
Abstract
Alcohol (ethanol) is a widely used and abused drug with approximately 90% of adults over the age of 18 consuming alcohol at some point in their lifetime. Alcohol exerts its actions through multiple neurotransmitter systems within the brain, most notably the GABAergic and glutamatergic systems. Alcohol’s actions on GABAergic and glutamatergic neurotransmission have been suggested to underlie the acute behavioral effects of ethanol. The striatum is the primary input nucleus of the basal ganglia that plays a role in motor and reward systems. The effect of ethanol on GABAergic and glutamatergic neurotransmission within striatal circuitry has been thought to underlie ethanol taking, seeking, withdrawal and relapse. This chapter reviews the effects of ethanol on GABAergic and glutamatergic transmission, highlighting the dynamic changes in striatal circuitry from acute to chronic exposure and withdrawal.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Alcoholism is a progressive and chronic relapsing disorder that ultimately leads to detrimental health outcomes. Studies have revealed adaptations to cortico-basal ganglia circuits that mediate the stages of the addiction cycle. This includes initial drug use to habitual and continued use despite negative outcomes. The transition to addiction involves neuroplasticity in these brain regions that begin in the mesolimbic dopamine region and transition to the dorsal striatum (Ito et al. 2002; Everitt and Robbins 2013, 2016). Although great progress has been made in ethanol pharmacology demonstrating that acute ethanol has only a few known primary targets (Vengeliene et al. 2008), it has long been proposed that the acute behavioral effects of ethanol are mediated principally by potentiation of γ-aminobutyric acid A (GABAA) receptors and/or inhibition N-methyl-d-aspartate (NMDA) receptors. This presumption is due to the similarities in behavioral effects between ethanol and benzodiazepines that act on GABAA receptors, as well as NMDA antagonists such as ketamine (Krystal et al. 2003). An imbalance in the striatum of GABAergic and glutamatergic transmission is thought to play a role in alcohol use and abuse. Understanding how ethanol alters GABAergic and glutamatergic systems through the progression to alcohol addiction within specific brain regions/circuits will provide valuable insights for developing finely targeted therapeutics.
2 GABAergic Synapses
GABA is the major inhibitory neurotransmitter in the brain. GABA is derived from glutamate by the enzyme glutamic acid decarboxylase (GAD). There are two GAD isoforms, GAD65 and GAD67, named for their molecular weights. Since GAD is required for the synthesis of GABA, it is commonly used as a marker for GABAergic neurons. Once synthesized, GABA is packaged into vesicles by the vesicular GABA transporter. The release of GABA is regulated by calcium concentration within the axon terminal, with increased concentration leading to vesicular release until basal calcium concentrations are restored.
GABA exerts its actions on transmission by GABAA ionotropic receptors and GABAB metabotropic receptors. GABAA receptors are heteropentomeric complexes that form a ligand-gated anion-selective channel that is permeable to chloride and bicarbonate. The GABAA receptor can be found both pre- and postsynaptically (Lovinger 2017). In mature neurons, the concentration of chloride is lower intracellularly leading to a net flow of anions into the neuron upon channel opening. Therefore, activation of GABAA receptors results in membrane hyperpolarization that decreases the excitability of a cell as chloride ions flow into the cell (Olsen and Sieghart 2008). In mammals, there are 19 different GABAA receptor subunits identified: α (1–6), β (1–3), γ (1–3), δ, ε, ρ (1–3), θ, and π (Olsen and Sieghart 2008, 2009). In addition to the 19 subunits, there are also subunit splice variants and phosphorylation states that can modify the activation of specific subunits. Most mammalian receptors consist of 2 α, 2 β, and 1 γ subunit. The subunits composing a GABAA receptor dictate their biophysical and pharmacological properties and location within the brain, as well as cellular distribution. Sensitivity to GABA is determined primarily by the α subunit expressed. GABAA receptors can possess a variety of allosteric modulatory sites also dependent on the subunits expressed, that allow modulators such as benzodiapines, neurosteroids, and barbiturates to alter their function. Most receptors containing the γ2 subunit are targeted to the synapse via their interaction with the scaffolding protein gephyrin (Farrant and Nusser 2005; Fritschy et al. 2012). The exception to this rule is α5βxγ2 receptors, which are targeted to the extrasynapse by the interaction of the α5 subunit and radixin (Loebrich et al. 2006). Receptors containing the δ subunit are localized exclusively in the extrasynapse (Walker and Semyanov 2008; Belelli et al. 2009; Herd et al. 2013). Posttranslational modifications of GABAA receptors regulate trafficking and stability. For example, Protein Kinase-A (PKA) phosphorylation of the β3 subunit leads to internalization of the receptor complex, whereas Protein Kinase C (PKC) phosphorylation of multiple subunits leads to membrane insertion (Kittler et al. 2005; Mele et al. 2014). Within the striatum, GABAA receptor subunits (α1–5, β1–3, γ1–3, and δ) are expressed to varying degrees, with the α2 and β3 subunits having the highest degree of immunoreactivity. The α2βxγ1/2 isoform is the most highly expressed isoform in the striatum and is located within the synaptic and extrasynaptic cellular compartments of striatal medium-sized GABAergic projection neurons (MSNs) (Schwarzer et al. 2001; Maguire et al. 2014). The α4βxδ isoform of the GABAA receptors is found exclusively extrasynaptically in MSNs and interneurons (Schwarzer et al. 2001; Maguire et al. 2014).
The GABAB receptors are Gi-protein-coupled receptors. When activated, they mediate inhibition by activating potassium channels and decreasing calcium conductance. There are two subtypes of the GABAB receptor, GABABR1 and GABABR2, that form homo- and heterodimers in the membrane. GABAB receptors can be located either pre- and postsynaptically. Presynaptic GABAB receptors serve as autoreceptors, regulating the release of GABA. Postsynaptically located GABAB receptors are primarily coupled to the activation of potassium channels (Misgeld et al. 2007), whose activation serves to hyperpolarize the cell, albeit at a slower time scale than GABAA receptors.
GABA transmission is terminated when GABA is cleared from the synapse. Reuptake of GABA is mediated by GABA transporters located on the plasma membrane of both neurons and glia. Following uptake by both cell types, GABA is degraded by the enzyme GABA transaminase into glutamate.
3 Glutamatergic Synapse
Glutamate is the major excitatory neurotransmitter in the brain. Glutamate is synthesized by two different mechanisms: the first is synthesis from glucose through the Krebs cycle by transamination of α-ketoglutarate. Alternatively, glutamate is formed directly from glutamine by the glutaminase enzyme. Glutamate is packaged into synaptic vesicles by vesicular glutamate transporters.
The postsynaptic actions of glutamate are mediated by ionotropic and metabotropic glutamate receptors. The ionotropic glutamate receptors, N-methyl-d-aspartate (NMDA), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), and kainic acid (KA) receptors, are glutamate-gated cation channels.
NMDA receptors are voltage-sensitive ionotropic glutamate receptors that when open allow for the flow of calcium and/or sodium ions, albeit at slower kinetics than AMPA and KA receptors. NMDA receptors are regulated by presynaptic glutamate release and postsynaptic mechanisms such as phosphorylation states, subunit expression, and membrane potential that all contribute to its roles in neuronal plasticity, stabilizing neuronal activity, and coincidence detection (Malenka and Nicoll 1999; Wang 1999; Yuste et al. 1999). The voltage sensitivity aspect of NMDA receptors is due to the blockade at resting membrane potentials by magnesium, which is removed by membrane depolarization. NMDA receptors are tetramers that consist of an obligatory NR1 subunit and regulatory NR2 (A–D) and/or NR3 (A–B) subunits. The regulatory subunits control the biophysical (conductance and open probability) and pharmacological properties of NMDA receptors (Wenzel et al. 1997; Traynelis et al. 2010). NMDA receptors are unique in that they require the binding of two different ligands, glutamate and glycine/d-serine, for activation. NR1 subunits bind glycine or d-serine, while the NR2 subunits bind glutamate (Gonda 2012). NMDA receptors play a role in synaptic plasticity mainly in the form of long-term potentiation that is associated with increases in membrane insertion of AMPA receptors, protein synthesis, spine formation, and the enlargement of existing spines (Malinow and Malenka 2002; Matsuzaki et al. 2004; Kasai et al. 2010). Phosphorylation state of NMDA receptors plays a role in their localization, activation state, and physiological properties (Traynelis et al. 2010).
The AMPA and KA glutamate receptors are also heterotetrameric protein complexes that form ligand-gated ion channels. AMPA receptors consist of GluR1–4 (also known as GluRA-D), GluRδ1, and GluRδ2 (Dingledine et al. 1999). Each GluR subunit contains a binding site for glutamate. Although AMPA receptors are capable of allowing the flow of calcium, sodium, and potassium, the majority of AMPA receptors contain the GluR2 subunit which renders the ion channel impermeable to calcium. AMPA receptors mediate most of the excitatory transmission in the brain due to its vast brain expression as well as lack of voltage sensitivity (as in the case of NMDA receptors). AMPA receptors have been well studied and shown to play a role in synaptic plasticity. KA receptor subunits include GluR5–7, KA1, and KA2 (Dingledine et al. 1999) and their activation leads to the flow of sodium and potassium ions, and consequently membrane depolarization.
Metabotropic glutamate receptors (mGluRs) are G-protein-coupled receptors. mGluRs are divided into three familial groups. Group I mGluRs (mGluR1 and mGluR5) are Gq-coupled receptors whose binding of glutamate leads to activation of the enzyme phospholipase C (PLC) that ultimately induces the release of calcium from intracellular stores and increases PKC activity (Kenny and Markou 2004). Group1 mGluRs are mainly located on the postsynaptic membrane. mGluR 1 has moderate expression in the dorsal striatum and low expression in the NAc, whereas mGluR5 is highly expressed in the entire striatum (Olive 2009; Pomierny-Chamiolo et al. 2014). Group II mGlurRs (mGluR2 and mGluR3) are Gi/o-coupled receptors whose activation decreases the activity of adenylyl cyclase, ultimately decreasing the intracellular concentration of cyclic adenosine monophosphate (cAMP) (Kenny and Markou 2004). They are present both pre- and postsynaptically. Lastly, Group III mGlurRs (mGluR4, GluR6, GluR7, and GluR8) are similar to Group II mGluRs in that they are Gi/o-coupled receptors (Kenny and Markou 2004). These receptors are primarily located presynaptically and play a role in regulating neurotransmitter release. Of the Group III mGluRs, mGluR4, mGluR7, and mGluR8 are expressed in the striatum (Corti et al. 2002; Messenger et al. 2002; Bragina et al. 2015).
Similar to GABA, glutamate transmission is terminated when glutamate is cleared from the synapse. Glutamate is predominately taken up through plasma membrane transporters that are located on glia, mainly astrocytes, and to a lesser extent, on neurons. Once transported intracellularly, glutamate is metabolized to glutamine.
Corticostriatal GABAergic synapses can undergo synaptic plasticity by both long-term potentiation (LTP) and long-term depression (LTD). Corticostriatal LTP requires the activation of NMDA and D1Rs (Calabresi et al. 2000; Kerr and Wickens 2001), while LTD requires activation of postsynaptic mGluRs and presynaptic CB1 receptors (Calabresi et al. 2000).
4 The Striatum and Action Control
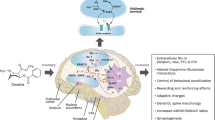
Cortico-basal ganglia loops play a role in the learning and selection of appropriate action sequences, as well as detecting deviations within the sequence and changes in the outcome. Cognitive control is required to guide the selection of appropriate actions based on an individual’s current goals and situation, and at the same time inhibiting unwanted actions. The striatum, the main input of the basal ganglia, is innervated from all regions of the cortex. The striatum is divided into the dorsomedial striatum (DMS, roughly equivalent to the caudate nucleus in primates), dorsolateral striatum (DLS, roughly equivalent to the putamen nucleus in primates), and ventral striatum. The ventral striatum can be divided into the nucleus accumbens (NAc) and the olfactory tubercle (Heimer and Wilson 1975). Although, the striatum receives input from all regions of the cortex, specific cortical regions send inputs to distinct striatal regions. The striatum then sends converging projections to the output nuclei, the internal globus pallidus (GPi) and substantia nigra pars reticulata (SNpr). The output nuclei project to the thalamus that sends projections back to the cortical input regions, completing the loop (Alexander et al. 1986). Cortico-basal ganglia circuits can be divided based on their cortical inputs and their roles in learning, control, and performance of actions (Haber et al. 2000). These parallel circuits compete for the control of behavior and are suggested to be connected in a spiraling manner whereby MSNs in one striatal region project to the ventral tegmental area (VTA)/SN that send projections to MSNs of another striatal region, starting from the NAc medial shell then to the NAc core/DMS and ultimately to the DLS (Fig. 1). Dopaminergic innervation of the DLS is thus under the influence of the NAc.

Model of cortico-striatal circuitry focusing on the limbic (green), associative (red), and sensorimotor (blue) circuits. The nucleus accumbens (NAc) receives glutamatergic input from the limbic cortices, hippocampus (yellow) and amygdala (yellow), and dopaminergic projections (gray dashed lines) from the ventral tegmental area (VTA). It then projects to the VTA. The dorsomedial striatum (DMS), or the caudate nucleus in primates, receives glutamatergic input from the associative cortices and dopaminergic input from the VTA and substantia nigra (SN). It then projects to the SN. The dorsolateral striatum (DLS), or the putamen nucleus in primates, receives glutamatergic input from the sensory and motor cortices and dopaminergic input from the SN. It then projects to the SN. The VTA projects back to the NAc and the DMS, while the SN sends projections back to the DMS as well as the DLS
Most alcohol users are casual consumers in which alcohol is consumed for its rewarding properties. In these individuals, alcohol consumption is thought to be a goal-directed action, in which alcohol use is dependent on the value of the outcome. Therefore, if alcohol consumption is devalued, for example it has become associated with unwanted intoxication/illness, alcohol use will cease. However with repeated, chronic use, alcohol drinking may transition to a habit in which an associated cue triggers voluntary alcohol consumption despite negative repercussions. This is highlighted by continued alcohol abuse by habitual alcoholics despite its negative outcomes (personal, social or financial). Drugs of abuse, including alcohol, are thought to induce abnormally strong consolidation of instrumental learning mechanisms, enhancing drug seeking behavior in response to cues or contexts that are associated with the drug. This may occur by drug-induced cortico-basal ganglia circuit plasticity that reinforces the connectivity within the “spiral,” leading to the recruitment of dorsal striatum circuitry proposed to underlie this transition from casual to habitual alcohol use and seeking (Koob and Volkow 2010).
4.1 Nucleus Accumbens: Motivation and Reinforcement
The limbic cortico-basal ganglia circuit is commonly referred to as the brain reward circuit (Haber 2011), with the nucleus accumbens (NAc) as its main basal ganglia input (Fig. 1 green pathway). The NAc can be divided into two subregions, the core and shell, based on MSN morphology, neurochemistry, projection patterns, and functions (Heimer et al. 1991; Zahm and Brog 1992; Meredith 1999).
The NAc core receives most of its input from glutamatergic projections from prelimbic medial prefrontal cortex (mPFC), hippocampus and amygdala (Groenewegen et al. 1999). It is also thought to be continuous with the DMS and therefore is implicated in conditioned responding, sensory motor integration, and emotional cues (Carlezon et al. 1995; Rodd-Henricks et al. 2002; Sellings and Clarke 2003; Ikemoto 2007; Guo et al. 2009; Suto et al. 2010). The NAc core sends GABAergic projections to the dorsolateral ventral pallidum and the substantia nigra (Zahm and Heimer 1990; Heimer et al. 1991; Zhou et al. 2003).
The NAc shell receives dopaminergic input from the VTA and glutamatergic projections from infralimbic mPFC, the basolateral amygdala, and ventral hippocampus (Britt et al. 2012; Papp et al. 2012). The NAc shell in conjunction with the bed nucleus of the stria terminalis and central amygdala have collectively been referred to as the extended amygdala complex, due to similarities in morphology and circuitry (Hopkins and Holstege 1978; Heimer and Alheid 1991). The extended amygdala complex has been implicated in the emotional processing of stimuli and drug addiction (Koob 2013). MSNs of the NAc shell project to the ventromedial ventral pallidum and the ventral tegmental area (Zahm and Heimer 1990; Heimer et al. 1991; Zhou et al. 2003). The NAc shell is implicated in reward processing, the control of motivation, behaviors by primary rewards, and behavioral inhibition such as aversion learning (Kravitz et al. 2012; Hikida et al. 2013).
With regards to alcohol use, ethanol increases the release of dopamine in the NAc. This increase in dopamine is believed to mediate the positive reinforcing effects of ethanol (Imperato and DiChiara 1986). The ventral striatum is also suggested to play a role in drug-induced increase in locomotion and cue-induced alcohol use.
4.2 DMS: Goal-Directed Behaviors
The DMS in the rodent, roughly equivalent to the primate caudate nucleus, is one of the subdivisions of the dorsal striatum that lines the lateral ventricle. It is part of the associative circuit that includes the prefrontal, entorhinal, and posterior parietal cortical projections to the medial striatum (Fig. 1, red pathway). The DMS also receives dopaminergic inputs from the substantia nigra and the VTA and glutamatergic inputs from the basolateral amygdala and thalamus (Haber et al. 2000; Ikemoto 2007; Pan et al. 2010; Corbit et al. 2013; Kupferschmidt et al. 2015). MSNs of the DMS project to the substantia nigra pars reticulate, subthalamic nucleus, and globus pallidus.
As part of the associative circuit, the DMS is thought to influence goal-directed actions based on the expected consequences or value of the actions, also referred to as action-outcome (Yin et al. 2005a, b; Gunaydin and Kreitzer 2016). Goal-directed actions are flexible in that their performance is sensitive to changes in value or motivation for the outcome as well as changes in the contingency between the action and outcome.
Studies using an outcome devaluation test that examines whether a goal-directed or habitual strategy is used to perform an instrumental action (i.e. nose poke or lever press) suggest that early alcohol use is a goal-directed action (Corbit et al. 2012, 2014). In this task, rodents are trained to perform an instrumental action for alcohol. During the outcome devaluation test, the value of the alcohol reward is changed by either pairing it with an aversive stimuli, satiation to alcohol, or changing the contingency between the action and alcohol reward. With extended exposure to the action-alcohol pair, self-administration shifts from a goal-directed action sensitive to the change in the value of the alcohol reward to a value-insensitive habitual action that occurs regardless of a change in the value of the alcohol reward. (Corbit et al. 2012; Lopez et al. 2014). Pharmacological inactivation of the DMS, but not the DLS, led to a loss in the sensitivity to alcohol devaluation in mice suggesting that the DMS is important for goal-directed alcohol self-administration (Corbit et al. 2012).
4.3 DLS: Habit Formation
The sensorimotor circuit includes primary and secondary sensory and motor cortices that project to the DLS (equivalent to the primate putamen nucleus), a subdivision of the dorsal striatum (Fig. 1, blue pathway). The DLS also receives convergent dopaminergic input from the substantia nigra. The proposed role of the sensorimotor circuit is the maintenance and execution of well-learned actions that rely on external and internal cues and less on the changes in the consequences of the action often referred to in conditioning terms as stimulus-response control over behavior (Webster 1961; McGeorge and Faull 1989; Yin et al. 2006). DLS MSNs, in turn, send GABAergic projections to the substantia nigra, globus pallidus, and subthalamic nucleus.
As part of the sensorimotor cortico-striatal loop, the DLS is involved in habit formation or stimulus-response learning. A habitual behavior is an overlearned action associated with a specific cue or context. When triggered by that cue or context the habit will be performed automatically regardless of the outcome. Habitual actions require less executive control than goal-directed actions (Dalley et al. 2004; Muller et al. 2007), and therefore allows the habit system to react quickly yet it also makes it inflexible.
The habit system is thought to be strengthened under alcohol conditions in which executive control over drug taking is decreased. The DLS is also implicated in the drug-induced stereotypies as well as habitual and compulsive alcohol seeking (Everitt et al. 2008; Corbit et al. 2012, 2014). Relapse is also thought to involve the DLS (Fuchs et al. 2006). During chronic alcohol use DLS output is potentiated while the circuits involved in associative learning, such as the DMS, are altered (Belin-Rauscent et al. 2012; Hogarth et al. 2013).
4.4 Subtypes of Striatal Neurons
The striatum contains medium-sized GABAergic spiny projection neurons (MSNs), GABAergic interneurons, and cholinergic interneurons. MSNs are the principal cells of the striatum, constituting 90% of the neuronal population in rodents and 70% in primates (Kemp and Powell 1971; Kita and Kitai 1988). As the sole output neurons of the striatum, MSNs process and integrate information from cortex, thalamus, limbic structures, VTA/SN, and neighboring striatal neurons.
MSNs can be further divided into two classes based on their axonal projections, dopamine receptor expression, peptide expression, and electrophysiological properties. MSNs that express the dopamine D1 receptor, co-express dynorphin, substance P, and M4 cholinergic receptors, and directly project to the substantia nigra, are referred to as direct pathway MSNs (dMSNs) (Augood et al. 1997; Gerfen and Surmeier 2011). MSNs that express the dopamine D2 receptor, co-express enkephalin and neurotensin, and indirectly projects to striatal output regions are referred to as indirect pathway MSNs (iMSNs) (Le Moine and Bloch 1995; Surmeier et al. 1996; Augood et al. 1997; Aubert et al. 2000; Gerfen and Surmeier 2011). A small proportion of MSNs co-express D1 and D2 receptors in rodents and primates (Le Moine and Bloch 1995; Aubert et al. 2000). Direct pathway MSNs promote actions by disinhibiting the thalamus and cortex, whereas iMSNs “stop” actions by indirectly disinhibiting the SN (Kravitz et al. 2010). The balance between dMSN and iMSN activity is required for normal reward-related behaviors (Fig. 2 top). The basal electrophysiological properties of dMSN and iMSN neurons differ in the striatum, such that iMSNs are more excitable than dMSNs (Kreitzer and Malenka 2008; Grueter et al. 2010; Planert et al. 2013). Similarly, there is an increase in glutamate release onto iMSNs as compared with dMSNs (Kreitzer and Malenka 2008; Grueter et al. 2010). Although both MSN subtypes can undergo LTD including those that are NMDA-dependent, endocannabinoid-mediated, and transient receptor potential cation channel (TRPV) 1-dependent, LTD is more robust in iMSNs (Kreitzer and Malenka 2007; Grueter et al. 2010). Dopamine also leads to differential actions on dMSNs and iMSNs. When dopamine is released in the striatum, D1 receptor activation on dMSNs results in long-term potentiation (LTP) of synaptic efficacy. At the same time, D2 receptor activation prevents LTP on these synapses (Reynolds and Wickens 2002).

Hypothetical model of the effects of acute and chronic alcohol on glutamatergic and GABAergic transmission onto the striatal subregions (left) and direct and indirect pathway MSNs of the striatum (right). Under normal conditions, glutamatergic (blue) and GABAergic transmission (orange) are balanced. This balance in output from the DLS and DMS/NAc as well as between dMSNs and iMSNs leads to normal action selection. Under acute alcohol exposure, data suggests that there is an overall hypoglutamatergic state in the striatum. However, there is a differential alcohol-induced effect on GABAergic transmission between striatal subregions: the acute ethanol-induced decrease in GABAergic transmission in the DLS and increase in GABAergic transmission in the DMS/NAc may lead to an imbalance in control of action selection between the striatal subregions. Under chronic alcohol exposure, there is a hyperglutamatergic state in dMSNs that is thought to lead to LTP of glutamatergic input onto dMSNs. In iMSNs, a decrease in NMDA receptors may lead to a decrease in glutamatergic transmission or LTD, concomitant with an increase in GABAergic transmission. Ultimately this is thought to bias towards activation of dMSNs and abnormal control over alcohol consumption
Although a majority of dMSNs and iMSNs are distinguished by expression and output regions, it should be noted that they are not entirely segregated. In the NAc, dMSNs and iMSNs are not as clearly segregated as in the dorsal striatum, with a larger proportion of MSNs co-expressing D1 and D2 receptors (Bertran-Gonzalez et al. 2008; Kupchik et al. 2015). dMSNs have been shown to send axon collaterals to the globus pallidus external and the ventral pallidum (Lu et al. 1998; Zhou et al. 2003; Fujiyama et al. 2011; Kupchik et al. 2015).
In addition to MSNs, approximately 4% of rodent striatal neurons are locally projecting GABAergic interneurons. Striatal GABAergic interneurons are medium sized, aspiny neurons and can be divided into three subtypes based on their electrophysiological properties and protein expression. Fast-spiking interneurons express the calcium binding protein parvalbumin and exhibit rapid and sustained firing (Cowan et al. 1990; Kawaguchi et al. 1995; Tepper and Bolam 2004). They provide the strongest input onto MSNs with approximately 100 connections targeting the soma and proximal dendrites of neighboring MSNs (Koós and Tepper 1999). Due to their high degree of synchronization, vast number of connections, and proximity of synapses close to or on the soma, fast-spiking interneurons can greatly control action potential firing in MSNs (Kita 1993; Bennett and Bolam 1994; Koós and Tepper 1999; Kubota and Kawaguchi 2000; Tunstall et al. 2002; Mallet et al. 2005). Low threshold-spiking interneurons co-express a combination of neuropeptide Y, somatostatin, and nitric oxide synthase (Smith and Parent 1986). They have lower action potential firing rates than fast-spiking interneurons and exhibit plateau potentials. Synapses formed between low threshold-spiking interneurons and MSNs tend to be more apical on MSN dendrites than is true of fast-spiking interneurons. The last subpopulation of striatal GABAergic interneurons express calretinin and are considered to be low-threshold spiking interneurons (Tepper and Bolam 2004). This group of interneurons are less understood due to their limited numbers and the overlapping expression of calretinin on a subset of MSNs. MSNs receive both feed forward and lateral inhibition from GABAergic interneurons and recurrent collaterals from neighboring MSNs, respectively.
The remaining striatal neurons are large cell bodied (20–50 μm) and aspiny interneurons that release the neurotransmitter acetylcholine (Bolam et al. 1984; Smith and Bolam 1990; Wilson et al. 1990). These interneurons are tonically active and have a relatively depolarized resting membrane potential (Wilson et al. 1990; Kawaguchi et al. 1995). Due to their large size and vast dendritic and axonal fields, they are suggested to integrate and modulate synaptic connections (Kawaguchi et al. 1995). Although cholinergic interneurons and cholinergic transmission play a large role in striatal circuitry and alcohol addiction (Clarke and Adermark 2015; Gonzales and Smith 2015), the effects of ethanol on cholinergic transmission are not discussed in this chapter.
5 Ethanol Actions on GABAergic Transmission
Ethanol has long been thought to exert its effects through potentiating the GABAergic system. This is due to similarities in the behavioral effects between ethanol and benzodiazepines, such as sedation, decreases in anxiety, and ataxia. The activity of both ionotropic and metabotropic GABA receptors have been shown to be important for ethanol reinforcement and relapse to ethanol seeking (Augier et al. 2017).
5.1 Acute Actions
There has been mixed data regarding the acute effects of ethanol on GABAA receptor function. Alcohol is considered to be an allosteric modulator of GABAA receptors that enhances GABAA receptor activity by increasing the probability of channel opening or increasing agonist affinity (Nestoros 1980; Suzdak et al. 1986; Tonner and Miller 1995; Zhou et al. 1998; Welsh et al. 2009; Soyka et al. 2016). Other studies, however, found a lack of ethanol effect on GABAA receptor properties (Gage and Robertson 1985; Siggins et al. 1987; White et al. 1990). The discrepancy between studies has been suggested to occur because of discrepant subunit composition, brain location, or phosphorylation state. In regards to subunit composition, it has been proposed that there is a specific ethanol binding site on the GABAA receptor located between the α and β3 subunit interface, or between the transmembrane region between TM2 and TM3. If this is the case, then it is safe to assume that GABAA receptors expressing a specific subunit composition profile will be sensitive to ethanol. Low concentrations of ethanol have been shown to act directly on extrasynaptic GABAA receptors containing α4/6β3δ (Wallner et al. 2006; Olsen et al. 2007), but this observation was contradicted elsewhere (Borghese et al. 2006; Botta et al. 2007). Several studies have demonstrated an involvement of PKC and its phosphorylation of subunits on the sensitivity of GABAA receptors to ethanol (Aguayo and Pancetti 1994; Weiner et al. 1994, 1997; Qi et al. 2007).
Failure to find a consistently observed direct ethanol effect on GABAA receptors suggests that the effect of ethanol on GABAergic transmission is perhaps at least partially via a presynaptic mechanism. Several studies have demonstrated an ethanol-induced increase in GABA release in various brain regions including the hippocampus, VTA, amygdala, spinal cord, cerebellum, and striatum (Crowder et al. 2002; Melis et al. 2002; Roberto et al. 2003; Ziskind-Conhaim et al. 2003; Ariwodola and Weiner 2004; Carta et al. 2004; Nie et al. 2004; Sanna et al. 2004; Siggins et al. 2005; Li et al. 2006; Ming et al. 2006; Zhu and Lovinger 2006; Criswell et al. 2008; Silberman et al. 2008; Theile et al. 2008; Wilcox et al. 2014). However not all studies have found an ethanol-induced alteration in GABA release (Proctor et al. 2006; Criswell et al. 2008). With regards to the striatum, the effects of GABAergic transmission appear to be subregion specific as it has been shown that acute ethanol increases the frequency of mIPSCs in the DMS and NAc but conversely decreases it in the DLS (Nie et al. 1997, 2000; Wilcox et al. 2014).
The regulation of GABA release by acute ethanol is thought to rely on activation of G-protein-coupled receptors including GABAB receptors (Peris et al. 1997; Nie et al. 2004; Wu et al. 2005; Silberman et al. 2009; Kelm et al. 2011). Acute ethanol has been shown to increase presynaptic GABAB receptor activity, suggesting its role in the ethanol-induced changes in GABA release. The presynaptic GABAB effect is thought to involve tonic inhibition of PKC. An ethanol-induced effect on postsynaptically located GABAB receptors has not been found (Ariwodola and Weiner 2004). The ethanol-induced alterations in presynaptic GABA release are further discussed in the chapter “Presynaptic Ethanol Actions: Potential Roles in Ethanol Seeking” found in this volume (Lovinger 2017).
5.2 Chronic Actions
Chronic ethanol exposure can result in tolerance to alcohol’s behavioral effects. This may reflect neural adaptations that ultimately decrease GABAergic transmission (Fig. 2, bottom). Changes in GABAA receptor subunit expression have been found in several brain regions coincident with chronic alcohol exposure (e.g., Papadeas et al. 2001; Cagetti et al. 2003; Floyd et al. 2004; Hemby et al. 2006; Jin et al. 2014). Specifically, in the NAc a decrease in the α4 subunit was found following 2 weeks of chronic ethanol drinking in rats compared to ethanol naïve rats (Papadeas et al. 2001). A different group demonstrated within the NAc a coordinated decrease in the protein and functional expression of the α1 and δ subunits, concomitant with an increase in α4, α5, and γ2 subunits, using a chronic intermittent ethanol exposure model followed by up to 40 days of withdrawal (Liang et al. 2014). This is suggestive of long-lasting adaptations in GABAergic transmission following chronic ethanol exposure. These differences may be due to length of exposure or withdrawal. In ethanol-dependent individuals engaged in prolonged withdrawal, lower GABAA receptor availability is found in the NAc of dependent individuals compared to controls (Lingford-Hughes et al. 2012). These adaptive changes in GABAA receptor subunit expression have also been found in other brain regions such as the hippocampus (Cagetti et al. 2003; Liang et al. 2004) and amygdala (Floyd et al. 2004; Roberto et al. 2004; Anderson et al. 2007). These alterations may induce changes in the functional properties of GABAA receptors leading to changes in affinity for GABA and allosteric modulators.
A few studies have reported that chronic ethanol exposure alters GABA release. An increase in GABA release was suggested to occur within the CeA and hippocampus of chronic ethanol treated rodents perhaps attributable to changes in the activity of GABAB autoreceptors (Tremwel et al. 1994; Peris et al. 1997; Roberto et al. 2004, 2008, 2010). Specifically, in the dorsal striatum a decrease in the frequency of mIPSCs was found in both the DLS of mice and in the putamen of monkeys (Cuzon Carlson et al. 2011, 2017; Wilcox et al. 2014). However, the role of GABAB receptors in modulating GABA release was not examined in these studies.
5.3 Pharmacotherapies for Alcohol Use Disorders That Target GABAergic Transmission
To date, there are only three medications approved by the US Food and Drug Administration (FDA) to treat alcohol use disorder: disulfiram, naltrexone, and acamprosate. Although these drugs have been shown to reduce alcohol consumption, their effects are modest and inconsistent. Therefore, there is a need to discover other pharmacological treatments need to be explored. The observed behavioral similarities between the effects of ethanol and benzodiazepines (Krystal et al. 2003) suggest that targeting the GABAergic system is a viable target for treating alcohol use disorder. There are three pharmacological agents that are thought to alter the GABAergic system that are currently being assessed for their ability to treat AUD: gabapentin, baclofen, and sodium oxybate. Gabapentin is structurally similar to the GABA, although it has no activity at GABA receptors. It is shown to have activity at voltage sensitive calcium channels and the ability to modulate GAD, increasing GABA synthesis (Taylor 1997). Gabapentin is currently FDA approved for the treatment of seizures, neuropathic pain, and restless leg syndrome. For alcohol use disorders, gabapentin has been shown to increase the rate of abstinence, decrease heavy drinking days, and decrease withdrawal symptoms (Voris et al. 2003; Mason et al. 2014). Baclofen is an agonist of the GABAB receptor and is currently FDA approved for muscle spasticity. Clinical studies have shown an increase in abstinence and a decrease in alcohol consumption in AUD individuals using baclofen over placebo (Addolorato et al. 2007; Muller et al. 2015; Beraha et al. 2016; Reynaud et al. 2017). Sodium oxybate is the salt version of gamma-hydroxybutyrate (GHB), an endogenous neurotransmitter that has agonist activity at GABA receptors (Kamal et al. 2016). Sodium oxybate is FDA approved for the treatment of narcolepsy and is currently approved for alcohol relapse prevention in Italy and Austria. Clinical trials suggest that sodium oxybate increases abstinence rates (Gallimberti et al. 1992; Caputo et al. 2007) but may lead to craving and abuse of GHB (Caputo et al. 2007).
6 Ethanol Actions on Glutamatergic Transmission
The glutamatergic system has been implicated in the acute intoxicating effects of ethanol, ethanol dependence, and withdrawal, and recently has been suggested to be a potential target for treatment of alcoholism. These acute intoxicating effect correlates with a decrease in glutamatergic function. There is evidence that changes in brain circuitry occurring as a result of chronic alcohol exposure leads to a hyperglutamatergic state (Fig. 2). The glutamatergic system plays a role in alcohol-associated dependence, including chronic alcohol seeking and relapse (Dahchour et al. 1998; Rossetti et al. 1999; Bäckström and Hyytiä 2004; Krupitsky et al. 2007; Nagy 2008; Alasmari et al. 2015).
6.1 Acute Actions
Acute ethanol has been shown to elicit both pre- and postsynaptic effects that ultimately lead to decreased glutamatergic transmission. Presynaptically, acute low concentrations of ethanol elevate glutamate levels in the striatum (Moghaddam and Bolinao 1994; Selim and Bradberry 1996; Lominac et al. 2006; Szumlinski et al. 2007; Soyka et al. 2016; Goodwani et al. 2017; Hopf 2017) whereas at acute higher concentrations ethanol can decrease extracellular glutamate concentrations (Moghaddam and Bolinao 1994; Piepponen et al. 2002; Tiwari et al. 2014). However, other studies suggest no acute ethanol effect on glutamate levels (Dahchour et al. 1994, 1996; Quertemont et al. 2002). This discrepancy may be due to strain differences, differences in ethanol sensitivity, concentration of ethanol examined, or brain location. Nevertheless, it is proposed that ethanol can alter extracellular glutamate levels by exerting an effect on glutamate uptake by astrocytes (Smith 1997; Othman et al. 2002; Melendez et al. 2005) or by the effect of high concentrations of ethanol inhibiting the release of glutamate by its action on NMDA receptors (Martin and Swartzwelder 1992; Woodward 1994).
Postsynaptically, ethanol has been shown to alter the functioning of ionotropic and metabotropic glutamate receptors. NMDA receptor, an ionotropic glutamate receptor, is one of the major targets of ethanol (Lovinger et al. 1989, 1990; Holmes et al. 2013) and inhibition of NMDA receptors by alcohol is thought to contribute to the intoxicating effects of alcohol (Hodge and Cox 1998). Acute ethanol at concentrations that mimic intoxicating levels in humans (5–50 mM or ~23–230 mg/dL) has been shown to inhibit the function of NMDA receptors in a concentration-dependent manner (Lovinger et al. 1989, 1990). This ethanol-induced inhibition of NMDA receptors is mediated by a decreased probability of channel opening and a decrease in mean open time (Lima-Landman and Albuquerque 1989; Weight et al. 1993).
The ethanol sensitivity of NMDA receptors is thought to be dependent on subunit composition, regional differences, phosphorylation state, and extracellular concentration of magnesium. NMDA receptors containing the NR2A or NR2B subunits are most potently affected by ethanol, with less potency for NMDA receptors containing the NR2C or NR3 subunits (Kuner et al. 1993; Yamakura et al. 1993; Masood et al. 1994; Chu et al. 1995; Mirshahi and Woodward 1995; Popp et al. 1998; Woodward 2000; Smothers and Woodward 2003). Fyn kinase, PKA, PKC, and DAARP-32 have been shown to phosphorylate NMDA receptors after ethanol administration (Moon et al. 1994; Snell et al. 1994; Miyakawa et al. 1997; Maldve et al. 2002; Li and Kendig 2003; Ferrani-Kile et al. 2003; Yaka et al. 2003) that may lead to the internalization of NR2 subunits (Suvarna et al. 2005). With regards to extracellular magnesium concentration, the inhibitory effect of ethanol on NMDA receptors correlates with the concentration of magnesium such that increasing concentrations of magnesium leads to an increase in ethanol-induced inhibition of NMDA receptors (Rabe and Tabakoff 1990; Martin et al. 1991; Morrisett et al. 1991; Calton et al. 1998). The ethanol sensitivity of NMDA receptors is also sensitive to glycine concentration, an allosteric modulator of the NMDA receptor. High concentrations of glycine (>10 μM) can decrease the ethanol-induced inhibition of NMDA receptors (Rabe and Tabakoff 1990). In the striatum, acute ethanol can inhibit the synaptic plasticity of excitatory postsynaptic currents in an ethanol concentration-dependent manner (Wang et al. 2007; Jeanes et al. 2011).
In addition to its effects on NMDA receptors, ethanol also inhibits the function of AMPA and kainate receptors (Moghaddam and Bolinao 1994; Costa et al. 2000; Crowder et al. 2002). AMPA and kainate ionotropic glutamate receptors are also sensitive to the acute effects of ethanol, albeit at concentrations exceeding those that block NMDA receptors (Lovinger et al. 1989; Dildy-Mayfield and Harris 1992; Costa et al. 2000; Moykkynen et al. 2003; Kalev-Zylinska and During 2007; Marty and Spigelman 2012; Santerre et al. 2014). A single 4-h two-bottle choice session in which mice had access to ethanol (20% v/v) and water leads to an increase in the protein level of the AMPA receptor subunit GluA1 (Beckley et al. 2016).
The role of Group I metabotropic glutamate receptors (mGluR1, mGluR5) in alcohol-related behaviors has been extensively studied. Gene variations in mGluR5 are associated with alcoholism risk (Schumann et al. 2008). Similarly, mGluR5 has been suggested to play a role in alcohol consumption and seeking as the blockade or deletion of mGluR5 specifically in the ventral striatum, attenuates these behaviors (Besheer et al. 2010; Cozzoli et al. 2012; Sinclair et al. 2012).
6.2 Chronic Actions
Chronic alcohol exposure leads to a hyperglutamatergic state in many brain regions, including the striatum (Ward et al. 2009; Ding et al. 2012, 2013; Das et al. 2015). In response to chronic ethanol, there is a potentiation of glutamatergic transmission (Fig. 2, bottom), potentially as a compensation to chronic blockade of NMDA by acute ethanol.
In postmortem brains of human alcoholics, studies have found an increase in NMDA receptor ligand binding, density, and affinity (Michaelis et al. 1993; Freund and Anderson 1996, 1999). Similar findings were observed in rodent studies of chronic ethanol exposure in which binding to the NMDA receptor was increased concomitant with an increase in the expression of the NR1, NR2A, and NR2B subunits of the NMDA receptor (Trevisan et al. 1994; Kumari and Ticku 2000; Kash et al. 2009; Obara et al. 2009; Wang et al. 2010). In addition to up-regulation of NMDA subunit expression, chronic ethanol also increases NMDA receptor function (Kalluri et al. 1998; Carpenter-Hyland et al. 2004; Carpenter-Hyland and Chandler 2006; Kash et al. 2009; Wang et al. 2007, 2010) and conductance (Iorio et al. 1992; Sanna et al. 1993; Chen et al. 1999; Floyd et al. 2003; Nagy et al. 2003; Nelson et al. 2005). In chronic ethanol-exposed mice that were undergoing withdrawal, high frequency stimulation induced NMDA receptor-dependent LTP of glutamatergic transmission in the striatum. This was in stark contrast to the ethanol naïve condition in which the same high frequency stimulation paradigm induced NMDAR-dependent LTD of glutamatergic transmission (Yamamoto et al. 1999; Jeanes et al. 2011). The increase in LTP may be facilitated by an increase in the response of NR2B containing NMDA receptors (Wang et al. 2007). A decrease in LTD of glutamatergic transmission was also found in the striatum of chronic ethanol exposed rodents potentially through decreased endocannabinoid cannabinoid 1 receptor signaling (Xia et al. 2006; DePoy et al. 2013). These changes in NMDA receptor expression and function can lead to hyperexcitability that may underlie the increased seizure susceptibility associated with early withdrawal from ethanol (Tsai et al. 1995; Tsai and Coyle 1998). In prolonged withdrawal, a down-regulation of NMDA subunit express, function, and LTD induction have been observed in the NAc (Abrahao et al. 2013).
A chronic ethanol-induced increase in AMPA receptor expression has been found in a number of brain regions, including the striatum (Chandler et al. 1999; Neasta et al. 2010; Ary et al. 2012; Wang et al. 2012). Specifically within the striatum, an increase in the expression and synaptic trafficking of GluA1 and GluA2 subunits of the AMPA receptor was found following chronic ethanol (Neasta et al. 2010; Ary et al. 2012). An increase in AMPA receptor-mediated excitatory postsynaptic currents was observed in the DMS and amygdala following chronic ethanol (Läck et al. 2007; Ma et al. 2017). The chronic ethanol-induced facilitation of LTP observed in striatal MSNs has been suggested to involve an increase in the synaptic insertion of AMPA receptors (Wang et al. 2010, 2015).
In addition to changes in the postsynaptic ionotropic glutamate receptors, an increase in the postsynaptic release of glutamate and its concentration in brain tissue have also been observed following chronic ethanol exposure in rodents and humans (Rossetti and Carboni 1995; Bauer et al. 2013). Elevated glutamate concentrations were measured in rodents after chronic ethanol exposure in several brain regions, and the glutamate concentration measured in the NAc, in particular, correlated with the severity of alcohol withdrawal (Fliegel et al. 2013). An increase in the spine density of MSNs as well as an increase in the frequency of excitatory postsynaptic currents were observed in the putamen of nonhuman primates (Cuzon Carlson et al. 2011). The observed chronic ethanol-induced increase in extracellular glutamate may be a result of alterations in vesicular glutamate transporters (Tsai and Coyle 1998; Fliegel et al. 2013). An increase in the expression and/or activity of mGluR1/5 may also increase glutamate release by way of activation of PLC (Obara et al. 2009; Meinhardt et al. 2013).
6.3 Pharmacotherapies of Alcohol Use Disorders That Target the Glutamatergic System
As stated above, there are currently only three FDA drugs for the treatment of AUD. Acamprosate was approved by the FDA in 2004 for the maintenance of abstinence in individuals with AUD (Maisel et al. 2013; Jonas et al. 2014; Donoghue et al. 2015). Although its exact mechanism of action is unknown, acamprosate is thought to reduce glutamate levels via antagonism of mGluR5 (Harris et al. 2003). Topiramate has shown potential as a potential AUD therapy drug. It is an antagonist of kainate and AMPA glutamate receptors (Gibbs et al. 2000) and is currently FDA approved to treat epilepsy. Topiramate has shown potential in reducing alcohol cravings and intake in human trials (Johnson et al. 2007; Blodgett et al. 2014; Martinotti et al. 2014). However, a number of adverse effects (including nausea, impaired cognitive function, and paraesthesia) has the potential to lead to noncompliance in some users as these adverse effects may outweigh the benefit of the drug (Kranzler et al. 2014).
7 Effects of Ethanol on Specific Neuronal Populations of the Striatum
Since it has been demonstrated that specific manipulation of either direct or indirect pathway MSNs leads to distinct downstream circuits mediating different behaviors (Kravitz et al. 2010; Carvalho Poyraz et al. 2016; Lambot et al. 2016), it is important to define these and other striatal cell types and highlight differences in alcohol-induced adaptations.
7.1 dMSN vs iMSNs
Evidence is mounting that GABAergic and glutamatergic transmissions within the striatum are modulated by ethanol in a cell-type specific manner. Repeated cycles of voluntary ethanol consumption and forced withdrawal selectively potentiate synaptic NMDA receptor activity in D1 receptor-expressing dMSNs but not in D2 receptor-expressing iMSNs of the DMS (Cheng et al. 2017). This imbalance in activity between dMSNs and iMSNs following chronic ethanol exposure occurs in conjunction with a loss in the ability to elicit LTD selectively in dMSNs, while that ability was observed only in iMSNs (Fig. 2, bottom). Although this was recovered by 2 weeks of withdrawal (Jeanes et al. 2014), it is not known whether the occlusion of LTD in dMSNs following chronic ethanol was due to an ethanol-induced floor effect in LTD whereby it could not be further generated, or whether ethanol altered synaptic function in some manner rendering it resistant to change. Another study suggests that the bias towards activation of dMSNs is a result of chronic ethanol-induced facilitation of NMDA receptor currents and LTP while concomitantly inhibiting NMDA receptors and eliciting LTD in iMSNs (Fig. 2, bottom; Renteria et al. 2017). A further explanation is the increase in GluN2B-containing NMDA receptors specifically in dMSNs may facilitate AMPA receptor plasticity, LTP and an overall increase in activity of dMSNs (Wang et al. 2012, 2015).
Interestingly, this potentiation of glutamatergic transmission onto dMSNs with ethanol exposure was found to be concomitant with an increase in GABAergic transmission onto iMSNs (Fig. 2, bottom; Cheng et al. 2017). This increase in GABAergic transmission in the striatum may result from an ethanol-induced increase in GABAergic interneuron connectivity onto D2-MSNs (Gittis et al. 2011).
On the whole, the literature suggests a strong bias towards activation of dMSNs following chronic ethanol exposure that may be responsible for facilitating continued, compulsive, or excessive alcohol intake (Berglind et al. 2006; Luo et al. 2011). In agreement with an ethanol-induced increase in dMSN output, genetic knockout or blockade of glutamate receptors specifically in dMSNs that results in a decrease in output from dMSNs, reduces the alcohol deprivation effect in which there is a temporary increase in voluntary alcohol consumption over baseline when ethanol access is reinstated following a period of withdrawal (Sinclair and Senter 1967, 1968; Eisenhardt et al. 2015).
7.2 Interneurons
There are conflicting reports as to how ethanol affects fast-spiking parvalbumin expressing interneurons of the striatum. An acute ethanol-induced depression of GABAergic synapses in the DLS was found to specifically involve the GABAergic synapses of fast-spiking parvalbumin-expressing GABAergic interneurons onto MSNs through modulation of opioid transmission (Patton et al. 2016). Conversely, acute application of ethanol led to a reversible membrane depolarization in striatal fast-spiking GABAergic interneurons through a reduction in cholinergic transmission (Blomeley et al. 2011). This difference in results may have been due to species differences or differences in recording parameters. In low-threshold spiking interneurons, brief application of ethanol led to membrane hyperpolarization (Blomeley et al. 2011), an effect attributed to the ethanol’s actions on potassium currents.
Acute ethanol also decreased the average action potential firing rate of cholinergic interneurons (Blomeley et al. 2011). Cholinergic interneurons have also been shown to facilitate the acute ethanol effect on long-term synaptic plasticity in the striatum (Adermark et al. 2011).
8 Implications of the Effect of Ethanol on Striatal GABAergic and Glutamatergic Transmission in the Progression to Addiction
Repeated cycles of ethanol consumption and withdrawal are thought to reinforce ethanol consumption, in some instances leading to pathologically excessive use of ethanol. Gaining insights into the detailed mechanisms that underlie the control of ethanol consumption by excitatory and inhibitory neurotransmission onto striatal neuronal subpopulations is a required step in elucidating prospective synaptic and neuronal therapeutic targets for the development of new approaches for the treatment of alcoholism. A further goal would be uncovering differences in the cortico-basal ganglia circuits (limbic, associative, and somatosensory) to determine what sets the stage for ethanol consumption progressing to excessive, compulsive intake in some individuals while others are spared.
Acute ethanol leads to a reduction in excitatory drive by way of the inhibition of NMDA receptors. This coupled with the enhancement of inhibitory GABAergic transmission may account for the sedating and dose-dependent depressant effects of ethanol intoxication. In the striatum, the acute effects of ethanol on GABAergic transmission appeared to be subregion specific with increases in presumably the release of GABA in the DMS and NAc but a decrease in the DLS (Nie et al. 1997, 2000; Wilcox et al. 2014). This may lead to a disruption in the normal processing of reward-related behaviors mediated by the limbic circuit, or action control mediated by the dorsal striatum that may begin a bias towards the somatosensory circuit. It is posited that abnormal reward-related learning and action selection for the consumption of alcohol is brought about by ethanol-induced changes in striatal synaptic strength and plasticity. This could ultimately prime the activity of the same striatal circuits in response to future alcohol exposure. The alcohol-induced changes in neuronal signaling within the NAc could explain the erroneous cue-induced associations that are made by individuals during alcohol exposure. The activity of the DMS appears to be required during the developmental phase of excessive alcohol drinking in which the action of alcohol intake is still dependent on the outcome of that action (Corbit et al. 2012). In a related fashion, the DMS may play a role in the relapse to alcohol seeking.
One consequence of chronic ethanol exposure appears to be a disinhibition of striatal output that facilitates the recruitment of certain cortico-striatal circuits. In particular, the sensorimotor circuit may increase habit formation and alcohol seeking (Corbit et al. 2012; Dickinson et al. 2002). The resulting recruitment of the sensorimotor loop and the DLS is thought to underlie compulsive drug use. This may be further exacerbated by the alcohol-induced bias in activation of dMSNs via increased AMPAR activity. The activation or potentiation of dMSN circuitry concomitant with a decrease in iMSN circuitry could reduce the threshold of alcohol-related sensory stimulation and enhance multisensory integration surrounding alcohol use.
References
Abrahao KP, Ariwodola OJ, Butler TR, Rau AR, Skelly MJ, Carter E, Alexander NP, McCool BA, Souza-Formigoni ML, Weiner JL (2013) Locomotor sensitization to ethanol impairs NMDA receptor-dependent synaptic plasticity in the nucleus accumbens and increases ethanol self-administration. J Neurosci 33:4834–4842
Addolorato G, Leggio L, Ferrulli A, Cardone S, Vonghia L, Mirijello A, Abenavoli L, D’Angelo C, Caputo F, Zambon A, Haber PS, Gasbarrini G (2007) Effectiveness and safety of baclofen for maintenance of alcohol abstinence in alcohol-dependent patients with liver cirrhosis: a randomized, double-blind controlled study. Lancet 370:1915–1922
Adermark L, Clarke RB, Söderpalm B, Ericson M (2011) Ethanol-induced modulation of synaptic output from the dorsolateral striatum in rats is regulated by cholinergic interneurons. Neurochem Int 58:693–699
Aguayo LD, Pancetti FC (1994) Ethanol modulation of the γ-aminobutyric acidA-and glycine-activated Cl-current in cultured mouse neurons. J Pharmacol Exp Ther 270:61–69
Alasmari F, Abuhamdah S, Sari Y (2015) Effects of ampicillin on cysteine/glutamate antiporter and glutamate transporter 1 isoforms as well as ethanol drinking in male P rats. Neurosci Lett 600:148–152
Alexander GE, DeLong MR, Strick PL (1986) Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 9:357–381
Anderson NJ, Daunais JB, Friedman DP, Grant KA, McCool BA (2007) Long-term ethanol self-administration by the nonhuman primate, Macaca fascicularis, decreases the benzodiazepine sensitivity of amygdala GABA(A) receptors. Alcohol Clin Exp Res 31:1061–1070
Ariwodola OJ, Weiner JL (2004) Ethanol potentiation of GABAergic synaptic transmission may be self-limiting: role of presynaptic GABA(B) receptors. J Neurosci 24:10679–10686
Ary AW, Cozzoli DK, Finn DA, Crabbe JC, Dehoff MH, Worley PF, Szumlinski KK (2012) Ethanol up-regulates nucleus accumbens neuronal activity dependent pentraxin (Narp): implications for alcohol-induced behavioral plasticity. Alcohol 46:377–387
Aubert I, Ghorayeb I, Mornamd E, Bloch B (2000) Phenotypical characterization of the neurons expressing the D1 and D2 dopamine receptors in the monkey striatum. J Comp Neurol 418:22–32
Augier E, Dulman RS, Damadzic R, Pilling A, Hamilton JP, Heilig M (2017) The GABAB positive allosteric modulator ADX71441 attenuates alcohol self-administration and relapse to alcohol seeking in rats. Neuropsychopharmacology 42:1789–1799
Augood SJ, Westmore K, Emson PC (1997) Phenotypic characterization of neurotensin messenger RNA-expressing cells in the neuroleptic-treated rat striatum: a detailed cellular co-expression study. Neuroscience 76:763–774
Bäckström P, Hyytiä P (2004) Ionotropic glutamate receptor agonists modulate cue-induced reinstatement of ethanol-seeking behavior. Alcohol Clin Exp Res 28(4):558–565
Bauer J, Pedersen A, Scherbaum N, Bening J, Patschke J, Kugel H, Heindel W, Arolt V, Ohrmann P (2013) Craving in alcohol-dependent patients after detoxification is related to glutamatergic dysfunction in the nucleus accumbens and the anterior cingulate cortex. Neuropsychopharmacology 38:1401–1408
Beckley JT, Laguesse S, Phamluong K, Morisot N, Wegner SA, Ron D (2016) The first alcohol drink triggers mTORC1-dependent synaptic plasticity in nucleus accumbens dopamine D1 receptor neurons. J Neurosci 36:701–713
Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, Cope DW (2009) Extrasynaptic GABAA receptors: form, pharmacology, and function. J Neurosci 29:12757–12763
Belin-Rauscent A, Everitt BJ, Belin D (2012) Intrastriatal shifts mediate the transition from drug-seeking actions to habits. Biol Psychiatry 72:343–345
Bennett BD, Bolam JP (1994) Localization of parvalbumin-immunoreactive structures in primate caudate-putamen. J Comp Neurol 347:340–356
Beraha EM, Salemink E, Goudriaan AE, Bakker A, de Jong D, Smits N, Zwart JW, Geest DV, Bodewits P, Schiphof T, Defoumy H, van Tricht M, van den Brink W, Wiers RW (2016) Efficacy and safety of high-dose baclofen for the treatment of alcohol dependence: a multicenter, randomized, double-blind controlled trial. Eur Neuropsychopharmacol 26:1950–1959
Berglind WJ, Case JM, Parker MP, Fuchs RA, See RE (2006) Dopamine D1 or D2 receptor antagonism within the basolateral amygdala differentially alters the acquisition of cocaine-cue associations necessary for cue-induced reinstatement of cocaine-seeking. Neuroscience 137:699–706
Bertran-Gonzalez J, Bosch C, Maroteaux M, Matamales M, Herve D, Valjent E, Girault JA (2008) Opposing patterns of signaling activation in dopamine D1 and D2 receptor-expressing striatal neurons in response to cocaine and haloperidol. J Neurosci 28:5671–5685
Besheer J, Grondin JJ, Cannady R, Sharko AC, Faccidomo S, Hodge CW (2010) Metabotropic glutamate receptor 5 activity in the nucleus accumbens is required for the maintenance of ethanol self-administration in a rat genetic model of high alcohol intake. Biol Psychiatry 67:812–822
Blodgett JC, Del Re AC, Maisel NC, Finney JW (2014) A meta-analysis of topiramate’s effects for individuals with alcohol use disorders. Alcohol Clin Exp Res 38:1481–1488
Blomeley CP, Cains S, Smith R, Bracci E (2011) Ethanol affects striatal interneurons directly and projection neurons through a reduction in cholinergic tone. Neuropsychopharmacology 36:1033–1046
Bolam JP, Wainer BH, Smith AD (1984) Characterization of cholinergic neurons in the rat neostriatum. A combination of choline acetyltransferase immunocytochemistry, Golgi-impregnataion and electro microscopy. Neuroscience 12:711–718
Borghese M, Storustovu S, Ebert B, Herd MB, Belelli D, Lambert JJ, Marshall G, Wafford KA, Harris RA (2006) The delta subunit of γ-aminobutyric acid type A receptors does not confer sensitivity to low concentrations of ethanol. J Pharmacol Exp Ther 316:1360–1368
Botta P, Radcliffe RA, Carta M, Mameli M, Daly E, Floyd KL, Dietrich RA, Valenzuela CF (2007) Modulation of GABAA receptors in cerebellar granule neurons by ethanol: a review of genetic and electrophysiological studies. Alcohol 41:187–199
Bragina L, Bonifacino T, Bassi S, Milanese M, Bonanno G, Conit F (2015) Differential expression of metabotropic glutamate and GABA receptors at neocortical glutamatergic and GABAergic axon terminals. Front Cell Neurosci 9:345
Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA, Bonci A (2012) Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron 76:790–803
Cagetti E, Liang J, Spigelman I, Olsen RW (2003) Withdrawal from chronic intermittent ethanol treatment changes subunit composition, reduces synaptic function and decreases behavioral responses to positive allosteric modulators of GABAA receptors. Mol Pharmacol 63:53–64
Calabresi P, Gubellini P, Centonze D, Picconi B, Bernardi G, Chergui K, Svenningsson P, Fienberg AA, Greengard P (2000) Dopamine and cAMP-regulated phosphoprotein 32kDa controls both striatal long-term depression and long-term potentiation, opposing forms of synaptic plasticity. J Neurosci 20:8443–8451
Calton JL, Wilson WA, Moore SD (1998) Magnesium-dependent inhibition of N-methyl-D-aspartate receptor-mediated synaptic transmission by ethanol. J Pharmacol Exp Ther 287:1015–1019
Caputo F, Addolorato G, Stoppo M, Francini S, Vignoli T, Lorenzini F, Del Re A, Comaschi C, Andreone P, Trevisani F, Bernardi M, Alcohol Treatment Study Group (2007) Comparing and combining gamma-hydroxybutyric acid (GHB) and naltrexone in maintaining abstinence from alcohol: an open randomized comparative study. Eur Neuropsychopharmacol 17:781–789
Carlezon WA Jr, Devine DP, Wise RA (1995) Habit-forming actions of nomifensine in nucleus accumbens. Psychopharmacology (Berl) 122:194–197
Carpenter-Hyland EP, Chandler LJ (2006) Homeostatic plasticity during alcohol exposure promotes enlargement of dendritic spines. Eur J Neurosci 24:3496–3506
Carpenter-Hyland EP, Woodward JJ, Chandler LJ (2004) Chronic ethanol induces synaptic but not extrasynaptic targeting of NMDA receptors. J Neurosci 24:7859–7868
Carta M, Mameli M, Valenzuela CF (2004) Alcohol enhances GABAergic transmission to cerebellar granule cells via an increase in Golgi cell excitability. J Neurosci 24:3746–3751
Carvalho Poyraz F, Holzner E, Bailey MR, Meszaros J, Kenney L, Kheirbek MA, Balsam PD, Kellendonk C (2016) Decreasing striatopallidal pathway function enhances motivation by energizing the initiation of goal-directed action. J Neurosci 36:5988–6001
Chandler LJ, Norwood D, Sutton G (1999) Chronic ethanol upregulates NMDA and AMPA, but not kinate receptor subunit proteins in rat primary cortical cultures. Alcohol Clin Exp Res 23:363–370
Chen X, Moore-Nichols D, Nguyen H, Michaelis EK (1999) Calcium influx through NMDA receptors, chronic receptor inhibition by ethanol and 2-amino-5-phosponopentanoic acid, and receptor protein expression. J Neurochem 72:1969–1980
Cheng Y, Huang CCY, Ma T, Wei X, Wang X, Lu J, Wang J (2017) Distinct synaptic strengthening of the striatal direct and indirect pathways drives alcohol consumption. Biol Psychiatry 81:918–929
Chu B, Anantharam V, Treistman SN (1995) Ethanol inhibition of recombinant heteromeric NMDA channels in the presence and absence of modulators. J Neurochem 65:140–148
Clarke R, Adermark L (2015) Dopaminergic regulation of striatal interneurons in reward and addiction: focus on alcohol. Neural Plast 2015:814567
Corbit LH, Nie H, Janak PH (2012) Habitual alcohol seeking: time course and the contribution of subregions of the dorsal striatum. Biol Psychiatry 72:389–395
Corbit LH, Leung BK, Balleine BW (2013) The role of the amygdala-striatal pathway in the acquisition and performance of goal-directed instrumental actions. J Neurosci 33:17682–17690
Corbit LH, Nie H, Janak PH (2014) Habitual alcohol seeking: time course and the contribution of subregions of the dorsal striatum. Biol Psychiatry 72:389–395
Corti C, Aldegheri L, Somogyi P, Ferraguti F (2002) Distribution and synaptic localization of the metabotropic glutamate receptor 4 (mGluR4) in the rodent CNS. Neuroscience 110:403–420
Costa ET, Soto EE, Cardoso RA, Olivera DS, Valenzuela CF (2000) Acute effects of ethanol on kainite receptors in cultured hippocampal neurons. Alcohol Clin Exp Res 24:220–225
Cowan RL, Wilson CJ, Emson PC, Heizmann CW (1990) Parvalbumin-containing GABAergic interneurons in the rat neostriatum. J Comp Neurol 302:197–205
Cozzoli DK, Courson J, Caruana AL, Miller BW, Greentree DI, Thompson AB, Wroten MG, Zhang PW, Xiao B, Hu JH, Klugmann M, Metten P, Worley PF, Crabbe JC, Szumlinski KK (2012) Nucleus accumbens mGluR5-associated signaling regulates binge alcohol drinking under drinking-in-the dark procedures. Alcohol Clin Exp Res 36:1623–1633
Criswell HE, Ming Z, Kelm MK, Breese GR (2008) Brain regional differences in the effect of ethanol on GABA release from presynaptic terminals. J Pharmacol Exp Ther 326:596–603
Crowder TL, Ariwodola OJ, Weiner JL (2002) Ethanol antagonizes kainite receptor-mediated inhibition of evoked GABA (A) inhibitor postsynaptic currents in the rat hippocampal CA1 region. J Pharmacol Exp Ther 303:937–944
Cuzon Carlson VC, Seabold GK, Helms CM, Garg N, Odarigi M, Rau AR, Daunais J, Alvarez A, Lovinger DM, Grant KA (2011) Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates. Neuropsychopharmacology 36:2513–2528
Cuzon Carlson VC, Grant KA, Lovinger DM (2017) Synaptic adaptations to chronic ethanol intake in male rhesus monkey dorsal striatum depend on age of drinking onset. Neuropharmacology 131:128–142. https://doi.org/10.1016/j.neuropharm.2017.12.010
Dahchour A, Quertemont E, De Witte P (1994) Acute ethanol increases taurine but neither glutamate nor GABA in the nucleus accumbens of male rats: a microdialysis study. Alcohol Alcohol 29:485–487
Dahchour A, Quertemont E, De Witte P (1996) Taurine increases in the nucleus accumbens microdialysate after acute ethanol administration to naïve and chronically alcoholised rats. Brain Res 735:9–19
Dahchour A, De Witte P, Bolo N, Nédélec JF, Muzet M, Durbin P, Macher JP (1998) Central effects of acamprosate: part I. Acamprosate blocks the glutamate increase in the nucleus accumbens microdialysate in ethanol withdrawn rats. Psychiatry Res 82:107–114
Dalley JW, Cardinal RN, Robbins TW (2004) Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci Biobehav Rev 28:771–784
Das SC, Yamamoto BK, Hristov AM, Sari Y (2015) Ceftriaxone attenuates ethanol drinking and restores extracellular glutamate concentration through normalization of GLT-1 in nucleus accumbens of male alcohol-preferring rats. Neuropharmacology 97:67–74
DePoy L, Daut R, Brigman JL, MacPherson K, Crowley N, Gunduz-Cinar O, Pickens CL, Cinar R, Saksida LM, Kunos G, Lovinger DM, Bussey TJ, Camp MC, Holmes A (2013) Chronic alcohol produces neuroadaptations to prime dorsal striatal learning. Proc Natl Acad Sci U S A 110:14783–14788
Dickinson A, Wood N, Smith JW (2002) Alcohol seeking by rats: action or habit? Q J Exp Psychol B 55:331–348
Dildy-Mayfield JE, Harris RA (1992) Comparison of ethanol sensitivity of rat brain kainate, DL-alpha-amino-3-hydroxy-5-methyl-4-isoxalone proprionic acid and N-methyl-d-aspartate receptors expressed in Xenopus oocytes. J Pharmacol Exp Ther 262:487–494
Ding ZM, Engelman EA, Rodd ZA, McBride WJ (2012) Ethanol increases glutamate neurotransmission in the posterior ventral tegmental area of female Wistar rats. Alcohol Clin Exp Res 36:633–640
Ding ZM, Rodd ZA, Engelman EA, Bailey JA, Lahiri DK, McBride WJ (2013) Alcohol drinking and deprivation alter basal extracellular glutamate concentrations and clearance in the mesolimbic system of alcohol-preferring (P) rats. Addict Biol 18:297–306
Dingledine R, Borges K, Bowie D, Traynelis SF (1999) The glutamate receptor ion channels. Pharmacol Rev 51:7–62
Donoghue K, Elzerbi C, Saunders R, Whittington C, Pilling S, Drummond C (2015) The efficacy of acamprosate and naltrexone in the treatment of alcohol dependence, Europe versus the rest of the world: a meta-analysis. Addiction 110:920–930
Eisenhardt M, Leixner S, Lujan R, Spanagel R, Bilbao A (2015) Glutamate receptors within the mesolimbic dopamine system mediate alcohol relapse behavior. J Neurosci 35:15523–15538
Everitt BJ, Robbins TW (2013) From the ventral to the dorsal striatum: devolving views of their roles in drug addiction. Neurosci Biobehav Rev 37:1946–1954
Everitt BJ, Robbins TW (2016) Drug addiction: updating actions to habits to compulsions ten years on. Annu Rev Psychol 67:23–50
Everitt BJ, Belin D, Economidou D, Pelloux Y, Dalley JW, Robbins TW (2008) Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits and addiction. Philos Trans R Soc Lond B Biol Sci 363:3125–3135
Farrant M, Nusser Z (2005) Variations on an inhibitory theme: phasic and tonic activation of GABA(A) receptors. Nat Rev Neurosci 6:215–229
Ferrani-Kile K, Randall PK, Leslie SW (2003) Acute ethanol affects phosphorylation state of the NMDA receptor complex: implication of tyrosine phosphatases and protein kinase A. Mol Brain Res 115:78–86
Fliegel S, Brand I, Soanagel R, Noori HR (2013) Ethanol-induced alterations of amino acids measured by in vivo microdialysis in rats: a meta-analysis. In Silico Pharmacol 1:7
Floyd DW, Jung KY, McCool BA (2003) Chronic ethanol ingestion facilitates N-methyl-D-aspartate receptor function and expression in rat lateral/basolateral amygdala neurons. J Pharmacol Exp Ther 307:1020–1029
Floyd DW, Friedman DP, Daunais JB, Pierre PJ, Grant KA, McCool BA (2004) Long-term ethanol self-administration by cynomolgus macaques alters the pharmacology and expression of GABAA receptors in basolateral amygdala. J Pharmacol Exp Ther 311:1071–1079
Freund G, Anderson KJ (1996) Glutamate receptors in the frontal cortex of alcoholics. Alcohol Clin Exp Res 20:1165–1172
Freund G, Anderson KJ (1999) Glutamate receptors in the cingulate cortex, hippocampus, and cerebellar vermis of alcoholics. Alcohol Clin Exp Res 23:1–6
Fritschy JM, Panzanelli P, Tyagarajan SK (2012) Molecular and functional heterogeneity of GABAergic synapses. Cell Mol Life Sci 69:2485–2499
Fuchs RA, Branham RK, See RE (2006) Different neural substrates mediate cocaine seeking after abstinence versus extinction training: a critical role for the dorsolateral caudate-putamen. J Neurosci 26:3584–3588
Fujiyama F, Sohn J, Nakano T, Furata T, Nakamura KC, Matsuda W, Kaneko T (2011) Exclusive and common targets of neostriatofugal projections of rat striosome neurons: a single neuron-tracing study using a viral vector. Eur J Neurosci 33:668–677
Gage PW, Robertson B (1985) Prolongation of inhibitory postsynaptic currents by pentobarbitone, halothane and ketamine in CA1 pyramidal cells in rat hippocampus. Br J Pharmacol 85:675–681
Gallimberti L, Ferri M, Ferrara SD, Fadda F, Gessa GL (1992) Gamma-hydroxybutyric acid in the treatment of alcohol dependence: a double-blind study. Alcohol Clin Exp Res 16:673–676
Gerfen CR, Surmeier DJ (2011) Modulation of striatal projection systems by dopamine. Annu Rev Neurosci 34:441–466
Gibbs JW 3rd, Sombati S, DeLorenzo RJ, Coulter DA (2000) Cellular actions of topiramate: blockade of kainate-evoked inward currents in cultured hippocampal neurons. Epilepsia 41:S10–S16
Gittis AH, Hang GB, LaDow ES, Shoenfeld LR, Atallah BV, Finkbeiner S, Kreitzer AC (2011) Rapid target-specific remodeling of fast-spiking inhibitory circuits after loss of dopamine. Neuron 71:858–868
Gonda X (2012) Basic pharmacology of NMDA receptors. Curr Pharm Des 18:1558–1567
Gonzales KK, Smith Y (2015) Cholinergic interneurons in the dorsal and ventral striatum: anatomical and functional considerations in normal and diseased conditions. Ann N Y Acad Sci 1349:1–45
Goodwani S, Saternos H, Alasmari F, Sari Y (2017) Metabotropic and ionotropic glutamate receptors as potential targets for the treatment of alcohol use disorder. Neurosci Biobehav Rev 77:14–31
Groenewegen HJ, Wright CI, Beijer AV, Voorn P (1999) Convergence and segregation of ventral striatal inputs and outputs. Ann N Y Acad Sci 877:49–63
Grueter BA, Brasnjo G, Malenka RC (2010) Postsynaptic TRPV1 triggers cell type-specific long-term depression in the nucleus accumbens. Nat Neurosci 13:1519–1525
Gunaydin LA, Kreitzer AC (2016) Cortico-basal ganglia circuit function in psychiatric disease. Annu Rev Physiol 78:327–350
Guo Y, Wang HL, Xiang XH, Zhao Y (2009) The role of glutamate and its receptors in mesocorticolimbic dopaminergic regions in opioid addiction. Neurosci Biobehav Rev 33:864–873
Haber SN (2011) Neuroanatomy of reward: a view from the ventral striatum. In: Gottfried JA (ed) Neurobiology of sensation and reward. CRC Press and Taylor & Francis, Boca Raton, pp 235–262
Haber SN, Fudge JL, McFarland NR (2000) Striatonigrostriatal pathways in primates form an ascending spiral from the shell to the dorsolateral striatum. J Neurosci 20:2369–2382
Harris BR, Gibson DA, Prendergast MA, Blanchard JA, Holley RC, Hart SR, Scotland RL, Foster TC, Pedigo NW, Littleton JM (2003) The neurotoxicity induced by ethanol withdrawal in mature organotypic hippocampal slices might involve cross-talk between metabotropic glutamate type 5 receptors and N-methyl-D-aspartate receptors. Alcohol Clin Exp Res 27:1724–1735
Heimer L, Alheid GF (1991) Piecing together the puzzle of basal forebrain anatomy. Adv Exp Med Biol 295:1–42
Heimer L, Wilson RD (1975) The subcortical projections of the allocortex: similarities in the neural associations of the hippocampus, the piriform cortex, and the neocortex. In: Santini M (ed) Golgi centennial symposium proceedings. Raven, New York, pp 177–193
Heimer L, Zahm DS, Churchill L, Kalivas PW, Wohltmann C (1991) Specificity in the projection patterns of accumbal core and shell in rat. Neuroscience 41:89–125
Hemby SE, O’Connor JA, Acosta G, Floyd D, Anderson N, McCool BA, Friedman D, Grant KA (2006) Ethanol-induced regulation of GABA-A subunit mRNAs in prefrontal fields of cynomolgus monkeys. Alcohol Clin Exp Res 30:1978–1985
Herd MB, Brown AR, Lambert JJ, Belelli D (2013) Extrasynaptic GABA(A) receptors couple presynaptic activity to postsynaptic inhibition in the somatosensory thalamus. J Neurosci 33:14850–14868
Hikida T, Yawata S, Yamaguchi T, Sanjo T, Sasaoka T, Wang Y, Nakanishi S (2013) Pathway-specific modulation of nucleus accumbens in reward and aversive behavior via selective transmitter receptors. Proc Natl Acad Sci 110:342–347
Hodge CW, Cox AA (1998) The discriminative stimulus effects of ethanol are mediated by NMDA and GABA(A) receptors in specific limbic brain regions. Psychopharmacology (Berl) 139:95–107
Hogarth L, Balleine BW, Corbit LH, Killcross S (2013) Associative learning mechanisms underpinning the transition from recreational drug use to addiction. Ann N Y Acad Sci 1282:12–24
Holmes A, Spanagel R, Krystal JH (2013) Glutamatergic targets for new alcohol medications. Psychopharmacology (Berl) 229:539–554
Hopf FW (2017) Do specific NMDA receptors subunits act as gateways for addictive behaviors? Genes Brain Behav 16:118–138
Hopkins DA, Holstege G (1978) Amygdaloid projections to the mesencephalon, pons and medulla oblongata in the cat. Exp Brain Res 32:529–547
Ikemoto S (2007) Dopamine reward circuitry: two projection systems from the ventral midbrain to the nucleus accumbens – olfactory tubercle complex. Brain Res Rev 56:27–78
Imperato A, DiChiara G (1986) Preferential stimulation of dopamine release in the nucleus accumbens of freely moving rats by ethanol. J Pharmacol Exp Ther 239:219–228
Iorio KR, Reinlib L, Tabakoff B, Hoffman PL (1992) Chronic exposure of cerebellar granule cells to ethanol results in increased N-methyl-D-asparate receptor function. Mol Pharmacol 41:1142–1148
Ito R, Dalley JW, Robbins TW, Everitt BJ (2002) Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J Neurosci 22:6247–6253
Jeanes ZM, Buske TR, Morrisett RA (2011) In vivo chronic intermittent ethanol exposure reverses the polarity of synaptic plasticity in the nucleus accumbens shell. J Pharmacol Exp Ther 336:155–164
Jeanes ZM, Buske TR, Morrisett RA (2014) Cell type-specific encoding of ethanol exposure in the nucleus accumbens shell. Neuroscience 277:184–195
Jin Z, Bhandage AK, Bazov I, Kononenko O, Bakalkin G, Korpi ER, Birnir B (2014) Expression of specific ionotropic glutamate and GABA-A receptor subunits is decreased in central amygdala of alcoholics. Front Cell Neurosci 8:288
Johnson BA, Rosenthal N, Capece JA, Wiegand F, Mao L, Beyers K, Ait-Daoud N, Anton RF, Ciraulo D, Kranzler HR, Mann K, O’Malley SS, Swift RM (2007) Topiramate for treating alcohol dependence: a randomized controlled trial. JAMA 298:1641–1651
Jonas DE, Amick HR, Feltner C, Bobashev G, Thomas K, Wilnes R, Kim MM, Shanahan E, Gass CE, Rowe CJ, Garbutt JC (2014) Pharmacotherapy for adults with alcohol use disorders in outpatient settings: a systemic review and meta-analysis. JAMA 311:1889–1900
Kalev-Zylinska ML, During MJ (2007) Paradoxical faciliatory effect of low-dose alcohol consumption on memory mediated by NMDA receptors. J Neurosci 27:10456–10467
Kalluri HS, Mehta AK, Ticky MK (1998) Up-regulation of NMDA receptor subunits in rat brain following chronic ethanol treatment. Brain Res Mol Brain Res 58:221–224
Kamal RM, van Noorden MS, Franzek E, Dijkstra BA, Loonen AJ, De Jong CA (2016) The neurobiological mechanisms of gamma-hydroxybutyrate dependence and withdrawal and their clinical relevance: a review. Neuropsychobiology 73:65–80
Kasai H, Fukuda M, Watanabe S, Hayashi-Takagi A, Noguchi J (2010) Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci 33:121–129
Kash TL, Baucum AJ 2nd, Conrad KL, Colbran RJ, Winder DG (2009) Alcohol exposure alters NMDAR function in the bed nucleus of the stria terminalis. Neuropsychopharmacology 34:2420–2429
Kawaguchi Y, Wilson CJ, Augood SJ, Emson PC (1995) Striatal interneurons: chemical, physiological and morphological characterization. Trends Neurosci 18:527–535
Kelm MK, Criswell HE, Breese GR (2011) Ethanol-enhanced GABA release: a focus on G-protein coupled receptors. Brain Res Rev 65:113–123
Kemp JM, Powell TP (1971) The structure of the caudate nucleus of the cat: light and electron microscopy. Phil Trans R Soc London 262:383–401
Kenny PJ, Markou A (2004) The ups and downs of addiction: role of metabotropic glutamate receptors. Trends Pharmacol Sci 25:265–272
Kerr JN, Wickens JR (2001) Dopamine D-1/D-5 receptor activation is required for long-term potentiation in the rat neostriatum in vitro. J Neurophysiol 85:117–124
Kita H (1993) GABAergic circuits of the striatum. Prog Brain Res 99:51–72
Kita H, Kitai ST (1988) Glutamate decarboxylase immunoreactive neurons in rat neostriatum: their morphological types and populations. Brain Res 447:346–352
Kittler JT, Chen G, Honing S, Bogdanov T, McAinsh K, Arancibia-Carcamo IL, Jovanovic JN, Pangalos MN, Haucke V, Yan Z, Moss SJ (2005) Phospho-dependent binding of the clathrin AP2 adaptor complex to GABAA receptors regulates the efficacy of inhibitory synaptic transmission. Proc Natl Acad Sci U S A 102:14871–14876
Koob GF (2013) Negative reinforcement in drug addiction: the darkness within. Curr Opin Neurobiol 23:559–563
Koob GF, Volkow ND (2010) Neurocircuitry of addiction. Neuropsychopharmacology 35:217–238
Koós T, Tepper JM (1999) Inhibitory control of neostriatal projection neurons by GABAergic interneurons. Nat Neurosci 2:467–472
Kranzler HR, Wetherill R, Feinn R, Pond T, Gelernter J, Covault J (2014) Posttreatment effects of topiramate treatment for heavy drinking. Alcohol Clin Exp Res 38:3017–3023
Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC (2010) Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 466:622–626
Kravitz AV, Tye LD, Kreitzer AC (2012) Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat Neurosci 15:816–818
Kreitzer AC, Malenka RC (2007) Endocannabinoid-mediated rescue of striatal LTD and motor deficits in Parkinson’s disease models. Nature 445:643–647
Kreitzer AC, Malenka RC (2008) Striatal plasticity and basal ganglia circuit function. Neuron 60:543–554
Krupitsky EM, Neznanova I, Masalov D, Burakov AM, Didenko T, Romanova T, Tsoy M, Bespalov A, Slavina TY, Grinenko AA, Petrakis IL, Pittman B, Gueorguieva R, Zvartau EE, Krystal JH (2007) Effect of memantine on cue-induced alcohol craving in recovering alcohol-dependent patients. Am J Psychiatry 164:519–523
Krystal JH, Petrakis IL, Krupitsky E, Schutz C, Trevisan L, D’Souza DC (2003) NMDA receptor antagonism and the ethanol intoxication signal: from alcoholism risk to pharmacotherapy. Ann N Y Acad Sci 1003:176–184
Kubota Y, Kawaguchi Y (2000) Dependence of GABAergic synaptic areas on the interneuron type and target size. J Neurosci 20:467–472
Kumari M, Ticku MK (2000) Regulation of NMDA receptors by ethanol. Prog Drug Res 54:151–189
Kuner T, Schoepfer R, Korpi ER (1993) Ethanol inhibits glutamate-induced currents in heteromeric NMDA receptor subtypes. Neuroreport 5:297–300
Kupchik YM, Brown RM, Heinsbroek JA, Lobo MK, Schwartz DJ, Kalivas PW (2015) Coding the direct/indirect pathways by D1 and D2 receptors is not valid for accumbens projections. Nat Neurosci 18:1230–1232
Kupferschmidt DA, Cody PA, Lovinger DM, Davis MI (2015) Brain BLAQ: posthoc thick-section histochemistry for localizing optogenetic consturcts in neurons and their distal terminals. Front Neuroanat 9:6
Läck AK, Diaz MR, Chappell A, DuBois DW, McCool BA (2007) Chronic ethanol and withdrawal differentially modulate pre-and postsynaptic function at glutamatergic synapses in rat basolateral amygdala. J Neurophysiol 98:3185–3196
Lambot L, Chaves Rodriguez E, Houtteman D, Li Y, Schiffmann SN, Gall D, de Kerchove d’Exaerde A (2016) Striatopallidal neuron NMDA receptors control synaptic connectivity, locomotor, and goal-directed behaviors. J Neurosci 36:4976–4992
Le Moine C, Bloch B (1995) D1 and D2 dopamine receptor gene expression in the rat striatum: sensitive cRNA probes demonstrate prominent segregation of D1 and D2 mRNAs in distinct neuronal populations of the dorsal and ventral striatum. J Comp Neurol 355:418–426
Li HF, Kendig JJ (2003) Ethanol withdrawal hyper-responsiveness mediated by NMDA receptors in spinal cord motor neurons. Br J Pharmacol 139:73–80
Li Q, Wilson WA, Swartzwelder HS (2006) Developmental differences in the sensitivity of spontaneous and miniature IPSCs to ethanol. Alcohol Clin Exp Res 30:119–126
Liang J, Cagetti E, Olsen RW, Spigelman I (2004) Altered pharmacology of synaptic and extrasynaptic GABAA receptors on CA1 hippocampal neurons is consistent with subunit changes in a model of alcohol withdrawal and dependence. J Pharmacol Exp Ther 310:1234–1245
Liang J, Lindemeyer AK, Suryanarayanan A, Meyer EM, Marty VN, Ahmad SO, Shao XM, Olsen RW, Spigelman I (2014) Plasticity of GABA(A) receptor-mediated neurotransmission in the nucleus accumbens of alcohol-dependent rats. J Neurophysiol 112:39–50
Lima-Landman MT, Albuquerque EX (1989) Ethanol potentiates and blocks NMDA-activated single-channel currents in rat hippocampal pyramidal cells. FEBS Lett 247:61–67
Lingford-Hughes A, Reid AG, Myers J, Feeney A, Hammers A, Taylor LG, Rosso L, Turkheimer F, Brooks DJ, Grasby P, Nutt DJ (2012) A [11C] Ro154513 PET study suggests that alcohol dependence in man is associated with reduced alpha 5 benzodiazepine receptors in limbic regions. J Psychopharmacol 26:273–281
Loebrich S, Bahring R, Katsuno T, Tsukita S, Kneussel M (2006) Activated radixin is essential for GABAA receptor alpha5 subunit anchoring at the actin cytoskeleton. EMBO J 25:987–999
Lominac KD, Kapasova Z, Hannun RA, Patterson C, Middaugh LD, Szumlinksi KK (2006) Behavioral and neurochemical interactions between group I mGluR antagonists and ethanol: potential insight into their anti-addictive properties. Drug Alcohol Depend 85:142–156
Lopez MF, Becker HC, Chandler LJ (2014) Repeated episodes of chronic intermittent ethanol promote insensitivity to devaluation of the reinforcing effect of ethanol. Alcohol 48:639–645
Lovinger DM (2017) Presynaptic ethanol actions: potential roles in ethanol seeking. Handb Exp Pharmacol. https://doi.org/10.1007/164_2017_76
Lovinger DM, White G, Weight FF (1989) Ethanol inhibits NMDA-activated ion current in hippocampal neurons. Science 243:1721–1724
Lovinger DM, White G, Weight FF (1990) NMDA receptor-mediated synaptic excitation selectively inhibited by ethanol in hippocampal slice from adult rat. J Neurosci 10:1372–1379
Lu XY, Ghasemzadeh MB, Kalivas PW (1998) Expression of D1 receptor, D2 receptor, substance P, and enkephalin messenger RNAs in the neurons projecting from the nucleus accumbens. Neuroscience 82:767–780
Luo Z, Volkow ND, Heintz N, Pan Y, Du C (2011) Acute cocaine induces fast activation of D1 receptor and progressive deactivation of D2 receptor striatal neurons: in vivo optical microprobe [Ca2+] imaging. J Neurosci 31:13180–13190
Ma T, Barbee B, Wang X, Wang J (2017) Alcohol induces input-specific aberrant synaptic plasticity in the rat dorsomedial striatum. Neuropharmacology 123:46–54
Maguire EP, Macpherson T, Swinny JD, Dixon CI, Herd MB, Belelli D, Stephens DN, King SL, Lambert JJ (2014) Tonic inhibition of accumbal spiny neurons by extrasynaptic alpha4betadelta GABAA receptors modulates the actions of psychostimulants. J Neurosci 34:823–838
Maisel NC, Blodgett JC, Wilbourne PL, Humphreys K, Finney JW (2013) Meta-analysis of naltrexone and acamprosate for treating alcohol use disorders: when are these medications most helpful? Addiction 108:275–293
Maldve RE, Zhang TA, Ferrani-Kile K, Schreiber SS, Lippmann MJ, Snyder GL, Fienberg AA, Leslie SW, Gonzales RA, Morrisett RA (2002) DARPP-32 and regulation of the ethanol sensitivity of NMDA receptors in the nucleus accumbens. Nat Neurosci 5:641–648
Malenka RC, Nicoll RA (1999) Long-term potentiation – a decade of progress? Science 285:1870–1874
Malinow R, Malenka RC (2002) AMPA receptor trafficking and synaptic plasticity. Annu Rev Neurosci 25:103–126
Mallet N, LeMoine C, Charpier S, Ganon F (2005) Feedforward inhibition of projection neurons by fast-spiking GABA interneurons in the rat striatum in vivo. J Neurosci 25:3857–3869
Martin D, Swartzwelder HS (1992) Ethanol inhibits release of excitatory amino acids from slices of hippocampal area CA1. Eur J Pharmacol 219:469–472
Martin D, Morrisett RA, Bian XP, Wilson WA, Swartzwelder HS (1991) Ethanol inhibition of NMDA mediated depolarizations is increased in the presence of Mg2+. Brain Res 546:227–234
Martinotti G, Di Nicola M, De Vita O, Hatzigakoumis DS, Guglielmo R, Santucci B, Aliotta F, Romanelli R, Verrastro V, Petruccelli F, Di Giannantonio M, Janiri L (2014) Low-dose topiramate in alcohol dependence: a single-blind, placebo-controlled study. J Clin Psychopharmacol 34:709–715
Marty VN, Spigelman I (2012) Long-lasting alterations in membrane properties K(+) currents, and glutamatergic synaptic currents of nucleus accumbens medium spiny neurons in a rat model of alcohol dependence. Front Neurosci 6:86
Mason BJ, Quello S, Goodell V, Shadan F, Kyle M, Begovic A (2014) Gabapentin treatment for alcohol dependence: a randomized clinical trial. JAMA Intern Med 174:70–77
Masood K, Wu C, Brauneis U, Weight FF (1994) Differential ethanol sensitivity of recombinant N-methyl-D-aspartate receptor subunits. Mol Pharmacol 45:324–329
Matsuzaki M, Honkura N, Ellis-Davies GC, Kasai H (2004) Structural basis of long-term potentiation in single dendirit spines. Nature 429:761–766
McGeorge AJ, Faull RLM (1989) The organization of the projection from the cerebral cortex to the striatum in the rat. Neuroscience 29:503–537
Meinhardt MW, Hansson AC, Perreau-Lentz S, Bauder-Wenz C, Stahlin O, Heilig M, Harper C, Drescher KU, Spanagel R, Sommer WH (2013) Rescue of infralimbic mGluR2 deficit restores control over drug-seeking behavior in alcohol dependence. J Neurosci 33:2794–2806
Mele M, Ribeiro L, Inacio AR, Wieloch T, Duarte CB (2014) GABA(A) receptor dephosphorylation followed by internalization is coupled to neuronal death in in vitro ischemia. Neurobiol Dis 65:220–232
Melendez RI, Hicks MP, Cagle SS, Kalivas PW (2005) Ethanol exposure decreases glutamate uptake in the nucleus accumbens. Alcohol Clin Exp Res 29:326–333
Melis M, Camarini R, Ungless MA, Bonci A (2002) Long-lasting potentiation of GABAergic synapses in dopamine neurons after a single in vivo ethanol exposure. J Neurosci 22:2074–2082
Meredith GE (1999) The synaptic framework for chemical signaling in nucleus accumbens. Ann N Y Acad Sci 877:140–156
Messenger MJ, Dawson LG, Duty S (2002) Changes in metabotropic glutamate messenger 1-8 gene expression in the rodent basal ganglia motor loop following lesion of the nigrostriatal tract. Neuropharmacology 43:261–271
Michaelis EK, Michaelis ML, Freed WJ, Foye J (1993) Glutamate receptor changes in brain synaptic membrane during chronic alcohol intake. Alcohol Alcohol Suppl 2:377–381
Ming Z, Criswell HE, Yu G, Breese GR (2006) Competing presynaptic and postsynaptic effects of ethanol on cerebellar purkinje neurons. Alcohol Clin Exp Res 30:1400–1407
Mirshahi T, Woodward JJ (1995) Ethanol sensitivity of heteromeric NMDA receptors: effects of subunit assembly, glycine and NMDAR1 Mg(2+)-insensitive mutants. Neuropharmacology 34:347–355
Misgeld U, Drew G, Yanovsky Y (2007) Presynaptic modulation of GABA release in the basal ganglia. Prog Brain Res 160:245–259
Miyakawa T, Yagi T, Kitazawa H, Yasuda M, Kawai N, Tsuboi K, Niki H (1997) Fyn-kinase as a determinant of ethanol sensitivity: relation to NMDA-receptor function. Science 278:698–701
Moghaddam B, Bolinao ML (1994) Biphasic effect of ethanol on extracellular accumulation of glutamate in the hippocampus and the nucleus accumbens. Neurosci Lett 178:99–102
Moon IS, Apperson ML, Kennedy MB (1994) The major tyrosine-phosphorylated protein in the postsynaptic density fraction is N-methyl-D-aspartate receptor subunit 2B. Proc Natl Acad Sci U S A 91:3954–3958
Morrisett RA, Martin D, Oetting TA, Lewis DV, Wilson WA, Swartzwelder HS (1991) Ethanol and magnesium ions inhibit N-methyl-D-aspartate-mediated synaptic potentials in an interactive manner. Neuropharmacology 30:1173–1178
Moykkynen T, Korpi ER, Lovinger DM (2003) Ethanol inhibits alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionc acid (AMPA) receptor function in central nervous system neurons by stabilizing desensitization. J Pharmacol Exp Ther 306:546–555
Muller J, Dreisbach G, Goschke T, Hensch T, Lesch KP, Brocke B (2007) Dopamine and cognitive control: the prospect of monetary gains influences the balance between flexibility and stability in a set-shifting paradigm. Eur J Neurosci 26:3661–3668
Muller CA, Geisel O, Pelz P, Higl V, Kruger J, Stickel A, Beck A, Wernecke KD, Hellweg R, Heinz A (2015) High-dose baclofen for the treatment of alcohol dependence (BACLAD study): a randomized, placebo-controlled trial. Eur Neuropsychopharmacol 25:1167–1177
Nagy J (2008) Alcohol related changes in regulation of NMDA receptor functions. Curr Neuropharmacol 6:39–54
Nagy J, Kolok S, Dezso P, Boros A, Szobathelyi Z (2003) Differential alterations in the expression of NMDA receptor subunits following chronic ethanol treatment in primary cultures of rat cortical and hippocampal neurons. Neurochem Int 42:35–43
Neasta J, Ben Hamida S, Yowell Q, Carnicells S, Ron D (2010) Role for mammalian target of rapamycin complex 1 signaling in neuroadaptations underlying alcohol-related disorders. Proc Natl Acad Sci U S A 107:20093–20098
Nelson TE, Ur CL, Gruol DL (2005) Chronic intermittent ethanol exposure enhances NMDA-receptor-mediated synaptic responses and NMDA receptor expression in hippocampal CA1 region. Brain Res 1048:69–79
Nestoros JN (1980) Ethanol specifically potentiates GABA-mediated neurotransmission in feline cerebral cortex. Science 209:708–710
Nie Z, Madamba S, Siggins G (1997) Ethanol enhances GABAergic transmission in nucleus accumbens: regulation by metabotropic mechanisms. Alcohol Clin Exp Res 21:74A
Nie Z, Madamba S, Siggins G (2000) Ethanol enhances γ-aminobutyric acid responses in a subpopulation of nucleus accumbens neurons; role of metabotropic glutamate receptors. J Pharmacol Exp Ther 293:654–661
Nie Z, Schweitzer P, Roberts AJ, Madamba SG, Moore SD, Siggins GR (2004) Ethanol augments GABAergic transmission in the central amygdala via CRF1 receptors. Science 303:1512–1514
Obara I, Bell RL, Goulding SP, Reyes CM, Larson LA, Ary AW, Truitt WA, Szumlinksi KK (2009) Differential effects of chronic ethanol consumption and withdrawal on homer/glutamate receptor expression in subregions of the accumbens and amygdala of P rats. Alcohol Clin Exp Res 33:1924–1934
Olive MF (2009) Metabotropic glutamate receptor ligands as potential therapeutics for addiction. Curr Drug Abuse Rev 2:83–98
Olsen RW, Sieghart W (2008) International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classification on the bass of subunit composition, pharmacology, and function. Update. Pharmacol Rev 60:243–260
Olsen RW, Sieghart W (2009) GABA A receptors: subtypes provide diversity of function and pharmacology. Neuropharmacology 56:141–148
Olsen RW, Hanchar HJ, Meera P, Wallner M (2007) GABAA receptor subtypes: the “one glass of wine” receptors. Alcohol 41:201–209
Othman T, Singlair CJ, Haughey N, Geiger JD, Parkinson FE (2002) Ethanol alters glutamate but not adenosine uptake in rat astrocytes: evidence for protein kinase C involvement. Neurochem Res 27:289–296
Pan WX, Mao T, Dudman JT (2010) Inputs to the dorsal striatum of the mouse of the mouse reflect the parallel circuit architecture of the forebrain. Front Neuroanat 4:147
Papadeas S, Grobin AC, Morrow AL (2001) Chronic ethanol consumption differentially alters GABA(A) receptor alpha 1 and alpha 4 subunit peptide expression and GABA(A) receptor-mediated 36 Cl(-) uptake in mesocorticolimbic regions of the rat brain. Alcohol Clin Exp Res 25:1270–1275
Papp E, Borhegyi Z, Tomioka R, Rockland KS, Mody I, Freund TF (2012) Glutamatergic input from specific sources influences the nucleus accumbens-ventral pallidum information flow. Brain Struct Funct 217:37–48
Patton MH, Roberts DM, Lovinger DM, Mathur BN (2016) Ethanol disinhibits dorsolateral striatal medium spiny neurons through activation of a presynaptic delta opioid receptor. Neuropsychopharmacology 41:1831–1840
Peris J, Eppler B, Hu M, Walker DW, Hunter BE, Mason K (1997) Effects of chronic ethanol exposure on GABA receptors and GABAB receptor modulation of 3H-GABA release in the hippocampus. Alcohol Clin Exp Res 21:1047–14052
Piepponen TP, Kiianmaa K, Ahtee L (2002) Effects of ethanol on the accumbal output of dopamine, GABA and glutamate in alcohol-tolerant and alcohol-nontolerant rats. Pharmacol Biochem Behav 74:21–30
Planert H, Berger TK, Silberberg G (2013) Membrane properties of striatal direct and indirect pathway neurons in mice and rat slices and their modulation by dopamine. PLoS One 8:e57054
Pomierny-Chamiolo L, Rup K, Pomierny B, Niedzielska E, Kalivas PW, Filip M (2014) Metabotropic glutamatergic receptors and their ligands in drug addiction. Pharmacol Ther 142:281–305
Popp RL, Lickteig R, Browning MD, Lovinger DM (1998) Ethanol sensitivity and subunit composition of NMDA receptors in cultured striatal neurons. Neuropharmacology 37:45–56
Proctor WR, DIao L, Freund RK, Browning MD, Wu PH (2006) Synaptic GABAergic and glutamatergic mechanisms underlying alcohol sensitivity in mouse hippocampal neurons. J Physiol 575:145–149
Qi ZH, Song M, Wallace MJ, Wang D, Newton PM, McMahon T, Chou WH, Zhang C, Shokat KM, Messing RO (2007) Protein kinase C epsilon regulates gamma-aminobutyrate type A receptors sensitivity to ethanol and benzodiazepines through phosphorylation of gamma2 subunits. J Biol Chem 282:33052–33063
Quertemont E, Linotte S, De Witte P (2002) Differential taurine responsiveness to ethanol in high- and low-alcohol sensitive rats: a brain microdialysis study. Eur J Pharmacol 444:143–150
Rabe CS, Tabakoff B (1990) Glycine site-directed agonists reverse the actions of ethanol at the N-methyl-D-aspartate receptor. Mol Pharmacol 38:753–757
Renteria R, Maier EY, Buske TR, Morrisett RA (2017) Selective alterations of NMDAR function and plasticity in D1 and D2 medium spiny neurons in the nucleus accumbens shell following chronic intermittent ethanol exposure. Neuropharmacology 112:164–171
Reynaud M, Aubin HJ, Trinquet F, Zakine B, Dano C, Dematteis M, Trojak B, Paille F, Detilleux M (2017) A randomized, placebo-controlled study of high-dose baclofen in alcohol-dependent patients – the ALPADIR study. Alcohol Alcohol 52:439–446
Reynolds JNJ, Wickens JR (2002) Dopamine-dependent plasticity of corticostriatal synapses. Neural Netw 15:507–521
Roberto M, Madamba SG, Moore SD, Tallent MK, Siggins GR (2003) Ethanol increases GABAergic transmission at both pre- and postsynaptic sites in rat central amygdala neurons. Proc Natl Acad Sci U S A 100:2053–2058
Roberto M, Madamba SG, Stouffer DG, Parsons LH, Siggins GR (2004) Increased GABA release in the central amygdala of ethanol-dependent rats. J Neurosci 24:10159–10166
Roberto M, Gilpin NW, O’Dell LE, Cruz MT, Morse AC, Siggins GR, Koob GF (2008) Cellular and behavioral interactions of gabapentin with alcohol dependence. J Neurosci 28:5762–5771
Roberto M, Cruz MT, Gilpin NW, Sabino V, Schweitzer P, Bajo M, Cottone P, Madamba SG, Stouffer DG, Zorilla EP, Koob GF, Siggins GR, Parsons LH (2010) Corticotropin releasing factor-induced amygdala gamma-aminobutyric acid release plays a key role in alcohol dependence. Biol Psychiatry 67:831–839
Rodd-Henricks ZA, McKinzie DL, Li TK, Murphey JM, McBride WJ (2002) Cocaine is self-administered into the shell but not the core of the nucleus accumbens of Wistar rats. J Pharmacol Exp Ther 303:1216–1226
Rossetti ZL, Carboni S (1995) Ethanol withdrawal is associated with increased extracellular glutamate in the rat striatum. Eur J Pharmacol 283:177–183
Rossetti Z, Carboni S, Fadda F (1999) Glutamate-induced increase of extracellular glutamate through N-methyl-D-aspartate receptors in ethanol withdrawal. Neuroscience 93:1135–1140
Sanna E, Serra M, Cossu A, Colombo G, Follesa P, Cuccheddu T, Concas A, Biggio G (1993) Chronic ethanol intoxication induces differential effects on GABAA and NMDA receptor function in the rat brain. Alcohol Clin Exp Res 17:115–123
Sanna E, Talani G, Busonero F, Pisu MG, Purdy RH, Serra M, Biggio G (2004) Brain steroidogenesis mediates ethanol modulation of GABAA receptor activity in rat hippocampus. J Neurosci 24:6521–6530
Santerre JL, Rogow JA, Kolitz EB, Pal R, Landin JD, Gigante E, Werner DF (2014) Ethanol dose-dependently elicits opposing regulatory effects on hippocampal AMPA receptor GluA2 subunits through a zeta inhibitory peptide-sensitive kinase in adolescent and adult Sprague-Dawley rats. Neuroscience 280:50–59
Schumann G, Johann M, Frank J, Preuss U, Dahmen N, Laucht M, Rieschel M, Rujescu D, Lourdusamy A, Clarke TK, Krause K, Dyer A, Depner M, Wellek S, Treutlein J, Szegdi A, Giegling I, Cichon S, Blomeyer D, Heinz A, Heath S, Lathrop M, Wodarz N, Soyka M, Spanagel R, Mann K (2008) Systematic analysis of glutamatergic neurotransmission genes in alcohol dependence and adolescent risky drinking behavior. Arch Gen Psychiatry 65:826–838
Schwarzer C, Berresheim U, Pirker S, Wieselthaler A, Fuchs K, Sieghart W, Sperk G (2001) Distribution for the major gamma-aminobutyric acid (A) receptor subunits in the basal ganglia and associated limbic brain areas of the adult rat. J Comp Neurol 433:526–549
Selim M, Bradberry CW (1996) Effect of ethanol on extracellular 5-HT and glutamate in the nucleus accumbens and prefrontal cortex: comparison between the Lewis and Fischer 344 rat strains. Brain Res 716:157–164
Sellings LH, Clarke PB (2003) Segregation of amphetamine reward and locomotor stimulation between nucleus accumbens medial shell and core. J Neurosci 23:6295–6303
Siggins GR, Pittman QJ, French ED (1987) Effects of ethanol on CA1 and CA3 pyramidal cells in the hippocampal slice preparation: an intracellular study. Brain Res 414:22–34
Siggins GR, Roberto M, Nie Z (2005) The tipsy terminal: presynaptic effects of ethanol. Pharmacol Ther 107:80–98
Silberman Y, Shi L, Brunso-Bechtold JK, Weiner JL (2008) Distinct mechanisms of ethanol potentiation of local and paracapsular GABAergic synapses in the rat basolateral amygdala. J Pharmacol Exp Ther 324:251–260
Silberman Y, Shi L, Ariwodola OJ, Weiner JL (2009) Differential effects of GABAB autoreceptor activation on ethanol potentiation of local and lateral paracapsular GABAergic synapses in the rat basolateral amygdala. Neuropharmacology 56:886–895
Sinclair JD, Senter RJ (1967) Increased preference for ethanol in rats following deprivation. Psychon Sci 8:11–12
Sinclair JD, Senter RJ (1968) Development of an alcohol-deprivation effect in rats. Q J Stud Alcohol 29:863–867
Sinclair CM, Cleva RM, Hood LE, Olive MF, Gass JT (2012) mGluR5 receptors in the basolateral amygdala and nucleus accumbens regulate cue-induced reinstatement of ethanol-seeking behavior. Pharmacol Biochem Behav 101:329–335
Smith TL (1997) Regulation of glutamate uptake in astrocytes continuously exposed to ethanol. Life Sci 61:2499–2505
Smith AD, Bolam JP (1990) The neural network of the basal ganglia as revealed by the study of synaptic connections of identified neurons. Trends Neurosci 13:259–265
Smith Y, Parent A (1986) Neuropeptide Y-immunoreactive neurons in the striatum of the cat and monkey: morphological characteristics, intrinsic organization and co-localization with somatostatin. Brain Res 372:241–252
Smothers CT, Woodward JJ (2003) Effect of the NR3 subunit on ethanol inhibition of recombinant NMDA receptors. Brain Res 987:117–121
Snell LD, Tabakoff B, Hoffman PL (1994) Involvement of protein kinase C in ethanol-induced inhibition of NMDA receptor function in cerebellar granule cells. Alcohol Clin Exp Res 18:81–85
Soyka M, Kranzler HR, Hesselbrock V, Kasper S, Muschler J, Moller HJ, WFSBP Task Force on Treatment Guidelines for Substance Use Disorders (2016) Guidelines for biological treatment of substance use and related disorders, part I: alcoholism, first revision. World J Biol Psychiatry 23:1–34
Surmeier DJ, Song WJ, Yan Z (1996) Coordinated expression of dopamine receptors in neostriatal medium spiny neurons. J Neurosci 16:6579–6591
Suto N, Ecke LE, You ZB, Wise RA (2010) Extracellular fluctuations of dopamine and glutamate in the nucleus accumbens core and shell associated with lever-pressing during cocaine self-administration, extinction, and yoked cocaine administration. Psychopharmacology 211:267–275
Suvarna N, Borgland SL, Wang J, Phamluong K, Aberson YP, Bonci A, Ron D (2005) Ethanol alters trafficking and functional N-methyl-D-aspartate receptor NR2 subunit ration via H-ras. J Biol Chem 280:31450–31459
Suzdak PD, Schwartz RD, Skolnick P, Paul SM (1986) Ethanol stimulates γ-aminobutyric acid receptor-mediated chloride transport in rat brain synaptoneurosomes. Proc Natl Acad Sci 83:4071–4075
Szumlinski KK, Diab ME, Friedman R, Henze LM, Lominac KD, Bowers MS (2007) Accumbens neurochemical adaptations produced by binge-like alcohol consumption. Psychopharmacology (Berl) 190:415–431
Taylor CP (1997) Mechanisms of action of gabapentin. Rev Neurol 153:S39–S45
Tepper JM, Bolam JP (2004) Functional diversity and specificity of neostriatal interneurons. Curr Opin Neurobiol 14:685–692
Theile JW, Morikawa H, Gonzales RA, Morrisett RA (2008) Ethanol enhances GABAergic transmission onto dopamine neurons in the ventral tegmental area of the rat. Alcohol Clin Exp Res 32:1040–1048
Tiwari V, Veeraiah P, Subramaniam V, Patel AB (2014) Differential effects of ethanol on regional glutamatergic and GABAergic neurotransmission pathways in mice brain. J Neurochem 128:628–640
Tonner PH, Miller KW (1995) Molecular sites of general anaesthetic action on acetylcholine receptors. Eur J Anaesthesiol 12:21–30
Traynelis SF, Wollmuth LP, McBain CJ, Menniti FS, Vance KM, Ogden KK, Hansen KB, Yuan H, Myers SJ, Dingledine R (2010) Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev 62:405–496
Tremwel MF, Hunter BE, Peris J (1994) Chronic ethanol exposure enhances [3H]GABA release and does not affect GABAA receptor mediated 36Cl uptake. Synapse 17:149–154
Trevisan L, Fitzgerald LW, Brose N, Gasic GP, Heinemann SF, Duman RS, Nestler EJ (1994) Chronic ingestion of ethanol upregulates NMDAR1 receptor subunit immunoreactivity in rat hippocampus. J Neurochem 62:1635–1638
Tsai G, Coyle JT (1998) The role of glutamatergic neurotransmission in the pathophysiology of alcoholism. Annu Rev Med 49:173–184
Tsai G, Gastfriend DR, Coyle JT (1995) The glutamatergic basis of human alcoholism. Am J Psychiatry 152:332–340
Tunstall MJ, Oorschot DE, Kean A, Wickens JR (2002) Inhibitory interactions between spiny projection neurons in the rat striatum. J Neurophysiol 88:1263–1269
Vengeliene V, Cilbao A, Molander A, Spanagel R (2008) Neuropharmacology of alcohol addiction. Br J Pharmacol 154:299–315
Voris J, Smith NL, Rao SM, Thorne DL, Flowers QJ (2003) Gabapentin for the treatment of ethanol withdrawal. Subst Abus 24:129–132
Walker MC, Semyanov A (2008) Regulation of excitability by extrasynaptic GABA(A) receptors. Results Probl Cell Differ 44:29–48
Wallner M, Hanchar HJ, Olsen RW (2006) Low dose acute alcohol effects on GABAA receptor subtypes. Pharmacol Ther 112:513–528
Wang XJ (1999) Synaptic basis of cortical persistent activity: the importance of NMDA receptors to working memory. J Neurosci 19:9587–9603
Wang J, Carnicella S, Phamluong K, Jeanblanc J, Ronesi JA, Chaudhru N, Janak PH, Lovinger DM, Ron D (2007) Ethanol induces long-term facilitation of NR2B-NMDA receptor activity in the dorsal striatum: implications for alcohol drinking behavior. J Neurosci 27:3593–3602
Wang J, Lanfranco MF, Gibb SL, Yowell QV, Carnicella S, Ron D (2010) Long-lasting adaptations of the NR2B-containing NMDA receptors in the dorsomedial striatum play a crucial role in alcohol consumption and relapse. J Neurosci 31:8163–8174
Wang J, Ben Hamida S, Darcq E, Zhu W, Gibb SL, Lanfranco MF, Carnicella S, Ron D (2012) Ethanol-mediated facilitation of AMPA receptor function in the dorsomedial striatum: implications for alcohol drinking behavior. J Neurosci 32:15124–15132
Wang J, Cheng Y, Wang X, Roltsch Hellard E, Ma T, Gil H, Ben Hamida S, Ron D (2015) Alcohol elicits functional and structural plasticity selectively in dopamine D1 receptor-expressing neurons of the dorsomedial striatum. J Neurosci 35:11634–11463
Ward RJ, Colivicchi MA, Allen R, Schol F, Lallemand F, de Witte P, Ballini C, Corte LD, Dexter D (2009) Neuro-inflammation induced in the hippocampus of ‘binge drinking’ rats may be mediated by elevated extracellular glutamate content. J Neurochem 111:1119–1128
Webster KE (1961) Corticostriate interrelations in the albino rat. J Anat 95:532–545
Weight FF, Peoples RW, Wright JM, Lovinger DM, White G (1993) Ethanol action on excitatory amino acid activated ion channels. Alcohol Alcohol Supp 2:353–358
Weiner JL, Zhang L, Carlen PL (1994) Potentiation of GABAA-mediated synaptic current by ethanol in hippocampal CA1 neurons: possible role of protein kinase C. J Pharmacol Exp Ther 268:1388–1395
Weiner JL, Valenzuela CF, Watson PL, Frazier CJ, Dunwiddie TV (1997) Elevation of basal protein kinase C activity increases ethanol sensitivity of GABAA receptors in rat hippocampus CA1 pyramidal neurons. J Neurochem 68:1949–1959
Welsh BT, Goldstein BE, Mihic SJ (2009) Single-channel analysis of ethanol enhancement of glycine receptor function. J Pharmacol Exp Ther 330:198–205
Wenzel A, Fritschy JM, Mohler H, Benke D (1997) NMDA receptor heterogeneity during postnatal development of the rat brain: differential expression of the NR2A, NR2B, and NR2C subunit proteins. J Neurochem 68:469–478
White G, Lovinger DM, Weight FF (1990) Ethanol inhibits NMDA-activated current but does not alter GABA-activated current in an isolated adult mammalian neuron. Brain Res 507:332–336
Wilcox MC, Cuzon Carlson VC, Sherazee N, Sprow GM, Bock R, Thiele TE, Lovinger DM, Alvarez VA (2014) Repeated binge-like ethanol drinking alters ethanol drinking patterns and depresses striatal GABAergic transmission. Neuropsychopharmacology 39:579–594
Wilson CJ, Chang HT, Kitai ST (1990) Firing patterns and synaptic potentials of identified giant aspiny interneurons in the rat neostriatum. J Neurosci 10:508–519
Woodward JJ (1994) A comparison of the effects of ethanol and the competitive glycine antagonist 7-chlorokynurenic acid on N-methyl-D-aspartic acid-induced neurotransmitter release from hippocampa slices. J Neurochem 62:987–991
Woodward JJ (2000) Ethanol and NMDA receptor signaling. Crit Rev Neurobiol 14:69–89
Wu PH, Poelchen W, Proctor WR (2005) Differential GABAB receptor modulation of ethanol effects on GABAA synaptic activity in hippocampal CA1 neurons. J Pharmacol Exp Ther 312:1082–1089
Xia JX, Li K, Zhou R, Zhang XH, Ge YB, Ru Yuan X (2006) Alterations of rat corticostriatal synaptic plasticity after chronic ethanol exposure and withdrawal. Alcohol Clin Exp Res 30:819–824
Yaka R, Phamluong K, Ron D (2003) Scaffolding of Fyn kinase to the NMDA receptor determines brain region sensitivity to ethanol. J Neurosci 23:3623–3632
Yamakura T, Mori H, Masaki H, Shimoji K, Mishina M (1993) Different sensitivities of NMDA receptor channel subtypes to non-competitive antagonists. Neuroreport 4:687–690
Yamamoto Y, Nakanishi H, Takai N, Shimazoe T, Watanabe S, Kita H (1999) Expression of N-methyl-D-aspartate-dependent long-term potentiation in the neostriatal neurons in an in vitro slice after ethanol withdrawal of the rat. Neuroscience 91:59–68
Yin HH, Knowlton BJ, Balleine BW (2005a) Blockade of NMDA receptors in the dorsomedial striatum prevents action-outcome learning in instrumental conditioning. Eur J Neurosci 22:505–512
Yin HH, Ostlund SB, Knowlton BJ, Balleine BW (2005b) The role of the dorsomedial stratum in instrumental conditioning. Eur J Neurosci 22:513–523
Yin HH, Knowlton BJ, Balleine BW (2006) Inactivation of dorsolateral striatum enhances sensitivity in action-outcome contingency in instrumental conditioning. Behav Brain Res 166:189–196
Yuste R, Majewska A, Cash SS, Denk W (1999) Mechanisms of calcium influx into hippocampal spines: heterogeneity among spines, coincidence detection by NMDA receptors, and optical quantal analysis. J Neurosci 19:1976–1987
Zahm DS, Brog JS (1992) On the significance of subterritories in the “accumbens” part of the rat ventral striatum. Neuroscience 50:751–767
Zahm DS, Heimer L (1990) Two transpallidal pathways originating in the rat nucleus accumbens. J Comp Neurol 302:437–446
Zhou Q, Verdoorn TA, Lovinger DM (1998) Alcohols potentiate the function of 5-HT3 receptor-channels on NCB-20 neuroblastoma cells by favoring and stabilizing the open channel state. J Physiol 507:335–352
Zhou L, Furuta T, Kaneko T (2003) Chemical organization of projection neurons in the rat accumbens nucleus and olfactory tubercle. Neuroscience 120:783–798
Zhu PJ, Lovinger DM (2006) Ethanol potentiates GABAergic synaptic transmission in a postsynaptic neuron/synaptic bouton preparation from basolateral amygdala. J Neurophysiol 96:433–441
Ziskind-Conhaim L, Gao BX, Hinckley C (2003) Ethanol dual modulatory actions on spontaneous postsynaptic currents in spinal motoneurons. J Neurophysiol 89:806–813
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Cuzon Carlson, V.C. (2018). GABA and Glutamate Synaptic Coadaptations to Chronic Ethanol in the Striatum. In: Grant, K., Lovinger, D. (eds) The Neuropharmacology of Alcohol . Handbook of Experimental Pharmacology, vol 248. Springer, Cham. https://doi.org/10.1007/164_2018_98
Download citation
DOI: https://doi.org/10.1007/164_2018_98
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-96522-2
Online ISBN: 978-3-319-96523-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)