
Abstract
In the adult central nervous system, neurons are endowed with a remarkable power: a continuous structural remodeling of their connections owing to minuscule changes of their axonal terminal branches and synaptic boutons, simultaneously with the growing and pruning of their distal dendritic branches and spines. In this review, we summarize the history of the concept of structural plasticity from its beginnings in the nineteenth century to its consolidation in our days, with the establishment of a clear relationship between the structural plasticity of neural circuits and memory and learning processes. After a short review of the morphological changes observed at all levels of highly activated neurons (dendritic spines, distal dendritic branches, axonal initial segments) in adult healthy animals, we cover a more classical topic by reporting the structural changes consecutive to neuronal lesions, particularly after axotomy or deafferentation. Finally, we discuss work done with grafts of embryonic central nervous tissue in adult mice with genetic or experimental brain lesions in order to determine the capability of the embryonic cells to substitute missing neurons in the adult animals. We present in particular the “adaptive rejuvenation,” i.e., the plasticity induced in the host adult cells by the grafted immature neural cells to allow their interaction and subsequent integration in the adult circuit. Our conclusion is the remarkable capacity for structural plasticity of adult neural cells, which still needs the full understanding of their genetic regulation.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
2.1 Introduction
The term plasticity was introduced in biology in the middle of the nineteenth century, referring to the adaptability of an organism to changes in its environment (Jones 2000). In the field of neuroscience, even though neuroscientists are very much familiar and commonly used the noun “plasticity,” they do not share a common definition. The term plasticity is a source of misunderstanding because of its unreliability. Indeed, plasticity does not have the same meaning depending on the context it is used, for instance, plasticity of phenotypic expressions, synaptic plasticity, morphological or functional plasticity, plasticity of sensorimotor coordination, behavioral plasticity, and postlesion plasticity. As questioned by Jacques Paillard (1976), page 33: “Are the realities covered by this term sufficiently precise so that we can hope to see it to become a heuristic concept generator of new hypotheses and new experiences, and thus a useful concept in neurobiology?”
It is not our intention to provide a long summary of the history of all concepts covered by the generic name neuroplasticity. On the contrary, as neuromorphologists, the focus of the present chapter will be very much reduced to morphological aspects of the plasticity, and many of our examples are taken from the cerebellum. We know that the review is far from being complete. There are whole sections that are not even mentioned such as developmental plasticity, critical periods, and phylogenetic aspects of the neural plasticity. Finally, the specific case of neural stem cell and its potential interest in nervous system repair have been treated in other chapters of this book or in numerous recent reviews (see, for instance, in Martino et al. 2011; Saha et al. 2012).
2.2 Origin of the Term “Plasticity”
2.2.1 The Importance of Psychologists in the Development of the Plasticity Concept
From his earliest origins, man has been interested in understanding the world around him and the nature of his constant interactions with that world. Because man has rapidly intuited that such interactions were possible, thanks to the interface that represents the brain, the mystery of how the nervous system was built and how it functions has passionately and simultaneously disturbed the human being. The aphorism “know thyself” has been the incentive that has driven scientists of all ages to try to unravel the mysteries of brain function.
One of the most amazing abilities of the brain is the mental process that allows us to learn and to keep in memory the learned tasks. During the second half of the nineteenth century, with the earliest scientific studies on learning and memory, it became obvious that during all his life the human being was able to learn and to remember and that the brain was the receptacle to accumulate new knowledge. The brain should be therefore provided with some kind of mechanism capable of fixing and preserving constant acquisitions. Due to the very poor knowledge of the brain constitution, organization, and function, a number of speculations began to appear trying to imagine how the brain was able to work. It is of interest to recall that the more accurate and fertile imagination appeared among psychologists. At the middle of the nineteenth century, Alexander Bain (1818–1903), an “associationist psychologist,” who tried to endorse psychology with experimental sensorimotor physiology, published his textbook, entitled “The Senses and the Intellect” (1855) where he started to expose his ideas on the possible mechanisms of memory. His devise was “There is nothing I wish more than so to unite psychology and physiology.” These ideas were completed in another book, “Mind and Body: The Theories of Their Relation,” published 18 years later, where he explained in Chap. 5, devoted to the intellect, that the mechanism of retention would pass through the growth of the number of cell junctions between the nerve cells, establishing precise new neural groupings. Using his own words, Bain wrote on page 91: “For every act of memory, every exercise of bodily aptitude, every habit, recollection, train of ideas, there is a specific grouping, or co-ordination, of sensations and movements, by virtue of specific growths in the cell junctions. For example, when I see a written word and, as a result of my education, pronounce it orally, the power lies in a series of definite groupings or connexions of nerve currents in the nerve and centres of the eye, with currents in motor nerves proceeding to the chest, larynx and mouth; and these groupings or connexions are effected by definite growths at certain proper or convenient cell crossings”.
What was the meaning of “cell junctions” or “cell crossing” in the central nervous system when Bain wrote the abovementioned sentences? In his publications, Bain provided descriptions of the organization of the nervous tissue based primarily upon the histological studies of Lionel S. Beale (1828–1906) who considered, like many others of his contemporaries, that the nervous system was structurally continuous.
Alexander Bain (1873, pp. 118–119) summarized Beale’s ideas published in 1862 and 1863, as following: “The manner of connexion of the nerve-fibres […] is conjectured and figured by Dr. Beale in a plan that facilitates our conception of the physical growths underlying memory and acquisition […] He observed, […] a series of hues passing across the body of the cell, and continuing into its branches, or communicating with the nerves. He considers these lines as the tracks of nervous action through the cell […] He couples with, this appearance the doctrine (maintained by him, although disputed by others) that the nerves terminate in loops, and consequently form an unbroken nervous circuit. He then suggests that the cell-crossing is the place where the inner bendings of a great many independent circuits come into close neighbourhood, and affect one another by a process of the nature of electrical induction. Any one of the circuits being active, or excited, would impart excitement to all that came near it in the same cell”.
It is, therefore, evident that the knowledge on the organization of the central nervous system gathered by Bain was inadequate. Although Beale published his morphological work 8 years before Gerlach (1871), their conception of the organization of the CNS (central nervous system) was similar. Therefore, the description of Beale was of the same nature than the wrong hypothesis later on enunciated by Joseph von Gerlach (1820–1896), who with a flawed gold-impregnation method decided that the fibers (axons and dendrites) emerging from different cells fused and formed a large interlocking plexus or network that occupied the whole of the gray matter (“reticular theory” or “reticularism”). Within this framework and knowing that the nerve impulses were electrical in nature, it is not difficult to imagine what the notion of Alexander Bain of a “cell junction”: the points of fibers intercrossing, considered like the specific sites for cell-to-cell interactions. In any case, in agreement with Stanley Finger (1994), Bain should be considered like one of the pioneers in postulating that the malleability of the nervous system—in what is referring to memory and learning—must go through structural changes in its circuits and, more importantly, that nerve activity should be responsible for such changes. The ancient metaphor that the brain was like a wax tablet, in which you could write or delete messages, began to have its biological counterpart.
Almost simultaneously, another psychologist William B Carpenter (1813–1885) in his book “Principles on Mental Physiology, with Their Applications to the Training and Discipline of the Mind and the Study of Its Morbid Conditions” (1874) used the term “plastic” on a quite similar sense, although no attempts to explain the mechanisms were done. In the chapter related to Habits (Habit of Self-discipline), Carpenter wrote page 289: “Whilst, then, every one admits the special strength of those early impressions which are received when the Mind is most ‘plastic,’—most fitted to receive and retain them, and to embody them (as it were) into its own Constitution, […] shaping that Mechanism, whose subsequent action mainly determines our Intellectual and Moral character, and, consequently, the whole course of our conscious lives”.
Somewhat more perspicacious was William James (1842–1910), who in his book on “The Principles of Psychology” (Vol. 1, Chap. 4 devoted to Habit, 1890) provided one of the earliest descriptions of the plastic nature of the nervous system and the changes of its circuits associated with the foundation of habits. He wrote on page 105: “The change of structure […] may be invisible and molecular, as when a bar of iron becomes magnetic or crystalline through the action of certain outward cause. […] Plasticity, then, in the wide sense of the word, means the possession of a structure weak enough to yield to an influence, but strong enough not to yield all at once. […] Organic matter, especially nervous tissue, seems endowed with a very extraordinary degree of plasticity of this sort; so that we may without hesitation lay down as our first proposition the following, that the phenomena of habit in living beings are due to the plasticity of the organic materials of which their bodies are composed”.
James, though aware of his ignorance on the real nature of the morphological substrate providing plasticity to the brain, described the nerve impulses as responsible for the induction of the plastic changes. However, the search of brain anatomy and cellular organization does not receive the required attention. Despite the importance of his assumptions and interpretations, James succumbed to the main weakness of those days’ psychology, when the CNS was simply regarded as a black box and only the quality of incoming and outgoing messages was of interest and not what might happen inside the box. Despite this weakness, he correctly understood that nervous activity was responsible for the elaboration of new paths within the CNS, as he put it on page 107: “If habits are due to the plasticity of materials to outward agents, we can immediately see to what outward influences, if to any, the brain-matter is plastic. […] The only impressions that can be made upon them are through the blood, on the one hand, and through the sensory nerve-roots, on the other; and it is to the infinitely attenuated currents that pour in through these latter channels that the hemispherical cortex shows itself to be so peculiarly susceptible. The currents, once in, must find a way out. In getting out they leave their traces in the paths which they take. The only thing they can do, in short, is to deepen old paths or to make new ones; and the whole plasticity of the brain sums itself up in two words when we call it an organ in which currents pouring in from the sense-organs make with extreme facility paths which do not easily disappear”.
Further in the book, when James developed his ideas about the concept of plasticity (page 566), he wrote: “Let us then assume as the basis of all our subsequent reasoning this law: When two elementary brain-processes have been active together or in immediate succession, one of them, on reoccurring, tends to propagate its excitement into the other”.
Envisaging that the simultaneous activity might not only reinforce the interactions between excited neurons but also create new interactive paths. Therefore, in spite of his poor anatomical vision of the organization of the brain, James advanced in his promulgated law a concept that, if it is interpreted by our current knowledge, turns out to be prophetic since it foresees the one of the “Hebbian synapses” as defined by Donald Hebb in 1949, almost 60 years later (see below). It is, therefore, justified that in many papers dealing with the history of neuroplasticity, William James is considered to be the first researcher in using the term plasticity from a modern perspective (Berlucchi and Buchtel 2009).
2.2.2 The “Neuron Doctrine” and Its Corollary, the Synaptic Contacts, Provided a Favorable Correlate to Explain Neuroplastic Changes
The difficulty to envision a more anatomical solution to the problem raised by the understanding of the learning and memory mechanisms began to be dispelled 2 years before the publication of James’ book, when Santiago Ramon y Cajal (1852–1934) started the study of the organization of the cortex of the cerebellum with the Golgi impregnation. In this study, practically completed in 3 years (Cajal 1988–1890), Cajal not only revealed the organization of the cerebellar circuits, but the study had much more general implications. Indeed, the presence of basket cells, with their peculiar pericellular baskets and “pinceaux formations” (brushlike), together with the correct interpretation of the developmental history of the climbing fibers, and their ultimate disposition coiling up along the main branches of the Purkinje cell (PC) dendrite (see in Sotelo 2008), incited Cajal to conceive that the diffuse nerve network supposed by Gerlach (1871) and later on (1873) by Golgi (1843–1926) did not exist. On the contrary, axons—as it was the case for dendrites—ended freely. A few years later, Cajal (1892) formulated his “neuron doctrine” stipulating that between neuronal processes there was no continuity, only contiguity. Within this framework, the CNS was composed of individual cells, like all the other tissues of the body, communicating between them through specific contact zones (the nervous articulations of Cajal), later on (1897) coined synapses by Sherrington (1857–1952). From now on, a new path was opened for a better comprehension of the mechanisms involved in the malleability of the nervous tissue during the processes of learning and memory and for a better definition of neuronal plasticity.
The newly opened path was rapidly taken by Eugenio Tanzi (1856–1934), one of the earliest and enthusiastic defenders of the “neuronal doctrine” proposed by Cajal, who in 1893 had the intuition that nerve current should find some resistance at the passage from one neuron to the next, throughout their cell junctions. The stretching of the arriving axon and/or the receiving dendrite, when the cell junctions are repetitively stimulated, similarly to what happens when exercising muscle fibers (hypertrophy), might overpass this resistance. This distance reduction would be the plastic change required for the phenomenon of learning through the facilitation of the passage of nerve impulses. In Tanzi’s words (1893), page 469: “If now we think that the interposed distance between the terminal arborisation of one neuron and the body of the next neuron constitute a resistance or […] a kind of difficult passage (‘mal passo’) that the nervous wave must overcome not without difficulty, it is evident that the conductibility of the nervous system must stand in an inverse relation of the interval between the two neurons. To the extent that exercise has the tendency to shorten the distances, it increases the conductibility of the neurons that is their functional capacity” (taken from the English translation published in the paper by Berlucchi and Buchtel 2009).
Another important actor in the history of what from now on we can call “synaptic plasticity” was Ernesto Lugaro (1870–1940), a clinical physician and psychiatrist. Between 1898 and 1906, when he published his treatise on psychiatry (translated to English in 1909), Lugaro expanded Tanzi’s hypothesis and coined the term “plastic activity of neurons.” Thus, after summarizing Cajal’s anatomical ideas, he wrote pages 94–95: “Let us now see if we can draw some deductions in psychology from these data regarding the anatomy and general physiology of the nervous system […] One of the problems which psychology has always put to anatomy is that of interpreting psychic evolution during development and adult life […] Anatomy gives us the general impression that organs are systems of unchangeable and rigid structure. How then can psychic plasticity be reconciled with this rigidity? The neurone doctrine is capable of giving us a satisfactory answer if we admit that between neurone and neurone there is some kind of constant or temporary discontinuity, no matter whether the relationship is established by contact or by means of plastic connections which undergo modifications most readily.”
It is remarkable how Lugaro explained that the nervous system could be considered like a kind of “neoteny” (a process during which some developmental features are preserved in adulthood). For him, adult life is simply a continuation of development (“If then we consider that the psychic maturity acquired in adult life is simply a continuation of development itself, the explanation will appear more clear and convincing to us.”), and, therefore for the brain, aging would be only a slowed down developmental processes. Finally, as clearly addressed by Berlucchi and Buchtel (2009), Lugaro succeeded in making a bridge between Tanzi’s modifiability of synapses and Cajal’s neurotropism, focusing again on the synapses as the main location of the neuronal plasticity, pages 96–97: “Neurotropism […] does not exclude the possibility that in certain organisms the connections between the various neurones are immutable […] Probably this holds in the case of the invertebrates. But where the structure of the nerve centres, although already highly organised is still capable of further perfection—especially in the cerebral cortex—it is clear that chemotropic activities can still be the factor in bringing about new anatomical relationships. Tanzi formerly explained the consolidation of memories and automatic actions which become habitual, by means of an ever expanding mechanism which tends to approximate and unite in a progressively more intimate manner elements connected with each other […] This plastic activity of the neurones, which is just a continuation of what is more accentuated in the embryo though in quite a different manner, is naturally more active in infancy and youth than in adult life. And that explains the particular plasticity of the young mind which education models so easily. To the same cause can be attributed the fact that after destruction of some cortical centres functional compensation—impossible in the adult—is possible in the child […] With the increase of years the plasticity of the neurones becomes quiescent, but very slowly, and in fact we see how the aged may become incapable of modifying their own convictions and of yielding to the suggestion of new experiences.”
2.2.3 Cajal’s Writings on Neuroplasticity
In a review concerning the drawings of Cajal, Sotelo wrote about the pros and cons of his inspiring role on the development of the neurosciences since 1888 (Sotelo 2003). Indeed, from his first review (Cajal 1892) about the histology of the nervous centers, he always tried to link his structural discoveries with their possible functional meaning. Although he was prophetic in many of his functional assumptions, sometimes he changed opinion during his long life of work, and other of his assumptions turned out to be wrong (Sotelo 2003). One of these undecided topics is the one of neural plasticity. After his deep morphological analyses of the degeneration and regeneration of the nerve fibers (Cajal 1913/1914), Cajal was stressed by the fact that peripheral axons regenerate while central axons were only able of abortive regeneration. This essential disparity results from differences in the cellular and molecular environment, making the CNS a hostile milieu for axon growth. Moreover, during his early years of research as a neuroembryologist, Cajal—together with Wilhelm His (1886, 1889), Camillo Golgi (1873), and Alfred Kölliker (1896)—reached the conclusion that differentiated neurons do not divide and only neuroblasts were able of mitotic activity. Since the latter are only present in developing periods, the CNS of adult mammals had completely loss their capacity to generate new neurons. These two important observations prompted Cajal (1928) to deduce that: “In the adult centers the nerve paths are something fixed, ended and immutable. Everything may die, nothing may be regenerated.” This sentence has been taken by many authors as the irrefutable proof that Cajal was not a supporter of neuroplasticity.
However, after the publications of Jones (1994), DeFelipe (2006), and many others, the importance of Cajal’s contributions to the study of the plasticity of the nervous system through his theoretical correlations between histological results and mental processes became obvious. In his first review published in Spanish (Cajal 1892), he reported his new concept on brain organization, first introducing the pyramidal neurons as the responsible for psychic functions, page 38: “With some restrictions, it can be affirmed that in the animal kingdom psychic functions are bound to the presence of pyramidal cells (psychic cells) […] The number of elements of different category connected to one cell can be measured by the extension and the degree of differentiation reached by its protoplasmic arborizations [dendrites] […] Therefore, it can be estimated as likely that the psychic cell plays a wider and usefully activity when bigger are the number of its protoplasmic, somatic and collateral expansions, and more copious, longer and more branched are the collaterals coming out its cylinder—axis [axon].”
A few lines below, on page 39, he reported his ideas about the morphological changes that should accompany the intellectual reasoning and the talent, resulting from what he called “cerebral gymnastics”: “These facts are: the notable increase in intellect observed among men dedicated to deep and continued mental exercise; and the coexistence of a notable talent and even of a true genius with a medium or smaller sized brain than those of normal weight and dimensions. In the first case, […] it can be supposed that cerebral gymnastics will lead to the development of protoplasmic processes and nervous collaterals beyond that normally observed, forcing the establishment of new and more extensive intracortical connections. […] In the second case, there is nothing to prevent us from accepting that certain brains, either because they inherit prior adaptations or through other causes, offer a notable development of all kind of collaterals in compensation for the smaller number of cells […]”.
Therefore, neurons can respond to functional activation like many other body tissues, especially muscle tissue. While strength training exercises in gymnastics lead to muscle hypertrophy, mental exercise increases the intellect in parallel with the increase in the number of cortical synaptic connections.
It is important to notice that at the bottom of the page, Cajal made reference to Alexander Bain (he did not provide the complete reference), by saying: “If understand is, as Bain says, to perceive similarities or differences between our ideas, the depth and breadth of judgment will be much larger, the greater the number of acquisitions or images will be used as material, and more extensive is the system of relationships that among them consents to establish the cellular substratum of the brain.” This sentence seems to sum up the ideas of Bain - expressed with Cajal terms- that the retention mechanism passes through growth in the number of cell junctions between nerve cells allowing the establishment of precise new neural groupings as previously discussed.
Indeed, 2 years later, at the occasion of his invitation by the Royal Society of London as speaker for the Croonian Lecture, Cajal (1894) delivered in French a new version of his ideas on the action of “cerebral gymnastics” on mental processes. He wrote pages 466–467: “Cerebral gymnastics are not able of improving the brain organization by increasing the number of cells, because, it is known, that the nerve cells have lost after the embryonic period the property of proliferation; but […] mental exercise leads to a higher development of the protoplasmic extensions [dendrites] and of nervous [axonal] collaterals in the regions of the brain in use. In this way, associations already established among certain groups of cells would be significantly strengthen by means of the multiplication of the small terminal branchlets of the protoplasmic appendages and nervous collaterals; but, in addition, totally new intercellular connections could be established thanks to the neoformation of [axonal] collaterals and protoplasmic expansions [dendrites]”.
Cajal started by considering that even though his ideas were not original, the main difference now is that “cell junctions” take their real value, since they concern the morphological contacts (later named the synapses) between neurons. Therefore, after exposing his theoretical considerations, he finished: “This anatomical-physiological hypothesis is not original in principle, because there are many physiologists and psychologists who have searched for the somatic characteristic of intelligence into the richness of cellular associations, but it has something new, it is based on positive facts on the brain structure and not on mere assumptions about the organization and the relationships of the nervous corpuscles.”
In 1895, Cajal published in Spanish an interesting article entitled “Some conjectures about the anatomical mechanism of ideation, association and attention,” which attempted to provide a mechanistic model of the phenomenon of consciousness, by explaining how the brain could pass from one stage of rest to another of activity. In this paper, Cajal first discussed the propositions of Hermann Rabl-Rückhard (1839–1905) who, in his publication on the study of a crustacean cerebral ganglion (1890), launched the hypothesis of amoeboid movements of neuronal processes (see Bawden 1900; DeFelipe 2006). This proposition was extended by Mathias Duval (1844–1907) to the nervous system of all living animals (1895). Accordingly, for Duval the amoeboid movement of neuronal processes would be able to cause their retraction, with the separation of cell contacts, or conversely, their moving toward each other to restore the contacts. These movements could explain the inactive state induced by sleep, anesthetics, etc. (contact separation), while their approach, allowing the restoration of contacts, would be responsible for the states of brain activity (consciousness). In the last part of the paper, Cajal developed his adaptation of the Mathias Duval hypothesis. Based upon his failure, despite repetitive experiments, to establish direct correlations between the way of animal’s sacrifice (chloroform, curare poisoning, bleeding) with the anatomy of neuronal processes, Cajal was forced to give up with this hypothesis. However, his observation on the cerebral cortex prompted him to adapt the hypothesis of amoeboid movements, from neuronal to astrocytic processes. He said: “In contrast, our studies of the cerebral cortex have led us to suspect that, during mental work the morphology some glia cells varies. In the grey matter of the brain, neuroglial cells are sometimes shrunken, provided with short and stout appendages, some other times exhibit long, abundant expansions studded with infinite secondary and tertiary branchlets (cells of penniform expansions of Retzius). Between the retracted and relaxed states all transitions can be seen”.
During mental activity, the amoeboid movements of neuroglial processes would be enough to allow the withdrawal of thin astrocytic processes between axon terminals and postsynaptic elements, thereby restoring contacts that were disconnected during the mental rest. Opposite, the inactivity would raise the movement of thin astrocytic lamellae that would pull apart the pre- and postsynaptic elements, disconnecting this way inactive neuronal circuits. It is obvious that, even though a similar kind of plasticity linked with the withdrawal of astrocytic covering of supraoptic hypothalamic neurons has been reported (it takes place during the control and coordination of oxytocin and vasopressin release during special conditions such as lactation and dehydration; see in Hatton 1997), the mechanism conceived by Cajal has not been validated.
2.2.4 Synapses and Plasticity
After a period of decline between 1915 and 1940, during which the hypothesis of the synaptic location of learning was questioned (see Berlucchi and Buchtel 2009), it made a strong comeback in 1948 and 1949 with the publications from the neurophysiologist Jerzy Konorski and the psychologist Donald O. Hebb. In his important monograph, “Conditioned Reflexes and Neuron Organization,” published in 1948, Konorski discussed the mechanisms underlying the behavioral changes induced by conditional reflexes. He foresaw that these mechanisms should be quite similar to those of the spinal reflexes, which are based upon the reorganization of synaptic connections. Donald O. Hebb (1949) in his book “The Organization of Behavior: A Neuropsychological Theory” proposed that the synaptic activity between two neurons strengthens when the neurons display simultaneous activity. Hebb reported his postulate as follows (Chap. 4, page 50): “When an axon of cell A is near enough to excite a cell B and repeatedly and persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A’s efficiency, as one of the cells firing B, is increased.” It is noteworthy to compare the similitude of this sentence of Hebb with the one of William James on page 566 of his book, written 59 years earlier (cited above).
The morphological correlate in this connectionist theory of learning is based upon the above-discussed principle developed by Tanzi, Lugaro, and Cajal of growth of synaptic connections (Berlucchi and Buchtel 2009), in Hebb’s words (Chap. 4, page 50): “When one cell repeatedly assists in firing another, the axon of the first cell develops synaptic knobs (or enlarges them if they already exist) in contact with the soma of the second cell (Soma refers to dendrites and body, or all of the cell except its axon).”
Advances in physiology and morphology achieved in the second half of the twentieth century have focused the synapse as the seat of learning and memory. The discovery in 1973 by Tim Bliss and Terje Lømo of the long-term potentiation (LTP) of synaptic transmission in hippocampal neurons produced in response to a burst of high-frequency stimuli (tetanic stimulation), activating the perforant pathway to dentate granule cells in the dentate gyrus, the major synaptic pathways in the hippocampus, provided a direct proof that a gain in strength and effectiveness in synaptic transmission can result from a sustained high activity as that produced by experience. Furthermore, the fact that the hyperpolarization of a neuron is sufficient to prevent its LTP (Lin and Glanzman 1994) showed the validity of the Hebb’s principle. Synaptic plasticity can be considered today as a major mechanism for memory formation.
2.3 Morphological Neuronal Plasticity: Remodeling of Synaptic Contacts and Anatomical Changes in Dendrites and Axonal Initial Segments (AIS) in Adult Non-lesioned Animals
As it is beyond the scope of this review, we will not discuss the integration of new neurons in adult circuits, although neurogenesis that occurs at least in rodent hippocampus and olfactory bulb is a beautiful example of morphological neuronal plasticity (see reviews in Deng et al. 2010 and Lepousez et al. 2013, respectively).
2.3.1 Dystrophic Axon Terminals and Dendritic Growth Cones
For years the morphologists have been exploring the CNS trying to find some clues supportive of the connectionist theory of learning. If the neuronal circuits would be in continuous remodeling, it should be possible to observe these morphological changes at the synaptic level. The advent of electron microscopy paved the path toward the observation of a possible synaptic remodeling that should result from some kind of degenerative process for the removal of disused presynaptic axon terminals, followed by the formation of newly formed synaptic contacts (synapse formation/elimination and remodeling of axons and dendrites). Sotelo and Palay (1971), in their electron microscope study of the synaptology of the lateral vestibular neurons of apparently normal rats, reported the occurrence of enlarged nerve endings containing a variety of unusual structures already noticed years before in humans (infantile neuroaxonal dystrophy or Seitelberger’s disease; Sandbank et al. 1970), as well as in experimental animals (Tellez and Terry 1968), and considered to be the consequence of pathological processes. The alterations seen mostly involve membranous organelles, almost exclusively appearing in axon terminals and preterminal fibers. Similar dystrophic axons have been also reported in the dorsal column nuclei (Hashimoto and Palay 1965), lateral vestibular nucleus of the cat (Mugnaini et al. 1967), hypothalamus (Dellman and Rodriguez 1970), superior cervical ganglion (Ceccarelli et al. 1971), deep cerebellar nuclei (Angaut and Sotelo 1973), and autonomic nerve endings (Townes-Anderson and Raviola 1978). It is important to underline that some of these dystrophic axons were in continuity through a thin neck with a small axon terminal of normal appearance, as if the small extension was originated from the large abnormal terminal by a budding of process (Fig. 25 of Sotelo and Palay 1971). Sotelo and Palay proposed that the dystrophic axons were degenerating, whereas the budding off would represent the regenerative counterpart.
Together with this axonic remodeling, Sotelo and Palay (1968) also reported the presence of large dendritic varicosities at the tip of some dendritic branches, which received synaptic inputs from nearby axon terminals. These dendritic profiles were crowded by a very large population of mitochondria and by glycogen particles, together with customary organelles. These profiles, more abundant in the dorsal than in the ventral regions of the nucleus, are not the attribute of the lateral vestibular nucleus and have been encountered in other regions such as the cerebellar nuclei (Sotelo and Angaut 1973), superior cervical ganglion (Ceccarelli et al. 1971), and autonomic nerve endings (Townes-Anderson and Raviola 1978). Their location, connection with the parent dendrite, and, mainly, correlation with Golgi-impregnated preparations (Sotelo and Angaut 1973) have allowed to conclude that these profiles might represent at least the distal segment of dendrites, if not their actual tips, and were considered as growth cones of a growing or regenerating nerve cell dendrite. The hypothetical occurrence of axon terminals in different stages in a cycle of degeneration and regeneration of nerve endings in the normal animal, together with the possibility of dendritic growth, was taken as indirect proof of a continuous remodeling of synaptic connections. This way, axons and dendrites might engage in such a process, reflecting the continued experience of the animal, making and discarding interneuronal connections by the activity of both pre- and postsynaptic elements. Although this hypothetical interpretation was tempting to investigators interested in finding a morphological substrate for the process of learning, these static results were insufficient to provide a morphological base to the connectionist hypothesis of learning and were forced to oblivion.
2.3.2 LTP and Dendritic Spines
One of the essential paradoxes to be handled by the brain is to preserve the right balance between two seemingly contradictory conditions: the stability of the already established neuronal circuits, required for preserving behaviors and long-term memory, and the constant structural changes needed for the synaptic plasticity underlying learning. Therefore, the morphological changes in normal learning conditions should be limited but should occur. The most broadly studied synaptic changes occur in excitatory axospinous synapses and are mainly characterized by the addition or removal of a small fraction of their postsynaptic dendritic spines, causing minor quantitative changes.
Cajal (1888), in his first study of the cerebellar cortex impregnated by the Golgi method, reported the existence of spines, as peculiar tiny excrescences emerging along the distal compartment of Purkinje cell dendrites. After long and tedious discussions to determine if the spines were only the result of artifactual precipitation of silver over the neuronal surface or belonged to true protrusions designed to increase the receptive surface area of the dendritic tree, its reality was firmly established by Cajal (1896) when he was able to replicate their appearance with a vital dye—the methylene blue method—in cerebral cortical neurons. George Gray (1959), 63 years later, corroborated the existence of dendritic spines during his electron microscope study of the rat visual cortex. Gray added two important attributes to spines: they almost systematically establish asymmetric synaptic contacts (a morphological marker of excitatory synapses) and have a specific organelle—the spine apparatus—of unknown function.
From the discovery of LTP (see above), neuroscientists have tried to correlate the long-term potentiated synapses with their presumptive morphological counterparts. Since the targets for the axons of the perforant path originated in the entorhinal cortex are the spines of the granule cell dendrites of the dentate gyrus, these dendritic spines have been for the last 37 years the matter of numberless quantifications. The analysis of several parameters tempted to provide a solid structural support to the observed functional modifications. So, changes in the preexisting synapses concerning either the length and width of the spines or of their active zones, the shape and size of postsynaptic differentiations, or changes in the number of synapses in potentiated compared to non-potentiated hippocampi were addressed mostly with electron microscopy (Van Harreveld and Fifkova 1975; Fifkova and Van Harreveld 1977; Fifková and Anderson 1981; Geinisman et al. 1989; Desmond and Levy 1990; Trommald et al. 1996; Harris et al. 2003; for a more detailed bibliography see in Bailey and Kandel 1993). After laborious quantifications, despite the solidity of some of the positive results, their inconsistency did not permit any definitive conclusion that LTP, prolonged for minutes and even hours, was able to provoke reproducible structural alterations of axospinous synapses on dentate granule cells. The problem was solved later on with the advancement of new imaging techniques (see below).
2.3.3 Recent Progress in Live Cell Imaging Has Finally Demonstrated the Occurrence of Synaptic Remodeling: Formation and Pruning of Spines
The progresses in physical sciences and informatics have generated new tools for the study of the organization of the brain and the ability to examine the dynamic aspects of the neuron. For instance, the development of computer-assisted image analysis has promoted new kinds of quantitative studies, such as three-dimensional morphology of Golgi-impregnated neurons with their dendritic and axonic fields or 3-D reconstruction of neuronal compartments from electron micrographs. The development of confocal laser scanning microscopy (CLSM) has provided an enhanced resolution, compared to classical fluorescence microscopy, and has allowed the 3-D reconstruction of thick sections. Advances in laser technology during the last 10 years have permitted the development of a new type of CLSM that, keeping all the advantages of the confocal microscopy (ability to penetrate scattering tissues), have added the advantage of greatly reducing phototoxicity and photobleaching. This is the two-photon laser scanning microscopy (TPLSM), where fluorescence excitation above or below the plane of study is practically nonexistent. Thus, it can be used to study thick brain slices in organotypic culture or even neuronal compartments located several hundreds of micrometers deep in the living brain. These recent improvements have opened a new epoch of neuromorphology, because the observations in living tissues or animals have added the dimension of time to this discipline. In fact, under favorable conditions, it is possible, after repeated examinations, to analyze the morphology of a neuron, its dendritic and axonal fields, as well as its dendritic spines over prolonged periods, allowing the detection of presumptive morphological changes.
Engert and Bonhoeffer (1999), by using local superfusion technique together with two-photon imaging, were pioneers to directly observe in living hippocampal slices that the induction of LTP in synapses of CA1 region was able to initiate neo-spinogenesis, whereas in non-stimulated regions of the same dendrite or, more interesting, in slices where long-term potentiation was blocked, spinogenesis did not occur. A similar high dynamic of dendritic spines was corroborated by numerous investigators (see, for instance, Leuner and Shors 2004; Kasai et al. 2010 for a review). These first data, reporting intense spine remodeling in hippocampal and cortical neurons, were later on reevaluated because they were obtained from young, still somewhat immature, neurons, and it is likely that spine lifetimes vary greatly. Therefore, even though neosynaptogenesis and synaptic pruning take place throughout the life span, after the critical period, when the brains are reaching maturity, spine dynamics slow down and become stable. This stability has been confirmed in many studies and with different experimental paradigms (Grutzendler et al. 2002, see in Bhatt et al. 2009). Moreover, age difference was quantified by Zuo et al. (2005), who working on spines on the apical dendrites of layer 5 pyramidal neurons of the barrel, motor, and frontal cortices reported in observations conducted over 2 weeks that while in 1-month-old mice 13–20 % of spines are eliminated and 5–8 % formed, in 4–6-month-old adult mice, the remodeling was much lesser since in 2 weeks only 3–5 % of the spines were eliminated and 5–8 % formed. The results were more impressive if the observation period is prolonged over 18 months, only 26 % of spines were eliminated and 19 % formed in the adult barrel cortex, testifying for the stability of spines in adult brains.
Finally, to close this section on dendritic spines, we believe that it is worthy to mention the recent paper by Lai et al. (2012). These authors have investigated, by transcranial two-photon microscopy, the structural modifications of neuronal circuits in the mouse frontal association cortex by analyzing changes in dendritic spines of layer-V pyramidal neurons during fear learning and extinction. These authors arrived to an important conclusion: fear conditioning, extinction, and reconditioning lead to opposing changes at the level of individual synapses because reconditioning after conditioning and extinction eliminates those newly formed spines stabilized by extinction. Therefore, stabilization appears as an essential key required for memory storage, and the persistence of new spines can be considered as engrams in learning.
2.3.4 Changes in Dendritic Spines Are Correlated with Changes in Presynaptic Axon Terminals. Examples from the Hippocampus
In a recently published paper by the group of Michael Frostcher (Zhao et al. 2012), structural changes of mossy fibers and their postsynaptic counterpart have been associated with chemically induced LTP in slice cultures, taken from newborn mouse hippocampi and kept for 14 days in vitro. Ten minutes before fixation, the cultures were treated with tetraethylammonium (TEA), a blocker of potassium channels, to induce LTP. The interest of this work is that in order to avoid all possible artifacts consecutive to the chemical fixation of the biological material, the authors have used high-pressure freezing physical fixation. In wild-type mouse hippocampi, the length of the membrane of the mossy fiber boutons was increased and associated with an increase in the number of active zones and of postsynaptic spines.
Pico Caroni and collaborators (Galimberti et al. 2006) analyzed the synaptic remodeling between granule cell axons and the dendritic spines of pyramidal cells in the hippocampal CA3 region. In control adult mice, each granule cell axon established 10–15 complex synaptic arrangements—the mossy fiber varicosities—that are changeable throughout life. More importantly, the quasi constant remodeling of these complex synapses was greatly enhanced just by keeping the mice in an enriched environment. Comparable results were obtained in organotypic cultures of the hippocampus, where it was possible to demonstrate that the generation and maintenance of the plastic changes required spiking activity in the slices and transmitter release from mGluR2-sensitive receptors, because blocking this release with an mGluR2 agonist reversed the structural plastic changes. Functionally speaking, the plasticity changes by increasing the size of active zones in presynaptic membrane that allows for a stronger excitatory response in the postsynaptic neurons, which could lead to an increased complexity of the mossy fiber circuits, the presumptive substrate for increased hippocampal learning. More recently, Pico Caroni and collaborators (Ruediger et al. 2011) were able to show in adult mice after one trial and incremental learning that mossy fibers in the hippocampus and cerebellum present long-lasting and reversible increases in the numbers of filopodial synapses onto fast-spiking interneurons that trigger feed-forward inhibition. From different types of experiments, the authors showed a causal relationship between the morphological data (increase in number of synapses) and the precision of learning and memory. Thus, they proposed that feed-forward inhibition growth at hippocampal mossy fibers fitted with the precision of hippocampus-dependent memories. The special interest of this last paper is that, although the majority of the work related to the structural plasticity of axospinous synapses has been focused on glutamatergic excitatory transmission, Ruediger et al. (2011) have shed light on the important role that local inhibitory circuits have in the acquisition of learning and memory.
2.3.5 Changes in Dendritic Tree Morphology
The group directed by Dale Purves was a pioneer in the analysis of dendritic trees in in vivo specimens. Indeed, with Robert Hardley and James Voyvodic (Purves et al. 1986), they described the methodology to visualize, over intervals from 1 day to up to 3 months, the same neuron in repetitive experiments. These early studies were performed with peripheral neurons, those of the superior cervical ganglion of young adult mice, by injection of nontoxic vital fluorescent dyes and laser scanning confocal microscopy. The obtained results corroborated the working hypothesis, the occurrence of extensive remodeling in dendritic trees, the postsynaptic elements in the sympathetic ganglion. The important changes consisted in growth of some of the branches, with retraction or even disappearance of others, and the de novo formation of still other dendrites. These results provided the idea for a relatively extensive dendritic remodeling in adult sympathetic neurons that does not correspond to the actual concept we have of the neuronal stability. Here again, the age of the animals could be responsible of the early results.
A few years later, Dale Purves started another important study directed to determine whether central neurons, in this case those involved in the formation of olfactory bulb glomeruli, were continuously changing both during development and in adulthood. For that purpose, newborn, juvenile, and adult mice were analyzed over intervals of several hours to several weeks (LaMantia et al. 1992). The results emphasize that while in newborn and juvenile mice the development is not finished, and new glomeruli are added progressively, in adult mice there is no longer addition of new glomeruli, and the neuronal population involved in glomerular formation remains stable, as it was the case for dendritic spines (see above). In conclusion, despite the impressive stability of dendritic processes and spines in adult brains, there is a slow pace remodeling throughout the whole life of the animals.
2.3.6 Changes in Length and Position of the AIS
The last neuronal region to be characterized with the electron microscope was the axon initial segment (AIS). It was only identified in 1968 (Palay et al. 1968), as a specialized and important region, which emerges from the axon hillock and separates the axonal from the somato-dendritc compartment, acting as selective tight molecular filter for the axoplasmic transport (Song et al. 2009). AIS is therefore the command region for axonal identity and responsible for neuronal polarization. It has been morphologically identified by the occurrence of three specific ultrastructural features: (a) fascicles of microtubules, (b) sparsely distributed clusters of ribosomes, and (c) a dense layer of finely granular material undercoating the plasma membrane. A similar undercoating occurs beneath the plasma membrane of myelinated axons at the nodes of Ranvier (Andres 1965, Fig. 5). This anatomical region is where the action potentials are initiated (Coombs et al. 1957) and is assumed to possess a membrane provided with a high density of voltage-gated sodium (Na 1) channels (Kole et al. 2008).
During the last 6 years, it has been established that the AISs, with their strategic location and important functions, are not static in structure but also subjected to plasticity. These studies started with the electrophysiological analysis of the auditory brainstem neurons in the newly hatched chicken (between P2 and P5) (Kuba et al. 2006). In a recent review, Hiroshi Kuba (2012) summarized his work explaining that in the nucleus magnocellularis (involved in a relay of timing information) the length of the AIS changes in relationship with sound frequency (increases with decreasing frequencies) to accommodate frequency-specific variations in synaptic inputs. In the nucleus laminaris (involved in integrating time from sounds received from both ears for their localization) the length and the location of the AISs vary depending on sound frequency; they are shorter and more remote for higher frequency. Juan Burrone and Matthew Grubb have corroborated and expanded the study of AIS plasticity (Grubb and Burrone 2010). They used dissociated cultures of hippocampal neurons taken from E18 rat fetuses. By increasing the levels of activity, these investigators were able to change the precise location of the AISs of the stimulated excitatory neurons. When the neurons, for example, were kept in vitro for 10–14 days in high extracellular potassium concentration (15 mM) to chronically depolarize them or in neurons transfected with the light-activated cation channel channelrhodopsin-2 and photostimulated, in these situations the molecular components of the AIS shifted up to 17 μm away from the neuronal perikaryon. These effects were reversible, and when the excitatory neurons were replaced in a 3.5 mM potassium medium, the AIS is relocated at its original place. Therefore, long-term changes in electrical activity can cause important changes in AIS location. Due to the essential role of AIS in the firing rate of the neurons and the correlation between its location and the thresholds for action potential spiking, shifting positions of the AIS appear as a mechanism for homeostatic plasticity for cell autonomous modulation of neuronal excitability (Grubb and Burrone 2010; Kuba et al. 2010).
2.3.7 The Influence of the Environment as a Motor Controlling the Number and the Plasticity of Synapses
One of the postulated principles of a plastic brain is that as a result of its constant exchanges with the exterior world, it should be constantly malleable by external causes. The principal causes encountered during animal life will be either changes in its physical environment or responses to injuries of its CNS. A fortuitous discovery of Mark Rosenzweig and collaborators was to observe modifications of brain circuitry through experience due to changes in the environment encountered by caged rats. As reported by Rosenzweig in a recent (2007) historical review, it was during the late 1950s and beginning of the 1960s that in collaboration with a neurochemist (Edwards Bennett) using acetylcholinesterase (AChE) activity as a parameter considered to be stable that they showed this value was partially dependent on the animal’s experience. The experienced animals as a consequence of their enriched environment were those kept in company with at least 10–12 other rats, in large cages provided with 5–6 objects to play, explore, and climb upon (e.g., wheels, ladders, and small mazes that were changed in location or replaced by new ones two to three times a week); they were considered rats raised in enriched environment. Control animals were raised in standard laboratory conditions, and the third group consisted of partially sensory-deprived rats either raised in isolation or maintained in groups up to three, in small empty cages. In rats kept in enriched environment, an augmentation in AChE activity was correlated with accrual depth and weight of the cerebral cortex, together with a decrease, per examined field, of neurons, glia, and capillaries suggestive of increased dendritic branching (Bennett et al. 1964; Diamond et al. 1964). Therefore, social interactions together with increased exercise were able to boost learning and memory in the subjected rats, as demonstrated with the behavioral tests used, and paralleled the augmentation of chemical and morphological parameters of the cerebral cortices of the experienced rats. More interesting, the behavioral, anatomical, and chemical changes do not only occur at weaning but also in adult full-grown rats after periods as long as 30 days in the enriched environment.
From the viewpoint of our chapter on structural plasticity, the work of William Greenough and his collaborators is also of great interest. While Marian Diamond was measuring parameters linked to the size of the cortex and its cellular elements, such as cortical thickness, diameter of neurons and of their nuclei, number of glial cells, and capillaries, as we referred to above, William Greenough and collaborators were more interested in parameters linked with synaptic connections such as dendritic dimensions and branching and dendritic spines, not only in the visual cortex where the encountered differences between control and experienced animals were the largest (Greenough and Volkmar 1973) but also in the hippocampus and entorhinal cortex (Fiala et al. 1978), basal ganglia (Comery et al. 1995, 1996), and cerebellar cortex (Greenough et al. 1986). The dendritic fields of the neurons of rats kept in enriched environments exceed by 20 % the size of those in control animals (Greenough and Volkmar 1973; Volkmar and Greenough 1972). Working on the adult rat visual cortex kept in the enriched environment (Juraska et al. 1980), they were able to show on Golgi-Cox-stained neurons that the size increase of the dendritic trees of the experienced rats was selective for layer IV stellate cells. In these neurons there was an increase in the number of first-order stem dendrites as well as in the fifth-order distal branches. Structural changes were, therefore, obtained as a result of life for 12 weeks in the enriched environment, but the change was not general but targeted to specific neurons. A few years later, the quantification was extended to the density of synaptic boutons (Turner and Greenough 1985) and the size of some synaptic components in layer IV, mainly the length of the synaptic complexes (Sirevaag and Greenough 1985). Greenough’s group has also shown that the resulting plasticity occurring from exposure to the enriched environment is not age dependent, since it can occur even in aging rats, with a similar increase in synapse number and dendritic branching (Greenough et al. 1986).
In addition, the work of Greenough has also corroborated and expanded on the results of the Rosenzweig team concerning the fact that not only neurons are able to change due to experience, since the enriched environment is also able to affect practically all classes of nonneuronal elements present in the CNS, and result in angiogenesis, increased myelination, and astrocytic hypertrophy with ensheathment of synapses (see in Markham and Greenough 2004).
The interactions between the environment and brain plasticity are very complex and far to be solved. Recently, the group of Gerd Kempermann demonstrated that factors unfolding or emerging during development contribute to individual differences in structural brain plasticity and behavior (Freund et al. 2013). They conclude their article by writing the ways in which living our lives makes us who we are.
2.4 Morphological Neuronal Plasticity Consecutive to Neuronal Lesions
We have just reviewed the evidence that despite the apparent stability of the brain circuits, the adult brain is characterized by constant synaptic remodeling, often accompanied with slight modifications in the size and branching of dendritic trees and of the somatic volume. Now, we want to discuss another kind of structural plasticity, the one taking place as a consequence of lesions that provoke the partial deafferentation of postsynaptic neurons.
It has been known for many centuries that persons suffering from cranial trauma or cerebrovascular problems, although they never recuperate their normal behavior, are capable of progressive improvement during the first months following the instauration of the neurological deficits. Since, contrary to what happens in the peripheral nerves that are able to regenerate, in the CNS, axonal regeneration is abortive (Cajal 1913, 1928), therefore, there must be other types of reparative processes responsible for the observed improvement. One of these possible mechanisms is likely the disposition showed by injured axons to grow locally, so-called axonal sprouting. Not only lesioned axons, if their neurons of origin do not degenerate (retrograde degeneration), are able to sprout, as reported already from the beginning of the twentieth century by Bielschowsky, Nageotte, and Marinesco (see Stahnisch 2003), but also intact nearby axons.
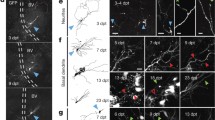
An excellent description of the after stroke improvement was provided by Alf Brodal (Professor of Anatomy at the Medical Faculty in Oslo) of his self observations after a left-sided hemiparesis without hemianopsia resulting from an embolic lesion of the posterior part of the right internal capsule (Brodal 1973). Alf Brodal finished his description saying: “It seems that some of the recovery after a brain lesion may be explained by reinnervation from remaining fibres of synaptic sites which have been denuded as a consequence of the interruption and degeneration of afferent fibres.” It is this process, first reported by Geoffrey Raisman (1969), that was named “terminal sprouting or terminal plasticity” (Sotelo 1975).
2.4.1 Terminal Sprouting
Work carried out with silver impregnation methods showed that the destruction of a source of afferent fibers to a central nucleus causes a compensatory sprouting of the remaining afferents synapsing on the partially denervated neurons (Liu and Chambers 1958). This compensatory process might occur either from collateral sprouting of remote fibers or from a terminal sprouting of nearby fibers. Geoffrey Raisman (1969) was able to settle this dilemma owing to the anatomical arrangement of the afferent fibers entering the septal nuclei. Indeed, these nuclei receive their afferents fibers from two main external sources, the medial forebrain bundle (MFB) and the fimbria. Ultrastructural studies using anterograde degeneration after specific lesions in either of these two kinds of afferent axons revealed that fibers from the MFB established synaptic connections on both dendrites and perikarya, whereas synapses from fimbrial axons were restricted to dendrites. The important observation was that several weeks after destroying one of these afferent fibers, the remaining afferents go beyond their normal boundaries and occupied postlesionally released postsynaptic differentiations (PSDs). Thus, the transection of the MFB induced the formation of axosomatic synapses by fimbrial axons that, according Raisman (1969), had moved from their locations by forming short terminal sprouts that ended by occupying the PSDs vacated after the MFB lesion. These neoformed terminals degenerated after a later destruction of the fimbria, corroborating their initial origin.
A prerequisite for this type of short-length axonal sprouting is that denervated postsynaptic membranes must withhold part of their molecular machinery (receptors and associated transmembrane and cytoplasmic molecules) when they are released from their presynaptic partners so that they would be able to attract intact synaptic boutons from their surroundings. Morphological evidence in favor of the possibility that some PSDs can retain their anatomical structure despite having lost their presynaptic elements was provided by the observation of free PSDs in somatic and dendritic membranes of partially deafferented neurons. Indeed, a frequent observation among electron microscopists trying to identify the origin of axon terminals by anterograde degeneration was the permanence of some normal-looking PSDs after the lesion, for instance, in the avian optic tectum after sectioning of the contralateral optic nerve (Gray and Hamlyn 1962) or after cutting the presynaptic fibers in the frog sympathetic ganglion (Hunt and Nelson 1965), where even the more complex PSDs provided with a subsynaptic bar can persist for over 12 days, strongly suggesting that these structures are not intimately related to the functional integrity of the axon terminals (Sotelo 1968). Examples of persistence of postlesional vacated PSDs in the CNS are numerous, such as the prepyriform cortex (Westrum 1969), the inferior colliculus (Lund 1969), the olfactory bulb (Pinching 1969), or the dorsal column nuclei, where normal-looking, vacated PSDs lasted for at least 6 months (Rustioni and Sotelo 1974).
The attraction exerted by the vacated PSDs and the consequent reinnervation cause the loss of one of the essential qualities of synaptic connections, their specificity, and, therefore, we termed the newly formed synapses “heterologous.” The problem with the presumptive loss of specificity was analyzed in two different central nuclei: the anterior ventral cochlear and the dorsal column nuclei, particularly in the nucleus gracilis (Gentschev and Sotelo 1973; Rustioni and Sotelo 1974). In the auditory nucleus, the primary sensory fibers ended as large axon terminals containing rounded vesicles, establishing asymmetrical synapses that correspond to excitatory terminals of the cochlear nerve. The intact axon terminals were smaller, contained flattened vesicles, and established symmetrical synapses in close proximity to the larger primary sensory terminals. The smaller terminals belonged to other systems of fibers (higher auditory centers, mainly the superior olivary complex with inhibitory action) that moved by a sliding process to reinnervate the vacated PSDs. It can be therefore suggested that this type of synaptic adjustment, with loss of the specificity of the nervous organization, could aggravate the unbalanced state of the primary deafferented neuron. In fact, the loss of excitatory inputs seems to generate an increase in inhibitory inputs on the anterior ventral cochlear neurons. As far as the morphological data can be interpreted, the persistence of free PSDs could therefore be harmful to the function of the neuron, if the newly formed synaptic contacts are functional. The situation is quite similar in the nucleus gracilis, although in this system the large primary sensory axon terminals systematically, and not only occasionally, receive serial synapses from the smaller boutons forming complex synaptic arrangements similar to those encountered in the glomeruli of the substantia gelatinosa of the spinal cord, where the axon terminals of the primary sensory fiber are postsynaptic to the surrounding flattened vesicle containing boutons. The transection of the dorsal roots induced the degeneration of the large primary sensory terminals and the sliding of their presynaptic boutons until their reoccupation of the vacated PSDs. The initial stage of the “reoccupation” of vacated PSDs is characterized by a partial apposition of the bouton-shaped terminal to the free postsynaptic sites; in some instances, the broken presynaptic membrane or a thin layer of astrocytic cytoplasm can be interposed between the deafferented zone and the invading terminal. Finally, the invading terminal develops a new presynaptic vesicular grid facing the deafferented PSD. This last stage is more easily identifiable when the two presynaptic terminals share the vacated PSD, both of them establishing active zones on either half of the PSD.
The fact that only a small fraction of the vacated PSDs persist and succeed in being reinnervated lets us assume the occurrence of a protective mechanism to avoid the harmful effect that, as discussed above, could provoke the formation of the heterologous synapses by unbalancing the required equilibrium between excitatory and inhibitory inputs. Gentschev and Sotelo (1973) reported that in the anterior ventral cochlear nucleus, the sequestration of vacated PSDs could be part of this protective mechanism. In fact, the most common fate of the vacated PSDs is their disappearance by engulfment into the postsynaptic cytoplasm by way of formation of small bags which contain some remnants of presynaptic organelles and which invaginate the surface of the postsynaptic element. From days 5 to 9 these pre- and postsynaptic remnants enter into the postsynaptic cytoplasm by a large pinocytosis, where the remnants of the synaptic complexes become totally engulfed in the neural cytoplasm until their disappearance probably by lysis through the lysosomal system. In the nucleus gracilis a similar mechanism to get rid of vacated PSDs was also disclosed (Rustioni and Sotelo 1974). This last result indicates that membrane sequestration is not specific of the deafferented anterior ventral cochlear nucleus and may be considered as a protective mechanism that exists in other nuclei to regulate the rate of axonal remodeling by terminal sprouting.
2.4.2 Collateral Sprouting from Axotomized and Non-axotomized Axons: Examples After Spinal Cord Injury
In the context of reparative research after traumatic injury, neuroscientists have been impregnated by the idea that axons do not regenerate within the mammalian adult CNS whereas they can in the PNS (Cajal 1928). Thus, until the beginning of this century, most studies dedicated to promote functional recovery after traumatic injury have been focused on how to promote axon regeneration in the adult mammalian CNS (Thuret et al. 2006). It is largely understood that neurons fail to regenerate their axons in the CNS for two reasons: (1) the lack of growth-permissive molecules and/or the presence of growth-inhibitory molecules in the environment of the severed neurons and (2) their inability to reactivate the genetic programs for axon growth (Dusart et al. 2005; Yiu and He 2006; Moore and Goldberg 2011). Thus, in the last decades, there has been a major effort to enhance spinal cord axonal regeneration through a variety of techniques including neutralization of neurite inhibition, administration of neurotrophic factors, and transplantation of a variety of cell types (see for reviews Filbin 2003; Rhodes and Fawcett 2004; Schwab 2004; Silver and Miller 2004; Thuret et al. 2006; Yiu and He 2006). Very surprisingly in many of these experiments, despite an effect on functional recovery, the number of axons that have been observed to regenerate is always low (between 1 % and 10 % when estimated) whatever the experimental approach (sciatic nerve, Aguayo 1985; ensheathing cells, Li et al. 1997; Nogo neutralization Schnell and Schwab 1993; and combination of treatments, Lu et al. 2004). Thus, functional recovery occurs in the presence of a very low number of regenerating axons. For instance, the group of Geoffrey Raisman has estimated that the presence of 1 % of the corticospinal axons beyond the lesion site is sufficient to produce significant functional recovery (Li et al. 1997). One difficulty to interpret these apparently controversial experimental results obtained after spinal cord injury is to determine the anatomical and functional bases of the recorded recovery. Indeed, important processes other than axon regeneration occur after spinal cord injury, either spontaneously or as a reaction to the applied treatment. For example, around the lesion site, demyelination of ascending and descending uninjured fibers has been observed, and over long periods after the lesion, spontaneous remyelination of these non-axotomized fibers by oligodendrocyte progenitors or by Schwann cells that invade the lesion area can contribute to the functional recovery by restoring conduction (Totoiu and Keirstead 2005).
Important components of functional recovery in the absence of regeneration of lesioned axons are the reorganization of circuits that have been spared by the lesion and the formation of new circuits through collateral sprouting of lesioned and non-lesioned axons (for review see Raineteau and Schwab 2001). The group of Martin Schwab has described the spontaneous formation of new circuits through collateralization of severed axons after spinal cord injury (Bareyre et al. 2004). Indeed, after incomplete spinal cord injury in rats, transected hindlimb corticospinal axons form collaterals into the cervical gray matter that contact short and long propriospinal axons. After 12 weeks, the contacts with the short propriospinal axons are no longer detected, whereas the ones with the long propriospinal axons that bridge the lesion site are maintained (Bareyre et al. 2004). Furthermore, treatment after rodent or primate spinal cord injury with Nogo-A neutralizing antibody (Nogo being one of the major myelin axon growth inhibitors present in the CNS) promoted not only axon regeneration of damaged fibers but also sprouting from intact fibers (Liebscher et al. 2005; Freund et al. 2006; Maier et al. 2009). Indeed, when a unilateral section of the corticospinal tract (CST) was performed at the level of the brainstem, axons grew from the contralateral non-lesioned CST and innervated the denervated side of the spinal cord in rats treated with a Nogo-A neutralizing antibody (Thallmair et al. 1998). Treatment after different types of injury with the bacterial enzyme chondroitinase ABC (ChABC) digesting the glycosaminoglycan chains on chondroitin sulfate proteoglycans (another major class of inhibitory molecules in the adult CNS) restores plasticity in adult CNS (Galtrey and Fawcett 2007; Carulli et al. 2010). Furthermore, chondroitinase ABC promotes compensatory sprouting of the intact corticospinal tract and recovery of forelimb function following unilateral pyramidotomy in adult mice (Starkey et al. 2012). Last, it has been recently described that the combination treatment caused a further increased of both axon regeneration and formation of collaterals from unsectioned axons (Zhao et al. 2013).
Although further works are necessary to unravel the importance of collateral formation in behavior recovery, it is an import point to consider when therapeutic treatments are proposed for spinal cord injury recovery.
2.5 Repair of Adult Central Nervous System by the Way of Cell Transplantations
2.5.1 Introduction
Despite the presence of neural stem cells in the CNS, from analysis of numerous genetic, traumatic, vascular, or experimental lesions, it is a certainty that a complete spontaneous restoration of the destroyed neurons and circuits along with disappearance of all neurological symptoms is not possible. For this reason, neuroscientists have been trying for over a century to bypass this limit by cell therapy, transplanting neurons—at the early times of transplantation—and stem cells during the last 20 years (immortalized cell lines, Renfranz et al. 1991, Snyder et al. 1992; adult neural stem cells, Arvidsson et al. 2002; and embryonic stem cells, McDonald et al. 1999, Björklund et al. 2002) to substitute for the missing neurons and palliate the caused impairments.
In the earliest experimental trials (Thompson 1890; Saltykow 1905; Del Conte 1907), the donor material was taken from postnatal animals, and a few days after the operations, all transplanted neurons had died, leaving only the nonneuronal supporting structures. Ranson (1909, 1914) reported the first successful transplantation of the dorsal root ganglia to the brain, even if the ganglia were taken from rats of at least 1 month of age. In his temporal longitudinal studies, Ranson showed that among the surviving cells the form of the spinal ganglion cell is not stable and fixed but is capable of undergoing marked alteration in a short space of time, similarly to what Nageotte (1906) described for pathological conditions of the human dorsal root ganglion cells in “tabes dorsalis” or in experimental animals after transplantation into the rabbit ear parenchyma (Nageotte 1907) in which the unipolar ganglion cells are transformed into multipolar irritated cells.
2.5.2 Transplantation of Embryonic Nervous Tissue into the Adult Brain
The real history of neural cell transplantation into the adult mammalian brain with a therapeutic aim started last century, at the middle of the 1970s (Stenevi et al. 1976; Björklund et al. 1976). The pioneering work of Gopal Das (Das and Altman 1971) and the work of Anders Björklund and Ulf Stenevi (1976, 1979) have defined the main conditions for the survival of transplanted cells (mainly the age of the donor cells that must be taken at the end of the proliferative period of the neuronal population to replace). Even though the results obtained with this approach (the grafting of embryonic solid pieces or isolated cell suspensions into the lesioned adult brain) have not been as successful as expected, for our purpose of the study of structural plasticity, they have been extremely useful. Indeed, this tool has allowed us to appraise the impact that embryonic neural cells can exert on mature neurons and vice versa. In normal conditions this confrontation of young neurons in a mature tissue only happens for the few neurons originating from the neural stem cells in the subventricular zone and the hippocampus. The main question to be solved was as follows: are embryonic neurons able to properly interact with adult neurons leading to their normal development and subsequent integration into the deficient adult neural circuits? But before addressing this question, let us make a short digression to discuss the normal way central neural circuits are organized in the mammalian brain.
2.5.2.1 Global Versus Point-to-Point Systems
Two organizational levels characterize the neuronal arrangement in the central nervous system. The vast majority of the central circuits, those involved in the transfer of specific patterned information such as sensory and motor systems, are organized following a plan of precise cell-to-cell connections that we define as “point-to-point” systems (Sotelo and Alvarado-Mallart 1986) and are based on the absolute specificity of synaptic contacts established between the constitutive neurons. Superimposed on this kind of basic circuits, there are some others considered to be modulatory or regulatory systems (concerning the peptidergic and particularly the monoaminergic system: noradrenergic, dopaminergic, serotonergic, and cholinergic) and that we coined “global systems” owing to their diffuse projection throughout the CNS (Sotelo and Alvarado-Mallart 1986). In addition, these “global systems” establish few morphologically detectable synaptic connections. For instance, in the serotonergic system of the cerebellar cortex, less than 9 % of its varicosities were involved in the establishment of “synaptic complexes”—the morphological correlate of the chemically transmitting synapses—and were considered as non-junctional terminals (Beaudet and Sotelo 1981) releasing their neurotransmitter in a paracrine fashion, as described for other regions of the CNS (Mobley and Greengard 1985). The neurotransmitter diffused through the extracellular space over large distances, acting on those neurons equipped with adequate receptors and located within its precise diffusional volume. Our “point-to-point” and “global” systems were renamed a few months later: “wiring transmission” and “volume transmission,” respectively (Agnati et al. 1986).
The interest of this distinction is that the grafted neurons in “global systems” exert their functional effects by paracrine release involving diffuse secretion of deficient neurochemicals in appropriate terminal areas within the host brain, without the necessity of the formation of specific synaptic connections between grafted and host neurons. By contrast, functional recovery following transplantation in “point-to-point” systems requires a much more precise morphological reconstruction involving precise regulation of the migration and the formation not only of afferent but mainly of efferent synaptic connections of grafted neurons, in such a way that the transplanted cells replace the missing cells. It is probably by this anatomical difference that transplantation has been much more broadly used in “global” than in “point-to-point” systems because the aims to fill are less constrictive in the former than in the latter. For instance, in animal models of Parkinson disease, grafting experiments have been over 10 times more numerous than in those of cerebellar ataxias.
The first indication that grafts can exert some positive effects in the improvement of the neurological symptoms originated in neurotoxic, traumatic, or genetic animal models of neurodegenerative diseases was obtained in experiments conducted in 1979 by Björklund and Stenevi. In these experiments, the “global system” constituted by the nigrostriatal pathway of adult rats was destroyed by the neurotoxin 6OHDA injected stereotaxically. This lesion reproduced symptoms corresponding to the clinical signs of the Parkinson disease. Björklund and Stenevi (1979) were able to show that grafts of DA-producing neural tissue (mesencephalic substantia nigra anlage) taken from rat fetuses, implanted into the striatum, could reduce some of the deficits caused by the 6OHDA intraparenchymal injection. For us, it is not casual that the history of modern neural cell transplantation started with the cell substitution in a “global system” because, as stated before, the possibility of functional replacement does not require a rebuilding of a mirror image of the destroyed neural circuit.
2.5.2.2 Transplantation in Murine Cerebellar Mutants as a Model of Grafting in “Point-to-Point” Systems
The relative simplicity and perfect geometrical arrangement of the cerebellar cortex, with few neuronal populations and inputs from various cell groups making synaptic connections at the precise site of the dendritic tree of the PCs—the unique output from the cortex—offer an excellent model to test the capacity of embryonic PCs to interact with the neural elements of the impaired adult circuit of the mutant cerebellum, to replace the missing PCs of the host. The model system we chose was the replacement of missing PCs in the cerebellum of the Purkinje cell degeneration (pcd) mutant strain of mice (Mullen et al. 1976). Nna1, a gene encoding a putative nuclear protein containing a zinc carboxypeptidase domain, is the mutated gene in the pcd mouse (Fernandez-Gonzalez et al. 2002). In this mutant, the PC degeneration takes place after weaning, when most of the developmental process of the cerebellar cortex and the corticonuclear connection are finished (Sotelo 2004). At 2 months, less than 1 % of the normal PC population remains, and by 4 months almost all have degenerated, remaining an average of 107 PCs in the whole cerebellum, mainly located in the nodulus (Wassef et al. 1986).
Sotelo and Alvarado-Mallart (1986) first described how grafted PCs migrate into the molecular layer of the host cerebellum replacing the missing neurons of the pcd mutant. Following the time course of the development of the embryonic day 12 (E12) cerebellar primordium into the postnatal day 60 (P60) cerebellum, it was possible to show the stages followed by the grafted PCs to be integrated in the cortical circuitry of the adult host cerebellum 4 days after transplantation (Sotelo and Alvarado-Mallart 1987). It was evident that the grafted PCs begin migrating to their proper domain into the host cerebellum, and even though they do not become aligned in a row as in the control cerebellum, they reach their proper environment dispersed within the host molecular layer. This migration reaches its maximal rate 2–3 days later, and the arrest of the migration takes place in 7 days, indicating that the attractive influences which manage the migration are present even in the adult cerebellum, but the migratory potential of the immature cells weakens in a precise developmental stage either during normal development or after grafting, as if provided with an internal clock. Once in the molecular layer, grafted PCs develop dendritic trees appropriately oriented, perpendicular to the parallel fibers of the host cerebellar cortex, and receive with precision the different inputs not only from cerebellar interneurons (parallel fibers, basket cell axons, and stellate cell axons) but also from extracerebellar climbing fibers with which they establish normal synaptic connections at the appropriated location of the dendrites.
The above-summarized results clearly show that neurons and axons of the host brain retain their capacity for synaptogenesis into adulthood and, similarly, that embryonic grafted neurons are able to pursue their inner tempo for migration and differentiation. In addition, the embryonic grafted PCs by interacting with the adult host neural cells (namely, Bergmann fibers, cerebellar interneurons, relay granule cells, and inferior olivary neurons) have the capacity to become synaptic partners for adult axon terminals. The newly formed synapses are functional as early as 10 days after grafting, when excitatory as well as inhibitory responses were already elicited in these cells by electrical stimulation of the host subcortical white matter (Gardette et al. 1990). Therefore, while the synaptogenesis with excitatory axons occurs in synchrony with that happening during normal development, the one with inhibitory axons occurs much faster. Despite this accelerated synaptogenesis PCs, like in normal development, pass through a phase of multiple innervation by climbing fibers (Crepel et al. 1976), transient phase that takes place also with an advanced time course, since by 15 days after grafting (equivalent to P8 aged PCs) it is already finished. These results clearly emphasize that the embryonic and adult neurons confronted in the transplantation experiments are provided with an extraordinary plastic capacity allowing the proper integration of grafted PCs in the deficient circuit of the pcd cerebellar cortex. With this great potentiality for plasticity, the obvious question is to determine if the cerebellar grafts can improve the motor ataxia in the pcd mice. The answer is that one essential prerequisite remained to be fulfilled before a functional and behavioral improvement could be expected. Indeed, although the deficient circuitry of the cerebellar cortex has been repaired with the synaptic integration of the grafted PCs, the cortico/nuclear projection remains constantly deficient, and only in a few occasions we have been able to disclose a few PC terminals synapsing upon deep cerebellar nuclear neurons.
Nevertheless, the obtained results concerning the integration of the grafted PCs in the cortical circuit are very interesting for our purposes of morphological neural plasticity. Indeed, they reveal a new kind of neural plasticity, the one required for the possible interactions among neural cells of similar phenotypes but of complete different ages, such as young postmitotic with adult neurons. The timing and the nature of the cellular interactions involved in the migration of grafted PCs into the host cerebellum should be similar to those that take place during normal ontogeny, suggesting that, in both cases, the developmental program is regulated autonomously by the participating cells, independent of external factors, as regulated by an internal clock (Sotelo and Alvarado-Mallart 1987; Sotelo et al. 1994). Although little is known about the molecular identity of the signals involved, it is clear that they are developmentally regulated and transiently expressed during ontogenesis, but what about mutually interacting molecular signals when embryonic neurons are grafted into adult brain? This raises the question as to whether the grafted embryonic PCs induce adult host cells, in particular the Bergmann glia, to transiently reexpress the molecular signals needed for their migration and synaptic integration into the host (Sotelo and Alvarado-Mallart 1987) or whether isochronic embryonic astrocytes, bearing the appropriate factors because of their age, also migrate from the graft and acquire the Bergmann fiber phenotype, providing this way substrate for migration of immature PCs. Transplant experiments carried out with donor embryonic cerebellum taken from a transgenic mouse line, which shows an ectopic expression of the hybrid Krox-20/lacZ gene in Bergmann fibers, allowed us to determine that the Bergmann fibers subserving radial migratory pathways to grafted PCs do not originate from the graft and, therefore, that they belong to the host (Sotelo et al. 1994). Moreover, in the developing cerebella the embryonic Bergmann fibers, the migratory substratum to immature PCs, like all other radial glia, express nestin an intermediate filament (Lendahl et al. 1990). This expression is transient, and once all the migratory processes are completed in the molecular layer (PCs, granule cells, and inhibitory interneurons), the expression of nestin is lost. Using a monoclonal antibody against nestin, we were able to reveal that only those Bergmann fibers within the cortical areas containing either the grafts or, particularly, grafted PCs migrating into the molecular layer reexpressed nestin.
The observed spatiotemporal correlation between grafted PC radial migration and nestin expression in the host molecular layer validates that these grafted embryonic neurons induce nestin expression in adult glial fibers. The molecular identities of the signals involved in such kind of interaction remain to be determined. Therefore, despite the actual absence of molecular mechanisms, it was possible to establish that grafted embryonic neural cells can interact with adult neural cells through a new and interesting plastic mechanism that regulates gene expression in neural cells of the adult host recapitulating transient phenotypes associated with specific stages of normal development. This new type of morphological plasticity was named “adaptive rejuvenation.” The occurring changes transiently generate a microenvironment permitting radial migration of the PCs, a necessary pre-requirement for their synaptic integration and the restoration of normal circuitry in the cerebellar cortex of pcd mice.
Transplantation of cerebral cortical embryonic cells into lesioned rodent cortex, initiated by Michel Roger and Afsaneh Gaillard (1998), has provided interesting results. Indeed, as published by Gaillart et al. (2007), when embryonic neocortex taken from transgenic mice overexpressing a green fluorescent protein was grafted into damaged murine adult motor cortex, grafted neurons were able to send axonal projections to specific target nuclei in the motor thalamus and even very long projections to the spinal cord. These important and unexpected results suggest that neural cell transplantation might promote the reconstruction of damaged circuits after adult brain injury. In addition, Pierre Vanderhaegen, Afsaneh Gaillard and collaborators have more recently shown that embryonic stem cells and induced pluripotent stem cells (iPSCs), cultured without added morphogens, can by themselves replace cortical neurons (Gaspard et al. 2008; Espuni-Camacho et al. 2013). These experiments open a new aspect of the requirements of transplanted precursor cells for the integration and repair of adult brain circuits.
2.6 Concluding Remarks
In conclusion, the data reported in this review, although only represent a low percentage of the very numerous interesting papers published on this topic, are aimed to emphasize how the important concept of structural neuronal plasticity has accompanied neuroscientists from the middle of the nineteenth century, even before the appearance of the “neuron doctrine,” considered as the milestone marking the beginning of modern neuroscience. The concept built up by the pioneer workers cited in this review implies that the nervous system is in a continuous remodeling process, by destroying and building up neuronal connections that allow us to consider the brain as a “neotenic” organ. Despite the enormous interest of the asked questions, and the major breakthroughs obtained in recent years as a result of the refinement of cellular imaging and transgenic methods, many of these old questions reviewed here still remain unanswered. For example, what is the real meaning of the maintenance of postsynaptic differentiations occurring only in a portion of the deafferented synapses? Are they indicative of the presence of neurotransmitter receptors or only of their scaffolding proteins? What are the possible deleterious effects of heterologous reafferentation by terminal axonal sprouting? How to get the matching of the new neurotransmitter, frequently GABA, and the former receptor that in most of the examples of eliminated synapses was a glutamate receptor? Hopefully with the arrival of new technologies, new vocations for the structural study of the nervous system will continue to emerge among young researchers, and these young researchers become interested in pursuing the evolution of these concepts and the resolution of old problems.
Abbreviations
- 6OHDA:
-
6-Hydroxydopamine
- AChE:
-
Acetylcholinesterase
- AIS:
-
Axonal initial segments
- ChABC:
-
Chondroitinase ABC
- CLSM:
-
Confocal laser scanning microscopy
- CNS:
-
Central nervous system
- CST:
-
Corticospinal tract
- DA:
-
Dopamine
- E:
-
Embryonic day
- LTP:
-
Long-term potentiation
- MFB:
-
Medial forebrain bundle
- P:
-
Postnatal day
- PC:
-
Purkinje cell
- Pcd:
-
Purkinje cell degeneration mutant strain of mice
- PSDs:
-
Postsynaptic differentiations
- TEA:
-
Tetraethylammonium
- TPLSM:
-
Two-photon laser scanning microscopy
References
Agnati LF, Fuxe K, Zoli M, Ozini I, Toffano G, Ferraguti F (1986) A correlation analysis of the regional distribution of central encephalin and beta-endorphin immunoreactive terminals and of opiate receptors in the adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol Scand 128:201–207
Aguayo AJ (1985) Axonal regeneration from injured neurons in the adult mammalian central nervous system. In: Cotman CW (ed) Synaptic plasticity. Guilford, New York, pp 457–538
Andres KH (1965) Uber die Feinstruktur besonderer Einrichtungen in markhaltigen Nervenfasern des Kleinhirns der Ratte. Z Zellforsch Mikroskop Anat 65:701
Angaut P, Sotelo C (1973) The fine structure of the cerebellar nuclei of the cat. II. Synaptic organization. Exp Brain Res 16:431–454
Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O (2002) Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med 8:963–970
Bailey CH, Kandel ER (1993) Structural changes accompanying memory storage. Annu Rev Physiol 55:397–426
Bain A (1855) The senses and the intellect. John W Parker and Son, London
Bain A (1873) Mind and body. The theories of their relation. D Appleton & Comp, New York
Bareyre FM, Kerschensteiner M, Raineteau O, Mettenleiter TC, Weinmann O, Schwab ME (2004) The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat Neurosci 7:269–277
Bawden HH (1900) A digest and criticism of the data upon which is based the theory of the amoeboid movements of the neurone. J Comp Neurol 10:243–264
Beale L (1862) Note on the minute structure of the grey matter of the convolutions of the brain of man, the sheep, cat, and dog. Proc R Soc Lond 12:671–673
Beale L (1963) Indications of the paths taken by the nerve-currents as they traverse the caudate nerve-cells of the spinal cord and encephalon. Proc R Soc Lond 13:386–392
Beaudet A, Sotelo C (1981) Synaptic remodeling of serotonin axon terminals in rat agranular cerebellum. Brain Res 206:305–329
Bennett EL, Diamond MC, Krech D, Rosenzweig MR (1964) Chemical and anatomical plasticity of brain changes in brain through experience, demanded by learning theories, are found in experiments with rats. Science 146:610–619
Berlucchi G, Buchtel HA (2009) Neuronal plasticity: historical roots and evolution of meaning. Exp Brain Res 192:307–319
Bhatt DH, Zhang S, Gao W-B (2009) Dendritic spines dynamics. Annu Rev Plant Physiol Plant Mol Biol 71:261–282
Björklund A, Stenevi U, Svendgaard NA (1976) Growth of transplanted monoaminergic neurones into the adult hippocampus along the perforant path. Nature 262:787–790
Björklund A, Stenevi U (1979) Reconstruction of the nigrostriatal dopamine pathway by intracerebral nigral transplants. Brain Res 177:555–560
Björklund LM, Sanchez-Pernaute R, Chung S, Andersson T, Chen IY, McNaught KS, Brownell AL, Jenkins BG, Wahlestedt C, Kim KS, Isacson O (2002) Embryonic stem cells develop into functional dopaminergic neurons after transplantation in a Parkinson rat model. Proc Natl Acad Sci U S A 99:2344–2349
Bliss TVP, Lømo T (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J Physiol 232:331–356
Brodal A (1973) Self-observations and neuro-anatomical considerations after a stroke. Brain 96:675–694
Cajal SR (1888) Estructura de los centros nerviosos de las aves. Rev Trim Histol Norm Patol 1:1–10
Cajal SR (1890) À propos des certains éléments bipolaires du cervelet avec quelques détails nouveaux sur l’évolution des fibres cérébelleuses. Int Msch Anat Physiol 7:12–30, French translation of the Spanish paper: Sobre ciertos elementos bipolares del cerebelo joven y algunos detalles más acerca del crecimiento y evolución de las fibras cerebelosas. Gaceta Sanitaria de Barcelona
Cajal SR (1892) Nuevo concepto de la histología de los centros nerviosos. Revista Ciencias Medicas de Barcelona 18:5–68
Cajal SR (1894) The Croonian Lecture: La fine structure des centres nerveux. Proc R Soc Lond 55:444–468
Cajal SR (1895) Algunas conjeturas sobre el mecanismo anatómico de la ideación, asociación y atención. Rev Med Cirug Práct 19:497–508
Cajal SR (1896) Las espinas colaterales de las células del cerebro teñidas con el azul de metileno. Rev Trim Micrográfica 1:123–136
Cajal SR (1913/1914) Estudios sobre la Degeneración y Regeneración del Sistema Nervioso. Tomo I (1913) Degeneración y Regeneración de los Nervios. Tomo II (1914) Degeneración y Regeneración de los Centros Nerviosos. Imprenta Hijos de Nicolas Moya (Madrid)
Cajal SR (1928) Degeneration and regeneration of the nervous system. Oxford Univ, Press (London)
Carpenter WB (1874) Principles of mental physiology, with their applications to the training and discipline of the mind, and study of its morbid conditions. D Appleton & Company, New York
Carulli D, Pizzorusso T, Kwok JCF, Putignano E, Poli A, Forostyak S, Andrews MR, Deepa SS, Glant TT, Fawcett JW (2010) Animals lacking link protein have attenuated perineuronal nets and persistent plasticity. Brain 133:2331–2347
Ceccarelli B, Clementi F, Mantegazza P (1971) Synaptic transmission in the superior cervical ganglion of the cat after reinnervation by vagus fibres. J Physiol Lond 216:87–98
Comery TA, Shah R, Greenough WT (1995) Differential rearing alters spine density on medium-sized spiny neurons in the rat corpus striatum: evidence for association of morphological plasticity with early response gene expression. Neurobiol Learn Mem 63:217–219
Comery TA, Stamoudis CX, Irwin SA, Greenough WT (1996) Increased density of multiple-head dendritic spines on médium-sized spiny neurons of the striatum in rats reared in complex environment. Neurobiol Learn Mem 66(2):93–96
Coombs JS, Curtis DR, Eccles JC (1957) The interpretation of spike potentials of motoneurons. J Physiol 139:198–231
Crepel F, Mariani J, Delhaye-Bouchaud N (1976) Evidence for e multiple innervation of Purkinje cells by climbing fibers in the immature rat cerebellum. J Neurobiol 7:567–578
Das GD, Altman J (1971) Transplanted precursors of nerve cells: their fate in the cerebellums of young rats. Science 173:637–638
DeFelipe J (2006) Brain plasticity and mental processes: cajal again. Nat Rev Neurosci 7:811–817
Del Conte G (1907) Einpfanzungen von Embryonalem Gewebe im Gehirn. Ziegler’s Beitrage zur pathol Anat, u z allg Pathol Bd 42(1):62–67
Dellman HD, Rodriguez EM (1970) Herring bodies; an electron microscopic study of local degeneration and regeneration of neurosecretory axons. Z Zellforsch 111:293–315
Deng W, Aimone JB, Gage FH (2010) New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nature Rev Neurosci 11:339–350
Desmond NL, Levy WB (1990) Morphological correlates of long-term potentiation imply the modification of existing synapses, not synaptogenesis, in the hippocampal dentate gyrus. Synapse 5:139–143
Diamond MC, Krech D, Rosenzweig MR (1964) The effects of an enriched environment on the rat cerebral cortex. J Comp Neurol 123:111–119
Dusart I, Ghoumari AM, Wehrle R, Morel MP, Bouslama-Oueghlani L, Camand E, Sotelo C (2005) Cell death and axon regeneration of purkinje cells after axotomy: challenges of classical hypotheses of axon regeneration. Brain Res Rev 49:300–316
Duval M (1895) Hypohèses sur la physiologie des centres nerveux: théorie histologique du sommeil. Comptes Rendus de la Société de Biologie 2 et 9 Février 47:74–77
Engert F, Bonhoeffer T (1999) Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399:66–70
Espuny-Camacho I, Michelsen K, Gall D, Linaro D, Hasche A, Bonnefont J, Bali C, Orduz D, Bilheu A, Herpoel A, Lambert N, Gaspard N, Péron S, Schiffmann SN, Giugliano M, Gaillard A, Vanderhaeghen P (2013) Pyramidal neurons derived from human pluripotent stem cells integrate efficiently into mouse brain circuits in vivo. Neuron 77:440–456
Fernandez-Gonzalez A, La Spada AR, Treadaway J, Higdon JC, Harris BS, Sidman RL, Morgan JI, Zuo J (2002) Purkinje cell degeneration (pcd) phenotypes caused by mutations in the axotomy-induced gene. Nna1 Sci 295:1904–1906
Fiala BA, Joyce JN, Greenough WT (1978) Environmental complexity modulates growth of granule cells dendrites in developing but not adult hippocampus of rats. Exp Neurol 59:372–383
Fifková E, Anderson CL (1981) Stimulation-induced changes in dimensions of stalks of dendritic spines in the centate molecular layer. Exp Neurol 74:621–627
Fifková E, Van Harreveld A (1977) Long-lasting morphological changes in dendritic spines of dentate granular cells following stimulation of the entorhinal area. J Neurocytol 6:211–230
Filbin MT (2003) Myelin-associated inhibitors of axonal regeneration in the adult mammalian CNS. Nat Rev Neurosci 4:703–713
Finger S (1994) Origins of neuroscience: a history of explorations into brain function. Oxford University Press, New York
Freund P, Schmidlin E, Wannier T, Bloch J, Mir A, Schwab ME, Rouiller EM (2006) Nogo-A-specific antibody treatment enhances sprouting and functional recovery after cervical lesion in adult primates. Nat Med 12:790–792
Freund J, Brandmaier AM, Lewenjohann L, Kirste I, Kritzler M, Krüger A, Sachser N, Lindenberger U, Kempermann G (2013) Emergence of individuality in genetically identical mice. Science 340:756–759
Gaillard A, Gaillard F, Roger M (1998) Neocortical grafting to newborn and adult rats: development, anatomical and functional aspects. Adv Anat Embryol Cell Biol 148:1–86
Gaillard A, Prestoz L, Dumartin B, Cantereau A, Morel F, Roger M, Jaber M (2007) Reestablishment of damaged adult motor pathways by grafted embryonic cortical neurons. Nat Neurosci 10:1294–1299
Galimberti I, Gogolla N, Alberi S, Santos AF, Muller D, Caroni P (2006) Long-term rearrangements of hippocampal mossy fiber terminal connectivity in the adult regulated by experience. Neuron 50:749–763
Galtrey CM, Fawcett JW (2007) The role of chondroitin sulfate proteoglycans in regeneration and plasticity in the central nervous system. Brain Res Rev 54:1–18
Gardette R, Crepel F, Alvarado-Mallart RM, Sotelo C (1990) Fate of grafted embryonic Purkinje cells in the cerebellum of the adult “Purkinje cell degeneration” mutant mouse. II. Development of synaptic responses : an in vitro study. J Comp Neurol 295:188–196
Gaspard N, Bouschet T, Hourez R, Dimidschstein J, Naeije G, van den Ameele J, Espuny- Camacho I, Herpoel A, Passante L, Schiffmann SN, Gaillard A, Vanderhaeghen P (2008) An intrinsic mechanism of corticogenesis from embryonic stem cells. Nature 455:351–357
Geinisman Y, Morrell F, deToledo-Morrell L (1989) Perforated synapses on double-headed dendritic spines: a possible structural substrate of synaptic plasticity. Brain Res 480:326–329
Gentschev T, Sotelo C (1973) Degenerative patterns in the ventral cochlear nucleus of the rat after primary deafferentation. An ultra-structural study. Brain Res 62:37–60
Gerlach J (1871) Von dem Rückenmark. In: Stricker S (ed) Handbuch der Lehre von den Geweben des Menschen und der Thiere. Wilhelm Engelmann, Leipzig, pp 665–693
Golgi C (1873) Sulla struttura della sostanza grigia del cervello. Gazzetta Medica Italiana Lombardia 33:244–246
Gray EG (1959) Electron microscopy of synaptic contacts on dendrite spines of the cerebral cortex. Nature 183:1592–1593
Gray EG, Hamlyn LH (1962) Electron microscopy of experimental degeneration in the avian optic tectum. J Anat 96:309–316
Greenough WT, Volkmar FR (1973) Pattern of dendritic branching in occipital cortex of rats reared in complex environments. Exp Neurol 40:491–504
Greenough WT, McDonald JW, Parnisari RM, Camel JE (1986) Environmental conditions modulate degeneration and new dendrite growth in cerebellum of senescent rats. Brain Res 380:136–143
Grubb MS, Burrone J (2010) Activity-dependent relocation of the axon initial segment fine-tunes neuronal excitability. Nature 465:1070–1074
Grutzendler J, Kasthuri N, Gan W-B (2002) Long-term dendritic spine stability in the adult cortex. Nature 420:812–816
Harris KM, Fiala JC, Ostroff L (2003) Structural changes at dendritic spines synapses during long-term potentiation. Phil Trans R Soc Lond B 358:745–748
Hashimoto PH, Palay SL (1965) Peculiar axons with enlarged endings in the nucleus gracilis. Anat Rec 151:454–455
Hatton GI (1997) Function-related plasticity in hypothalamus. Annu Rev Neurosci 20:375–397
Hebb DO (1949) The organization of behavior. A neuropsychological theory. Chapter 4, “the first stage of perception: growth of the assembly”. Wiley, New York, pp 60–78
His W (1886) Zur Geschichte des menschlichen Rückenmarkes und der Nervenwurzeln. Abhandl Kgl sächsGes. Wissensch math-phys Kl 13:479–513
His W (1889) Die Neuroblasten und deren Entstehung in embryonalen Mark. Abhandl Kgl sächs Ges. Wissensch math-phys K1 15:313–372
Hunt CC, Nelson PG (1965) Structural and functional changes in the frog sympathetic ganglion following cutting of the presynaptic nerve fibres. J Physiol 177:1–20
James W (1890) The principles of psychology, vol 1. Macmillan, London
Jones EG (1994) Santiago Ramón y cajal and the croonian lecture, march 1894. Trends Neurosci 17:190–192
Jones EG (2000) Plasticity and neuroplasticity. J Hist Neurosci 9:37–39
Juraska JM, Greenough WT, Elliott C, Mack KJ, Berkowitz R (1980) Plasticity in adult rat visual cortex: an examination of several cell populations after differential rearing. Behav Neural Biol 29:157–167
Kasai H, Hayama T, Ishikawa M, Watanabe S, Yagishita S, Noguchi J (2010) Learning rules and persistence of dendritic spines. Eur J Neurosci 32:241–249
Koelliker A (1896) Handbuch der Gewebelehre des Menschen und der Thiere, vol II, 6th edn. Wilhelm Engelmann Verlag, Leipzig
Kole MH, Ilschner SU, Kampa BM, Williams SR, Ruben PC, Stuart GJ (2008) Action potential generation requires a high sodium channel density in the axon initial segment. Nat Neurosci 11:178–186
Konorski J (1948) Conditioned reflexes and neuron organization. Cambridge Univ. Press, Cambridge, p 267, Reprinted with a supplementary chapter in 1968, New York: Hafner Publ. Co
Kuba H (2012) Structural tuning and plasticity of the axon initial segment in auditory neurons. J Physiol 590:5571–5579
Kuba H, Ishii TM, Ohmori H (2006) Axonal site of spike initiation enhances auditory coincidence detection. Nature 444:1069–1072
Kuba H, Oichi Y, Ohmori H (2010) Presynaptic activity regulates Na + channel distribution at the axon initial segment. Nature 465:1075–1078
Lai CS, Franke TF, Gan WB (2012) Opposite effects of fear conditioning and extinction on dendritic spine remodeling. Nature 483:87–91
LaMantia AS, Pomeroy SL, Purves D (1992) Vital imaging of glomeruli in the mouse olfactory bulb. J Neurosci 12:976–988
Lendahl U, Zimmerman LB, McKay RD (1990) CNS stem cells express a new class of intermediate filament protein. Cell 60:585–595
Lepousez G, Valley MT, Lledo PM (2013) The impact of adult neurogenesis on olfactory bulb circuits and computations. Annu Rev Physiol 75:339–363
Leuner B, Shors TJ (2004) New spines, new memories. Mol Neurobiol 29:117–130
Li Y, Field PM, Raisman G (1997) Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science 277:2000–2002
Liebscher T, Schnell L, Schnell D, Scholl J, Schneider R, Gullo M, Fouad K, Mir A, Rausch M, Kindler D, Hamers FPT, Schwab ME (2005) Nogo-A antibody improves regeneration and locomotion of spinal cord-injured rats. Ann Neurol 58:706–719
Lin XY, Glanzman DL (1994) Long-term potentiation in Aplysia sensorimotor synapses in cell culture: regulation by postsynaptic voltage. Proc Biol Sci 255:113–118
Liu CN, Chambers WW (1958) Intraspinal sprouting of dorsal root axons; development of new collaterals and preterminals following partial denervation of the spinal cord in the cat. AMA Arch Neurol Psychiatry 79:46–61
Lu P, Yang H, Jones LL, Filbin MT, Tuszynski MH (2004) Combinatorial therapy with neurotrophins and cAMP promotes axonal regeneration beyond sites of spinal cord injury. J Neurosci 24:6402–6409
Lugaro E (1909) Modern problems in psychiatry (English translation of the book published in 1906). The University Press, Manchester
Lund RD (1969) Synaptic patterns of the superficial layers of the superior colliculus of the rat. J Comp Neurol 135:179–208
Maier IC, Ichiyama RM, Courtine G, Schnell L, Lavrov I, Edgerton VR, Schwab ME (2009) Differential effects of anti-nogo-A antibody treatment and treadmill training in rats with incomplete spinal cord injury. Brain 132:1426–1440
Markham JA, Greenough WT (2004) Experience-driven brain plasticity: beyond the synapse. Neuron Glia Biol 1:351–363
Martino G, Pluchino S, Bonfanti L, Schwartz M (2011) Brain regeneration in physiology and pathology: the immune signature driving therapeutic plasticity of neural stem cells. Physiol Rev 91:1281–1304
McDonald JW, Liu XZ, Qu Y, Liu S, Mickey SK, Turetsky D, Gottlieb DI, Choi DW (1999) Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nat Med 5:1410–1412
Mobley P, Greengard P (1985) Evidence for widespread effects of noradrenaline on axon terminals in the rat frontal cortex. Proc Natl Acad Sci U S A 82:945–947
Moore DL, Goldberg JL (2011) Multiple transcription factor families regulate axon growth and regeneration. Devel Neurobio 12:1186–1211
Mugnaini E, Walberg F, Hauglie-Hanssen E (1967) Observations on the fine structure of the lateral vestibular nucleus (Deiters' nucleus) in the cat. Exp Brain Res 4:146–186
Mullen RJ, Eicher EM, Sidman RL (1976) Purkinje cell degeneration, a new neurological mutation in the mouse. Proc Natl Acad Sci U S A 73:208–212
Nageotte J (1906) Note sur la régénération collatérale des neurones radiculaires postérieurs dans le tabes. Compt Rend Soc Biol Paris 60:745
Nageotte J (1907) Greffe de ganglions rachidiens survie des éléments nobles et transformation des cellules unipolaires en cellules multipolaires. Compt Rend Soc Biol Paris 62:63
Paillard J (1976) Réflexions sur l’usage du concept de plasticité en neurobiologie. J Psychol Norm Pathol 1:33–47
Palay SL, Sotelo C, Peters A, Orkand PM (1968) The axon hillock and the initial segment. J Cell Biol 38:193–201
Pinching AJ (1969) Persistence of post-synaptic membrane thickenings after degeneration of olfactory nerves. Brain Res 16:277–281
Purves D, Hadley RD, Voyvodic JT (1986) Dynamic changes in the dendritic geometry of individual neurons visualized over periods up to three months in the superior cervical ganglion of living mice. J Neurosci 6:1051–1060
Rabl-Rückhard H (1890) Sind die Ganglienzellen amöboid? Eine Hypothese zur Mechanik psychischer Vorgänge. Neurologische Centralblatt 7:199–200
Raisman G (1969) Neuronal plasticity in the septal nuclei of the adult rat. Brain Res 14:25–48
Raineteau O, Schwab ME (2001) Plasticity of motor systems after incomplete spinal cord injury. Nat Rev Neurosci 2:263–273
Ranson SW (1909) Transplantation of the spinal ganglion to the brain. Quart Bull Northwestern Univ Med School 11:176
Ranson SW (1914) Transplantation of the spinal ganglion, with observations on the significance of the complex types of spinal ganglion cells. J Comp Neurol 24:547–558
Renfranz PJ, Cunningham MG, McKay RD (1991) Region-specific differentiation of the hippocampal stem cell line HiB5 upon implantation into developing mammalian brain. Cell 66:713–729
Rhodes KE, Fawcett JW (2004) Chondroitin sulphate proteoglycans: preventing plasticity or protecting the CNS? J Anat 204:33–48
Rosenzweig MR (2007) Modification of brain circuits through experience. In: Bermudez-Rattoni F (ed) Neural plasticity and memory: from genes to brain imaging. CRC Press, Boca Raton
Ruediger S, Vittori C, Bednarek E, Genoud C, Strata P, Sacchetti B, Caroni P (2011) Learning-related feedforward inhibitory connectivity growth required for memory precision. Nature 473:514–518
Rustioni A, Sotelo C (1974) Some effects of chronic deafferentation on the ultrastructure of the nucleus gracilis of the cat. Brain Res 28:527–533
Saha B, Jaber M, Gaillard A (2012) Potentials of endogenous neural stem cells in cortical repair. Frontiers Cell Neurosci. doi:10.3389/fncel.2012.00014
Saltykow S (1905) Versuche über Gehirnplantation, zugleich ein Beitrag zur Kenntnis der Vorgänge an den zelligen Gehirne-elementen. Arch Psychiatr Nervenkr 40:329–388
Sandbank U, Lerman P, Geifman M (1970) Infantile neuroaxonal dystrophy: cortical axonic and presynaptic changes. Acta Neuropathol 16:342–352
Schnell L, Schwab ME (1993) Sprouting and regeneration of lesioned corticospinal tract fibres in the adult rat spinal cord. Eur J Neurosci 5:1156–1171
Schwab ME (2004) Nogo and axon regeneration. Curr Opin Neurobiol 14:118–124
Sherrington CS (1897) The central nervous system. In: Foster M (ed) A textbook of physiology, vol III, 7th edn. Macmillan, London
Silver J, Miller JH (2004) Regeneration beyond the glial scar. Nat Rev Neurosci 5:146–156
Sirevaag AM, Greenough WT (1985) Differential rearing effects on Rat visual cortex synapses. II. Synaptic morphometry. Dev Brain Res 19:215–226
Snyder EY, Deitcher DL, Walsh C, Arnold-Aldea S, Hartwieg EA, Cepko CL (1992) Multipotent neural cell lines can engraft and participate in development of mouse cerebellum. Cell 66:33–51
Song A-H, Wang D, Chen G, Li Y, Luo J, Duan S, M-m P (2009) A selective filter for cytoplasmic transport at the axon initial segment. Cell 136:1148–1160
Sotelo C (1968) Permanence of postsynaptic specializations in the frog sympathetic ganglion cells after denervation. Exp Brain Res 6:294–305
Sotelo C (1975) Synaptic remodeling in mutants and experimental animals. In: Vital-Durand F, Jeannerod M (eds) Aspects of neuronal plasticity. Editions de l’INSERM, Paris, pp 167–190, Colloques de l’ INSERM Vol 43
Sotelo C (2003) Viewing the brain through the master hand of Ramón y Cajal. Nat Rev Neurosci 4:71–77
Sotelo C (2004) Cellular and genetic regulation of the development of the cerebellar system. Prog Neurobiol 72:295–339
Sotelo C (2008) Viewing the cerebellum through the eyes of Ramón y Cajal. Cerebellum 7:517–522
Sotelo C, Alvarado-Mallart RM (1986) Growth and differentiation of cerebellar suspensions transplanted into the adult cerebellum of mice with heredodegenerative ataxia. Proc Natl Acad Sci U S A 83:1135–1139
Sotelo C, Alvarado-Mallart RM (1987) Embryonic and adult neurons interact to allow Purkinje cell replacement in mutant cerebellum. Nature 327:421–423
Sotelo C, Angaut P (1973) The fine structure of the cerebellar nuclei of the cat. I. Neurons and neuroglial cells. Exp Brain Res 16:410–430
Sotelo C, Palay SL (1968) The fine structure of the lateral vestibular nucleus in the rat. Neurons and neuroglial cells. J Cell Biol 36:151–179
Sotelo C, Palay SL (1971) Altered axons and axon terminals in the lateral vestibular nucleus of the rat. Possible example of axonal remodeling. Lab Investig 25:653–671
Sotelo C, Alvarado-Mallart RM, Frain M, Vernet M (1994) Molecular plasticity of adult Bergmann fibers is associated with radial migration of grafted Purkinje cells. J Neurosci 14:124–133
Stahnisch FW (2003) Making the brain plastic: early neuroanatomical staining techniques and the pursuit of structural plasticity, 1910–1970. J Hist Neurosci 12:413–435
Starkey ML, Bartus K, Barritt AW, Bradbury EJ (2012) Chondroitinase ABC promotes compensatory sprouting of the intact corticospinal tract and recovery of forelimb function following unilateral pyramidotomy in adult mice. Eur J Neurosci 36:3665–3678
Stenevi U, Björklund A, Svendgaard NA (1976) Transplantation of central and peripheral monoamine neurons to the adult rat brain: techniques and conditions for survival. Brain Res 114:1–20
Tanzi E (1893) I fatti e le induzioni dell’odierna istologia del sistema nervoso. Riv Sper Fren Med Leg 19:419–472
Tellez I, Terry RD (1968) Fine structure of early changes in the vestibular nuclei of the thiamine-deficient rat. Am J Pathol 54:777–794
Thallmair M, Metz GA, Z’Graggen WJ, Raineteau O, Kartje GL, Schwab ME (1998) Neurite growth inhibitors restrict plasticity and functional recovery following corticospinal tract lesions. Nat Neurosci 1:124–131
Thompson WG (1890) Successful brain grafting. NY Medical J 51:701–702
Thuret S, Moon LD, Gage FH (2006) Therapeutic interventions after spinal cord injury. Nat Rev Neurosci 7:628–643
Totoiu MO, Keirstead HS (2005) Spinal cord injury is accompanied by chronic progressive demyelination. J Comp Neurol 486:373–383
Townes-Anderson E, Raviola G (1978) Degeneration and regeneration of autonomic nerve endings in the anterior part of the rhesus monkey ciliary muscle. J Neurocytol 7:583–600
Trommald M, Hulleberg G, Andersen P (1996) Long-term potentiation is associated with new excitatory spine synapses on rat dentate granule cells. Learn Mem 3:218–228
Turner AM, Greenough WT (1985) Differential rearing effects on rat visual cortex synapses. I. Synaptic and neuronal density per neuron. Brain Res 329:195–203
Van Harreveld A, Fifková E (1975) Swelling of dendritic spines in the fascia dentata after stimulation of the perforant fibers as a mechanism of post-tetanic potentiation. Exp Neurol 49:736–749
Volkmar FR, Greenough WT (1972) Rearing complexity affects branching of dendrites in the visual cortex of the Rat. Science 176:1445–1447
Wassef M, Simons J, Tappaz ML, Sotelo C (1986) Non-Purkinje cell GABAergic innervation of the deep cerebellar nuclei : a quantitative immunocytochemical study. Brain Res 399:125–135
Westrum LE (1969) Electron microscopy of degeneration in the lateral olfactory tract and plexiform layer of the prepyriform cortex of the rat. Z Zellforsch 98:157–187
Yiu G, He Z (2006) Glial inhibition of CNS axon regeneration. Nat Rev Neurosci 7:617–627
Zhao RR, Andrews MR, Wang D, Warren P, Gullo M, Schnell L, Schwab ME, Fawcett JW (2013) Combination treatment with anti-Nogo-A and chondroitinase ABC is more effective than single treatments at enhancing functional recovery after spinal cord injury. Eur J Neurosci. doi:10.1111/ejn.12276
Zhao S, Studer D, Chai X, Graber W, Brose N, Nestel S, Young C, Rodriguez EP, Saetzler K, Frotscher M (2012) Structural plasticity of hippocampal mossy fiber synapses as revealed by high-pressure freezing. J Comp Neurol 520:2340–2351
Zuo Y, Lin A, Chang P, Gan W-B (2005) Development of long-term dendritic spine stability in diverse regions of cerebral cortex. Neuron 46:181–189
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Sotelo, C., Dusart, I. (2014). Structural Plasticity in Adult Nervous System: An Historic Perspective. In: Junier, MP., Kernie, S. (eds) Endogenous Stem Cell-Based Brain Remodeling in Mammals. Stem Cell Biology and Regenerative Medicine. Springer, Boston, MA. https://doi.org/10.1007/978-1-4899-7399-3_2
Download citation
DOI: https://doi.org/10.1007/978-1-4899-7399-3_2
Published:
Publisher Name: Springer, Boston, MA
Print ISBN: 978-1-4899-7398-6
Online ISBN: 978-1-4899-7399-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)