
Abstract
Formation of stable soil organic matter is typically the result of a relatively slow series of decomposition processes that can be constrained in early successional sites. Alternatively, compounds such as glomalin, a glycoprotein produced by arbuscular mycorrhizal fungi, may form relatively early during soil development and improve aggregate stabilization, water infiltration, and carbon and nitrogen storage. After 31 years of development, significant amounts of soil C, N, and BRSP, an indicator of glomalin, had accrued in pyroclastic deposits, in patterns affected by both plant community type and soil depth. Mycorrhizal fungi are important, but incompletely understood, drivers of pedogenic processes during primary succession and may exert disproportionate effects on soil processes and plant development prior to the accumulation of humified soil organic matter.
Glossary terms are shown in bold italic face.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Ectomycorrhiza
- Endomycorrhiza
- Glomalin
- Mycorrhizal fungi
- Organic matter
- Soil ecosystem development
- Plant communities
- Soil-plant relationships
- Pyroclastic-flow deposit
5.1 Introduction
Plant and soil community assembly on Mount St. Helens volcanic substrates has been a subject of considerable interest for more than 30 years. Early colonists such as lupines ( Lupinus latifolius and L. lepidus ) were successful because local source populations possessed morphological and physiological adaptations to harsh conditions (Braatne and Bliss 1999), including the ability to fix atmospheric nitrogen via symbiosis with Rhizobium (Halvorson et al. 1992). Organic matter, nutrient concentrations, microbial biomass, and other indicators of soil development increased relatively rapidly under lupines. These factors, together with biotic interactions such as competition with other plants or insect herbivory, abiotic erosive processes, and stochastic environmental drivers like the period of unusually wet conditions recorded for the pyroclastic site from 1995 through 1999, helped shape emerging patterns of plant community structure and soil characteristics (Bishop et al. 2005; Halvorson et al. 2005; del Moral et al. 2012).
One unexpected observation of early studies was evidence for significant accumulations of glomalin in pyroclastic deposits , especially under lupines (Halvorson and Smith 2009). Glomalin, produced by arbuscular mycorrhizal fungi (AMF), is thought to be an important pool of decomposition-resistant organic matter (Wright and Upadhyaya 1996, 1998; Treseder and Turner 2007; Singh 2012; Walley et al. 2013). As a mixture of compounds (Gillespie et al. 2011), glomalin is a glycoproteinaceous material defined operationally by alkaline salt extraction at high temperature and pressure, which can be classified as Bradford-reactive soil protein (BRSP) , glomalin-related soil protein (GRSP) , total glomalin (TG) , easily-extractable glomalin (EEG) , immunoreactive TG (IRTG) , immunoreactive EEG (IREEG) , immunoreactive GRSP , or immunofluorescent material (e.g., Rillig et al. 2003; Lovelock et al. 2004b; Singh 2012; Koide and Peoples 2013; Nichols et al. 2013). Despite issues concerning the exact molecular composition of glomalin, potential co-extractants measured during the protein analysis (Halvorson and Gonzales 2006; Gillespie et al. 2011), and variations in molecular structure with different arbuscular mycorrhizal species or decomposition processes in the soil, glomalin concentration is frequently correlated with water-stable aggregation, mycorrhizal activity, and soil organic C and N concentrations (Nichols and Wright 2004, 2005; Treseder and Turner 2007; Koide and Peoples 2013).
Arbuscular mycorrhizal fungi (AMF) are often found in early primary-successional substrates (Kikvidze et al. 2010), including those deposited by the 1980 eruption of Mount St. Helens (Allen et al. 2005; Titus et al. 2007), but their role in soil ecosystem development and interactions with plants is likely to be complex and variable (Singh et al. 2011; Kardol et al. 2013) and is still incompletely understood (Dickie et al. 2013). Diversion of photosynthate from growth to support the AMF-plant relationship may increase the availability of nutrients to plants (Van Der Heijden and Horton 2009; Neumann and George 2010). Mycorrhizal-plant associations are often associated with phosphorus acquisition or availability (Richardson and Simpson 2011; Turner et al. 2013), but their role in nitrogen acquisition and cycling is increasingly appreciated (Smith and Smith 2011; Veresoglou et al. 2012; Hodge and Storer 2015). The efficiency of the trade-off of plant-derived carbon for mycorrhizally enhanced nutrient access has been used to differentiate whether AMF is a parasite or a mutualist (Johnson et al. 1997), but this distinction is still debated (Smith and Smith 2011). In several instances, AMF associations have produced no growth benefits in early successional species (Titus and del Moral 1998; Sikes et al. 2012).
Less is understood about the nature and strength of feedbacks between AMF, plants, and soil conditions. Fungal community structure has been found to develop along with aboveground vegetation, increasing in diversity with age of the terrain (Cutler et al. 2014) and affecting plant community composition and thus aboveground productivity (Rillig et al. 2014). Other research suggests that fungal activity may respond more to soil conditions than to plant identity (Sikes et al. 2014).
Aside from a paucity of critical plant nutrients like nitrogen, the biggest barriers plants face in pyroclastic substrates are the lack of water and soil organic matter. Glomalin and glomalin-like biomolecules produced by AMF may be instrumental in surmounting these barriers (Wu et al. 2013; Miransari 2014). In addition to boosting nutrient retention by providing organic binding sites (González-Chávez et al. 2002), the roles of these molecules in building soil structure and creating micro- and macro-sites for better water-holding capacity by stimulating aggregate formation may be critical to soil formation in pyroclastic substrates (Rillig and Mummey 2006; Nichols et al. 2013). As a glycoprotein, glomalin is hydrophilic, particularly where the glycosyl groups attach, but amino acid analysis plus water-stable aggregation analysis demonstrates that it has hydrophobic character as well, owing to more hydrophobic amino acids (K. Nichols, personal communication). Glycoproteins commonly exhibit different folding patterns, exposing hydrophilic sides in aqueous solutions and hydrophobic sides for binding and other activities. Glomalin may thus increase water repellency in soils (Wright and Upadhyaya 1998; Rillig 2005; Young et al. 2012). Increased water repellency is often related to aggregate stability and may enhance water and carbon storage in pyroclastic soils and affect mineralization of soil organic matter (Atanassova et al. 2014). Glomalin production may be stimulated in response to chemical, physical, or even biological soil factors (Purin and Rillig 2007). For example, Rillig and Steinberg (2002) observed less total hyphal length but more glomalin produced when roots were grown in small glass beads (<106 μm) simulating less-aggregated soil, compared with those grown in larger glass beads (710–1180 μm) simulating an aggregated soil. Changes in community structure of AMF and production of glomalin were induced as a response to toxic heavy metals like Al, Cr, or Cu, or to salinity (Na) (González-Chávez et al. 2004; Hammer and Rillig 2011; Seguel et al. 2013; Gil-Cardeza et al. 2014; Krishnamoorthy et al. 2014).
The objectives of this study were to gather information about organic matter that has accumulated in Mount St. Helens pyroclastic substrates since the 1980 eruption. This information is consistent with our long-term studies of soil development , and whereas previous studies emphasized simple binary comparisons between lupine-influenced zones and barren substrates, this work better reflects contemporary successional patterns across the landscape by comparing soil characteristics along a gradient of plant community complexity. A further objective was to measure the quantity and distribution of BRSP, hereafter referred to as glomalin, both relative to depth and in association with different plant community types, and to estimate its contribution to total soil carbon and nitrogen pools . We suggest that glomalin forms relatively early during soil development, and that even small amounts of such microbially produced substances may have important effects on soil processes and thus vegetation development prior to the accumulation of humified soil organic matter.
5.2 Materials and Methods
5.2.1 Site Descriptions and Sampling
We collected soil on the Mount St. Helens pyroclastic-flow deposits (Swanson and Major 2005) (46.23°N, 122.16°W) in September 2011 near study sites used extensively as part of a long-term investigation of small-mammal community reassembly on the Pumice Plain (Crisafulli et al. 2005) (Fig. 5.1). We sampled from (1) herb-dominated upland areas that varied in their abundance of bryophytes (mosses), grasses, and forbs, with diversity characterized as low (LDH), intermediate (IDH), or high (HDH), (2) dense shrub-dominated thickets of upland green alder ( Alnus viridis ) (SDU), and (3) riparian areas supporting dense thickets of willow ( Salix sitchensis ) (SDR) (Fig. 5.2a–e). The LDH, IDH, and HDH communities occupy largely intact surfaces created during the eruption and are well represented in the majority of the landscape. The shrub-dominated communities are more limited in their coverage, occupying areas adjacent to stream courses (shrub-dominated riparian, SDR ) or areas subjected to post-eruption erosional processes with lower surface elevations and high topographic moisture (shrub-dominated upland, SDU ). Collectively, these two shrub community types occupy <10% of the Pumice Plain. Litter accumulation in the herb-dominated sites relates to overall plant cover and decreases from the HDH to the LDH community types. Distinct organic layers are present in both of the shrub community types.

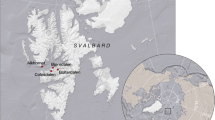
Locations of study sites where soil samples were collected. (Cartography by Kelly Christiansen, USDA Forest Service, Pacific Northwest Research Station).

Examples of (a) a low-diversity upland herb community (LDH) characterized by low species richness and cover values (5–10%) of forbs and grasses and an occasional small-stature shrub; (b) an intermediate-diversity upland herb community (IDH) characterized by moderate species richness and cover values (20–30%) of forbs and grasses and low shrub cover; (c) a high-diversity upland herb community (HDH) characterized by high species richness and cover values (~50%) of forbs and grasses with numerous scattered Salix sitchensis and Alnus viridis shrubs; (d) a shrub-dominated upland thicket (SDU) characterized by mesic upland sites that support dense monotypic stands of A. viridis; and (e) a shrub-dominated riparian thicket (SDR) characterized by wet/mesic seep and streamside sites that support dense growth of S. sitchensis and A. viridis. For both SDU and SDR, canopy height of 2–5 m and cover >70% create heavily shaded understories with low-diversity herb communities and contribute to high litter accumulation. (Photos by C.M. Crisafulli).
We also sampled in two late-seral forest stands unaffected by the 1980 eruption and used these as reference sites to compare with the early primary-successional sites on the Pumice Plain. The reference sites are located in the Pacific silver fir zone (Franklin and Dyrness 1988) about 40 km northeast of Mount St. Helens, at 1120 m (ASL). Dominant overstory species in these stands are Abies amabilis and Tsuga heterophylla , with co-dominant Pseudotsuga menziesii and Thuja plicata . Mean stem density and basal area per hectare are 625 and 145.7 m2, respectively. Understory community dominants are ericaceous shrubs (Vaccinium spp. ) with high cover values (58%), Abies amabilis saplings, and a variety of herbs (e.g., Linnaea borealis , Cornus canadensis , Rubus lasiococcus ).
These reference forest sites were selected because they were of similar age and vegetative structure to forest growing in the Spirit Lake basin (now the Pumice Plain) prior to the 1980 eruption (USDA Forest Service, 1979 unpublished data) and had similar physical site characteristics , such as slope gradient, elevation, and climate. Based on these similarities, we assume that these reference sites supported soils representative of our study area and more broadly similar to late-seral forest of the southern Washington Cascade Range, including the Mount St. Helens vicinity. Forest soils in this region are andisols of volcanic origin (Dahlgren et al. 2004), and a common feature of these soils is their development on top of fresh volcanic surfaces. Indeed, soil pits at our reference forest sites reveal this typical pattern of tephra deposits interleaved with organic soils that developed between volcanic deposition events (see Fig. 11.5 in Allen et al., Chap. 11, this volume). Thus, our reference forest sites provide a relevant comparison to evaluate differences in key soil metrics (i.e., nutrients, organic matter, and glomalin) between undisturbed late-seral forest soils typical of the region and those developing on the Pumice Plain. Finally, the potential natural vegetation of the Pumice Plain is coniferous forest, and we assume that in the absence of additional volcanic activity, both vegetation and soils on our site will eventually converge toward something roughly similar to pre-eruption conditions, although this process could take centuries.
For each plant community , we collected mineral soil with a trowel at three depths (surface to 5, 5–10, and 10–20 cm) from three randomly located pits at two or three study sites. For SDU, we collected samples in nine randomly selected alder thickets. We also sampled the distinct organic layer present on top of the soil for SDU and SDR, but did not consider these samples in our analyses. In the lab, we sieved samples (2 mm sieve), dried them to a constant mass at 55 °C, and stored them at room temperature pending further analysis. Dried samples averaged less than 0.3% H20 by weight. We estimated particle bulk density as the average mass of both “freely settled” (poured) and “tapped” soil needed to fill a container of known volume (25 cm3).
5.2.2 Vegetation Structure
We distinguished vegetation structure at each study site using a modified “foliage height diversity” (FHD) method (MacArthur and MacArthur 1961). This method provides information on plant physiognomy by measuring the vertical distribution of vegetation by life form (see below). At each of our study sites, we arrayed nine transects at 15-m intervals. We fixed transect length at 30 m for LDH, IDH, and HDH sites, but varied the length at the SDR and SDU sites based on the width of those habitats (range 7.9–30 m). We placed FHD measurement stations (n = 66–144) every 2 m along each transect, and at each station we vertically positioned a 3-m graduated PVC pole and assessed the presence (contact with pole) of bryophytes, graminoids, forbs, shrubs, and trees in each of the following height classes: 0–10 cm, 11–25 cm, 26–50 cm, 51–100 cm, 101–150 cm, 151–200 cm, 200–300 cm, and >300 cm. Thus, across sites there were between 582 and 1152 points in space where we assessed vegetation presence. At each site, we calculated the product of the number of FHD stations on each transect and the eight height classes to derive the total number of points sampled for vegetation. Then we summed the number of intercepts with vegetation present (positive hits). We divided the number of positive hits along each transect by the total number of possible points along that transect to calculate the proportion of vegetation present.
5.2.3 Total and Soluble Soil Carbon (C) and Nitrogen (N)
We determined total soil C and N after dry oxidation (Nelson and Sommers 1996) using a FlashEA 1112 NC Analyzer (CE Elantech, Lakewood, New Jersey, USA) with CE Elantech NC reference soil (3.5% C and 0.37% N) and K-factor calibration with aspartic acid as a standard. We extracted water-soluble organic carbon (WSC) and nitrogen (WSN) using a sequential cool- (23 °C) and hot-water (80 °C) extraction procedure and analyzed the solutions with a Shimadzu TOC-VCPN analyzer equipped with a TNM-1 module (Shimadzu Scientific Instruments, Columbia, Maryland, USA) (Halvorson et al. 2013).
5.2.4 Glomalin (Bradford-Reactive Soil Protein)
We extracted subsamples (2 g) from each soil for glomalin with 100 mM tetrasodium pyrophosphate, pH 9.0, for 1-h cycles at 121 °C in an autoclave (Wright et al. 2006). We repeated the 1-h extraction cycle 2–15 (mean = 4) times until the extraction solution was colorless. After each 1-h extraction cycle, we centrifuged samples to pellet the soil and remove extract solution. We combined all extract solutions per subsample and measured the total volumes. We used the Bradford total protein assay as a measure of glomalin, with bovine serum albumin (BSA) as the standard (Bradford 1976; Nichols and Wright 2004). We added an aliquot of 100 mM tetrasodium pyrophosphate to the BSA standard to remove background cross-reactivity with the dye solution. We calculated total protein values as mg Bradford-reactive soil protein (BRSP) g−1 soil.
5.3 Data Analysis
5.3.1 Vegetation Structure
We examined the effect of site and plant community type on the mean proportion of vegetation using a two-way ANOVA. There was no site effect on the proportion of vegetation (P = 0.28), and there was strong evidence that the mean proportion of vegetation differed between at least two of the plant communities (P < 0.001 from a one-way ANOVA F-test). This finding was our justification for pooling all sites for a given community type. We also summarized the mean proportion of vegetation for each life form. We calculated the mean using the proportion of each life form intersecting a transect at eight heights.
5.3.2 Soil
We conducted data analysis using SAS/STAT software, version 9.2 of the SAS System for Windows (SAS Institute 2002–2008). We used a linear mixed model with repeated measures (SAS PROC MIXED, SAS Institute, Cary, North Carolina, USA) to analyze patterns of total and soluble carbon and nitrogen and glomalin, and selected a model that contained both fixed (plant community) and random (sample location) effects, with depth as a repeated measure. We used the KR (Kenward-Roger) option to calculate degrees of freedom and selected covariance structures to minimize Akaike’s information criterion . We performed multiple pairwise comparisons of means using Fisher’s protected least significant difference test using a value of 5% (P < 0.05) as the minimum criterion for significance unless otherwise noted. We evaluated assumptions of normality and identified appropriate data transformations with SAS/ASSIST.
Using SAS PROC CORR, we characterized bivariate relationships between glomalin and the other soil properties by Spearman correlations. Unlike Pearson correlations, Spearman correlations do not assume bivariate normality and are less biased by outliers. Values given in text and graphs are the arithmetic mean ± the standard error of the mean expressed on an oven-dry soil basis.
5.4 Results and Discussion
Early work (through about 2005) was based on a simple binary sampling design comparing lupine-influenced zones to barren substrates. Beginning in 1990, living and dead lupines were delineated from each other and sample depths were standardized (0–5, 5–10 cm). By 2010, it was clear that there was a need to compare the initial herbaceous colonists (lupines) to other nitrogen-fixing species (upland alder) because woody species were becoming increasingly dominant in the landscape. This initial shift in woody-plant abundance has several important potential implications for plant succession (e.g., species turnover), microclimate (e.g., altered light regime, decreased wind, increased water retention), and soil genesis (e.g., heavy litter input, Frankia nitrogen influence). The current sampling approach, adopted in 2011, moved away from simple binary comparisons to multiple comparisons among several sites representative of a gradient of plant community complexity.
5.4.1 Vegetation Structure
At the coarsest level, vegetation analyses revealed an unsurprising dichotomy between structurally simple herb-dominated communities and comparatively complex shrub-dominated communities (Fig. 5.3). A significant difference was apparent between LDH and all other sites, and IDH and HDH were significantly different from both shrub community types, but not from one another. Shrub sites were similar to each other. However, when looking at the mean proportion of different life forms present at the sites, a more complex pattern emerged (Fig. 5.4). Bryophyte values were highest at IDH and HDH sites, lowest at shrub sites, and intermediate at LDH sites (Fig. 5.4a). Graminoids followed a more complex pattern, with the highest value in SDU followed in decreasing proportion from HDH > LDH > SDR > IDH (Fig. 5.4b). Forb values generally increased from LDH to SDU (Fig. 5.4c). All herb-dominated sites had low shrub values, whereas SDR and SDU had a similar mean proportion of shrubs (Fig. 5.4d).

Vegetation structural complexity across five plant community types on the Pumice Plain, Mount St. Helens, USA. Low-diversity herb (LDH), intermediate-diversity herb (IDH), high-diversity herb (HDH), shrub-dominated riparian (SDR), and shrub-dominated upland (SDU). Data points are mean values with error bars representing one standard error.

Mean proportion of vegetation by life form across five plant community types on the Pumice Plain, Mount St. Helens, USA. Data points are mean values with error bars representing one standard error.
The plant communities reported in this chapter are more complex in terms of species richness and physical structure as compared with the lupine communities studied in the first decades after the eruption (Halvorson et al. 1991, 2005; Halvorson and Smith 2009). In many locations on the Pumice Plain, the sequence of plant community development in upland habitats involved L. lepidus establishment, followed by rapid growth and spread of plants in expanding rings around the mother plant and maintenance of a near monoculture for a few to several years. In this regard, lupine communities spread in something analogous to an advancing army front. These young and vigorously growing lupine populations appear to inhibit the establishment of other plant taxa (Morris and Wood 1989) until natural senescence ensues or is hastened by herbivores (Bishop et al. 2005). Once a dieback event occurs, numerous wind-dispersed herbs and graminoids establish, and lupine rarely attains dominance, yet tends to persist as a community member for decades. The upland communities in this study represent different states along this general trajectory.
5.4.2 Soil
Soil bulk density, total and soluble carbon and nitrogen, and glomalin were strongly affected by plant community type, but this effect also depended on sample depth (Table 5.1). In general, differences among the community types were most apparent at the 0–5 cm depth, but there was little distinction among sites at the 10–20 cm depth. Most soil characteristics varied with depth, but these changes were often least evident in the LDH.
5.4.2.1 Bulk Density
Bulk density increased significantly with depth under most community types. The lower values of bulk density near the surface were likely associated with inputs of organic matter from vegetation and were particularly evident at SDR. Conversely, soil bulk density for LDH did not vary with depth and showed less influence of organic inputs at the 0–5 cm depth. Values of average bulk density at the 10–20 cm depth were similar for all locations, 1.42 ± 0.02 Mg m−3.
Surface values of bulk density in the LDH, IDH, and SDU community types were similar to those previously observed for barren pyroclastic sites, whereas those for the HDH and SDR sites were consistent with those found under lupine colonists (Halvorson and Smith 2009). Values of bulk density at the 10–20 cm depth changed little from those reported for young pyroclastic-flow deposits (Wilson and Head 1981).
5.4.2.2 Total Soil C and N
Concentrations of total soil carbon and nitrogen decreased significantly with depth under all community types, especially the SDR. Differences among the community types were most evident at the 0–5 cm depth, with the highest concentrations of C and N in SDR soil and the lowest in the LDH soil. We also observed significant differences among community types for total soil C at the 5–10 cm depth, but not at the 10–20 cm depth (average 1.46 ± 0.26 g kg−1). Total soil nitrogen was less variable with depth and indistinguishable among community types at 5–10 and 10–20 cm, 0.16 ± 0.02 and 0.12 ± 0.02 g kg−1, respectively.
Soil C/N ratios varied with main effects of depth (P < 0.0001) and plant community (P < 0.01). Higher ratios (13.3 ± 0.5) occurred at the surface, compared with 11.6 ± 0.4 and 11.7 ± 0.4 at the 5–10 and 10–20 cm depths, respectively, and ratios were arrayed among the different plant community types in association with larger amounts of organic matter : SDR (15.1 ± 0.4) > SDU (12.5 ± 0.5) ≥ HDH (12.2 ± 0.4) ≥ IDH (10.7 ± 0.4) ≥ LDH (9.4 ± 0.4). In comparison, C/N ratios in the reference forest sites were much higher, 37.5 ± 1.5, 40.7 ± 3.3, and 36.7 ± 3.2 at the 0–5, 5–10, and 10–20 cm depths, respectively. Increasing C/N ratios along the gradient of plant communities from LDH to reference forest sites suggest the accumulation of recalcitrant soil organic carbon, a result of greater inputs from vegetation over time, but may also derive from changes in fungal effects on decomposition during succession. Such changes may be associated with the increasing prevalence of ectomycorrhizal and ericoid mycorrhizal fungal associations, which produce enzymes (Phillips et al. 2014) that may result in greater competition for nitrogen between free-living decomposers and plant hosts. Recently, Averill et al. (2014) demonstrated that soils dominated by plants associated with ectomycorrhizal and ericoid mycorrhizal fungi store 1.7 times more C per unit N than do soils dominated by plants associated with arbuscular mycorrhizal fungi.
The LDH and HDH community types correspond most closely to the barren and lupine (L. lepidus)-influenced sites contrasted in earlier studies and show that organic matter has continued to accrue in developing pyroclastic soils since the eruption in 1980 (Fig. 5.5). From initial values near 0 in barren pyroclastic substrates (Engle 1983), concentrations of total C and N in the 0–5 cm depth of LDH sites increased after 31 years by an average of 81.3 ± 20.9 and 8.1 ± 2.0 mg kg−1 year−1, almost twice as fast as those reported previously for bare sites (Halvorson and Smith 2009). In comparison, by 2011, the concentrations of total C and N increased in HDH sites by an average 183 ± 35 and 14.8 ± 2.9 mg kg−1 year−1, more comparable with earlier estimates under lupines (Halvorson et al. 2005; Halvorson and Smith 2009). The highest accumulations of total C and N in surface soil , equivalent to 665 ± 171 and 37.5 ± 9.1 mg kg−1 year−1, were associated with the SDR community type.

Average values from the current study in relation to historic trends of (a) total soil carbon and (b) soil nitrogen in lupine-influenced (from under live or dead Lupinus lepidus) and barren pyroclastic soil. Pre-2011 values are averages with 95% confidence intervals determined by dry combustion. For barren samples n = 11, 18, 30, 10, 8, and 8 for 1990, 1997, 2000, 2005, 2009, and 2010, respectively. For lupine-influenced samples n = 22, 28, 29, 20, 8, and 8 for 1990, 1997, 2000, 2005, 2009, and 2010, respectively. Data for 1990–2005 are from Halvorson and Smith (2009). Data for 2009 and 2010, from Jonathan Halvorson (unpublished), are from soil samples collected on 1 September 2009 and 21 September 2010 in the same vicinity as the previous studies. Samples were collected, processed, and analyzed using methods and equipment identical to the current study.
Relatively high concentrations of carbon and nitrogen observed in surface samples from SDU and SDR sites undoubtedly relate to significant accumulations of organic matter from leaf litter at these shrub-dominated sites and perhaps to the difficulty in clearly delineating the surface of the mineral soil. Total carbon and nitrogen at both sites were about 380 and 19 g kg−1 in the coarse fractions of the organic layer (>2 mm) and about 200 and 11.5 g kg−1 in the finer fraction (<2 mm), respectively (Jonathan Halvorson, unpublished data).
5.4.2.3 Water-Extractable C and N
Cool-water-extractable C and N decreased with depth, especially in the SDU and SDR community types. The highest concentrations were extracted from the shrub-dominated community types, whereas lower concentrations were extracted from the herb-dominated communities, especially LDH. Differences among community types diminished with depth and were absent at 10–20 cm, averaging 36.5 ± 2.5 and 2.6 ± 0.2 mg kg−1 for extractable C and N, respectively. Hot water extracted two to three times more C and N in patterns similar to cool water; the highest concentrations and greatest variability among community types occurred at the 0–5 cm depth, and shrub-dominated community types had higher concentrations compared to LDH. However, unlike total or cool-water-extractable C and N, higher concentrations of hot-water-extractable C and N were found at the 5–10 and 10–20 cm depths under the shrub-dominated and HDH communities than under the LDH.
The C and N extracted from soil with cool water may correlate to soluble plant residues, whereas C and N recovered after hot-water incubation correlate with soil microbial biomass C and N, mineralizable N , and total carbohydrates (Ghani et al. 2003; Curtin et al. 2006). Both cool- and hot-water-extractable C have been found to be related to soil microbial activity in patterns affected by soil and plants (Uchida et al. 2012). Earlier studies of Mount St. Helens pyroclastic deposits by Halvorson and Smith (2009) reported higher water-extractable C and N values, decreasing with depth, under lupines, as compared with barren sites. More importantly, they found that the amount of hot-water-soluble C was indistinguishable from microbial biomass C measured by substrate-induced respiration. Estimates of soil microbial biomass C at 0–10 cm depth under herb-dominated community types , if still comparable to hot-water-extractable C, were slightly higher than those reported in 2005 and equivalent to about 5% of the total soil C.
5.4.2.4 Glomalin
Average concentrations of glomalin were greatest in the 0–5 cm depth but varied among community types with highest values for SDR and lowest for LDH. Concentrations of glomalin decreased with depth to a similar low value across all sites at the 10–20 cm depth (0.24 g kg−1 soil). Glomalin was strongly and positively correlated with total and soluble soil C and N at each depth (Table 5.2), but negatively correlated with soil bulk density. With the exception of LDH, estimates of glomalin in 0–5 cm depth were within the broad range of values reported for temperate and boreal forests (0.6–5.8 g kg−1) (Singh 2012). However, these values were lower than at two old-growth reference forests located about 40 km to the NE of Mount St. Helens that ranged from 8.4 to 4.0 g kg−1 at 0–5 and 10–20 cm (Table 5.1).
In other studies (Rillig et al. 2003; Emran et al. 2012; Gispert et al. 2013; Vasconcellos et al. 2013), glomalin has been found to be positively correlated with soil C and N but negatively correlated with soil bulk density. Glomalin production is generally higher under conditions that favor AMF activity (Ryan and Graham 2002) or diversity (Helgason et al. 1998), such as minimal physical disruption of the soil (Wright et al. 1999; Borie et al. 2000; Wright and Anderson 2000), higher levels of plant diversity, or low to moderate soil fertility (Treseder and Allen 2002; Lovelock et al. 2004a; Treseder 2004). Changes in glomalin have also been related to the amount of physical disruption of soil or to shifting plant communities (often afforestation) (e.g., Treseder and Turner 2007; Fokom et al. 2012). For example, sites under arable land use showed an exponential decrease in glomalin levels by ~60% after 110 years (Spohn and Giani 2010, 2011). Conversely, glomalin in soil increased with the amount of time under conservation management , including reduced tillage, elimination of fallow periods, use of perennial crops, and greater annual crop diversity (Wright and Anderson 2000; Rillig et al. 2001; Harner et al. 2004; Staddon 2005).
5.4.2.5 Standing Pools of Total C, N, and Glomalin
Standing stocks of organic matter, including glomalin, are the net result of processes that affect its production, such as increased net primary productivity, and those that affect its decomposition, such as soil microbial community structure, moisture, temperature, or disturbance (Treseder and Turner 2007). Stocks of total C, N, and glomalin in the top 20 cm, calculated from data in Table 5.1, varied with plant community type (Fig. 5.6) and were similarly arrayed (SDR ≥ SDU ≥ HDH ≥ IDH ≥ LDH). Values ranged from 1406 ± 238, 84 ± 13, and 241 ± 36 g m−2 at SDR sites for total C, N, and glomalin, respectively, to 358 ± 89, 36 ± 7, and 62 ± 8 g m−2 at LDH sites. Vegetative complexity/diversity data showed no meaningful difference between SDU and SDR (Fig. 5.3), a pattern consistent with total C and N (Fig. 5.6a, b). However, higher glomalin values for SDR sites (Fig. 5.6c) may relate to the age of the plant communities. The SDR sites were among the first to establish (circa late 1980s), whereas the SDU sites initiated later (about 2005). Thus, SDR sites had been accumulating organic matter in comparatively large quantities for 20–25 years. In addition, SDU sites had formerly been colonized by forb and grass species that are slowly waning but still present, whereas in the SDR sites, the transition to shrubs has been underway for decades. The SDU community type was notable for the relatively large pool of total C and N at 10–20 cm depth, not observed in other community types. Increasing C and N with depth suggests the presence of substantial amounts of soil organic matter at depths greater than those examined in this study. Such patterns can result from deeper root distribution under upland shrubs, together with reduced rates of decomposition with depth related to changes in the activity and composition of microbial communities (Gill and Burke 2002; Fierer et al. 2003), and point to the need for a more complete sampling of the soil column in future studies. Similarly, the comparatively large pools of total C, N, and glomalin in the reference forest sites also increased with depth , to about 7455 ± 734, 189 ± 13, and 864 ± 71 g per m−2, respectively.

Average (standard error) total pool sizes of (a) soil carbon, (b) soil nitrogen, and (c) glomalin (BRSP) in mineral soil under different plant community types growing on Mount St. Helens pyroclastic substrates in 2011. Values are depth-weighted and calculated from data in Table 5.1 (n = 6–9). Letters denote differences among community types at the 0–20 cm depth (log-10 transformed data, protected LSD, P ≤ 0.05). Data for reference forest sites (REF) are shown for comparative purposes but not included in the statistical analyses. Note the use of log-10 scales on the y-axes.
5.4.2.6 Glomalin Carbon and Nitrogen
The carbon content of glomalin is known to vary with depth or concentration and to be affected by extraction method, ranging between 27% and 43%, although glomalin nitrogen is more narrowly defined, about 2–4% (Nichols and Wright 2005; Halvorson and Gonzalez 2006). Estimates based on an average composition of 35% C and 4% N showed little variation among community types but demonstrated that glomalin differentially accounted for about 6.2 ± 0.3 < 7.3 ± 0.3 ≤ 8.0 ± 0.5% of the total soil C at 0–5, 5–10, and 10–20 cm depths or 0.52 ± 0.09, 0.13 ± 0.02, and 0.09 ± 0.01 g kg−1, respectively. Glomalin accounted for about 9.6 ± 0.3% (0.028 ± 0.004 g kg−1) of soil N, irrespective of depth . Though affected by the method used to extract glomalin, these numbers are in reasonable agreement with earlier values for Mount St. Helens pyroclastic substrates (Halvorson and Smith 2009) and some other studies (Woignier et al. 2014), but higher than the 3–5% of the total C and N pools typical of temperate and tropical sites (Rillig et al. 2001, 2003; Nichols 2003; Lovelock et al. 2004a; Halvorson and Gonzalez 2006; Jorge-Araújo et al. 2015).
Enrichment of glomalin-C with depth can be attributed to older and more recalcitrant forms of organic matter that may be co-extracted with glomalin (Woignier et al. 2014) but can also be associated with soil disturbance or changes in plant communities. Studies by Spohn and Giani (2010, 2011) found that the ratio of glomalin-related soil protein to total organic carbon increased over time after conversion from forest soils to arable soils. A plot of the proportion of glomalin-C against total soil C (Fig. 5.7a) suggests production or retention of glomalin relative to other pools of soil organic matter may be especially significant when organic matter is low. The concentrations of glomalin observed in surface samples at pyroclastic sites (0–5 cm) accounted for a nearly constant proportion of the total pool of soil C.

Estimated average (standard error) values for (a) glomalin-C as a function of total soil C and (b) glomalin-N as a function of total soil N. Values were derived from measurements of BRSP assuming 35% C and 4% N. Note log-10 scales on the x-axes.
Although different trends were observed among the sites, the proportion of glomalin-N also appeared to decrease as the concentration of total soil N increased to 0.2–0.3 g kg−1, particularly at the 10–20 cm depth. However, there was also evidence for increasingly higher proportions of glomalin-N accompanying higher concentrations of soil N observed in surface (0–5 cm) samples from the pyroclastic sites and the reference forest sites (Fig. 5.7b). Relatively high enrichment of glomalin-N in pyroclastic substrates may reflect strong demands for a limited pool of soil N by plants that induce AMF production (Johnson 2010) and thus the amount of N stabilized in glomalin. Nitrogen incorporated into the structure of glomalin would be stabilized against microbial mineralization but likely still functionally active with respect to glomalin’s effects on soil structure (Rillig et al. 2007). However, although buffered from losses, it is not clear whether the nitrogen in glomalin itself or occluded in glomalin-related aggregates would be accessible to plants. Thus, sequestration of N in the hyphae of symbiotic AMF might remove it from the pool of nutrients readily available to the host plant but could also form the basis of an exclusionary competitive mechanism beneficial to the host by limiting access to nutrients by competitors.
5.5 Conclusions
Through their effects on plant physiology, soil ecological interactions, and soil engineering, mycorrhizal fungi are important but incompletely understood drivers of pedogenic processes during primary succession (e.g., Rillig and Steinberg 2002; Rodriguez et al. 2009; Torres et al. 2012). Formation of stable soil organic matter is typically the result of a relatively slow series of decomposition processes that can be constrained, in early successional sites, by the availability of exogenous inputs or by environmental conditions that limit microbial activities. Alternatively, compounds such as glomalin, a glycoprotein produced by arbuscular mycorrhizal fungi, may form relatively early during soil development and improve aggregate stabilization, water infiltration, and carbon and nitrogen storage.
The presence of glomalin in Mount St. Helens pyroclastic substrates is further evidence of the positive feedbacks between plants, microorganisms, and soils thought to play a central role in early successional communities . Our data suggest that C, N, and glomalin concentrations increase together with vegetation complexity, and in this sense there appears to be a coupling between plant community development and soil genesis during early primary succession although the two may not synchronize. Although establishment of woody plants (shrubs) is an ecological threshold for plant community change and development, changes in soil genesis likely lag behind vegetative community structure. Even small amounts of microbially produced substances like glomalin may have disproportionate effects on soil processes and plant development prior to the accumulation of humified soil organic matter. However, the details of these effects, together with a more complete understanding of the evolving roles of fungi during primary succession, remain to be elucidated.
References
Allen, M.F., C.M. Crisafulli, S.J. Morris, L.M. Egerton-Warburton, J.A. MacMahon, and J.M. Trappe. 2005. Mycorrhizae and Mount St. Helens: Story of a symbiosis. In Ecological responses to the 1980 eruption of Mount St. Helens, ed. V.H. Dale, F.J. Swanson, and C.M. Crisafulli, 221–231. New York: Springer.
Atanassova, I.D., S.H. Doerr, and G.L. Mills. 2014. Hot-water-soluble organic compounds related to hydrophobicity in sandy soils. In Soil carbon, ed. A.E. Hartemink and K. McSweeney, 137–146. Switzerland: Springer International Publishing.
Averill, C., B.L. Turner, and A.C. Finzi. 2014. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 505: 543–545.
Bishop, J.G., W.F. Fagan, J.D. Schade, and C.M. Crisafulli. 2005. Causes and consequences of herbivory on prairie lupine (Lupinus lepidus) in early primary succession. In Ecological responses to the 1980 eruption of Mount St. Helens, ed. V.H. Dale, F.J. Swanson, and C.M. Crisafulli, 151–161. New York: Springer.
Borie, F.R., R. Rubio, A. Morales, and C. Castillo. 2000. Relationships between arbuscular mycorrhizal hyphal density and glomalin production with physical and chemical characteristics of soils under no-tillage. Revista Chilena de Historia Natural 73: 749–756.
Braatne, J.H., and L.C. Bliss. 1999. Comparative physiological ecology of lupines colonizing early successional habitats on Mount St. Helens. Ecology 80: 891–907.
Bradford, M.M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248–254.
Crisafulli, C.M., J.A. MacMahon, and R.R. Parmenter. 2005. Small-mammal survival and colonization on the Mount St. Helens volcano: 1980–2002. In Ecological responses to the 1980 eruption of Mount St. Helens, ed. V.H. Dale, F.J. Swanson, and C.M. Crisafulli, 199–218. New York: Springer.
Curtin, D., C.E. Wright, M.H. Beare, and F.M. McCallum. 2006. Hot water-extractable nitrogen as an indicator of soil nitrogen availability. Soil Science Society of America Journal 70: 1512–1521.
Cutler, N.A., D.L. Chaput, and C.J. van der Gast. 2014. Long-term changes in soil microbial communities during primary succession. Soil Biology and Biochemistry 69: 359–370.
Dahlgren, R.A., M. Saigusa, and F.C. Ugolini. 2004. The nature, properties and management of volcanic soils. In Advances in Agronomy No. 82, ed. D. Sparks, 113–182. Amsterdam: Academic Press, Elsevier.
del Moral, R., L.A. Thomason, A.C. Wenke, N. Lozanoff, and M.D. Abata. 2012. Primary succession trajectories on pumice at Mount St. Helens, Washington. Journal of Vegetation Science 23: 73–85.
Dickie, I.A., L.B. Martínez-García, N. Koele, G.A. Grelet, J.M. Tylianakis, D.A. Peltzer, and S.J. Richardson. 2013. Mycorrhizas and mycorrhizal fungal communities throughout ecosystem development. Plant and Soil 367: 11–39.
Emran, M., M. Gispert, and G. Pardini. 2012. Patterns of soil organic carbon, glomalin and structural stability in abandoned Mediterranean terraced lands. European Journal of Soil Science 63: 637–649.
Engle, M. 1983. Carbon, nitrogen and microbial colonization of volcanic debris on Mount St. Helens. Environmental science and regional planning. Pullman: Washington State University.
Fierer, N., J.P. Schimel, and P.A. Holden. 2003. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry 35: 167–176.
Fokom, R., S. Adamou, M.C. Teugwa, A.D. Begoude Boyogueno, W.L. Nana, M.E.L. Ngonkeu, N.S. Tchameni, D. Nwaga, G. Tsala Ndzomo, and P.H. Amvam Zollo. 2012. Glomalin related soil protein, carbon, nitrogen and soil aggregate stability as affected by land use variation in the humid forest zone of South Cameroon. Soil and Tillage Research 120: 69–75.
Franklin, J.F., and C.T. Dyrness. 1988. Natural vegetation of Oregon and Washington. Corvallis: Oregon State University Press.
Ghani, A., M. Dexter, and K.W. Perrott. 2003. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biology and Biochemistry 35: 1231–1243.
Gil-Cardeza, M.L., A. Ferri, P. Cornejo, and E. Gomez. 2014. Distribution of chromium species in a Cr-polluted soil: Presence of Cr(III) in glomalin related protein fraction. Science of the Total Environment 493: 828–833.
Gill, R.A., and I.C. Burke. 2002. Influence of soil depth on the decomposition of Bouteloua gracilis roots in the shortgrass steppe. Plant and Soil 241: 233–242.
Gillespie, A.W., R.E. Farrell, F.L. Walley, A.R.S. Ross, P. Leinweber, K.-U. Eckhardt, T.Z. Regier, and R.I.R. Blyth. 2011. Glomalin-related soil protein contains non-mycorrhizal-related heat-stable proteins, lipids and humic materials. Soil Biology and Biochemistry 43: 766–777.
Gispert, M., M. Emran, G. Pardini, S. Doni, and B. Ceccanti. 2013. The impact of land management and abandonment on soil enzymatic activity, glomalin content and aggregate stability. Geoderma 202: 51–61.
González-Chávez, C., J. D’Haen, J. Vangronsveld, and J. Dodd. 2002. Copper sorption and accumulation by the extraradical mycelium of different Glomus spp. (arbuscular mycorrhizal fungi) isolated from the same polluted soil. Plant and Soil 240: 287–297.
González-Chávez, M.C., R. Carrillo-González, S.F. Wright, and K.A. Nichols. 2004. The role of glomalin, a protein produced by arbuscular mycorrhizal fungi, in sequestering potentially toxic elements. Environmental Pollution 130: 317–323.
Halvorson, J.J., and J.M. Gonzalez. 2006. Bradford reactive soil protein in Appalachian soils: Distribution and response to incubation, extraction reagent and tannins. Plant and Soil 286: 339–356.
Halvorson, J., and J. Smith. 2009. Carbon and nitrogen accumulation and microbial activity in Mount St. Helens pyroclastic substrates after 25 years. Plant and Soil 315: 211–228.
Halvorson, J.J., J.L. Smith, and E.H. Franz. 1991. Lupine influence on soil carbon, nitrogen and microbial activity in developing ecosystems at Mount St. Helens. Oecologia 87: 162–170.
Halvorson, J.J., E.H. Franz, J.L. Smith, and R.A. Black. 1992. Nitrogenase activity, nitrogen fixation and nitrogen inputs by lupines at Mount St. Helens. Ecology 73: 87–98.
Halvorson, J.J., J.L. Smith, and A.C. Kennedy. 2005. Lupine effects on soil development and function during early primary succession at Mount St. Helens. In Ecological responses to the 1980 eruption of Mount St. Helens, ed. V.H. Dale, F.J. Swanson, and C.M. Crisafulli, 243–254. New York: Springer.
Halvorson, J.J., J.M. Gonzalez, and A.E. Hagerman. 2013. Retention of tannin-C is associated with decreased soluble nitrogen and increased cation exchange capacity in a broad range of soils. Soil Science Society of America Journal 77: 1199–1213.
Hammer, E.C., and M.C. Rillig. 2011. The influence of different stresses on glomalin levels in an arbuscular mycorrhizal fungus—Salinity increases glomalin content. PLoS One 6: e28426.
Harner, M.J., P.W. Ramsey, and M.C. Rillig. 2004. Protein accumulation and distribution in floodplain soils and river foam. Ecology Letters 7: 829–836.
Helgason, T., T.J. Daniell, R. Husband, A.H. Fitter, and J.P.W. Young. 1998. Ploughing up the wood-wide web? Nature 394: 431.
Hodge, A., and K. Storer. 2015. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant and Soil 386: 1–19.
Johnson, N.C. 2010. Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytologist 185: 631–647.
Johnson, N.C., J.H. Graham, and F.A. Smith. 1997. Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytologist 135: 575–585.
Jorge-Araújo, P., H. Quiquampoix, P.T. Matumoto-Pintro, and S. Staunton. 2015. Glomalin-related soil protein in French temperate forest soils: Interference in the Bradford assay caused by co-extracted humic substances. European Journal of Soil Science 66: 311–319.
Kardol, P., G.B. De Deyn, E. Laliberté, P. Mariotte, and C.V. Hawkes. 2013. Biotic plant–soil feedbacks across temporal scales. Journal of Ecology 101: 309–315.
Kikvidze, Z., C. Armas, K. Fukuda, L. Martínez-García, M. Miyata, A. Oda-Tanaka, F. Pugnaire, and B. Wu. 2010. The role of arbuscular mycorrhizae in primary succession: Differences and similarities across habitats. Web Ecology 10: 50–57.
Koide, R.T., and M.S. Peoples. 2013. Behavior of Bradford-reactive substances is consistent with predictions for glomalin. Applied Soil Ecology 63: 8–14.
Krishnamoorthy, R., K. Kim, C. Kim, and T. Sa. 2014. Changes of arbuscular mycorrhizal traits and community structure with respect to soil salinity in a coastal reclamation land. Soil Biology and Biochemistry 72: 1–10.
Lovelock, C.E., S.F. Wright, D.A. Clark, and R.W. Ruess. 2004a. Soil stocks of glomalin produced by arbuscular mycorrhizal fungi across a tropical rain forest landscape. Journal of Ecology 92: 278–287.
Lovelock, C.E., S.F. Wright, and K.A. Nichols. 2004b. Using glomalin as an indicator for arbuscular mycorrhizal hyphal growth: An example from a tropical rain forest soil. Soil Biology & Biochemistry 36: 1009–1012.
MacArthur, R.H., and J.W. MacArthur. 1961. On bird species diversity. Ecology 42: 594–598.
Miransari, M. 2014. Mycorrhizal fungi to alleviate salinity stress on plant growth. In Use of microbes for the alleviation of soil stresses, ed. M. Miransari, 77–86. New York: Springer.
Morris, W.F., and D.M. Wood. 1989. The role of lupine in succession on Mount St. Helens: Facilitation or inhibition? Ecology 70: 697–703.
Nelson, D.W., and L.E. Sommers. 1996. Total carbon, organic carbon and organic matter. In Methods of soil analysis part 3: Chemical methods. No 5. in the Soil Science Society of America Books Series, ed. D.L. Sparks, A.L. Page, P.A. Helmke, R.H. Loeppert, P.N. Soltanpour, M.A. Tabatabai, C.T. Johnson, and M.E. Sumner, 961–1010. Madison: Soil Science Society of America.
Neumann, E., and E. George. 2010. Nutrient uptake: The arbuscular mycorrhiza fungal symbiosis as a plant nutrient acquisition strategy. In Arbuscular mycorrhizas: Physiology and function, ed. H. Koltai and Y. Kapulnik, 137–167. Amsterdam: Springer.
Nichols, K.A. 2003. Characterization of glomalin: A glycoprotein produced by arbuscular mycorrhizal fungi. Ph.D. dissertation. College Park: University of Maryland.
Nichols, K.A., and S.F. Wright. 2004. Contributions of soil fungi to organic matter in agricultural soils. In Functions and management of soil organic matter in agro-ecosystems, ed. F. Magdoff and R. Weil, 179–198. Washington, DC: CRC.
Nichols, K., and S. Wright. 2005. Comparison of glomalin and humic acid in eight native US soils. Soil Science 170: 985–997.
Nichols, K.A., J.J. Halvorson, and T. Caesar. 2013. Roles of biology, chemistry, and physics in soil macroaggregate formation and stabilization. Open Agriculture Journal 7: 107–117.
Phillips, L.A., V. Ward, and M.D. Jones. 2014. Ectomycorrhizal fungi contribute to soil organic matter cycling in sub-boreal forests. ISME Journal 8: 699–713.
Purin, S., and M.C. Rillig. 2007. The arbuscular mycorrhizal fungal protein glomalin: Limitations, progress, and a new hypothesis for its function. Pedobiologia 51: 123–130.
Richardson, A.E., and R.J. Simpson. 2011. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology 156: 989–996.
Rillig, M.C. 2005. A connection between fungal hydrophobins and soil water repellency? Pedobiologia 49: 395–399.
Rillig, M.C., and D.L. Mummey. 2006. Mycorrhizas and soil structure. New Phytologist 171: 41–53.
Rillig, M.C., and P.D. Steinberg. 2002. Glomalin production by an arbuscular mycorrhizal fungus: A mechanism of habitat modification? Soil Biology and Biochemistry 34: 1371–1374.
Rillig, M.C., S.F. Wright, K.A. Nichols, W.F. Schmidt, and M.S. Torn. 2001. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant and Soil 233: 167–177.
Rillig, M.C., P.W. Ramsey, S. Morris, and E.A. Paul. 2003. Glomalin, an arbuscular-mycorrhizal fungal soil protein, responds to land-use change. Plant and Soil 253: 293–299.
Rillig, M., B. Caldwell, H.B. Wösten, and P. Sollins. 2007. Role of proteins in soil carbon and nitrogen storage: Controls on persistence. Biogeochemistry 85: 25–44.
Rillig, M.C., S. Wendt, J. Antonovics, S. Hempel, J. Kohler, J. Wehner, and T. Caruso. 2014. Interactive effects of root endophytes and arbuscular mycorrhizal fungi on an experimental plant community. Oecologia 174: 263–270.
Rodriguez, R.J., J.F. White Jr., A.E. Arnold, and R.S. Redman. 2009. Fungal endophytes: Diversity and functional roles. New Phytologist 182: 314–330.
Ryan, M.H., and J.H. Graham. 2002. Is there a role for arbuscular mycorrhizal fungi in production agriculture? Plant and Soil 244: 263–271.
SAS Institute. 2002–2008. SAS/STAT user’s guide, version 9.2. Cary: SAS Institute.
Seguel, A., J. Cumming, K. Klugh-Stewart, P. Cornejo, and F. Borie. 2013. The role of arbuscular mycorrhizas in decreasing aluminium phytotoxicity in acidic soils: A review. Mycorrhiza 23: 167–183.
Sikes, B.A., H. Maherali, and J.N. Klironomos. 2012. Arbuscular mycorrhizal fungal communities change among three stages of primary sand dune succession but do not alter plant growth. Oikos 121: 1791–1800.
Sikes, B., H. Maherali, and J. Klironomos. 2014. Mycorrhizal fungal growth responds to soil characteristics, but not host plant identity, during a primary lacustrine dune succession. Mycorrhiza 24: 219–226.
Singh, L.P., S.S. Gill, and N. Tuteja. 2011. Unraveling the role of fungal symbionts in plant abiotic stress tolerance. Plant Signaling & Behavior 6: 175–191.
Singh, P.K. 2012. Role of glomalin related soil protein produced by arbuscular mycorrhizal fungi: A review. Agricultural Science Research Journal 2: 119–125.
Smith, S.E., and F.A. Smith. 2011. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annual Review of Plant Biology 62: 227–250.
Spohn, M., and L. Giani. 2010. Water-stable aggregates, glomalin-related soil protein, and carbohydrates in a chronosequence of sandy hydromorphic soils. Soil Biology and Biochemistry 42: 1505–1511.
———. 2011. Impacts of land use change on soil aggregation and aggregate stabilizing compounds as dependent on time. Soil Biology and Biochemistry 43: 1081–1088.
Staddon, P.L. 2005. Mycorrhizal fungi and environmental change: The need for a mycocentric approach. New Phytologist 167: 635–637.
Swanson, F.J., and J.J. Major. 2005. Physical events, environments, and geological-ecological interactions at Mount St. Helens: March 1980–2004. In Ecological responses to the 1980 eruption of Mount St. Helens, ed. V.H. Dale, F.J. Swanson, and C.M. Crisafulli, 27–43. New York: Springer.
Titus, J., and R. del Moral. 1998. The role of mycorrhizal fungi and microsites in primary succession on Mount St. Helens. American Journal of Botany 85: 370–375.
Titus, J.H., S. Whitcomb, and H.J. Pitoniak. 2007. Distribution of arbuscular mycorrhizae in relation to microsites on primary successional substrates on Mount St. Helens. Canadian Journal of Botany 85: 941–948.
Torres, M.S., J.F. White Jr., X. Zhang, D.M. Hinton, and C.W. Bacon. 2012. Endophyte-mediated adjustments in host morphology and physiology and effects on host fitness traits in grasses. Fungal Ecology 5: 322–330.
Treseder, K.K. 2004. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytologist 164: 347–355.
Treseder, K.K., and M.F. Allen. 2002. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: A model and field test. New Phytologist 155: 507–515.
Treseder, K.K., and K.M. Turner. 2007. Glomalin in ecosystems. Soil Science Society of America Journal 71: 1257–1266.
Turner, B., H. Lambers, L. Condron, M. Cramer, J. Leake, A. Richardson, and S. Smith. 2013. Soil microbial biomass and the fate of phosphorus during long-term ecosystem development. Plant and Soil 367: 225–234.
Uchida, Y., S. Nishimura, and H. Akiyama. 2012. The relationship of water-soluble carbon and hot-water-soluble carbon with soil respiration in agricultural fields. Agriculture, Ecosystems & Environment 156: 116–122.
Van Der Heijden, M.G.A., and T.R. Horton. 2009. Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. Journal of Ecology 97: 1139–1150.
Vasconcellos, R.L., J.A. Bonfim, D. Baretta, and E.J. Cardoso. 2013. Arbuscular mycorrhizal fungi and glomalin-related protein as indicators of soil quality in a recuperation gradient of the Atlantic forest in Brazil. Land Degradation & Development 27: 325–334.
Veresoglou, S.D., B. Chen, and M.C. Rillig. 2012. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biology and Biochemistry 46: 53–62.
Walley, F.L., A.W. Gillespie, A.B. Adetona, J.J. Germida, and R.E. Farrell. 2013. Manipulation of rhizosphere organisms to enhance glomalin production and C sequestration: Pitfalls and promises. Canadian Journal of Plant Science 94: 1025–1032.
Wilson, L., and J.W. Head. 1981. Morphology and rheology of pyroclastic flows and their deposits, and guidelines for future observations. In The 1980 eruptions of Mount St. Helens, Washington. Professional Paper 1250, ed. P.W. Lipman and D.R. Mullineaux, 513–524. Washington, DC: U.S. Geological Survey.
Woignier, T., P. Etcheverria, F. Borie, H. Quiquampoix, and S. Staunton. 2014. Role of allophanes in the accumulation of glomalin-related soil protein in tropical soils (Martinique, French West Indies). European Journal of Soil Science 65: 531–538.
Wright, S.F., and R.L. Anderson. 2000. Aggregate stability and glomalin in alternative crop rotations for the central Great Plains. Biology and Fertility of Soils 31: 249–253.
Wright, S.F., and A. Upadhyaya. 1996. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Science 161: 575–586.
———. 1998. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant and Soil 198: 97–107.
Wright, S.F., J.L. Starr, and I.C. Paltineanu. 1999. Changes in aggregate stability and concentration of glomalin during tillage management transition. Soil Science Society of America Journal 63: 1825–1829.
Wright, S.F., K.A. Nichols, and W.F. Schmidt. 2006. Comparison of efficacy of three extractants to solubilize glomalin on hyphae and in soil. Chemosphere 64: 1219–1224.
Wu, Q.-S., A.K. Srivastava, and Y.-N. Zou. 2013. AMF-induced tolerance to drought stress in citrus: A review. Scientia Horticulturae 164: 77–87.
Young, I.M., D.S. Feeney, A.G. O’Donnell, and K.W.T. Goulding. 2012. Fungi in century old managed soils could hold key to the development of soil water repellency. Soil Biology and Biochemistry 45: 125–127.
Acknowledgments
The authors thank T. Blackman, E. Denton, J. Harrah, T. Robertson A. Smits, C. Thorson, and D. Wetch for sample collection and technical assistance. Photo credits: C.M. Crisafulli. Funding for CMC was provided by the USDA Forest Service, Pacific Northwest Research Station, and the National Science Foundation (LTREB Program DEB-0614538). This work is dedicated to Jeffrey L. Smith.
Disclaimer
The US Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and, where applicable, sex, marital status, family status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program (not all prohibited bases apply to all programs). USDA is an equal opportunity provider and employer. Mention of commercial products and organizations in this manuscript is solely to provide specific information. It does not constitute endorsement by USDA-ARS over other products and organizations not mentioned.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Glossary
- Arbuscular mycorrhizal fungi
-
A plant-fungal relationship, found in over 90% of vascular plants, in which the fungus penetrates the cortical cells of the roots and forms highly branched arbuscules and, in some species, vesicles. These fungi provide nutrients and water to plants as well as assisting in soil structure and disease prevention.
- Ectomycorrhizal fungi
-
A plant-fungal relationship, found in about 2% of plant species, particularly woody plants, in which the fungus, primarily basidiomycetes and some ascomycetes, forms a dense hyphal sheath, known as the mantle, around the root surface. The hyphal network may extend several centimeters into the surrounding soil to aid in water and nutrient uptake, often helping the host plant to survive adverse conditions, in exchange for carbohydrates.
- Ericoid mycorrhizal fungi
-
A plant-fungal relationship between members of the plant family Ericaceae, such as blueberries, cranberries, and Rhododendron, and several lineages of fungi in acidic and nutrient poor soils found in boreal forests, bogs, and heathlands. The fungus establishes loose hyphal networks around the outside of hair roots, from which they penetrate the walls of cortical cells to form intracellular coils that can densely pack individual plant cells. The coils function only for a period of a few weeks before the plant cell and fungal hyphae begin to degrade. The coils are where nutrients are exchanged for carbohydrates, and these fungi also have enzymatic capabilities to break down complex organic molecules and act as saprophytes, living on dead or decaying organic matter.
- Mycorrhiza(e)
-
A symbiotic association between a fungus and the roots of a host plant where the body of the fungi are thread-like hyphae that extend the root system. The mycorrhizal association is generally mutualistic, playing important roles in soil biology, chemistry, and physics, but in particular species or in particular circumstances, mycorrhizae may be variously pathogenic in host plants.
- Photosynthate
-
A chemical product of photosynthesis—carbohydrates that are synthesized from carbon dioxide and a source of hydrogen (usually water), using light as an energy source.
Rights and permissions
Copyright information
© 2018 Springer Science+Business Media LLC
About this chapter

Cite this chapter
Halvorson, J.J., Nichols, K.A., Crisafulli, C.M. (2018). Soil Carbon and Nitrogen and Evidence for Formation of Glomalin, a Recalcitrant Pool of Soil Organic Matter, in Developing Mount St. Helens Pyroclastic Substrates. In: Crisafulli, C., Dale, V. (eds) Ecological Responses at Mount St. Helens: Revisited 35 years after the 1980 Eruption. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-7451-1_5
Download citation
DOI: https://doi.org/10.1007/978-1-4939-7451-1_5
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-7449-8
Online ISBN: 978-1-4939-7451-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)