
Abstract
The European Environment Agency (The Weybridge + 15 (1996–2011) report. EEA Technical report, vol 2. Copenhagen, 2012) and the United Nations Environment programme together with the World Health Organisation (State of the science of endocrine disrupting chemicals-2012. Geneva, Switzerland) both recently published major and highly authoritative reviews of endocrine disrupting chemicals in the natural environment and their effects on reproduction and health in both humans and wildlife. One surprising conclusion to emerge from these reviews was that there are relatively few well documented reports of endocrine disruption (ED) in wild mammals, mainly because much of the available evidence is correlative and does not conclusively demonstrate that the chemicals in question cause the physiological and phenotypic problems attributed to them. However, based on strong evidence from studies of wild birds, reptiles, invertebrates, and laboratory animals, it is difficult to imagine that wild mammals would be the exception. This chapter is therefore included to emphasize the point that the role of reproductive science within wildlife conservation is much broader than a narrow focus on artificial breeding technologies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The last decade has seen an increase in evidence for relationships between exposure to certain man-made chemicals and endocrine disruption in wildlife UNEP/WHO (2012); of particular concern are flame retardants, organochlorine pesticides and polychlorinated biphenyls (PCBs), which are a class of compounds now banned due to their harmful effects but historically used in various industrial applications. A range of toxic persistent organic pollutants (POPs), such as PCBs, polychlorinated dibenzop-dioxins (PCDDs), polychlorinated dibenzofurans (PCDFs), dichlorodiphenyldichloroethylene (p,p′-DDE) and polybrominated diphenyl ethers (PBDEs), continue to contaminate the environment due to past and present human activities. Many of the endocrine disrupting effects reported in wildlife can be reproduced in laboratory studies where animals are experimentally exposed to EDCs, adding support to the hypothesis that exposure to these chemicals is associated with adverse health effects. Here we briefly describe some of the recent comparative research findings, which are of interest to reproductive biologists in the context of a changing environment. These chemicals represent another form of environmental stress, adding to the stresses that wildlife already experience because of human population growth, habitat degradation and climate change.
In addition to the chemicals mentioned above, attention is being increasingly paid to the effects on reproduction of the ever growing number of antimicrobial compounds used in both human and veterinary medicine. Between 1992 and 1999, over 700 antibacterial products, the majority of which contained triclosan (TCS) entered the consumer market. A major review of TCS, which described its structure, occurrence, degradation products and both toxicity and endocrine disrupting effects (Dann and Hontela 2011) illustrated the biological background of the compound and provided figures to show that, in Sweden alone, about 2 t are used in personal care products and toothpaste and that global production of TCS has now exceeded 1,500 t per year. A considerable amount of this material eventually finds its way into sewage, where it exerts endocrine disrupting effects on algae and other aquatic invertebrates. Experimentally it also exerts endocrine disrupting effects, both estrogenic and androgenic, on fish (Pinto et al. 2013), although fortunately at doses that are usually higher than those typically found in the environment. Although TCS is noted for its toxicity, endocrine disrupting effects mediated via its resemblance to thyroid hormones have recently been described in a Pacific tree frog, Pseudacris regilla, (Marlatt et al. 2013) and endocrine disrupting effects in mice have been attributed to its estrogenic action (Crawford and de Catanzaro 2012). These examples highlight the growing realisation that pharmaceutical compounds, as well as industrial chemicals, are becoming more prevalent in the environment. Fortunately regulatory authorities are becoming more aware of the attendant problems and need for vigilance, and a new field of “ecopharmacovigilance” has recently emerged (Holm et al. 2013). Authorities concerned with reproductive aspects of human clinical medicine have also recently voiced their concern. For example, the American College of Obstetricians and Gynaecologists and the American Society for Reproductive Medicine have issued a joint Committee Opinion (ACOG/ASRM 2013) advocating for government policy changes to identify and reduce exposure to toxic environmental agents. It included the following statement: “Every pregnant woman in America is exposed to many different chemicals in the environment; prenatal exposure to certain chemicals is linked to miscarriages, stillbirths, and birth defects. Many chemicals that pregnant women absorb or ingest from the environment can cross the placenta to the fetus. Exposure to mercury during pregnancy, for instance, is known to harm cognitive development in children. Toxic chemicals in the environment harm our ability to reproduce, negatively affect pregnancies, and are associated with numerous other long-term health problems.”
Understanding of the effects of EDCs on animal development and reproduction can only be achieved by increasing our knowledge of basic reproductive biology. Determining how compounds exert their influence across the whole spectrum of life on the planet requires comparative knowledge of developmental and reproductive mechanisms in everything from marine invertebrates around coastal waters to large terrestrial mammals. Indeed, as these species are intimately linked to each other within complex food webs, there is a myriad of ways in which these man-made chemicals and their products can influence reproductive processes.
2 Overview of Comparative Effects
Research into the effects of environmental chemical exposure in both laboratory and wild species has been ongoing for decades, but was brought into sharp focus by observations in the 1960s that organochlorine pesticides, mainly dichlorodiphenyltrichloroethane (DDT), caused eggshell thinning and reduced reproductive success in birds (for review, see Hellou et al. 2013). For example, organochlorines endangered the populations of a top predator, the peregrine falcon (Falco peregrinus) in the northern hemisphere and led to its extinction in the most heavily impacted areas of North America and Europe between the 1950s and 1970s. Similar observations (Hernandez et al. 2008) have been made in other top predators, including the Spanish Imperial Eagle, an iconic but threatened Iberian species. The reasons behind these effects, especially pronounced in top predators through bioaccumulation from dietary exposure, included disruption to calcium metabolism, neurotoxicity and behavioural changes. This type of research ultimately prompted a ban on the use of DDT in North America and Europe, leading to a reduction in body burdens in birds, an improvement in eggshell thickness, and the subsequent recovery of many of the affected populations. Recent studies have shown encouraging evidence of long-term declines in organochlorine concentrations within the eggs of these birds and correlative increases in shell thickness (Vorkamp et al. 2009; Falk et al. 2006).
Nevertheless, high concentrations of pesticides have occasionally been released by accident into the environment, with disastrous consequences. When industrial pesticides (DDT and its metabolites DDD and DDE, plus the acaricide dicofol; p, p′-dichlorodiphenyl-2,2,2,-trichloroethanol) were accidentally spilled into a tributary of Lake Apopka in Florida, USA in 1980, dramatic declines were observed in juvenile recruitment in the resident American alligator (Alligator mississippiensis) population. Abnormal ovarian morphology, large numbers of polyovular follicles and polynuclear oocytes were reported in female alligators. Reduced phallus size and altered plasma testosterone concentrations also were seen in males. Plasma estradiol concentrations were almost double those in female alligators from a reference lake (Guillette and Moore 2006; Guillette et al. 1996). Contaminant exposure was regarded as the most likely explanation for the abnormalities observed. Although unfortunate for the American alligators affected by the chemical spillage, the event stimulated considerable interest in the response of reptiles to endocrine disruption because of the plasticity and variety of their sex determination mechanisms (Sarre et al. 2004, 2011). The turtle, Trachemys scripta elegans, has subsequently been proposed as a laboratory model for assaying the estrogenic effects of exogenous chemicals (Gale et al. 2002) because the species depends on both environmental temperature and the hormonal environment for sex determination. Although this is an interesting idea, regulatory authorities may dispute its effectiveness because endocrine disruption might be confounded by the effects of temperature.
It is, however, important to recognise the conflict that occurs when chemicals are banned and the original purposes of these chemicals are still required. For example, DDT has had a major impact on the control of insect borne diseases, such as malaria, across the world, but unless suitable alternatives are available, the diseases will continue to ravage human populations. In fact, the apparent absence of suitable alternatives has resulted in the reintroduction of DDT in South Africa for use in controlling mosquito populations near Johannesburg. This step, which involves enforced spraying within the houses of the local human populations had had devastating reproductive effects on the humans themselves, i.e. reduced sperm quality and external urogenital birth defects, and the local wildlife (Bornman and Bouwman 2012; de Jager et al. 2002), with the reappearance of disorders related to abnormal sexual differentiation.
In contrast to DDT and PCBs, some other EDCs are increasing, or at least not showing signs of decrease, in the environment (e.g. perfluorinated alkyl compounds and replacements for banned brominated flame retardants). Perfluoroalkyl acids (PFAAs), a sub-class of fluorochemicals with fully fluorinated carbon chains, have gained increasing attention as an emerging category of pollutants (Jensen and Leffers 2008; Joensen et al. 2009). As such, they have become the target for risk evaluation and reduced production. These synthetic fluorinated organic chemicals have extensive industrial applications, including as surfactants and emulsifiers often in the production of other fluorinated chemicals, as well as grease and stain-repellents, friction reducers (wiring, computers), water-proofing and insulating agents, and in fire extinguishing foam (Benskin et al. 2012). Recent studies (White et al. 2011) have shown that these compounds are found globally in human tissues, including human milk and human cord blood from individuals in North America, China, and various European countries. White et al. (2011) also pointed out the probable interactions between the presence of chemicals such as PFAAs during embryonic and postnatal development in mammals, and the influence of the maternal intrauterine environment on the health of adults later in life. Galatius et al. (2011) studied temporal trends in harbour porpoises from the Danish North Sea collected between 1980 and 2005, and found no evidence of a population decline. This is encouraging, but it is nevertheless likely that more sensitive studies might find evidence of correlations between adult fitness in porpoises and early life exposure to endocrine disrupters.
There is growing evidence for long-term interactions between the foetal environment and adult health, i.e. that the original sources of adult disease can often be attributed to what happened “in utero” or even earlier during gamete development (Thornburg et al. 2010). The relationships between the intrauterine environment and the embryo during the early life of mammals and the onset of adult diseases, such as cardiovascular disease, hypertension and diabetes were initially identified from epidemiological observations on human populations, but have since been confirmed in experimental mammals. These observations, often collectively known as the Barker hypothesis (Barker 1995), repeatedly show that if embryos undergo different forms of “stress” during early development, children are likely to be underweight at birth and will then show phenotypic symptoms of disease as they develop into adults. Most attention has been paid to dietary stress, where the embryo initially seems to adapt its metabolic functions to make the most of the limited resources available. If conditions improve later in life, this individual cannot cope with the better lifestyle and tends to become obese and a develop a suite of late onset diseases (Barker et al. 2010; Barker 1995). Logically the presence of EDCs within the foetal environment, embryo, newborn or juveniles and the female reproductive tract could exert additional stress or have important influences on embryonic growth and development in wild species. In support of this hypothesis we can point to recent evidence from a Swedish human population showing that prenatal exposure to PCBs was associated with higher birth weight, and PBDE exposure with lowered birth weights (Lignell et al. 2013). Although these effects are complex and difficult to clarify, such human population studies suggest that wild species, whose body burden of such chemicals is often higher, will also be affected; extensive and detailed data on contaminant concentrations in arctic wild species, including mammals, birds and fishes, reported by Letcher et al. (2010) lend support to this idea. In fact, these authors commented that evidence of defective neurological development in some polar bears might be attributed to such long-term effects, but they could not be sure because of the difficulties involved in obtaining relevant data.
This information becomes more pertinent when it is considered that chemical analyses of several large mammals (seals, porpoises, whales and polar bears) have demonstrated high body burdens of hydrophobic contaminants, such as PCBs and brominated flame retardants (for review, see Sonne 2010). Typically, these species acquire pollutants via their diet and bioaccumulate EDCs, which tend to be lipophilic compounds, within their body fat so that the concentrations increase together with increasing age. When females begin to suckle their offspring, their milk is enriched with EDCs, and the EDCs are transferred to their newborns, with probable effects on survival and reproductive development (Hall et al. 2009). Surprisingly, however, it has proven difficult to demonstrate that PCBs and other lipophilic compounds actually cause impaired reproductive development (Letcher et al. 2010), although population and pharmacokinetic modelling studies of East Greenland polar bears, backed up by experimental studies of Greenland sledge dogs, predict the imminent occurrence of negative population impacts (Sonne 2010). Evidence for reduced female reproductive efficacy has also been found in studies of the European harbour porpoise and short-beaked common dolphin (Munson et al. 1998). In harbour porpoises, high persistent organic pollutant burdens tended to be associated with few ovarian scars, suggesting that high contaminant levels may be inhibiting ovulation; however, the significance of ovarian scars has recently been re-evaluated (Dabin et al. 2008). Because there was no evidence of an age-related increase, the authors cast some doubt on the usefulness of this parameter. Several species of pinnipeds have experienced recent population declines; Alaskan populations of northern fur seal, the Galápagos sea lion, Zalophus wollebaeki (Alava et al. 2009) and the Steller sea lion (Eumetopias jubatus) (Trites and Donnelly 2003) have all suffered from reduced pupping rates. In the fur seal these reproductive failures are suspected to be associated with bioaccumulation of environmental contaminants in maternal body tissues (Fillman et al. 2007).
Many studies have been published on the potential impacts of contaminants on thyroid function in various large marine mammals. Schnitzler et al. (2008) studied thyroid histology in relation to trace metals (Cd, Fe, Zn, Cu, Se, and Hg) and showed that there were largely negative relationships between concentrations of cadmium, selenium and copper and thyroid fibrosis. They concluded that there was insufficient evidence from their study to support the hypothesis that these elements have adverse effects on thyroid function. Nevertheless, the European Environment Agency report drew attention to several reports that individual contaminants (including PCBs, dieldrin and chlordane) negatively affect thyroid function in seals (Routti et al. 2008), sea lions (Debier et al. 2005), beluga whales in the St Lawrence estuary (Deguise et al. 1995) and polar bears (Braathen et al. 2004). The relationships between thyroid function and reproduction are complex; they interact with other components of the endocrine system, are involved in growth and bone formation, and contaminants with ED activity will undoubtedly exert a broad range of physiological effects.
Wild amphibians are known to be sensitive to water-borne endocrine disruption because of their highly permeable integument and the possibility of exposure during critical periods of development (embryonic and larval). Owing to the importance of thyroid function during amphibian metamorphosis there is a growing body of work relating to thyroid disrupting chemicals, including Organization for Economic Co-operation and Development (OECD) methodology for detecting EDCs (Pickford 2010). Like similarly affected fish species (see section below), amphibians showing gonadal intersex, feminisation of secondary sexual characteristics and altered sex hormone concentrations have been observed at sites contaminated by agricultural pesticides across Italy, South Africa, parts of Florida, Ontario and Michigan (Carr and Patino 2011; Norris 2011; Papoulias et al. 2013). The exact causes are still regarded as uncertain, but extensive research efforts have been invested in the use of amphibian species as laboratory models for studying and detecting endocrine disruption in these species (Miyata and Ose 2012; Olmstead et al. 2012).
2.1 Endocrine Disruption in Fish
Searching Web of Science (in August 2013) with the terms “fish + endocrine +disruption” resulted in the retrieval of 1,147 scientific papers, and refining the search using the additional term “reproduction” found 301 papers. The oldest papers related to reproduction dated back to 1995, thus demonstrating how much this particular field has advanced in less than two decades. Within this short review it is impossible to cite all of the available work in this taxa, so we will summarise by focusing on major reviews; 70 in total, including recent ones by McNair et al. (2012), Soffker and Tyler (2012) and Waye and Trudeau (2011). The European Environment Agency report (2012) also provides a useful and succinct summary of this area.
Endocrine disruption in fish is clearly widespread; the best studied example is that of feminised male roach (Rutilus rutilus), a cyprinid (carp) fish in United Kingdom rivers (Rodgers-Gray et al. 2001), and a second example, the gudgeon (Gobio gobio) was later also identified (van Aerle et al. 2001). These males exhibited abnormal reproductive characteristics associated with exposure to effluents from sewage treatment works, including elevated plasma vitellogenin (a female-specific egg laying protein) concentrations, and many had eggs developing in their testes (intersex) or feminised sperm ducts. Following a nationwide survey, these effects were attributed to natural and synthetic estrogens in the sewage effluent (Jobling et al. 2006). Nonylphenol was also identified as an important EDC in some locations, and eventually an effect map that related the incidence of sexually disrupted fish to estrogenic activities in more than 2,000 sewage effluent outlet locations was constructed (Williams et al. 2009). The map showed that 39 % of the modelled river reaches in the UK were predicted not to be at risk from ED, and most of the remainder were predicted to be at risk (with 1–3 % were predicted to be at high risk).
Feminised fish have also been found in other European countries, such as Denmark, France, Italy, Germany and the Netherlands. Again, these were associated with sewage effluent outlets. Studies in North America support the European findings to a certain extent. A study of 16 species in nine river basins found that only 3 % of the fish, from four species examined at 111 sites, exhibited intersex (Hinck et al. 2009).
Although feminisation has now been widely reported in aquatic systems, the converse effect of female masculinisation has also been described both experimentally and in field studies. A study of Eastern mosquitofish (Gambusia holbrooki) in the St John river, Florida (Bortone and Cody 1999) found that females showed significant elongation of the anal fin and the gonopodium, an anal fin that is modified into an intromittent organ in males of the poecilidae, such as this. A similar study in China that focused on another mosquitofish (Gambusia affinis) (Hou et al. 2011) found evidence of masculinising effects on the anal fin but also detected increased testis mass. Interestingly, these study sites both received effluents from a local paper mill. Paper mill effluents have since been implicated as causing female masculinization at other sites (Deaton and Cureton 2011), such as changes in female mating behaviour, altered offspring sex ratios, diminished body size in masculinized females and lower fecundity (for review, see Soffker and Tyler 2012). A laboratory study in which both of these species were exposed to the androgen agonist 17-β trenbolone (TB) (Brockmeier et al. 2013) detected masculinising effects after exposure to 0.1, 1 or 10 μg TB/L, including a series of gene activation effects in the tip of the anal fin. Although Kovacs et al. (2013) concluded that chemical mixtures in paper mill effluents were too complex to understand in terms of physiological effects, most evidence strongly suggests that they do contain steroid analogues that directly interfere with sexual differentiation.
Endocrine disruption has also been reported in marine fish: Kirby et al. (2004) detected intersex and elevated vitellogenin concentrations in flounder (Platichthys flesus) in many coastal regions, especially estuaries receiving effluents from industrial and domestic sources. Similar effects have been noted in various marine species, for example bigeye tuna (Thunnus obesus) around the coast of Japan (Hashimoto et al. 2003), killifish (Fundulus heteroclitus) (Bugel et al. 2010) in Newark bay, New Jersey in the USA and marine top predators in the Mediterranean sea (Fossi et al. 2007).
Understanding precisely which causative agents are responsible for feminisation in fish presents something of a puzzle. 17β-Ethinyl estradiol (EE2), which is a component of the human contraceptive pill, has been studied extensively and is known to possess powerful estrogenic action. Although efficient water purification systems are used widely to extract it before water is recycled back into rivers, detailed studies have shown that even some advanced water purification systems do not eliminate all oestrogenic activities (Baynes et al. 2012). Many other chemicals possess endocrine disrupting actions. Apart from the pollutants mentioned above, these include nonylphenol, octylphenol ethoxylate surfactants, bisphenol A, phthalates, phyto-oestrogens and endogenous estrogens excreted from women. Moreover, work by Jobling et al. (2009) has suggested that estrogenic compounds (such as steroid hormones) in association with anti-androgenic activity (measured by in vitro techniques, and as yet of undetermined source) are statistically correlated to intersex induction in the UK. This illustrates the complexity of the problem and highlights the difficulties involved in attempting mitigation strategies.
3 Endocrine Disruption in Invertebrates
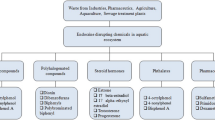
Aquatic invertebrates are key parts of food webs that underpin the life of all other species and are highly abundant in the world’s oceans. For example, the density of a polychaete (Nereis diversicolor) living around coastal and estuarine habitats has been recorded as 3,700 m3 (Scaps 2002). Their importance in marine ecology has been outlined by Lawrence and Soame (2009), who pointed out that as suspension feeders in fjords, populations filter the whole water mass up to three times per day and reduce the phytoplankton biomass by 50 % in less than 5 h. This species and others are therefore in a prime position for exposure to pollutants with varying effects. Their evolutionary histories are, however, very different and because they have developed different endocrinological signalling systems, their responses to pollutants are diverse and not necessarily predictable. Endocrine disruption is therefore a distinct possibility in some species, while others seem more tolerant to known vertebrate EDCs. The whole field is so diverse that it is impossible to provide detailed information about all classes of invertebrates; however, some of the main effects that have been described are summarised in Table 4.1.
4 What Should Be Done?
The title of this subsection is unashamedly copied from a review by the late Dr Stuart Rhind (Rhind 2009) who presented it by invitation at a symposium of the Zoological Society of London in 2009 organized by one of the present authors (WVH). Apart from providing an excellent overview of the entire field, Dr Rhind memorably discussed an experiment in which sheep grazed on land that had been fertilized twice yearly using sewage sludge were compared with sheep that were grazed on untreated grass. Analyses showed that soil levels of contaminants such as phthalate and alkyl phenol PCB and PBDE were initially very low and were increased only minimally by the sewage treatment (Rhind et al. 2002). Nevertheless, when the reproductive performance of the experimentally exposed sheep was investigated, it was found that the testes of their fetuses contained fewer Leydig and Sertoli cells than the controls, coupled with lower blood concentrations of the hormones testosterone and inhibin (Paul et al. 2005). There were also fewer oocytes in the fetal ovaries (Fowler et al. 2008) and an altered balance of pro- and anti-apoptotic proteins towards apoptosis. This remarkable outcome can be regarded as a “real world” effect that probably applies not only to grazing domestic sheep but also many other terrestrial species, especially those whose habitats are likely to have suffered any form of airborne or waterborne pollution. Subtleties such as the reduction of oocyte and Sertoli cell production (which would both result in lowered gamete production) by mammalian fetuses are likely to be undetectable in wild and threatened species, because, by definition, these species are not intensely studied. Nevertheless, the outcome of such effects will ultimately be reflected in lowered fertility, an undesirable outcome under the circumstances.
As discussed elsewhere in this book, however, the way in which different species are affected cannot necessarily be predicted, given the huge diversity of reproductive mechanisms that have evolved to cope with different, and often very adverse, conditions. Improving our understanding of comparative reproductive mechanisms is therefore as essential in this, as it is in related fields. The outcomes of many field observations, especially those involving complex mixtures of chemicals, underline the crudeness of our understanding of mixtures, and the way in which they affect reproductive mechanisms. This is understandable because experimental laboratory scientists typically prefer to make sure they understand the variables in their treatments. Although regulatory initiatives such as that introduced in 2006 by the European Union, namely Registration, Evaluation, Authorization and Restriction of chemical substances (REACH), will provide basic toxicity data on the all chemicals produced in Europe or imported into Europe in amounts that exceed 100 t per annum, the enormous number of chemicals that REACH is expected to evaluate (143,000 were pre-registered with REACH in 2008) will preclude all but the most limited of testing regimes. In fact, under the REACH protocols all substances are only tested once. This is a massive undertaking and it is interesting to see that the policy itself has been criticised because of the extensive need for animal testing (Hartung and Rovida 2009); these authors suggested that 54 million vertebrate animals would be used under REACH and that the costs would be around €9.5 billion.
One conclusion to be drawn is that there is a pressing need for the further development of reliable tests that can be used in vitro to assess the toxicity of chemicals, thereby avoiding animal use. Some authors such as Schrattenholz et al. (2012) have considered that multifactorial systems biology may be useful for this purpose because of the possibility of integrating data across transcriptomics, proteomics, epigenomics and metabolomics. Others such as Lee et al. (2012) have proposed the use of whole embryo culture and mouse embryonic stem cells as alternative models for the study of developmental toxicology. Focusing on species of most ecological relevance has led some authors, such as Scholz et al. (2013), to concentrate on fish and amphibian cells for toxicity testing, while others have applied the same principle to the evaluation of chemicals that would be particularly relevant in terms of marine species such as corals (Shafir et al. 2003: Howe et al. 2012).
The studies cited in this short chapter underline and emphasise the vast amount of work that has been carried out over the past few decades, and it is apparent that although international regulatory authorities are now taking note of the need to prevent some of the worst chemicals from reaching the environment, the problems are global, multifactorial and difficult.
References
ACOG, ASRM. Exposure to toxic environmental agents. Fertil Steril. 2013. doi: 10.1016/j.fertnstert.2013.08.043.
Alava JJ, Ikonomou MG, Ross PS, Costa D, Salazar S, Aurioles-Gamboa D, Gobas F. Polychlorinated biphenyls and polybrominated diphenyl ethers in Galapagos sea lions (Zalophus wollebaeki). Environ Toxicol Chem. 2009;28(11):2271–82. doi:10.1897/08-331.s1.
Armoza-Zvuloni R, Kramarsky-Winter E, Rosenfeld H, Shore LS, Segal R, Sharon D, Loya Y. Reproductive characteristics and steroid levels in the scleractinian coral Oculina patagonica inhabiting contaminated sites along the Israeli Mediterranean coast. Mar Pollut Bull. 2012;64(8):1556–63. doi:10.1016/j.marpolbul.2012.05.020.
Barker DJ, Gelow J, Thornburg K, Osmond C, Kajantie E, Eriksson JG. The early origins of chronic heart failure: impaired placental growth and initiation of insulin resistance in childhood. Eur J Heart Fail. 2010;12(8):819–25. hfq069 [pii] 10.1093/eurjhf/hfq069.
Barker DJP. Fetal origins of coronary heart disease. BMJ. 1995;311(6998):171–4.
Baynes A, Green C, Nicol E, Beresford N, Kanda R, Henshaw A, Churchley J, Jobling S. Additional treatment of wastewater reduces endocrine disruption in wild fish-a comparative study of tertiary and advanced treatments. Environ Sci Technol. 2012;46(10):5565–73. doi:10.1021/es204590d.
Benskin JP, Li B, Ikonomou MG, Grace JR, Li LY. Per- and polyfluoroalkyl substances in landfill leachate: patterns, time trends, and sources. Environ Sci Technol. 2012;46(21):11532–40. doi:10.1021/es302471n.
Bornman MS, Bouwman H. Environmental pollutants and diseases of sexual development in humans and wildlife in South Africa: harbingers of impact on overall health? Reprod Domest Anim. 2012;47:327–32. doi:10.1111/j.1439-0531.2012.02094.x.
Bortone SA, Cody RP. Morphological masculinization in poeciliid females from a paper mill effluent receiving tributary of the St. Johns River, Florida, USA. B Environ Contam Tox. 1999; 63(2):150–6.
Braathen M, Derocher AE, Wiig O, Sormo EG, Lie E, Skaare JU, Jenssen BM. Relationships between PCBs and thyroid hormones and retinol in female and male polar bears. Environ Health Perspect. 2004;112(8):826–33. doi:10.1289/ehp.6809.
Brockmeier EK, Ogino Y, Iguchi T, Barber DS, Denslow ND. Effects of 17 beta-trenbolone on Eastern and Western mosquitofish (Gambusia holbrooki and G affinis) anal fin growth and gene expression patterns. Aquat Toxicol. 2013;128:163–70. doi:10.1016/j.aquatox.2012.12.007.
Bugel SM, White LA, Cooper KR. Impaired reproductive health of killifish (Fundulus heteroclitus) inhabiting Newark Bay, NJ, a chronically contaminated estuary. Aquat Toxicol. 2010;96(3):182–93. doi:10.1016/j.aquatox.2009.10.016.
Carr JA, Patino R. The hypothalamus-pituitary-thyroid axis in teleosts and amphibians: endocrine disruption and its consequences to natural populations. Gen Comp Endocrinol. 2011;170(2):299–312. doi:10.1016/j.ygcen.2010.06.001.
Chesman BS, Langston WJ. Intersex in the clam Scrobicularia plana: a sign of endocrine disruption in estuaries? Biol Lett. 2006;2(3):420–2. doi:10.1098/rsbl.2006.0482.
Crawford BR, de Catanzaro D. Disruption of blastocyst implantation by triclosan in mice: impacts of repeated and acute doses and combination with bisphenol-A. Reprod Toxicol. 2012;34(4):607–13. doi:10.1016/j.reprotox.2012.09.008.
Dabin W, Cossais F, Pierce G, Ridoux V. Do ovarian scars persist with age in all Cetaceans: new insight from the short-beaked common dolphin (Delphinus delphis Linnaeus, 1758). Mar Biol. 2008;156(2):127–39. doi:10.1007/s00227-008-1070-4.
Dann AB, Hontela A. Triclosan: environmental exposure, toxicity and mechanisms of action. J Appl Toxicol. 2011;31(4):285–311. doi:10.1002/jat.1660.
de Jager C, Bornman MS, Bailey JL. Male reproductive health and DDT: sufficient evidence to discontinue its use? Proceeding of 9th international symposium on spermatology. Capetown, South Africa. Monduzzi Editore, Bologna. 2002;191–6.
Deaton R, Cureton JC. Female masculinization and reproductive life history in the western mosquitofish (Gambusia affinis). Environ Biol Fish. 2011;92(4):551–8. doi:10.1007/s10641-011-9878-z.
Debier C, Ylitalo GM, Weise M, Gulland F, Costa DP, Le Boeuf BJ, de Tillesse T, Larondelle Y. PCBs and DDT in the serum of juvenile California sea lions: associations with vitamins A and E and thyroid hormones. Environ Pollut. 2005;134(2):323–32. doi:10.1016/j.envpol. 2004.07.012.
Deguise S, Martineau D, Beland P, Fournier M. Possible mechanisms of action of environmental contaminants on St Lawrence Beluga whales (Delphinapterus leucas). Environ Health Perspect. 1995;103:73–7.
Dinan L, Bourne P, Whiting P, Dhadialla TS, Hutchinson TH. Screening of environmental contaminants for ecdysteroid agonist and antagonist activity using the Drosophila melanogaster B(II) cell in vitro assay. Environ Toxicol Chem. 2001a;20(9):2038–46.
Dinan L, Bourne PC, Meng Y, Sarker SD, Tolentino RB, Whiting P. Assessment of natural products in the Drosophila melanogaster B(II) cell bioassay for ecdysteroid agonist and antagonist activities. Cell Mol Life Sci. 2001b;58(2):321–42.
European Environment Agency. The impact of endocrine disrupters on wildlife, people and their environments. The Weybridge +15 (1996–2011) report. EEA technical report, vol 2. Copenhagen; 2012.
Falk K, Moller S, Mattox WG. A long-term increase in eggshell thickness of Greenlandic Peregrine Falcons Falco peregrinus tundrius. Sci Total Environ. 2006;355(1–3):127–34. doi:10.1016/j.scitotenv.2005.02.024.
Fillmann G, Hermanns L, Fileman TW, Readman JW. Accumulation patterns of organochlorines in juveniles of Arctocephalus australis found stranded along the coast of Southern Brazil. Environ Pollut. 2007;146(1):262–7. doi:10.1016/j.envpol.2006.02.034.
Forbes VE, Warbritton R, Aufderheide J, Van Der Hoeven N, Caspers N. Effects of bisphenol a on fecundity, egg hatchability, and juvenile growth of Marisa cornuarietis. Environ Toxicol Chem. 2008;27(11):2332–40. doi:10.1897/07-622.1.
Fossi MC, Casini S, Marsill L. Potential toxicological hazard due to endocrine-disrupting chemicals on Mediterranean top predators: state of art, gender differences and methodological tools. Environ Res. 2007;104(1):174–82. doi:10.1016/j.envres.2006.06.014.
Fowler PA, Dora NJ, McFerran H, Amezaga MR, Miller DW, Lea RG, Cash P, McNeilly AS, Evans NP, Cotinot C, Sharpe RM, Rhind SM. In utero exposure to low doses of environmental pollutants disrupts fetal ovarian development in sheep. Mol Hum Reprod. 2008;14(5):269–80. doi:10.1093/molehr/gan020.
Galatius A, Dietz R, Riget FF, Sonne C, Kinze CC, Lockyer C, Bossi R. Temporal and life history related trends of perfluorochemicals in harbor porpoises from the Danish North Sea. Mar Poll Bull. 2011;62(7):1476–83. doi:10.1016/j.marpolbul.2011.04.011.
Gale RW, Bergeron JM, Willingham EJ, Crews D. Turtle sex determination assay: mass balance and responses to 2,3,7,8-tetrachlorodibenzo-p-dioxin and 3,3',4,4',5-pentachlorobiphenyl. Environ Toxicol Chem. 2002;21(11):2477–82.
Gomes T, Gonzalez-Rey M, Bebianno MJ. Incidence of intersex in male clams Scrobicularia plana in the Guadiana Estuary (Portugal). Ecotoxicology. 2009;18(8):1104–9. doi:10.1007/s10646-009-0359-5.
Guillette Jr LJ, Moore BC. Environmental contaminants, fertility, and multioocytic follicles: a lesson from wildlife? Semin Reprod Med. 2006;24(3):134–41. doi:10.1055/s-2006-944419.
Guillette Jr LJ, Pickford DB, Crain DA, Rooney AA, Percival HF. Reduction in penis size and plasma testosterone concentrations in juvenile alligators living in a contaminated environment. Gen Comp Endocrinol. 1996;101(1):32–42.
Hall AJ, Thomas GO, McConnell BJ. Exposure to persistent organic pollutants and first-year survival probability in gray seal pups. Environ Sci Technol. 2009;43(16):6364–9. doi:10.1021/es9004398.
Hartung T, Rovida C. Chemical regulators have overreached. Nature. 2009;460(7259):1080–1. doi:10.1038/4601080a.
Hashimoto S, Kurihara R, Strussmann CA, Yamasaki T, Soyano K, Hara A, Shiraishi H, Morita M. Gonadal histology and serum vitellogenin levels of bigeye tuna Thunnus obesus from the Northern Pacific Ocean–absence of endocrine disruption bio-indicators. Mar Poll Bull. 2003;46(4):459–65. doi:10.1016/S0025-326X(02)00460-5.
Hellou J, Lebeuf M, Rudi M. Review on DDT and metabolites in birds and mammals of aquatic ecosystems. Environ Rev. 2013;21(1):53–69. doi:10.1139/er-2012-0054.
Hernandez M, Gonzalez LM, Oria J, Sanchez R, Arroyo B. Influence of contamination by organochlorine pesticides and polychlorinated biphenyls on the breeding of the Spanish Imperial Eagle (Aquila adalberti). Environ Toxicol Chem. 2008;27(2):433–41. doi:10.1897/07-308r.1.
Hinck JE, Blazer VS, Schmitt CJ, Papoulias DM, Tillitt DE. Widespread occurrence of intersex in black basses (Micropterus spp.) from U.S. rivers, 1995–2004. Aquat Toxicol. 2009;95(1):60–70. doi:10.1016/j.aquatox.2009.08.001.
Holm G, Snape JR, Murray-Smith R, Talbot J, Taylor D, Sorme P. Implementing ecopharmacovigilance in practice: challenges and potential opportunities. Drug Saf. 2013;36(7):533–46. doi:10.1007/s40264-013-0049-3.
Horiguchi T. Masculinization of female gastropod mollusks induced by organotin compounds, focusing on mechanism of actions of tributyltin and triphenyltin for development of imposex. Environ Sci. 2006;13(2):77–87.
Hou LP, Xie YP, Ying GG, Fang ZQ. Developmental and reproductive characteristics of western mosquitofish (Gambusia affinis) exposed to paper mill effluent in the Dengcun River, Sihui, South China. Aquat Toxicol. 2011;103(3–4):140–9. doi:10.1016/j.aquatox.2011.02.018.
Howe PL, Reichelt-Brushett AJ, Clark MW. Aiptasia pulchella: a tropical cnidarian representative for laboratory ecotoxicological research. Environ Toxicol Chem. 2012;31(11):2653–62. doi:10.1002/etc.1993.
Jensen AA, Leffers H. Emerging endocrine disrupters: perfluoroalkylated substances. Int J Androl. 2008;31(2):161–9. doi:10.1111/j.1365-2605.2008.00870.x.
Jobling S, Casey D, Rogers-Gray T, Oehlmann J, Schulte-Oehlmann U, Pawlowski S, Baunbeck T, Turner AP, Tyler CR. Comparative responses of molluscs and fish to environmental estrogens and an estrogenic effluent. Aquat Toxicol. 2004;66(2):207–22.
Jobling S, Williams R, Johnson A, Taylor A, Gross-Sorokin M, Nolan M, Tyler CR, van Aerle R, Santos E, Brighty G. Predicted exposures to steroid estrogens in U.K. rivers correlate with widespread sexual disruption in wild fish populations. Environ Health Perspect. 2006;114 Suppl 1:32–9.
Jobling S, Burn RW, Thorpe K, Williams R, Tyler C. Statistical modeling suggests that antiandrogens in effluents from wastewater treatment works contribute to widespread sexual disruption in fish living in english rivers. Environ Health Perspect. 2009;117(5):797–802. doi:10.1289/ehp.0800197.
Joensen UN, Bossi R, Leffers H, Jensen AA, Skakkebaek NE, Jorgensen N. Do perfluoroalkyl compounds impair human semen quality? Environ Health Perspect. 2009;117(6):923–7. doi:10.1289/ehp.0800517.
Keay J, Thornton JW. Hormone-activated estrogen receptors in annelid invertebrates: implications for evolution and endocrine disruption. Endocrinology. 2009;150(4):1731–8. doi:10.1210/en.2008-1338.
Kirby MF, Allen YT, Dyer RA, Feist SW, Katsiadaki I, Matthiessen P, Scott AP, Smith A, Stentiford GD, Thain JE, Thomas KV, Tolhurst L, Waldock MJ. Surveys of plasma vitellogenin and intersex in male flounder (Platichthys flesus) as measures of endocrine disruption by estrogenic contamination in United Kingdom estuaries: temporal trends, 1996 to 2001. Environ Toxicol Chem. 2004;23(3):748–58. doi:10.1897/03-166.
Kovacs TG, Martel PH, O’Connor BI, Hewitt LM, Parrott JL, McMaster ME, MacLatchy DL, Van der Kraak GJ, Van den Heuvel MR. A survey of Canadian mechanical pulp and paper mill effluents: insights concerning the potential to affect fish reproduction. J Environ Sci Health A Tox Hazard Subst Environ Eng. 2013;48(10):1178–89. doi:10.1080/10934529.2013.776440.
Lawrence AJ, Soame JM. The endocrine control of reproduction in Nereidae: a new multi-hormonal model with implications for their functional role in a changing environment. Phil Trans Roy Soc Lond B Biol Sci. 2009;364(1534):3363–76. doi:10.1098/rstb.2009.0127.
Lee HY, Inselman AL, Kanungo J, Hansen DK. Alternative models in developmental toxicology. Syst Biol Reprod Med. 2012;58(1):10–22. doi:10.3109/19396368.2011.648302.
Letcher RJ, Bustnes JO, Dietz R, Jenssen BM, Jorgensen EH, Sonne C, Verreault J, Vijayan MM, Gabrielsen GW. Exposure and effects assessment of persistent organohalogen contaminants in arctic wildlife and fish. Sci Total Environ. 2010;408(15):2995–3043. doi:10.1016/j.scitotenv. 2009.10.038.
Lignell S, Aune M, Darnerud PO, Hanberg A, Larsson SC, Glynn A. Prenatal exposure to polychlorinated biphenyls (PCBs) and polybrominated diphenyl ethers (PBDEs) may influence birth weight among infants in a Swedish cohort with background exposure: a cross-sectional study. Environ Health. 2013;12. doi:44 10.1186/1476-069x-12-44.
Marlatt VL, Veldhoen N, Lo BP, Bakker D, Rehaume V, Vallee K, Haberl M, Shang DY, van Aggelen GC, Skirrow RC, Elphick JR, Helbing CC. Triclosan exposure alters postembryonic development in a Pacific tree frog (Pseudacris regilla) Amphibian Metamorphosis Assay (TREEMA). Aquat Toxicol. 2013;126:85–94. doi:10.1016/j.aquatox.2012.10.010.
Matozzo V, Gagne F, Marin MG, Ricciardi F, Blaise C. Vitellogenin as a biomarker of exposure to estrogenic compounds in aquatic invertebrates: a review. Environ Int. 2008;34(4):531–45. doi:10.1016/j.envint.2007.09.008.
McNair A, Lim JN, Nakagawa S. The fitness consequences of environmental sex reversal in fish: a quantitative review. Biol Rev. 2012;87(4):900–11. doi:10.1111/j.1469-185X.2012.00230.x.
Miyata K, Ose K. Thyroid hormone-disrupting effects and the amphibian metamorphosis assay. J Toxicol Pathol. 2012;25(1):1–9. doi:10.1293/tox.25.1.
Munson L, Calzada N, Kennedy S, Sorensen TB. Luteinized ovarian cysts in Mediterranean striped dolphins. J Wildl Dis. 1998;34(3):656–60.
Norris DO. Endocrine disruption of reproduction in amphibians. Hormones and reproduction of vertebrates, Vol 2: Amphibians. In: Norris DO, Lopez KH, editors. London: Academic Press. 2011;2:203–211.
Oehlmann J, Schulte-Oehlmann U, Bachmann J, Oetken M, Lutz I, Kloas W, Ternes TA. Bisphenol A induces superfeminization in the ramshorn snail Marisa cornuarietis(Gastropoda: Prosobranchia) at environmentally relevant concentrations. Environ Health Perspect. 2006;114 Suppl 1:127–33.
Olmstead AW, Kosian PA, Johnson RN, Blackshear PE, Haselman J, Blanksma C, Korte JJ, Holcombe GW, Burgess E, Lindberg-Livingston A, Bennett BA, Woodis KK, Degitz SJ. Trenbolone causes mortality and altered sexual differentiation in Xenopus tropicalis during larval development. Environ Toxicol Chem. 2012;31(10):2391–8. doi:10.1002/etc.1965.
Papoulias DM, Schwarz MS, Mena L. Gonadal abnormalities in frogs (Lithobates spp.) collected from managed wetlands in an agricultural region of Nebraska, USA. Environ Pollut. 2013;172:1–8. doi:10.1016/j.envpol.2012.07.042.
Pascoal S, Carvalho G, Vasieva O, Hughes R, Cossins A, Fang Y, Ashelford K, Olohan L, Barroso C, Mendo S, Creer S. Transcriptomics and in vivo tests reveal novel mechanisms underlying endocrine disruption in an ecological sentinel, Nucella lapillus. Mol Ecol. 2013;22(6):1589–608. doi:10.1111/mec.12137.
Paul C, Rhind SM, Kyle CE, Scott H, McKinnell C, Sharpe RM. Cellular and hormonal disruption of fetal testis development in sheep reared on pasture treated with sewage sludge. Environ Health Perspect. 2005;113(11):1580–7.
Pickford DB. Screening chemicals for thyroid-disrupting activity: a critical comparison of mammalian and amphibian models. Crit Rev Toxicol. 2010;40(10):845–92. doi:10.3109/10408444.2010.494250.
Pinto PIS, Guerreiro EM, Power DM. Triclosan interferes with the thyroid axis in the zebrafish (Danio rerio). Toxicol Res. 2013;2(1):60–9. doi:10.1039/c2tx20005h.
Rhind SM. Anthropogenic pollutants: a threat to ecosystem sustainability? Phil Trans Roy Soc Lond B Biol Sci. 2009;364(1534):3391–401. doi:10.1098/rstb.2009.0122.
Rhind SM, Smith A, Kyle CE, Telfer G, Martin G, Duff E, Mayes RW. Phthalate and alkyl phenol concentrations in soil following applications of inorganic fertiliser or sewage sludge to pasture and potential rates of ingestion by grazing ruminants. J Environ Monit. 2002;4(1):142–8.
Rodgers-Gray TP, Jobling S, Kelly C, Morris S, Brighty G, Waldock MJ, Sumpter JP, Tyler CR. Exposure of juvenile roach (Rutilus rutilus) to treated sewage effluent induces dose-dependent and persistent disruption in gonadal duct development. Environ Sci Technol. 2001;35(3):462–70.
Routti H, Nyman M, Jenssen BM, Backman C, Koistinen J, Gabrielsen GW. Bone-related effects of contaminants in seals may be associated with vitamin D and thyroid hormones. Environ Toxicol Chem. 2008;27(4):873–80. doi:10.1897/07-139.1.
Sarre SD, Ezaz T, Georges A. Transitions between sex-determining systems in reptiles and amphibians. Annu Rev Genomics Hum Genet. 2011;12:391–406. doi:10.1146/annurev-genom-082410-101518.
Sarre SD, Georges A, Quinn A. The ends of a continuum: genetic and temperature-dependent sex determination in reptiles. BioEssays. 2004;26(6):639–45. doi:10.1002/bies.20050.
Scaps P. A review of the biology, ecology and potential use of the common ragworm Hediste diversicolor (O.F. Muller) (Annelida: Polychaeta). Hydrobiologia. 2002;470(1–3):203–18. doi:10.1023/a:1015681605656.
Schnitzler JG, Siebert U, Jepson PD, Beineke A, Jauniaux T, Bouquegneau JM, Das K. Harbor porpoise thyroids: histologic investigations and potential interactions with environmental factors. J Wildl Dis. 2008;44(4):888–901.
Scholz S, Renner P, Belanger SE, Busquet F, Davi R, Demeneix BA, Denny JS, Leonard M, McMaster ME, Villeneuve DL, Embry MR. Alternatives to in vivo tests to detect endocrine disrupting chemicals (EDCs) in fish and amphibians–screening for estrogen, androgen and thyroid hormone disruption. Crit Rev Toxicol. 2013;43(1):45–72. doi:10.3109/10408444.2012.737762.
Schrattenholz A, Soskic V, Schopf R, Poznanovic S, Klemm-Manns M, Groebe K. Protein biomarkers for in vitro testing of toxicology. Mutat Res. 2012;746(2):113–23. doi:10.1016/j.mrgentox.2012.02.008.
Shafir S, Van Rijn J, Rinkevich B. The use of coral nubbins in coral reef ecotoxicology testing. Biomol Eng. 2003;20(4–6):401–6.
Soffker M, Tyler CR. Endocrine disrupting chemicals and sexual behaviors in fish – a critical review on effects and possible consequences. Crit Rev Toxicol. 2012;42(8):653–68. doi:10.3109/10408444.2012.692114.
Sonne C. Health effects from long-range transported contaminants in Arctic top predators: an integrated review based on studies of polar bears and relevant model species. Environ Int. 2010;36(5):461–91. doi:10.1016/j.envint.2010.03.002.
Tarrant AM. Hormonal signaling in cnidarians: do we understand the pathways well enough to know whether they are being disrupted? Ecotoxicology. 2007;16(1):5–13. doi:10.1007/s10646-006-0121-1.
Thornburg KL, Shannon J, Thuillier P, Turker MS. In utero life and epigenetic predisposition for disease. Adv Genet. 2010;71:57–78. doi:10.1016/B978-0-12-380864-6.00003-1.
Trites AW, Donnelly CP. The decline of Steller sea lions Eumetopias jubatus in Alaska: a review of the nutritional stress hypothesis. Mammal Rev. 2003;33(1):3–28. doi:10.1046/j.1365-2907.2003.00009.x.
UNEP/WHO. State of the science of endocrine disrupting chemicals — 2012. Geneva, Switzerland: United Nations Environment Programme/World Health Organization;2012.
van Aerle R, Nolan TM, Jobling S, Christiansen LB, Sumpter JP, Tyler CR. Sexual disruption in a second species of wild cyprinid fish (the gudgeon, Gobio gobio) in United Kingdom freshwaters. Environ Toxicol Chem. 2001;20(12):2841–7.
Vorkamp K, Thomsen M, Moller S, Falk K, Sorensen PB. Persistent organochlorine compounds in peregrine falcon (Falco peregrinus) eggs from South Greenland: levels and temporal changes between 1986 and 2003. Environ Int. 2009;35(2):336–41. doi:10.1016/j.envint.2008.08.008.
Waye A, Trudeau VL. Neuroendocrine disruption: more than hormones are upset. Toxicol Environ Health B Crit Rev. 2011;14(5–7):270–91. doi:10.1080/10937404.2011.578273.
White SS, Fenton SE, Hines EP. Endocrine disrupting properties of perfluorooctanoic acid. J Steroid Biochem Mol Biol. 2011;127(1–2):16–26. doi:10.1016/j.jsbmb.2011.03.011.
Williams RJ, Keller VD, Johnson AC, Young AR, Holmes MG, Wells C, Gross-Sorokin M, Benstead R. A national risk assessment for intersex in fish arising from steroid estrogens. Environ Toxicol Chem. 2009;28(1):220–30. doi:10.1897/08-047.1.
Acknowledgements
The authors are grateful to Dr Alice Baynes (Brunel University, Uxbridge, UK) for her constructive comments during preparation of this article.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Kumar, E., Holt, W.V. (2014). Impacts of Endocrine Disrupting Chemicals on Reproduction in Wildlife. In: Holt, W., Brown, J., Comizzoli, P. (eds) Reproductive Sciences in Animal Conservation. Advances in Experimental Medicine and Biology, vol 753. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-0820-2_4
Download citation
DOI: https://doi.org/10.1007/978-1-4939-0820-2_4
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-0819-6
Online ISBN: 978-1-4939-0820-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)