 2014). These sites are enriched in K+ relative to NO3 − compared to lower elevation stream sites. The stoichiometric deviation at the high elevation stream sites in W5 is consistent with the pattern showing higher K+ to NO3 − in Oa solutions with decreasing values in the Bh and Bs solutions. Bs solutions in W5 are similar to stream values (although the concentrations of both solutes are markedly lower in stream water). There is also some deviation in soil solutions from the ice storm , which in contrast are enriched in NO3 − relative to K+. This deviation probably reflects lower mobilization of K+ due to limited release from dead roots given the much lower tree mortality in the ice storm in comparison to cutting treatments.

Overall response in the response of the change in K+ relative to the change in NO3 − in solutions to disturbances of different watersheds at the HBEF
Despite the remarkable consistency in the stoichiometric behavior of K+ and NO3 − (Fig. 22.9), a highly variable degree of coupling between NO3 − and K+ concentrations in the stream water draining the mature forest watersheds, W1 and W6, at the HBEF suggests that different mechanisms contribute to the pulses of high NO3 − leaching observed over the 45 years of record. An important consideration is the distribution of anions and their changes over time. Cation leaching is closely coupled with anion loss due to constraints in solution electroneutrality (Cronan et al. 1978). Under most conditions sulfate is the dominant anion in HBEF drainage waters due to elevated inputs from atmospheric deposition (Likens et al. 2002) coupled with low mineral weathering rates and limited production of HCO3 −. Note that atmospheric deposition and stream sulfate concentrations have been significantly decreasing (−0.79 and −0.49 μmol L−1 year−1, respectively) throughout the biogeochemical record at Hubbard Brook, due to emission controls (Likens et al. 2002; Fuss et al. 2015). The long-term decline in stream sulfate and NO3 − concentrations undoubtedly drive the long-term stream decline in K+ concentration. However, superimposed on this pattern of chronic sulfate and nitrate leaching are periodic forest disturbance events (forest cutting, climatic disturbance) which add pulsed inputs of NO3 −. Elevated anion leaching will facilitate cation leaching . The degree to which K+ loss occurs under these conditions depends on the net supply of this nutrient controlled by mobilization and immobilization process (mineralization, plant uptake, cation adsorption/desorption). In most cases the behavior of W1 and W6 was largely concordant in terms of this coupling during years of high NO3 − concentrations. In particular, the NO3 − and K+ concentrations were strongly coupled during four years of high stream NO3 − concentrations (1981, 2000, 2008, 2015), whereas they were clearly decoupled in 1973 (W6 only) and in 1991 (Fig. 22.2). The flushes of NO3 − from the Hubbard Brook watersheds have been attributed to reductions in plant uptake owing to disturbance by soil freezing (Mitchell et al. 1996) and ice storms (Houlton et al. 2003) and insect damage to the canopy (Aber et al. 2002). Clearly, a reduction in plant uptake of nitrogen has a much greater effect on stream NO3 − flux than for K+ due to the greater nutritional requirement coupled with the fact that K+ leaching can be facilitated by elevated sulfate in addition to NO3 −; see, for example, the relative responses of these solutes to the forest harvest experiments (Table 22.1) as well as the degree of reduction during the growing season (Fig. 22.3). Therefore, if reduced root uptake is the principal cause of elevated NO3 − flux, we might not expect a very tight coupling between these solutes in high NO3 − concentration years; other mechanisms (sulfate leaching, cation exchange) may be equally or more important.
The coupling between NO3 − and K+ concentrations following large-scale disturbance experiments in W2, W4, and W5 provides further insights into mechanisms contributing to leaching fluxes. In W5 during the second summer after harvest (1986), concentrations of K+ peaked several weeks earlier than NO3 − in Oa horizon soil solutions presumably due to a delay in peak rates of nitrification compared with rapid K+ leaching from dead fine roots. In contrast, K+ and NO3 − concentrations in Bs soil solution were strongly coupled over this interval, with both delayed relative to Oa solutions (Fig. 22.7); this pattern could reflect either or both delayed leaching of K+ from dead mineral soil roots or retention on the cation exchange complex. Notably exchangeable K+ nearly doubled in 1986 in comparison with pre-treatment levels (Romanowicz et al. 1996). However, the seasonal pattern of stream NO3 − and K+ during this peak year of leaching (1986, Fig. 22.6) exhibited weak coupling, probably reflecting the lack of NO3 − retention due to both minimal root uptake and lack of anion exchange in soil. In W4, where the forest was progressively cut over a four-year period, the flush of K+ was strongly delayed in comparison to NO3 −; for example, NO3 − concentrations returned to background levels even before the peak in K+ concentration was observed in 1980–1981 (Fig. 22.5a). The causes of this extended delay in K+ flux are not clear, nor is the extended interval of elevated stream K+ until the early 2000s. Deeper soils and the larger watershed area on W4 may contribute. It is notable that following the 1999 ice storm , which caused considerable canopy damage on W4, a coupled increase in NO3 − and K+ concentrations occurred in 2000–2001 but the magnitude of the NO3 − flush greatly exceeded that for K+ (0.013 mol K+ (mol NO3 −)−1; Fig. 22.5a).
There is a curious seasonal coupling of increased NO3 − and K+ concentrations in Oa soil solutions on W5 during a five-year interval of high K+ leaching from those surface horizons (Figs. 22.7 and 22.8). This appears to be controlled by NO3 − (r2 = 0.58) leaching in the nongrowing season which drives the release of K+ presumably from the cation exchange complex.
The most recent flush of NO3 − from the uncut watersheds at the HBEF (W1, W6) was larger and of longer duration in W1 than W6 (Fig. 22.2). The accompanying flush of K+ also differed between these two catchments. In 1999 W1 was treated with a calcium silicate (wollastonite) which increased pH in soils, soil solutions, and surface waters (Cho et al. 2012), causing a marked recovery of forest health and productivity (Battles et al. 2014). The cause of the NO3 − pulse in 2015 has not been determined; neither extensive soil freezing nor canopy damage was observed at this time. However, a reduction in forest floor mass in W1 (Johnson et al. 2014; Rosi-Marshall et al. 2016) may have contributed to this NO3 − flush via enhanced mineralization and nitrification. The 3-year peak in NO3 − concentration (2013, 2014, 2015) on W1 was much greater than the coincident K+ flush (Fig. 22.2), perhaps supporting this mechanism. However, the high K+ flush that accompanied the NO3 − peak in W6 (Fig. 22.2) may suggest otherwise.
6 Suggestions for Future Research
In this study, we evaluated the mobilization of potassium and nitrate in response to disturbances to forest ecosystems (ice storm and harvest) at the Hubbard Brook Experimental Forest, New Hampshire. We envision that the mobility of other nutrient cations such as calcium and magnesium could also be influenced by various discrete natural and anthropogenic disturbance events including windstorms, wildfire, ice storms , insects, and pathogen irruptions and harvest. Calcium is known to be in short supply in these acid soils due to inherently low weathering rates coupled with legacy effects of acidic deposition and might be considered as a limiting nutrient due to its control of the acid base status of the ecosystems (Likens et al. 1998). Understanding the consequences of various disturbances on calcium and magnesium availability is therefore research interests that should be pursued in the future.
Here, we examined the influence of disturbances on nutrient mobility over both immediate and longer periods. Most atmospheric inputs to the study ecosystem have been relatively constant over the long term; however, inputs and leaching of both nitrate and sulfate have substantially decreased in recent years due to air pollution controls. It would be useful to examine the interactive effects of changes in strong anion inputs and land disturbances on the ecosystem behavior. For example, we suspect that aluminum mobilization following disturbance was greater during the historical period when acidic deposition was more severe and mobile anion leaching was greater.
As we discussed in this chapter, Groffman et al. (2018) hypothesized that climate change is driving an “oligotrophication” of these forested watersheds. Our unpublished observations suggest that nitrate leaching following natural disturbances like ice storms and windstorms is lower now than just two decades ago. The mechanisms contributing to this hypothesized oligotrophication could be elucidated by conducting a forest harvest experiment today in the same manner as the earlier experiments to examine differences in the stoichiometry of leaching .
At Hubbard Brook Experimental Forest, long-term monitoring of gauged watersheds has provided a quantitative basis for evaluating disturbance effects on stream water chemistry. Other sites with long-term records and whole-catchment disturbance experiments, for example, Coweeta Hydrologic Laboratory and HJ Andrews Experimental Forest, would be informative choices to conduct similar analysis. We envision different responses to disturbances given differences in soils, vegetation, climate, and atmospheric inputs. For example, in soils with elevated pools of amorphous iron and aluminum oxides and high sulfate adsorption capacity, such as the non-glaciated regions of the southeastern USA (Coweeta), historical atmospheric sulfate deposition has been retained in soil to a greater extent than at HBEF. We hypothesize that in response to discrete events such as forest harvest, abrupt changes in soil acidity could release sulfate previously retained in soil to surface waters and cause more severe episodic acidification. In contrast, in the relatively pristine Pacific Northwest, lower mobility of nitrate might be expected to result in different stoichiometry of K and N following disturbance at HJ Andrews. We believe that a better understanding of the complex controls on the stoichiometry of nutrient mobility would be forthcoming with a broader evaluation of disturbance responses across a global array of forested watersheds.
A major driver of changes in soil, soil solution, and stream chemistry at the HBEF has been long-term decreases in leaching by sulfate as a result of decreases in acid deposition and the apparent and unexplained “tightening” or oligotrophication of the nitrogen cycle (Groffman et al. 2018). A critical research question is what is driving the long-term decrease in NO3 − leaching at Hubbard Brook and other forested watersheds in the Northeast United States (e.g., Driscoll et al. 2016). What is the role of decreases in NO3 − concentration and acid deposition in this phenomenon, or do other factors such as climate change contribute? Do these long-term changes in the biogeochemistry at Hubbard Brook affect the pulsed response of watersheds to climatic or harvesting disturbance? Related questions are: To what extent do decreases in sulfate and NO3 − leaching affect cation loss and the status of nutrient cations in soil? And do these changes alter the species composition and vigor of the forest community and soil development processes? If so, what are the implications for long-term ecosystem function?
References
Aber JD, Nadelhoffer KJ, Steudler P, Melillo JM (1989) Nitrogen saturation in northern forest ecosystems. BioScience 39:378–286. https://doi.org/10.2307/1311067
Aber JD, Ollinger SV, Driscoll CT, Likens GE, Holmes RT, Freuder RJ et al (2002) Inorganic nitrogen losses from a forested ecosystem in responseto physical, chemical, biotic, and climatic perturbations. Ecosystems 5:0648–0658. https://doi.org/10.1007/s10021-002-0203-8
Attiwill PM (1968) The loss of elements from decomposing litter. Ecology 49:142–145. https://doi.org/10.2307/1933568
Bailey SW, Brousseau PA, McGuire KJ, Ross DS (2014) Influence of landscape position and transient water table on soil development and carbon distribution in a steep, headwater catchment. Geoderma 226:279–289. https://doi.org/10.1016/j.geoderma.2014.02.017
Battles JJ, Cleavitt NL, Saah DS, Poling BT, Fahey TJ (2017) Ecological impact of a microburst windstorm in a northern hardwood forest. Can J For Res 47:1695–1701. https://doi.org/10.1139/cjfr-2017-0206
Battles JJ, Fahey TJ, Driscoll CT Jr, Blum JD, Johnson CE (2014) Restoring soil calcium reverses forest decline. Environ Sci Technol Lett 1:15–19. https://doi.org/10.1021/ez400033d
Bormann FH, Likens GE (1967) Nutrient cycling. Science 155:424–429. https://doi.org/10.1126/science.155.3761.424
Bormann FH, Likens GE (1979) Catastrophic disturbance and the steady state in northern hardwood forests: A new look at the role of disturbance in the development of forest ecosystems suggests important implications for land-use policies. Am Sci 67:660–669
Bowden WB, Bormann FH (1986) Transport and loss of nitrous oxide in soil water after forest clear-cutting. Science 233:867–869. https://doi.org/10.1126/science.233.4766.867
Buso DC, Likens GE, Eaton JS (2000) Chemistry of precipitation, streamwater, and lakewater from the Hubbard Brook Ecosystem Study: a record of sampling protocols and analytical procedures. Gen Tech Rep NE-275 Newtown Sq PA US Dep Agric For Serv Northeast Res Stn 52 P 275
Campbell JL, Mitchell MJ, Mayer B, Groffman PM, Christenson LM (2007) Mobility of nitrogen-15-labeled nitrate and sulfur-34-labeled sulfate during snowmelt. Soil Sci Soc Am J 71:1934–1944. https://doi.org/10.2136/sssaj2006.0283
Campbell JL, Socci AM, Templer PH (2014) Increased nitrogen leaching following soil freezing is due to decreased root uptake in a northern hardwood forest. Glob Change Biol 20:2663–2673. https://doi.org/10.1111/gcb.12532
Cho Y, Driscoll CT, Johnson CE, Blum JD, Fahey TJ (2012) Watershed-level responses to calcium silicate treatment in a northern hardwood forest. Ecosystems 15:416–434. https://doi.org/10.1007/s10021-012-9518-2
Cleavitt NL, Fahey TJ, Groffman PM, Hardy JP, Henry KS, Driscoll CT (2008) Effects of soil freezing on fine roots in a northern hardwood forest. Can J For Res 38:82–91. https://doi.org/10.1139/X07-133
Cronan CS, Reiners WA, Reynolds RC, Lang GE (1978) Forest floor leaching: contributions from mineral, organic, and carbonic acids in New Hampshire subalpine forests. Science 200:309–311. https://doi.org/10.1126/science.200.4339.309-a
Dahlgren RA, Driscoll CT (1994) The effects of whole-tree clear-cutting on soil processes at the Hubbard Brook Experimental Forest, New Hampshire, USA. Plant Soil 158:239–262. https://doi.org/10.1007/BF00009499
Dittman JA, Driscoll CT, Groffman PM, Fahey TJ (2007) Dynamics of nitrogen and dissolved organic carbon at the Hubbard Brook Experimental Forest. Ecology 88:1153–1166. https://doi.org/10.1890/06-0834
Driscoll C, Whitall D, Aber J, Boyer E, Castro M, Cronan C et al (2003) Nitrogen pollution: Sources and consequences in the US northeast. Environ Sci Policy Sustain Dev 45:8–22. https://doi.org/10.1080/00139150309604553
Driscoll CT, Driscoll KM, Fakhraei H, Civerolo K (2016) Long-term temporal trends and spatial patterns in the acid-base chemistry of lakes in the Adirondack region of New York in response to decreases in acidic deposition. Atmos Environ 146:5–14. https://doi.org/10.1016/j.atmosenv.2016.08.034
Fahey TJ, Battles JJ, Wilson GF (1998) Responses of early successional northern hardwood forests to changes in nutrient availability. Ecol Monogr 68:183–212. https://doi.org/10.2307/2657200
Fahey TJ, Hughes JW, Pu M, Arthur MA (1988) Root decomposition and nutrient flux following whole-tree harvest of northern hardwood forest. For Sci 34:744–768. https://doi.org/10.1093/forestscience/34.3.744
Federer CA, Hornbeck JW, Tritton LM, Martin CW, Pierce RS, Smith CT (1989) Long-term depletion of calcium and other nutrients in eastern US forests. Environ Manage 13:593–601. https://doi.org/10.1007/BF01874965
Fuss CB, Driscoll CT, Campbell JL (2015) Recovery from chronic and snowmelt acidification: Long-term trends in stream and soil water chemistry at the Hubbard Brook Experimental Forest, New Hampshire, USA. J Geophys Res Biogeosciences 120:2360–2374. https://doi.org/10.1002/2015JG003063
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP et al (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226. https://doi.org/10.1007/s10533-004-0370-0
Gannon JP, Bailey SW, McGuire KJ (2014) Organizing groundwater regimes and response thresholds by soils: A framework for understanding runoff generation in a headwater catchment. Water Resour Res 50:8403–8419. https://doi.org/10.1002/2014WR015498
Groffman PM, Driscoll CT, Durán J, Campbell JL, Christenson LM, Fahey TJ et al (2018) Nitrogen oligotrophication in northern hardwood forests. Biogeochemistry 141:523–539. https://doi.org/10.1007/s10533-018-0445-y
Hewlett JD, Hibbert AR (1967) Factors affecting the response of small watersheds to precipitation in humid areas. In: Sopper WE, Lull HW (eds) Forest hydrology. Pergamon Press, New York, pp 275–290
Hirsch RM, Slack JR, Smith RA (1982) Techniques of trend analysis for monthly water quality data. Water Resour Res 18:107–121. https://doi.org/10.1029/WR018i001p00107
Hornbeck JW, Martin CW, Pierce RS, Bormann FH, Likens GE, Eaton JS (1986) Clearcutting northern hardwoods: effects on hydrologic and nutrient ion budgets. For Sci 32:667–686. https://doi.org/10.1093/forestscience/32.3.667
Houlton BZ, Driscoll CT, Fahey TJ, Likens GE, Groffman PM, Bernhardt ES et al (2003) Nitrogen dynamics in ice storm-damaged forest ecosystems: implications for nitrogen limitation theory. Ecosystems 6:431–443. https://doi.org/10.1007/s10021-002-0198-1
Irland LC (1998) Ice storm 1998 and the forests of the Northeast: A preliminary assessment. J For 96:32–40. https://doi.org/10.1093/jof/96.9.32
Johnson CE (2002) Cation exchange properties of acid forest soils of the northeastern USA. Eur J Soil Sci 53:271–282. https://doi.org/10.1046/j.1365-2389.2002.00441.x
Johnson CE, Driscoll CT, Blum JD, Fahey TJ, Battles JJ (2014) Soil chemical dynamics after calcium silicate addition to a northern hardwood forest. Soil Sci Soc Am J 78:1458–1468. https://doi.org/10.2136/sssaj2014.03.0114
Johnson DW, Cole DW (1980) Anion mobility in soils: relevance to nutrient transport from forest ecosystems. Environ Int 3:79–90. https://doi.org/10.1016/0160-4120(80)90040-9
Johnson DW, Todd DE (1983) Relationships among iron, aluminum, carbon, and sulfate in a variety of forest soils 1. Soil Sci Soc Am J 47:792–800. https://doi.org/10.2136/sssaj1983.03615995004700040035x
Kendall C, Campbell DH, Burns DA et al (1995) Tracing sources of nitrate in snowmelt runoff using the oxygen and nitrogen isotopic compositions of nitrate. In: Tonnessen KA, Williams MW, Tranter M (eds) Biogeochemistry of seasonally snow-covered catchments, IAHS Publ No. 228. IAHS, Wallingford, UK, pp 339–348
Likens GE (2013) Biogeochemistry of a forested ecosystem. Springer, New York
Likens GE, Bormann FH, Johnson NM, Fisher DW, Pierce RS (1970) Effects of forest cutting and herbicide treatment on nutrient budgets in the Hubbard Brook Watershed-Ecosystem. Ecol Monogr 40:23–47. https://doi.org/10.2307/1942440
Likens GE, Bormann FH, Johnson NM, Pierce RS (1967) The calcium, magnesium, potassium, and sodium budgets for a small forested ecosystem. Ecology 48:772–785. https://doi.org/10.2307/1933735
Likens GE, Bormann FH, Pierce RS, Reiners WA (1978) Recovery of a deforested ecosystem. Science 199:492–496. https://doi.org/10.1126/science.199.4328.492
Likens GE, Driscoll CT, Buso DC, Siccama TG, Johnson CE, Lovett GM et al (1998) The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 41:89–173. https://doi.org/10.1023/A:1005984620681
Likens GE, Driscoll CT, Buso DC, Mitchell MJ, Lovett GM, Bailey SW et al (2002) The biogeochemistry of sulfur at Hubbard Brook. Biogeochemistry 60:235–316. https://doi.org/10.1023/A:1020972100496
Likens GE, Driscoll CT, Buso DC, Siccama TG, Johnson CE, Lovett GM et al (1994) The biogeochemistry of potassium at Hubbard Brook. Biogeochemistry 25:61–125. https://doi.org/10.1007/BF00000881
Lloret J, Valiela I (2016) Unprecedented decrease in deposition of nitrogen oxides over North America: the relative effects of emission controls and prevailing air-mass trajectories. Biogeochemistry 129:165–180. https://doi.org/10.1007/s10533-016-0225-5
Lovett GM, Goodale CL, Ollinger SV, Fuss CB, Ouimette AP, Likens GE (2018) Nutrient retention during ecosystem succession: a revised conceptual model. Front Ecol Environ 16:532–538. https://doi.org/10.1002/fee.1949
Lovett GM, Nolan SS, Driscoll CT, Fahey TJ (1996) Factors regulating throughfall flux in a New Hampshire forested landscape. Can J For Res 26:2134–2144. https://doi.org/10.1139/x26-242
Matson PA, Vitousek PM, Ewel JJ, Mazzarino MJ, Robertson GP (1987) Nitrogen transformations following tropical forest felling and burning on a volcanic soil. Ecology 68:491–502. https://doi.org/10.2307/1938454
McColl JG, Cole DW (1968) A mechanism of cation transport in a forest soil. Northwest Sci 42:134–141
Merrens EJ, Peart DR (1992) Effects of hurricane damage on individual growth and stand structure in a hardwood forest in New Hampshire, USA. J Ecol 80:787–795. https://doi.org/10.2307/2260866
Mitchell MJ, Driscoll CT, Kahl JS, Murdoch PS, Pardo LH (1996) Climatic control of nitrate loss from forested watersheds in the Northeast United States. Environ Sci Technol 30:2609–2612. https://doi.org/10.1021/es9600237
Morse JL, Durán J, Groffman PM (2015) Soil denitrification fluxes in a northern hardwood forest: the importance of snowmelt and implications for ecosystem N budgets. Ecosystems 18:520–532. https://doi.org/10.1007/s10021-015-9844-2
Mou P, Fahey TJ, Hughes JW (1993) Effects of soil disturbance on vegetation recovery and nutrient accumulation following whole-tree harvest of a northern hardwood ecosystem. J Appl Ecol 30:661–675. https://doi.org/10.2307/2404245
Nodvin SC, Driscoll CT, Likens GE (1986a) The effect of pH on sulfate adsorption by a forest soil. Soil Sci 142:69–75
Nodvin SC, Driscoll CT, Likens GE (1986b) Simple partitioning of anions and dissolved organic carbon in a forest soil. Soil Sci 142:27–35
Parker GG (1983) Throughfall and stemflow in the forest nutrient cycle. Adv Ecol Research 13:57–133. https://doi.org/10.1016/S0065-2504(08)60108-7
Peters SC, Blum JD, Driscoll CT, Likens GE (2004) Dissolution of wollastonite during the experimental manipulation of Hubbard Brook Watershed 1. Biogeochemistry 67:309–329. https://doi.org/10.1023/B:BIOG.0000015787.44175.3f
Rhoads AG, Hamburg SP, Fahey TJ, Siccama TG, Hane EN, Battles J et al (2002) Effects of an intense ice storm on the structure of a northern hardwood forest. Can J For Res 32:1763–1775. https://doi.org/10.1139/x02-089
Romanowicz RB, Driscoll CT, Johnson CE, Fahey TJ, Likens GE, Siccama TG (1996) Changes in the biogeochemistry of potassium following a whole-tree harvest. Soil Sci Soc Am J 60:1664–1674. https://doi.org/10.2136/sssaj1996.03615995006000060009x
Rosi-Marshall EJ, Bernhardt ES, Buso DC, Driscoll CT, Likens GE (2016) Acid rain mitigation experiment shifts a forested watershed from a net sink to a net source of nitrogen. Proc Natl Acad Sci 113:7580–7583. https://doi.org/10.1073/pnas.1607287113
Runkle JR (1982) Patterns of Disturbance in some old-growth mesic forests of eastern North America. Ecology 63:1533–1546. https://doi.org/10.2307/1938878
Ryan DF, Bormann FH (1982) Nutrient resorption in northern hardwood forests. BioScience 32:29–32. https://doi.org/10.2307/1308751
Sen PK (1968) On a class of aligned rank order tests in two-way layouts. Ann Math Stat 39:1115–1124. https://doi.org/10.1214/aoms/1177698236
Tierney GL, Fahey TJ, Groffman PM, Hardy JP, Fitzhugh RD, Driscoll CT (2001) Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry 56:175–190. https://doi.org/10.1023/A:1013072519889
Van Miegroet H, Cole DW (1984) The impact of nitrification on soil acidification and cation leaching in a red alder ecosystem 1. J Environ Qual 13:586–590. https://doi.org/10.2134/jeq1984.00472425001300040015x
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler PW et al (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750. https://doi.org/10.1890/1051-0761(1997)007[0737:HAOTGN]2.0.CO;2
Yanai RD, Vadeboncoeur MA, Hamburg SP, Arthur MA, Fuss CB, Groffman PM et al (2013) From missing source to missing sink: long-term changes in the nitrogen budget of a northern hardwood forest. Environ Sci Technol 47:11440–11448. https://doi.org/10.1021/es4025723
Acknowledgments
This is a contribution of the Hubbard Brook Ecosystem Study. Support for this work was largely provided by a number of grants from the National Science Foundation, particularly through the Long-Term Ecological Research program. Support is also provided by the US Forest Service. The Hubbard Brook Experimental Forest is administered by the US Forest Service. We thank Gene Likens and John Campbell for providing us chemistry of streamwater and streamflow data measured in the Hubbard Brook watersheds.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Fakhraei, H., Fahey, T.J., Driscoll, C.T. (2020). The Biogeochemical Response of Nitrate and Potassium to Landscape Disturbance in Watersheds of the Hubbard Brook Experimental Forest, New Hampshire, USA. In: Levia, D.F., Carlyle-Moses, D.E., Iida, S., Michalzik, B., Nanko, K., Tischer, A. (eds) Forest-Water Interactions. Ecological Studies, vol 240. Springer, Cham. https://doi.org/10.1007/978-3-030-26086-6_22
Download citation
DOI: https://doi.org/10.1007/978-3-030-26086-6_22
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-26085-9
Online ISBN: 978-3-030-26086-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)