
Abstract
A key strategy in agroecology is to restore functional biodiversity in agroecosystems at field and landscape levels. Biodiversity performs essential ecological services, and, if correctly assembled in time and space, it can lead to agroecosystems capable of sponsoring their own crop protection. Diversity can be enhanced in crop fields by means of cover crops, intercropping, agroforestry, crop/livestock mixtures, and, in the surrounding landscapes, using shelterbelts, hedgerows, corridors, etc. Correct agroecosystem diversification strategies usually regulate pests by restoring the natural control of insect pests. This chapter provides a number of studies performed around the world, including Neotropical regions, about the effects of crop diversity on the enhancement of natural enemies and its impacts on the densities of herbivore pests in various crop systems.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
About 80% of the 1.5 billion hectares devoted to agriculture in the planet are occupied by industrial large-scale monocultures of maize, soybean, rice, and wheat. Due to their genetic homogeneity, these systems have proven to be highly vulnerable to insect invasions and disease epidemics (Heinemann et al. 2013). To keep pests at bay, about 2.3 billion kg of pesticides are applied worldwide per year, and today there are about 450 insecticide-resistant insects, indicating that the technology is reaching its limits. Moreover, secondary pest outbreaks are common in pesticide-loaded crops due to elimination of natural enemies (Hayes and Hansen 2017). Total removal of pesticides can restore natural enemy diversity and lead to renewed biological control of specific pests. Within 2 years, virtually, all banana insect pests in Golfito, Costa Rica, dropped to below economic threshold levels due to enhanced parasitization and predation after stopping insecticide (dieldrin and carbaryl) sprays. Similarly, in walnut orchards of California, natural biological control of the frosted scale and the calico scale was soon achieved by encyrtid parasitoids after removal of dichlorodiphenyltrichloroethyne (DDT) sprays (Croft 1990).
In insecticide-free agroecosystems, such as organic farms, communities of predators tend to be more even exerting stronger pest control pressure (Crowder et al. 2010). Despite the fact that organic farming potentially offers a means of augmenting natural pest control, most commercial organic farms practice monoculture under input-substitution management, which does not offer optimal environmental conditions (habitat, refuge, alternative food sources, etc.) to natural enemies. Many researchers have shown that by adding plant diversity to monocultures, it is possible to exert changes in habitat complexity which in turn favors natural enemy abundance and effectiveness due to enhanced availability of alternate prey, nectar sources, and suitable microhabitats (Altieri and Nicholls 2004).
A key strategy in agroecology is to restore functional biodiversity at field and landscape levels. Biodiversity performs key ecological services and if correctly assembled in time and space can lead to agroecosystems capable of sponsoring their own soil fertility, crop protection, and productivity (Altieri 1999). Diversity can be enhanced in crop fields with cover crops, intercropping, agroforestry, crop/livestock mixtures, and in the surrounding landscapes via shelterbelts, hedgerows, corridors, etc. Generally, correct agroecosystem diversification strategies result in pest regulation through restoration of natural control of insect pests (Altieri and Nicholls 2014).
2 Manipulating Biodiversity at the Landscape Level
One key characteristic of modern agricultural systems is the large size and homogeneity of crop monocultures that fragment the natural landscape, directly affecting abundance and diversity of natural enemies. Such simplification can have serious ecological implications for biological control, such as in the case of four US Midwest states where recent biofuel-driven growth in maize and soybean planting resulted in lower landscape diversity, decreasing the supply of pest natural enemies to maize and soybean fields and reducing biocontrol services by 24%. This loss of biocontrol services cost soybean and maize producers in these states an estimated $58 million per year in reduced yield and increased pesticide use (Landis et al. 2008).
One way to reintroduce biodiversity into large-scale monocultures is by establishing vegetationally diverse field margins and/or hedgerows that may serve as biological corridors. There is wide acceptance of the importance of field margins as reservoirs of the natural enemies of crop pests, as these habitats provide overwintering sites, increased resources such as alternative prey/hosts, pollen, and nectar for parasitoids and predators (Gurr et al. 1998).
2.1 Effects of Hedges and Surrounding Natural Vegetation
Many studies have demonstrated increased abundance of natural enemies and more effective biological control where crops are bordered by wild vegetation from which natural enemies colonize adjacent crop fields. Parasitism of armyworm, Pseudaletia unipunctata (Haworth) (Noctuidae), was significantly higher in maize fields embedded in a complex landscape than in maize fields surrounded by simpler habitats. In a 2-year study, researchers found higher parasitism of Ostrinia nubilalis (Hübner) (Crambidae) larvae by the parasitoid Eriborus terebrans (Gravenhorst) (Ichneumonidae) in edges of maize fields adjacent to wooded areas than in interior fields (Landis et al. 2000).
Similarly, in Germany, parasitism of rape pollen beetle was about 50% at the edge of the fields, while at the center of the fields, parasitism dropped significantly to 20% (Thies and Tscharntke 1999). In Hawaii, the presence of nectar-source plants in sugarcane field margins allowed population levels to rise and increased the efficiency of the sugarcane weevil parasite, Lixophaga sphenophori (Villeneuve) (Tachinidae) (Thopham and Beardsley 1975). The authors suggest that the effective range of the parasite within cane fields is limited to about 45–60 meters from nectar sources present in the field margins. Continuous herbicidal elimination of field margin nectar source plants had a detrimental effect on populations of Lixophaga and, therefore, led to a decrease in the efficiency of the parasite as a biocontrol agent of the weevil. Maier (1981) observed higher parasitization rates of apple maggot [Rhagoletis pomonella (Walsh) (Tephritidae)] by braconids in apple and hawthorn orchards in northern Connecticut, where plants such as blueberry (Vaccinium spp.), dogwood (Cornus spp.), and winterberry (Ilex ciliata) commonly grew nearby. These plants support populations of several frugivorous Tephritidae that serve as alternate hosts to braconids.
In many cases, weeds and other natural vegetation around crop fields harbor alternate hosts/prey for natural enemies, thus providing seasonal resources to bridge the gaps in the life cycles of entomophagous insects and crop pests. A classic case is that of the egg parasitoid wasp Anagrus epos Girault (Mymaridae), whose effectiveness in regulating the grape leafhopper, Erythroneura elegantula Osborn (Cicadellidae), was increased greatly in vineyards near areas invaded by wild blackberry (Rubus sp.). This plant supports an alternative host leafhopper [Dikrella cruentata (Gillette)], which breeds in its leaves in winter (Doutt and Nakata 1973). Recent studies have shown that prune trees planted next to vineyards also allowed early-season buildup of A. epos. After surviving the winter on an alternate host, the prune leafhopper, Anagrus, wasps move into the vineyard in the spring, providing grape leafhopper control up to a month earlier than in vineyards not near prune tree refuges (Pickett and Bugg 1998).
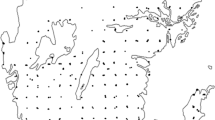
Murphy et al. (1998) completed a rigorous evaluation of the effectiveness of French prune trees in increasing control of the grape leafhopper. Results from this study indicated that there was a consistent and significant pattern of higher parasitism in grape vineyards with adjacent prune tree refuges than in vineyards lacking refuges. Researchers now recommend that trees should always be planted upwind from the vineyard, but otherwise can be managed as a typical commercial prune orchard; and to plant as many trees as is economically feasible, since the more trees there are, the more productive the refuge is likely to be (Corbett and Rosenheim 1996) (Fig. 1.1).

Hypothesized sources of Anagrus colonizing vineyards early in the season. Anagrus colonize vineyards from adjacent French prune tree refuges. Anagrus also colonize from external overwintering sites. The windbreak effect generated by prune trees causes increased colonization by external Anagrus immediately downwind of refuges. (Corbett and Rosenheim 1996)
2.2 Corridors
One way to introduce the beneficial biodiversity from surrounding landscapes into large-scale monocultures is by establishing vegetationally diverse corridors that allow the movement and distribution of useful arthropod biodiversity into the center of monocultures. Nicholls et al. (2001) established a vegetational corridor that connected to a riparian forest and cut across a vineyard monoculture. The corridor allowed natural enemies emerging from the riparian forest to disperse over large areas of otherwise monoculture vineyard systems. The corridor provided a constant supply of alternative food for predators effectively decoupling predators from a strict dependence on grape herbivores and avoiding a delayed colonization of the vineyard.
This complex of predators continuously circulated into the vineyard interstices, establishing a set of trophic interactions leading to a natural enemy enrichment, which also led to lower numbers of leafhoppers and thrips on vines. Generalist predators in the families Coccinellidae, Chrysopidae, Nabidae, and Syrphidae exhibited a density gradient in the vineyard, indicating that the abundance and spatial distribution of these insects were influenced by the presence of the corridor that channeled dispersal of the insects into adjacent vines. Adult leafhoppers exhibited a clear density gradient reaching lowest numbers in vine rows near the corridor and forest and increasing in numbers toward the center of the field, away from the adjacent vegetation. The highest concentration of leafhoppers occurred after the first 20–25 rows (30–40 m) downwind from the corridor.
Researchers in Switzerland introduced successional strips of annual flowering plants into cereal fields (Lys and Nentwig 1992). Significantly higher predator activity was found in the strip-managed fields than in the control monocultures, especially for carabid beetles such as Poecilus cupreus L., Carabus granulates L., and Pterostichus melanarius Illiger. Several observations led to the conclusion that this higher activity was generally due to a prolongation of the reproductive period in the strip-managed area. Besides the marked increase in activity and density, a large increase in the diversity of ground beetle species was observed. After 3 years of research, the authors concluded that flower strips offer not only higher food availability but also more suitable overwintering sites. Flower strips increase the chance of survival of many carabid species in arable ecosystems thus counteracting the faunal impoverishment trends promoted by monocultures. Nentwig (1988) found similar effects with 3–9-m-wide sown weed strips dividing large fields in small parts so that the distance between strips does not exceed 50–100 m. A favorite plant to be used as strips within or around fields is Phacelia tanacetifolia.
3 Manipulating Plant Biodiversity at the Field Level
3.1 Ecological Theory
It is accepted by many entomologists that inter-species plant diversity reduces crop vulnerability to insect pests. There is a large body of literature documenting that diversification of cropping systems (variety mixtures, polycultures, agroforestry systems, etc.) often lead to reduced herbivore populations (Risch et al. 1983; Altieri and Nicholls 2004). Two hypotheses have been offered to explain such reductions (Andow 1991).
The natural enemy hypothesis predicts that there will be a greater abundance and diversity of natural enemies of pest insects in polycultures than in monocultures. Predators tend to be polyphagous and have broad habitat requirements, so they would be expected to encounter a greater array of alternative prey and microhabitats in a heterogeneous environment (Russell 1989).
A study that supports the enemy hypothesis was conducted in tropical corn/bean/squash systems where Letourneau (1987) studied the importance of parasitic wasps in mediating the differences in pest abundance between simple and complex crop arrangements. A squash-feeding caterpillar, Diaphania hyalinata L. (Crambidae), occurred at low densities on intercropped squash in tropical Mexico (Letourneau 1987). Part of the effect of the associated maize and bean plants may have been to render the squash plants less apparent to ovipositing moths. Polyculture fields also harbored greater numbers of parasitic wasps than did squash monocultures. Malaise trap captures of parasitic wasps in monoculture consisted of one-half the number of individuals caught in mixed culture. The parasitoids of the target caterpillars were also represented by higher numbers in polycultures throughout the season. Not only were parasitoids more common in the vegetationally diverse traditional system; also the parasitization rates of D. hyalinata eggs and larvae on squash were higher in polycultures. Approximately 33% of the eggs in polyculture samples over the season were parasitized and only 11% of eggs in monocultures. Larval samples from polycultures showed an incidence of 59% parasitization for D. hyalinata larvae, whereas samples from monoculture larval specimens were 29% parasitized.
The resource concentration hypothesis is based on the fact that insect populations can be influenced directly by the concentration and spatial dispersion of their food plants. Many herbivores, particularly those with narrow host ranges, are more likely to find and remain on hosts that are growing in dense or nearly pure stands and which are thus providing concentrated resources and monotonous physical conditions (Andow 1991).
One study that supports this hypothesis (Risch 1981) looked at the population dynamics of six chrysomelid beetles in monocultures and polycultures of maize/bean/squash (Cucurbita pepo). In polycultures containing at least one nonhost plant (maize), the number of beetles per unit was significantly lower relative to the numbers of beetles on host plants in monocultures. Measurement of beetle movements in the field showed that beetles tended to emigrate more from polycultures than from host monocultures.
Apparently, this was due to several factors: (a) beetles avoided host plants shaded by maize, (b) maize stalks interfered with flight movements of beetles, and (c) as beetles moved through polycultures, they remained on nonhost plants for a significantly shorter time than on host plants. There were no differences in rates of parasitism or predation of beetles between systems (Risch 1981).
A second study (Bach 1980) examined the effects of plant diversity on the cucumber beetle, Acalymma vittata (Fabricius). Population densities were significantly greater in cucumber (Cucumis sativus) monocultures than in polycultures containing cucumber and two nonhost species. Bach also found greater tenure time of beetles in monocultures than in polycultures. She also determined that these differences were caused by plant diversity per se and not by differences in host plant density or size. Nevertheless, she did not reveal if differences in numbers of herbivores between monocultures and polycultures are due to diversity or rather to the interrelated and confounding effects of plant diversity, plant density, and host plant patch size.
3.2 Research Evidence
Over the last 40 years, many studies have evaluated the effects of crop diversity on densities of herbivore pests and have tried to prove one or both ecological hypothesis. An early review by Risch et al. (1983) summarized 150 published studies on the effect of diversifying an agroecosystem on insect pest abundance; 198 total herbivore species were examined in these studies. Fifty-three percent of these species were found to be less abundant in the more diversified system, 18% were more abundant in the diversified system, 9% showed no difference, and 20% showed a variable response.
Eight years later, Andow (1991) analyzed results from 209 studies involving 287 pest species and found that compared with monocultures, the population of pest insects was lower in 52% of the studies, i.e., 149 species and higher in 15% of the studies, i.e., 44 species. Of the 149 pest species with lower populations in intercrops, 60% were monophagous and 28% polyphagous. The population of natural enemies of pests was higher in the intercrop in 53% of the studies and lower in 9%. The reduction in pest numbers was almost twice for monophagous insects (53.5% of the case studies showed lowered numbers in polycultures) than for polyphagous insects (33.3% of the cases).
In a meta-analysis of 21 studies comparing pest suppression in polyculture versus monoculture, Tonhasca Jr. and Byrne (1994) found that polycultures significantly reduced pest densities by 64%. In a later meta-analysis, Letourneau et al. (2011) found a 44% increase in abundance of natural enemies (148 comparisons), a 54% increase in herbivore mortality, and a 23% reduction in crop damage on farms with species-rich vegetational diversification systems than on farms with species-poor systems.
Unequivocally, earlier reviews and recent meta-analyses suggest that diversification schemes generally achieve significant positive outcomes including natural enemy enhancement, reduction of herbivore abundance, and reduction of crop damage from a combination of bottom-up and top-down effects.
A study conducted in Kenya at the International Center of Insect Physiology and Ecology (ICIPE) added a new dimension to the above studies by showing that interactions in polycultures are mediated by the relationship between chemical ecology and agrobiodiversity. Scientists developed a habitat management system to control the stem borer, which uses two kinds of crops that are planted together with maize: a plant that repels these borers (the push) and another that attracts (pulls) them (Khan et al. 2000).
The plant chemistry responsible for stemborer control involves release of attractive volatiles from the trap plants and repellent volatiles from the intercrops. Two of the most useful trap crops that pull in the natural enemies of borers, such as the parasitic wasp [Cotesia sesamiae (Cameron) (Braconidae)], are Napier grass and Sudan grass, both important fodder plants; these are planted in a border around the maize.
Two excellent borer-repelling crops, which are planted between the rows of maize, are molasses grass, which also repels ticks, and the leguminous silverleaf (Desmodium), which in addition can suppress the parasitic weed Striga by a factor of 40 compared to maize monocrop. The N-fixing ability of Desmodium increases soil fertility, leading to a 15–20 percent increase in maize yield (Kahn et al. 1998). The push-pull strategy was adopted by more than 10,000 households in 19 districts in Kenya, 5 districts in Uganda, and 2 districts in Tanzania helping participating farmers to increase their maize yields by an average of 20% in areas where only stemborers are present and by more than 50% in areas where both stemborers and Striga weed are problems.
Participating farmers in the breadbasket of Trans-Nzoia reported a 15–20% increase in maize yield. In the semiarid Suba district – plagued by both stemborers and striga – a substantial increase in milk yield has occurred in the last 4 years, with farmers now being able to support grade cows on the fodder produced by Desmodium and other plants. When farmers plant maize, napier, and desmodium together, a return of US$ 2.30 for every dollar invested is made.
4 Conclusions
A community of organisms in an agroecosystem becomes more complex when a larger number of different kinds of plants are included, leading to more interactions among arthropods and microorganisms, components of above and below ground food webs. As diversity increases, so do opportunities for coexistence and beneficial interference between species that can enhance agroecosystem sustainability.
Diverse systems encourage complex food webs, which entail more potential connections and interactions among members, creating many alternative paths for energy and material flow. For this reason, a more complex community exhibits more stable production and less fluctuations in the numbers of undesirable organisms. By enhancing functional biodiversity, a major goal of the agroecological conversion process is achieved: strengthening the weak ecological functions in the agro-ecosystem, allowing farmers to gradually eliminate inputs altogether by relying instead on ecosystem functions (Nicholls et al. 2016).
Hundreds of studies show that complementary interactions occur between crops grown in polycultures and between adjacent cultivated and uncultivated vegetational components of agroecosystems. These interactions can have positive or negative, direct or indirect effects on the biological control of specific crop pests.
The exploitation of these interactions in real situations involves agroecosystem design and management and requires an understanding of the numerous relationships among plants, herbivores, and natural enemies (Altieri and Nicholls 2014). One of the major problems has been predicting which plant biodiversity spatial arrangements will lead to reduced pest abundance, since not all combinations of crops will produce the desired effect and blind adherence to the principle that a more diversified system will reduce pest infestation is clearly inadequate (Gurr et al. 1998).
Therefore, there is a need for greater understanding of the mechanisms involved to explain how, where, and when pest reduction occur and of identifying the type of biodiversity that is desirable to maintain and/or enhance in order to carry out ecological services and then to determine the best practices that will encourage the desired biodiversity components. Regardless of the need for more research in this area, several farmers and Integrated Pest Management (IPM) practitioners have identified various combinations of crops, trees, and natural vegetation that bolster biological control, and many use them commercially, such as introducing flowering cover crops in vineyards or deploying corridors of alyssum in vegetable crops, phacelia strips and beetle banks in cereal crops, etc.
There are many agricultural practices and designs that have the potential to enhance functional biodiversity, and others that negatively affect it. The idea is to apply the best management practices in order to enhance or regenerate the kind of biodiversity that can best subsidize the sustainability of agroecosystems by providing ecological services such as biological pest control, nutrient cycling, water and soil conservation, etc. (Fig. 1.2).

The effects of agroecosystem management and associated cultural practices on the biodiversity of natural enemies and the abundance of insect pests. (Altieri and Nicholls 2004)
The role of agroecologists should be to encourage those agricultural practices that increase the abundance and diversity of above- and below-ground organisms, which in turn provide key ecological services to agroecosystems (Altieri and Nicholls 1999). In order for this diversification strategy to be more rapidly implemented, a much better understanding of the ecology of parasitoids and predators within and outside of the cultivated habitat and identifying those resources that are necessary for their survivorship and reproduction (Altieri and Letourneau 1982). It is also important to determine to what extent populations within the crop contribute to the overall natural enemy metapopulation in subsequent years. If these contributions are minor, then investments in habitat management should be oriented specifically to increasing the source populations outside the crop to ensure a greater number of immigrants each year, an action parallel to increasing the dosage of a chemical biocide. However, if the subpopulations within the cropping system contribute significantly to the year-to-year metapopulation dynamics, then habitat modifications should not only consider tactics fostering immigration into the crop but also those augmenting the probability of successional emigration when this habitat becomes unsuitable. Such actions could include the addition of plant species to provide alternate hosts and/or food sources, habitats as suitable overwintering sites, or the provision of corridors within the cropping system to facilitate movement between the subcomponents of the metapopulation (Pickett and Bugg 1998).
From a management perspective, there are four key issues to consider when implementing habitat management: (a) the selection of the most appropriate plant species and their spatial/temporal deployment; (b) identify the natural enemy complexes associated with such plant arrangements, the predator/parasitoid behavioral mechanisms that are influenced by the manipulation; (c) the spatial scale over which the habitat enhancement operates; and (d) the potential negative aspects associated with adding new plants to the agroecosystem, as obviously proposed habitat management techniques must fit existing cropping systems and adapt to the needs and circumstances of farmers.
References
Altieri MA (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74(1/3):19–31
Altieri MA, Letourneau DK (1982) Vegetation management and biological control in agroecosystems. Crop Prot 1(4):405–430
Altieri MA, Nicholls CI (1999) Biodiversity, ecosystem function and insect pest management in agricultural systems. In: Collins WW, Qualset CO (eds) Biodiversity in agroecosystems. CRC Press, Boca Raton, pp 69–84
Altieri MA, Nicholls CI (2004) Biodiversity and pest management in agroecosystems, 2nd edn. Waworth Press, New York
Altieri MA, Nicholls CI (2014) Manage insects in your farm: a guide to ecological strategies, sustainable agriculture research and education handbook series, vol 7. Sustainable Agriculture Network, College Park
Andow DA (1991) Vegetational diversity and arthropod population response. Annu Rev Entomol 36:561–586
Bach CE (1980) Effects of plant density and diversity on the population dynamics of a specialist herbivore, the striped cucumber beetle, Acalymma vittata (Fab.). Ecology 61(6):1515–1530
Corbett A, Rosenheim JA (1996) Impact of a natural enemy overwintering refuge and its interaction with the surrounding landscape. Ecol Entomol 21(2):155–164
Croft BA (1990) Arthropod biological control agents and pesticides. Wiley, New York
Crowder DW, Northfield TD, Strand MR et al (2010) Organic agriculture promotes evenness and natural pest control. Nature 466(7302):109–112
Doutt RL, Nakata J (1973) The rubus leafhopper and its egg parasitoid: an endemic biotic system useful in grape-pest management. Environ Entomol 2(3):381–386
Gurr GM, Van Emden HF, Wratten SD (1998) Habitat manipulation and natural enemy efficiency: implications for the control of pests. In: Barbosa P (ed) Conservation biological control. Academic Press, New York, pp 155–183
Hayes TB, Hansen M (2017) From silent spring to silent night: agrochemicals and the anthropocene. J Elementa Sci Anthrop. https://doi.org/10.1525/elementa.246. Accessed 2 Nov 2017
Heinemann JA, Massaro M, Coray DS et al (2013) Sustainability and innovation in staple crop production in the US Midwest. Int J Agr Sustain 12(1):71–88
Kahn ZR, Ampong-Nyarko K, Hassanali A et al (1998) Intercropping increases parasitism of pests. Nature 388:631–632
Khan ZR, Pickett JA, Van den Berg J et al (2000) Exploiting chemical ecology and species diversity: stemborer and striga control for maize and sorghum in Africa. Pest Manag Sci 56:957–962
Landis DA, Gardiner MM, Van der Werf W et al (2008) Increasing corn for biofuel production reduces biocontrol services in agricultural landscapes. Proc Natl Acad Sci USA 105:20552–20557
Landis DA, Wratten SD, Gurr GA (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
Letourneau D (1987) The enemies hypothesis: tritrophic interactions and vegetational diversity in tropical agroecosystems. Ecology 68(6):1616–1622
Letourneau DK, Armbrecht I, Rivera BS et al (2011) Does plant diversity benefit agroecosystems?: a synthetic review. Ecol Appl 21(1):9–21
Lys JA, Nentwig W (1992) Augmentation of beneficial arthropods by strip-management: 4. surface activity, movements and density of abundant carabid beetles in a cereal field. Oecologia 92(3):373–382
Maier CT (1981) Parasitoids emerging from puparia of Rhagoletis pomonella (Diptera: Tephritidae) infesting hawthorn and apple in Connecticut. Can Entomol 113(9):867–870
Murphy BC, Rosenheim JA, Dowell RV et al (1998) Habitat diversification tactic for improving biological control: parasitism of the western grape leafhopper. Entomol Exp Appl 87(3):225–235
Nentwig W (1988) Augmentation of beneficial arthropods by strip management: 1. Succession of predaceous arthropods and long-term change in the ratio of phytophagous and predaceous species in a meadow. Oecologia 76(4):597–606
Nicholls CI, Altieri MA, Vazquez L (2016) Agroecology: principles for the conversion and redesign of farming systems. J Ecosys Ecograph S5:10
Nicholls CI, Parrella MP, Altieri MA (2001) The effects of a vegetational corridor on the abundance and dispersal of insect biodiversity within a northern California organic vineyard. Landsc Ecol 16(2):133–146
Pickett CH, Bugg RL (1998) Enhancing biological control: habitat management to promote natural enemies of agricultural pests. University of California Press, Berkeley
Risch SJ (1981) Insect herbivore abundance in tropical monocultures and polycultures: an experimental test of two hypotheses. Ecology 62(5):1325–1340
Risch SJ, Andow D, Altieri MA (1983) Agroecosystem diversity and pest control: data, tentative conclusions, and new research directions. Environ Entomol 12(3):625–629
Russell EP (1989) Enemies hypothesis: a review of the effect of vegetational diversity on predatory insects and parasitoids. Environ Entomol 18(4):590–599
Thies C, Tscharntke T (1999) Landscape structure and biological control in agroecosystems. Science 285(5429):893–895
Thopham M, Beardsley JW (1975) An influence of nectar source plants on the New Guinea sugarcane weevil parasite, Lixophaga sphenophori (Villeneuve). Proc Hawaii Entomol Soc 22:145–155
Tonhasca A Jr, Byrne DN (1994) The effects of crop diversification on herbivorous insects: a meta-analysis approach. Ecol Entomol 19(3):239–244
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Altieri, M.A., Nicholls, C.I. (2019). Vegetational Designs to Enhance Biological Control of Insect Pests in Agroecosystems. In: Souza, B., Vázquez, L., Marucci, R. (eds) Natural Enemies of Insect Pests in Neotropical Agroecosystems. Springer, Cham. https://doi.org/10.1007/978-3-030-24733-1_1
Download citation
DOI: https://doi.org/10.1007/978-3-030-24733-1_1
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-24732-4
Online ISBN: 978-3-030-24733-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)