
Abstract
Endocrine disrupting chemicals (EDCs) known to be present in the marine environment, have great potential to influence reproductive status in aquatic organisms. A widespread incidence of intersex linked with EDCs exposure has been reported in molluscs and fish species. The species Scrobicularia plana is inherently gonochoristic; nonetheless, the feminisation of male clams was described in estuaries from the United Kingdom. Thus, this study reports the presence of oocytes in the testis of male clams of S. plana collected in the Guadiana River (Portugal). Samples were collected between June and September (2007), and gonad tissue analysed by light microscopy. Intersex with several degrees of intensity was detected during the first three months of sampling, in which the percentage of male clams affected varied between 5.9, 58.3 and 71.4%. Little is known about the causes of intersex in S. plana, being the urban, industrial and agricultural discharges possible major sources of induction. In the Guadiana River, the presence of several EDCs may be directly linked to the incidence of the intersex condition detected in Scrobicularia plana.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The aquatic environment is principally vulnerable to pollution, partly because of the significant release of chemicals into rivers, lakes, and the sea, mainly through effluents from sewage treatment works (STW) and industries (Sumpter 2005). Municipal effluents, even after an appropriate treatment, are known to release diverse pollutants into the aquatic environment, including endocrine disruptor chemicals (EDCs), which comprise natural or synthetic compounds, such as polychlorinated biphenyls, organochlorine pesticides, plasticizers, surfactants, pharmaceutical agents and miscellaneous steroids. Many known EDCs are estrogenic, with the ability to mimic or block the action of hormones or to influence their concentration in tissues, as seen in several aquatic organisms, for example, fish (e.g. Rutilus rutilus, Xiphias gladius, Acipenser gueldenstaedtii) and invertebrate species, as Mya arenaria, marine and freshwater mussels (e.g. Mytilus galloprovincialis, Tapes philippinarium, Elliptio complanata) (Minier et al. 2000; Jobling et al. 2002; De Metrio et al. 2003; Gagné and Blaise 2003; Canesi et al. 2004; Langston et al. 2005; Sumpter 2005; Bjerregaard et al. 2006; Jackson et al. 2006; Porte et al. 2006; Almeida et al. 2007).
The majority of xenobiotic estrogens and their metabolites are able to bioaccumulate and biomagnify in different environmental compartments, including marine biota, due to their lipophilic and constant nature (Porte et al. 2006). Several ecological effects have been identified, such as altered sexual differentiation, malformation, feminisation, alteration of immune function and decreased fertility in birds, fish and mammals. Estrogens in particular (nonylphenol, estradiol, etc.) possess the ability to increase the number of intersexed animals following periods of chronic exposure, as well as induction of vitellogenin (Jobling et al. 2002; Gagné and Blaise 2003; Sumpter 2005; Bjerregaard et al. 2006; Jackson et al. 2006; Porte et al. 2006; Ford et al. 2007). Intersexuality is a condition whereby an individual possesses oocytes or different stages of spermatogonia, at varying degrees of development, within the normal gonad of the opposite gender (i.e. spermatocytes in the ovary or oocytes in the testis), however, little is know about the physiological mechanisms or gonodal differentiation in spontaneous intersexual individuals of gonochoristic species. Fish species, from both freshwater and marine environments, present the most well documented cases of feminisation of males, characterized by the occurrence of female gonodal characteristics in males gonads (ovotestis) (Minier et al. 2000; Jobling et al. 2002; De Metrio et al. 2003; Bjerregaard et al. 2006; Jackson et al. 2006).
In invertebrate species, only a limited number of studies have focused the effects of EDCs, mainly due to a lack of knowledge on basic endocrine physiology of most invertebrate groups (Depledge and Billinghurst 1999; Langston et al. 2005; Sumpter 2005; Porte et al. 2006). Bivalve molluscs have been used worldwide as bioindicators to assess the impact of pollutants in coastal marine ecosystems and their health status (Bebianno and Serafim 1998; Bocchetti and Regoli 2006). Sedentary bivalves such as the estuarine bivalve Scrobicularia plana (Da Costa 1778) are potentially exposed to a wide range of contaminants normally for extensive periods of time, becoming highly tolerant to physical and chemical changes in the sediment (ultimate sink for pollutants). This, together with the ability to bioaccumulate pollutants and a deposit-feeding behaviour, makes S. plana an ideal biomonitor for investigating the effects of environmental contamination (Langston et al. 1998; Wootton and Pipe 2003; Casagranda and Boudouresque 2005; Langston et al. 2005). This bivalve is a burrowing deposit-feeding organism, inhabiting soft bottoms of clay or mud in the inter-tidal zone of estuaries. Geographically, it ranges from the Norwegian and Baltic Seas, south to the Iberian Peninsula, into the Mediterranean, along the Atlantic coast of Morocco to the Canary Isles and south to Senegal (Sola 1997; Guerreiro 1998; Rodríguez-Rúa et al. 2003; Wootton and Pipe 2003; Raleigh and Keegan 2006).
The Portuguese coast and marine areas are impacted by human activities, influencing the structure and dynamics of its estuaries, fundamentally through urban, agricultural and harbour effluents and discharges (Almeida et al. 2007). The Guadiana River is the fourth most important river of the Iberian Peninsula, located along the southern border between Portugal and Spain, close to the city of Vila Real de Santo António and next to the Gulf of Cádiz (Cravo et al. 2006; Almeida et al. 2007). Being one of the largest rivers in the south of the Iberian Peninsula, with 810 km in length, its estuary is 70 km long with a maximum width of 550 m and maximum depth between 5 and 17 m (Cravo et al. 2006; Wolanski et al. 2006; Caetano et al. 2006). In order to evaluate the impact of the presence of EDCs in the Guadiana estuary, the presence of intersex in male clams S. plana, was investigated.
Materials and methods
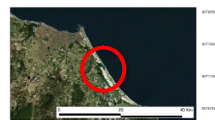
Samples of S. plana were collected in the Guadiana River estuary (Fig. 1), located in the south coast of Portugal, during the period of June to September 2007. 30 individuals were sampled each month, and maintained in aerated 50% seawater during 48 h. For each specimen, a small portion of the gonad was excised and examined under light microscope (Axiovert S100, Zeiss) with a digital still camera incorporated (Sony), at a 100× magnification, in order to detect gender and reproductive development stage. Oocyte sizes were measured using a graticulate eyepiece with a micrometric scale. In order to test differences in the frequency of intersex and sex ratio between months, Fisher Exact analysis was used, with a level of significance set at P < 0.05. Statistical analyses were carried out in SigmaStat version 3.5 (Systat Software, CA, USA).

Sampling site in the Guadiana Estuary (south coast of Portugal)
Results and discussion
The clam S. plana is considered inherently gonochoristic (distinct male and female characteristics) (Rodríguez-Rúa et al. 2003; Raleigh and Keegan 2006), however the presence of intersex has been shown to be a widespread phenomenon in some populations in the Southwest of the United Kingdom (Langston et al. 2007). In this study, intersex condition was characterized by the occurrence of oocytes among normal testicular tissue (ovotestis), in individuals of S. plana collected during June to September (2007) in the Guadiana Estuary. In Portugal, a phenomenon designated as ‘hermaphroditism’ has already been identified in some individuals of S. plana in the Tagus River, where the male testicular tissue was well developed with the existence of few oocytes, and also several oocytes and only few portions of the male tissue (Paes-da-Franca 1956). This could be the first evidence of the occurrence of a condition related/similar to intersex in S. plana in a Portuguese River.
Scrobicularia plana is a species whose reproductive cycle varies with latitude, with southern populations exhibiting more than one annual reproductive period (Sola 1997; Rodríguez-Rúa et al. 2003; Raleigh and Keegan 2006). Paes-da-Franca (1956) and Guerreiro (1998) showed that S. plana has two reproduction cycles, March-April and June-September, that preceded two periods of spawning in April-July and in September. Additionally, in the Guadalquivir estuary in Spain, populations of this species presented the same periods of sexual activity and spawning (Rodríguez-Rúa et al. 2003). Accordingly, the sampling time selected for this preliminary study was one of the periods of maximum sexual activity for this species in European southern regions. In fact, the spawning period occurred in September, where gonodal tissue was undifferentiated, becoming impossible to determine the gender of individuals.
For the 30 individuals of S. plana sampled each month, the occurrence of ovotestis increased in frequency until the spawning period (Fig. 2a). The intersex incidence showed a lower prevalence in June (P < 0.05), with 5.9% of males affected by intersex (3.3% of the total population), contrasting with the 58.3% in July (23.3% of the total population) and 71.4% (33.3% of the total population) in August (Fig. 2b). The higher frequency of intersex in S. plana is in agreement with the periods of higher sexual activity, with a peak in August (P < 0.05), as demonstrated in the populations from southwest United Kingdom (Langston et al. 2007) and in the Tagus estuary population, referenced as ‘hermaphroditism’ (Paes-da-Franca 1956).

a Intersex frequency, expressed as the percentage of males with oocytes in the testicular tissue and sex ratio of ‘unaffected’ males; b Percentage of males, females and intersex in the three months
The proportion of females was very similar between the 3 months (June to August), ranging from 43.3 to 60%, while the proportion of ‘unaffected’ males dropped from 53.3 to 16.7% and 13.3%, respectively (Fig. 2b). Consequently, the sex ratio between males and females decreased significantly (P < 0.05). Chesman and Langston (2006) detected a ratio of males (without intersex) to females lower than the unity in the S. plana population from the Avon Estuary (United Kingdom), thus confirming the result obtained in this study. In the studies that reported the existence of ‘hermaphrodites’ in this species, the sex ratio indicated a slight predominance of females to males, however with no significant differences (Paes-da-Franca 1956; Raleigh and Keegan 2006). In normal populations, equal proportions of males and females were recorded in the Mira and Tagus estuary in Portugal and in the Bidasoa estuary in Spain (Sola 1997; Guerreiro 1998).
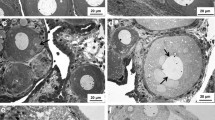
Different degrees of intersex in S. plana were observed, with an increasing severity throughout the sampling months (Fig. 3). Adopting the ranking system implemented by Chesman and Langston (2006) to categorise the different degrees of intersex in gonads of Scrobicularia plana (from a predominantly male gonad: single oocyte present in a field of view to a predominantly female gonad with follicles containing bundles of sperm), the condition of the males from Guadiana River did not cover the span of the scoring index used. In contrast to the situation found in the southwest of the United Kingdom population, in S. plana from the Guadiana River only four of the six scores were detected, ranging from a single oocyte in a normal testicular tissue to more than five closely associated oocytes. In June, only the first score was observed, with normal male gonad showing a single oocyte in a field of view (Fig. 3c), while in July a mixture of score one and two was detected, with one and two oocytes (Fig. 3d). As for August, two more phases of intersex were detected, the third and fourth and scores, with predominantly male tissue with one to four closely associated oocytes (Fig. 3e) or with more than five closely associated oocytes (Fig. 3f), therefore representing the higher severity of intersex noticed.

Light micrographs of the portion of the gonads of S. plana a normal female ovarian tissue, b normal male testicular tissue, c to f male testicular tissue showing the presence of oocytes, with different degrees of intersex. Size of oocytes ≈1.5 to 2.8 μm
The incidence of intersex in gonochoristic populations of S. plana has raised several questions, namely its principal source. The presence of the two genders in the same individual (intersexed gonads) has been reported as a normal condition in gonochoristic species (Lammens 1967), nevertheless, their existence has also been associated to the vicinity to polluted environments, as seen in wild populations of gonochoristic teleosts (Jackson et al. 2006). In Scrobicularia plana, intersex was also hypothesised as a normal condition, nonetheless, Langston et al. (2007) has shown that it can be induced after laboratorial exposure to (xeno)estrogens (17 β-oestradiol, 17α-ethinyloestradiol, octylphenol and nonylphenol), with increased ovotestis condition in male clams. Admitting that several other factors can influence the presence of intersex and sex ration in invertebrates, like metals (cadmium) and parasitism (Morley 2006; Smaoui-Damak et al. 2006; LeBlanc 2007), none of these were found significant. Parasites were not observed and metal levels in this species were not significantly relevant to induce this phenomenon (Gomes, unpublished data). Therefore, the influence of endocrine disrupting compounds on intersex condition appears to be an important factor affecting this species.
The presence of endocrine disrupting compounds in the aquatic environment is well documented, being the outfalls from sewage treatment plants, agricultural and cattle breeding the major inputs (Minier et al. 2000; Gagné and Blaise 2003; Langston et al. 2005; Sumpter 2005; Bjerregaard et al. 2006; Almeida et al. 2007). In the case of the Guadiana waters, only few studies have focused on the presence of these compounds. Almeida et al. (2007) detected significant levels of EDCs in the Guadiana River, not only in water samples, but also in sediments. The major compounds found were pesticides and their metabolites (atrazine, simazine, prometryn, among others), polycyclic aromatic hydrocarbons (phenantherene, pyrene, etc.) and herbicides as diuron. Bisphenol A (a plastic monomer) was also found, although in small amounts. The presence of EDCs in the Guadiana Estuary area is clear, with several point and diffuse sources resulting from urbanisation, agriculture, cattle breeding, olive and vineyard crops. The intensive agricultural activities and pesticide treatments that occur in the Spanish areas close to its hydrological basin also influences the discharges in the river (Wolanski et al. 2006; Almeida et al. 2007). Additionally, sewage discharges from the two densely populated cities near the river mouth, Vila Real de Santo António and Ayamonte (Spanish side), can be associated to the presence of EDCs (Serafim et al. 2006; Almeida et al. 2007), namely Vila Real Santo António, that has no sewage treatment plant for its effluents, being the sewage directly discharged into the river.
In sediment samples, two organotic compounds, dibutyltin and tributyltin (TBT), were found in considerable levels, essentially in the ‘Castro Marim’ marshes near the S. plana sampling sites. Being a river with high maritime traffic (mainly in Spring and Summer), one of the major sources of organotin compounds in the Guadiana River is still the antifouling paint, whose use was forbiden in Portugal since 1992 (Barroso et al. 2000). Due to its deposit-feeding behaviour, this species is potentially susceptible to exposure to a extensive range of contaminants (Sola 1997; Coelho et al. 2006). Therefore, the hypothesis that the presence of endocrine-disrupting chemicals in the waters of the Guadiana River is directly related to the intersex condition detected in S. plana cannot be discharged.
Two other examples of alterations of the endocrine system of marine invertebrates have been studied in the Guadiana River. The phenomenon of imposex (superimposition of male characters onto the female) has been recorded in the prosobranch Ocenebra erinacea, associated with the presence of TBT in the river (Coelho PhD Dissertation; Veiga unpublished data). Also, the levels of vitellogenin-like (Vg-like) proteins were quantified in Mytilus galloprovincialis by the indirect assay termed alkali-labile phosphate assay (ALP), showing significant levels of Vg-like proteins, probably related to EDCs in the Guadiana waters (Pereira, data not published).
In conclusion, intersex was identified in S. plana from the Guadiana River, in a less severe degree when compared to that observed in the populations from the United Kingdom. The present study provides important information to the question as to how geographically widespread is the presence of intersex in S. plana is, but not a response to which precise factors lead to this phenomenon. The next steps to be accomplished are a more complex characterization of the levels of EDCs in the Guadiana waters and S. plana tissues in order to relate them to the incidence of intersex in these clams and apply other endocrine disruption methodologies to complement the information about the influence of EDCs in this species.
References
Almeida C, Serôdio P, Florêncio MH, Nogueira JMF (2007) New strategies to screen for endocrine-disrupting chemicals in the Portuguese marine environment utilizing large volume injection-capillary gas chromatography-mass spectrometry combined with retention time locking libraries (LVI-GC-MS-RTL). Anal Bioanal Chem 387:2569–2583
Barroso CM, Moreira MH, Gibbs PE (2000) Comparison of imposex and intersex development in four prosobranch species for TBT monitoring of a southern European estuarine system (Ria de Aveiro, NW Portugal). Mar Ecol Prog Ser 201:221–232
Bebianno MJ, Serafim MA (1998) Comparison of metallothionein induction in response to cadmium in the gills of the bivalve molluscs Mytilus galloprovincialis and Ruditapes decussatus. Sci Total Environ 214:123–131
Bjerregaard LB, Korsgaard B, Bjerregaard P (2006) Intersex in wild roach (Rutilus rutilus) from Danish sewage effluent-receiving streams. Ecotoxicol Environ Saf 64:321–328
Bocchetti R, Regoli F (2006) Seasonal variability of oxidative biomarkers, lysosomal parameters, metallothioneins and peroxisomal enzymes in the Mediterranean mussel Mytilus galloprovincialis from Adriatic Sea. Chemosphere 65:913–921
Caetano M, Vale C, Falcão M (2006) Particulate trace metal distribution in Guadiana estuary punctuated by flood episodes. Estuar Coast Shelf Sci 70:109–116
Canesi L, Lorusso LC, Ciacci C, Betti M, Zampini M, Gallo G (2004) Environmental estrogens can affect the function of mussel hemocytes through rapid modulation of kinase pathways. Gen Comp Endocrinol 138:158–165
Casagranda C, Boudouresque CF (2005) Abundance, population structure and production of Scrobicularia plana and Abra tenuis (Bivalvia: Scrobicularidae) in a Mediterranean Brackish Lagoon, Lake Ichkeul, Tunisia. Int Rev Hydrobiol 90:376–391
Chesman BS, Langston WJ (2006) Intersex in the clam Scrobicularia plana: a sign of endocrine disruption in estuaries? Biol Lett 2:420–422
Coelho JP, Rosa M, Pereira E, Duarte A, Pardal MA (2006) Pattern and annual rates of Scrobicularia plana mercury bioaccumulation in a human induced mercury gradient (Ria de Aveiro, Portugal). Estuar Coast Shelf Sci 69:629–635
Cravo A, Madureira M, Felícia H, Rita F, Bebianno MJ (2006) Impact of outflow from the Guadiana River on the distribution of suspended particulate matter and nutrients in the adjacent coastal zone. Estuar Coast Shelf Sci 70:63–75
De Metrio G, Corriero A, Desantis S, Zubani D, Cirillo F, Deflorio M, Bridges CR, Eicker J, de la Serna JM, Megalofonou P, Kime DE (2003) Evidence of a high percentage of intersex in the Mediterranean swordfish (Xiphias gladius L.). Mar Pollut Bull 46:358–361
Depledge MH, Billinghurst Z (1999) Ecological significance of endocrine disruption in marine invertebrates. Mar Pollut Bull 39:32–38
Ford AT, Martins I, Fernandes TF (2007) Population level effects of intersexuality in the marine environment. Sci Total Environ 374:102–111
Gagné F, Blaise C (2003) Effects of municipal effluents on serotonin and dopamine levels in the freshwater mussel Elliptio complanata. Comp Biochem Physiol Part C 136:117–125
Guerreiro J (1998) Growth and production of the bivalve Scrobicularia plana in two southern European estuaries. Vie Milieu 48(2):121–131
Jackson K, Hurvitz A, Din SY, Goldberg D, Pearlson O, Degani G, Levavi-Sivan B (2006) Anatomical, hormonal and histological description of captive Russian sturgeon (Acipenser gueldenstaedtii) with intersex gonads. Gen Comp Endocrinol 148:359–367
Jobling S, Coey S, Whitmore JG, Kime DE, Van Look KJW, McAllister BG, Beresford N, Henshaw AC, Brighty G, Tyler CR, Sumpter JP (2002) Wild intersex roach (Rutilus rutilus) have reduced fertility. Biol Reprod 67:515–524
Lammens JJ (1967) Growth and reproduction in a tidal flat population of Macoma balthica (L.). Neth J Sea Res 3:315–382
Langston W, Bebianno MJ, Burt G (1998) Metal handling strategies in molluscs. In: Langston W, Bebianno MJ (eds) Metal metabolism in aquatic environments. Kluwer, New York, pp 219–283
Langston WJ, Burt GR, Chesman BS, Vane CH (2005) Partitioning, bioavailability and effects of oestrogens and xeno-oestrogens in the aquatic environment. J Mar Biol Assoc UK 85:1–31
Langston WJ, Burt GR, Chesman BS (2007) Feminisation of male clams Scrobicularia plana from estuaries in Southwest UK and its induction by endocrine-disrupting chemicals. Mar Ecol Prog Ser 333:173–184
LeBlanc GA (2007) Crustacean endocrine toxicology: a review. Ecotoxicol 16:61–81
Minier C, Caltot G, Leboulanger F, Hill EM (2000) An investigation of the incidence of intersex fish in Seine-Maritime and Sussex regions. Analusis 28(9):801–806
Morley NJ (2006) Parasitism as a source of potential distortion in studies on endocrine disrupting chemicals in molluscs. Mar Pollut Bull 52:1330–1332
Paes-da-Franca ML (1956) Variação sazonal das gónadas em Scrobicularia plana da Costa. Arq Mus Bocage 27:107–124
Porte C, Janer G, Lorusso LC, Ortiz-Zarragoitia M, Cajaraville MP, Fossi MC, Canesi L (2006) Endocrine disruptors in marine organisms: approaches and perspectives. Comp Biochem Physiol Part C 143:303–315
Raleigh J, Keegan BF (2006) The gametogenic cycle of Scrobicularia plana (Mollusca: Bivalvia) in Mweeloon Bay (Galway, west coast of Ireland). J Mar Biol Assoc UK 86:1157–1162
Rodríguez-Rúa A, Prado MA, Romero Z, Bruzón M (2003) The gametogenic cycle of Scrobicularia plana (Da Costa, 1778) (Mollusc: Bivalve) in Guadalquivir estuary (Cádiz, SW Spain). Aquac 217:157–166
Serafim A, Company R, Cravo A, Bebianno MJ (2006) Impact of metals in bivalve species from Guadiana River and Estuary. Met Ions Biol Med 9:457–462
Smaoui-Damak W, Rebai T, Berthet B, Hamza-Chaffai A (2006) Does cadmium pollution affect reproduction in the clam Ruditapes decussates? A one-year case study. Comp Biochem Physiol Part C 143:252–261
Sola JC (1997) Reproduction, population dynamics, growth and production of Scrobicularia plana Da Costa (Pelecypoda). Neth J Aquat Ecol 30(4):283–296
Sumpter JP (2005) Endocrine disrupters in the aquatic environment: an overview. Acta Hydrochem Hydrobiol 33:9–16
Wolanski E, Chicharo L, Chicharo MA, Morais P (2006) An ecohydrology model of the Guadiana Estuary (South Portugal). Estuar Coast Shelf Sci 70:132–143
Wootton EC, Pipe RK (2003) Structural and functional characterisation of the blood cells of the bivalve molluscs, Scrobicularia plana. Fish Shellfish Immunol 15:249–262
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gomes, T., Gonzalez-Rey, M. & Bebianno, M.J. Incidence of intersex in male clams Scrobicularia plana in the Guadiana Estuary (Portugal). Ecotoxicology 18, 1104–1109 (2009). https://doi.org/10.1007/s10646-009-0359-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-009-0359-5