
Abstract
Adenosine triphosphate (ATP) plays major roles in cell metabolism as an energy supplier and as a substrate for enzymatic reactions. While ATP is well known for its role as an intracellular energy carrier, recent studies have found that ATP exists not only in the cytoplasm, but also in the extracellular matrix. Cytoplasmic ATP can be secreted into the apoplast through wound leakage, secretory vesicles, or transporters in the plasma membrane. As a signaling molecule, extracellular ATP (eATP) regulates plant metabolism, growth and development, and responses to biotic and abiotic stimuli. eATP binds to receptors in the plasma membrane, where it triggers the generation of second messengers, including Ca2+, NO, and reactive oxygen species. These second messengers induce expression of a series of functional genes that promote changes in the cellular structure and physiological function of plant cells. Here, we discuss the progress in research on the function and signaling properties of this novel apoplastic messenger.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
As an energy-carrying molecule, adenosine triphosphate (ATP) plays key roles in energy metabolism and as a substrate in signal transduction pathways. While ATP had historically been considered an intracellular molecule, its unexpected detection in the extracellular matrix (ECM) of animal and human cells (Drury and Szent-Györgyi 1929) led to extensive investigation of its existence, downstream signaling pathways, and physiological functions. It is now known that extracellular ATP (eATP), which can be found in the blood and tissue fluid, participates in a range of functions, including neuron transmission, blood pressure regulation, cell differentiation, smooth muscle contraction, exocrine and endocrine secretion, inflammation, platelet aggregation, pain, and cardiac function modulation. eATP is secreted from the cell through secretory vesicles generated in the Golgi apparatus or through transporters in the plasma membrane (PM). Deregulation of secretion can lead to abnormal levels of eATP, which in turn causes cell death, uncontrolled cell division, and aberrant cell differentiation, and can result in serious illness in animals and humans (reviewed by Ralevic and Burnstock 1998; Burnstock 2006, 2007; Burnstock et al. 2013; Idzko et al. 2014; Kennedy 2015; Cavaliere et al. 2015; Ferrari et al. 2015).
ATP influences cellular processes by binding to purinergic receptors in the PMs of target cells. There are two classes of purinergic receptors for ATP: P2X receptors and P2Y receptors. P2X receptors are ligand-gated ion channels and P2Y receptors are heterotrimeric G-protein-coupled receptors. The binding of ATP to these receptors in target cells triggers signaling cascades that lead to transmembrane signal transduction and the generation of second messengers. These second messengers include cytosolic Ca2+, nitrogen monoxide (NO), reactive oxygen species (ROS), hydrogen peroxide (H2O2), inositol triphosphate (IP3), and diacetyl glycerol (DAG). A series of biochemical reactions occurs downstream of second messengers, including the phosphorylation and dephosphorylation of structural and catalytic proteins, as well as the activation of transcription factors and the genes they promote. These reactions induce changes in cell metabolism and development, which can eventually lead to changes in the structure and function of the entire body (reviewed by Ralevic and Burnstock 1998; Burnstock 2006, 2007; Burnstock et al. 2013; Idzko et al. 2014; Kennedy 2015; Cavaliere et al. 2015; Ferrari et al. 2015).
In the 1970s, plant biologists first investigated the effects of exogenous ATP on plant metabolism and the responses of plants to stimuli. These early studies found that addition of ATP promotes fast closing of venus fly trap leaves (Jaffe 1973), endonuclease synthesis (Udvardy and Farkas 1973), and stomatal opening (Raghavendra 1981; Nejidat et al. 1983). At the time, these effects were attributed ATP absorption and increased intracellular energy supply. Lüttge et al. (1974) reported that ATP promoted K+ absorption of cut oat leaves by chelating inhibitory divalent cations (e.g., Ca2+ and Mg2+). The possibility that ATP could act as a messenger in the extracellular space was realized only decades later, in the 1990s. Due in large part to the ideas and methodologies established in research on the roles of eATP in animals, research on eATP in plants has made some advances during the past two decades. ATP is now known to be prevalent in the apoplast of various plant species. In addition, eATP level is correlated with the viability and growth rate of plant cells, suggesting it is a key regulator of cellular metabolism. Results of pharmacological experiments revealed that cytoplasmic ATP is secreted via presumed ATP transporters, by secretory vesicles originating in the Golgi apparatus, or during the stress response caused by a wound to the PM. eATP signal transduction has been actively investigated in the last decade. Work from these studies has shown that binding of eATP to its receptors stimulates transmembrane signaling events, including the generation of second messengers. Unsurprisingly, most of the second messengers involved in eATP signaling in animal cells are also involved in eATP-induced physiological responses in plant cells. These messengers directly or indirectly regulate metabolic reactions and gene expression, resulting in changes in structure, function, growth, and development of plant cells under different conditions (reviewed by Roux and Steinebrunner 2007; Clark and Roux 2009, 2011; Tanaka et al. 2010a, 2014; Chivasa and Slabas 2012; Cao et al. 2014). So far, research on eATP has concentrated on three fields: its physiological function, its secretion mechanisms, and its signal transduction. These three fields will be the focus of this review.
2 The Physiological Function of eATP
Researchers have manipulated ATP levels in experimental systems as a means by which to investigate the roles of eATP in plant cell growth, development, and stress responses. A simple way to elevate eATP level is by adding ATP into an experimental system. Generally, to ensure eATP acts as a signaling molecule rather than as an energy molecule, the effects of weakly hydrolyzable ATP analogs must be investigated after determining the effects of added ATP. Three widely used ATP analogs are ATPγS (adenosine 5′-O-(3-thio) triphosphate), Bz-ATP (3′-O-(4-benzoyl) benzoyl adenosine 5′-triphosphate), and 2me-ATP (2-methylthio-adenosine 5′-triphosphate). Since these analogs can only weakly be hydrolyzed by ATPase, they are unlikely to participate in energy-consuming or substrate phosphorylation reactions. Therefore, if the effects of ATP analogs are similar to those of exogenous ATP, it can be concluded that ATP participates in these reactions as a signaling molecule. Moreover, to verify the specificity of ATP in the physiological functions of plant cells, the effects of other nucleotides, including ADP (adenosine di-phosphate), AMP (adenosine mono-phosphate), adenosine, GTP (guanosine-5′-triphosphate), CTP (cytidine triphosphate), and TTP (thymidine triphosphate) should also be investigated.
ATP hydrolysis enzymes such as apyrase (ATP-diphosphatase) and glucose hexokinase can be used to decrease eATP levels. In early experiments, these enzymes were added into the bath solution of cultured plant cells or smeared onto the surface of plant organs. The effects of these enzymes were presumed to result from eATP depletion-induced physiological reactions. In later studies, levels of extracellular apyrase have been manipulated through transgenic regulation. Phenotypic differences between wild-type and apyrase null mutants or overexpression lines were thought to result from changes in eATP levels. However, Schiller et al. (2012) found that an apyrase in Arabidopsis thaliana, AtAPY1, localized to the Golgi apparatus rather than to the ECM. Activity assays revealed that AtAPY1 prefers to use GDP and UDP as substrates, rather than ATP. These results indicated that not all apyrases localize to the ECM and participate in eATP turnover-related physiological responses. Therefore, the effects of transgenic modification of apyrases must be very carefully evaluated before concluding the result from eATP degradation.
Despite the challenges associated with manipulating ATP levels, findings from a large body of research have led to the conclusion that eATP is involved in the regulation of the following processes as a multifunctional messenger:
2.1 Cell Viability
Chivasa et al. (2005) reported that eATP is necessary for maintaining the viability of plant cells. They found that suspension-cultured A. thaliana cells released ATP into the bathing medium. Addition of apyrase and glucose–hexokinase led to reduced levels of ATP in the medium and to the death of cultured cells, indicating that deprivation of eATP compromised cell viability. Likewise, addition of AMP-PCP (β,γ-methylene adenosine 5′-triphosphate), a weakly hydrolyzable ATP analog that competitively inhibits binding of eATP to its receptor, also led to death of cultured cells, providing further evidence that eATP signaling is required for cell viability. When AMP-PCP or ATP hydrolyzing enzymes were smeared onto the surface of growing leaves of A. thaliana, maize, tobacco, and soybean, the tissues within the treated zone collapsed, while tissues outside of the treated zone retained a healthy appearance (Fig. 1). These results suggest that blockade or impairment of eATP signaling leads to cell death, and led the authors to conclude that eATP could be involved in maintaining key metabolic processes in plant cells.

ATP hydrolysis enzymes and analogs lead to death of Arabidopsis tissues. Apyrase, glucose–hexokinase (Glc-Hk), or AMP-PCP was smeared on localized area (top row) or entire leaves (bottom row) of Arabidopsis thaliana leaves. Leaf cells being treated eventually died (Chivasa et al. 2005)
To verify the physiological function of eATP depletion-induced cell death, Chivasa et al. (2005) investigated the effect of fumonisin B1 (FB1), a programmed cell death-eliciting mycotoxin, on eATP level and cell viability of A. thaliana. FB1 treatment remarkably decreased eATP level and led to severe injury or death of plant cells. Exogenous ATP markedly rescued plant cells from FB1-induced cell death (Fig. 2). These results suggest that this mycotoxin causes cell death partially through promoting eATP degradation, and that conversely, exogenous ATP stimulates protective reactions, thereby rescuing plant cells.

ATP rescues Arabidopsis plants from FB1-induced death. Arabidopsis thaliana plants were transferred and being cultured on nutrient agar containing 1 μM FB1 or 1 μM FB1 mixed with 1 mM ATP for 5 days. Top row, retardation of growth in all treatments containing FB1. Bottom row, close-up photographs of representative plants from corresponding plates in the top row (Chivasa et al. 2005)
Findings from another research group were at odds with a protective role for ATP, revealing instead that eATP induces programmed cell death (PCD). Sun et al. (2012a, b) reported that eATP triggered PCD of cultured Populus euphratica cells in a dose-dependent manner: high concentrations (≥0.5 mM) of ATP induced PCD while low concentrations of eATP did not. Specifically, high levels of ATP led to an increase in DNA fragmentation, chromatin condensation, and caspase activity, which are all typical features of PCD. It is not excluded that high apoplastic eATP concentrations of about 0.5 mM have physiological functions, because cytoplasmic ATP concentration is 1–2 mM and ATP can be released from a wound after injury. The disparate conclusions from Sun et al. (2012a, b) and Chivasa et al. (2005) were not due to differences in ATP levels in the culture medium, as both studies used similar concentrations (around 1 mM). Instead, their results may indicate that different plant species respond to eATP in distinct manners. Sun et al. (2012a, b) suggest a physiological function of eATP-induced PCD in which biotic or abiotic stresses stimulate ATP release, leading to transient or sustained increases in eATP level. Then, eATP-induced PCD may eliminate injured or infected cells to prevent further injury of adjacent cells, and may be an important step in the plant response to stress.
2.2 Vegetative Growth
A large body of work supports a role for eATP in promoting vegetative growth. Thomas et al. (1999) reported that exogenous apyrase promotes growth of Arabidopsis cells. In their experiments, apoplastic ATP, which can be degraded by apyrase to release phosphate, was regarded as a source of inorganic phosphate (Pi), a nutrient important for plant growth. Lew and Dearnaley (2000) reported that exogenous ATP, ADP, and GTP promoted root hair growth and depolarized the PM potential of growing root hairs. In addition, Kim et al. (2006) reported that eATP localization and concentration are positively correlated with cell expansion. Specifically, eATP concentration is higher in the walls of elongating cells such as root epidermal cells in the elongation zone and division zone (Fig. 3). In growing root hairs, a tip to base eATP gradient was detected and is thought to be the basis for root hair elongation, since this gradient is less prominent or missing in the cell walls of non-growing root hairs. Finally, Tonón et al. (2010) reported that etiolated Arabidopsis seedlings growing in reduced medium containing glutathione (GSH) or dithiothreitol (DTT) show decreased hypocotyl elongation rate. Addition of ATP effectively restores the normal growth of GSH-suppressed hypocotyl elongation. Additional data suggests that eATP-stimulated ROS generation plays a basic role in maintaining plant cell growth rate.

The distribution of eATP on the root surface of Medicago truncatula. CBD (cell wall binding domain peptide)-luciferase reporter was used to visualize eATP. Intensity-coded (inset range in c, where white areas show the strongest ATP concentration and black is the background) images showing ATP to exist in interstitial spaces at growing regions of the root [including meristematic region (a), elongation region (b), root hairs (b), and etiolated hypocotyl (d)] (Kim et al. 2006)
Although the above findings demonstrate that eATP positively regulates plant growth, still other findings suggest that eATP suppresses plant cell growth. Wu et al. (2007) reported that the expression levels of two A. thaliana apyrase genes, APY1 and APY2, are correlated with growth rate. Expression of APY1 and APY2 was markedly higher in rapidly growing tissue and cell types such as root tip, root cap, columella cells, and the root–hypocotyl junction than in other tissues and cells. Expression of apyrases significantly affected growth rate of seedlings both in light and in darkness. In apy1/apy2 double-knockout mutants, the growth rates of the primary roots of seedlings growing in light and the hypocotyls of etiolated seedlings growing in darkness were both markedly lower than those in wild-type plants. Furthermore, Wolf et al. (2007) found that in A. thaliana apy1/apy2 double-knockout mutants, root and shoot meristems were abnormal and far less functional, and the morphological features of cotyledons were different from wild-type plants. Some apy1/apy2 double-knockout mutants were seedling lethal or had developmental defects in cell division and growth. Riewe et al. (2008) reported that apyrase located in the apoplast of potato cells plays essential roles in tuber generation and development. Suppression of apyrase expression led to increased tuber number and decreased tuber size. In addition, tubers of the apyrase null mutants had more longitudinal shapes than wild-type tubers. Decreased apyrase expression in the tubers also changed their gene expression patterns. Specifically, increased expression was observed in genes encoding cell wall proteins related to cell elongation, carbon and energy provision for starch biosynthesis, and mitochondrial ATP/ADP translocators, whereas decreased expression was observed in key enzymes related to starch synthesis. These data indicate that sufficient apyrase molecules are needed for normal growth of vegetative organs and reproductive cells. Since the main function of apoplast apyrase is to hydrolyze ATP, it can be concluded that eATP accumulation may not always be beneficial for cell growth, and that eATP hydrolysis may be necessary for maintaining stable growth and development.
Consistent with this idea, Clark et al. (2010) demonstrated that levels of eATP affect its physiological functions. They reported that eATP is involved in growth of cotton fibers. When cotton fibers enter their rapid growth phase, there is a significant increase in the expression of two extracellular apyrases in cotton: GhAPY1 and GhAPY2. In a cultured ovule system, growing fibers released ATP into culture medium. Inhibition of apyrase activity by apyrase inhibitor or apyrase antibodies led to increased eATP level in the medium and inhibited fiber growth. Although addition of ATPγS at a low concentration (30 μM) promoted fiber growth, addition at a high concentration (150 μM) suppressed fiber growth (Fig. 4). These data suggest that eATP released by ovule cells is required for cotton fiber growth. However, since sustained fiber growth requires steady, low eATP levels, the timely hydrolysis of ATP by apyrase is necessary in order to prevent eATP over accumulation.

High concentration (150 μM) of ATPγS decreased cotton fiber growth while low concentration (30 μM) of ATPγS increased it (Clark et al. 2010)
2.3 Pollen Germination and Pollen Tube Growth
Steinebrunner et al. (2003) found that pollen germination (PG) and pollen tube growth (PTG) of A. thaliana were suppressed by apyrase inhibitors, and completely blocked by double-knockout mutagenesis of APY1 and APY2. However, complementation with either one of these apyrases rescued PG in the double knockouts. These results indicate that apyrases are required for PG and PTG. Moreover, they suggested that quenching of an eATP signal may be one possible reason for apyrase suppressed PG and PTG. Furthermore, Reichler et al. (2009) reported that addition of ATPγS markedly inhibited PG and PTG of wild-type A. thaliana. Nevertheless, the dynamics and function of endogenous eATP in pollen grain cell wall are still uncertain. Whether there is any positive regulatory role of eATP in PG and PTG needs to be clarified.
2.4 Stomatal Movement
Thirty years ago, it was reported that exogenous ATP promoted stomatal opening in Commelina benghalensis (Raghavendra 1981) and Commelina communis (Nejidat et al. 1983). In those studies, ATP-promoted stomatal opening was thought to result from increased energy supply. However, recent research revealed that eATP may function as a signaling molecule, rather than as an energy carrier, to promote stomatal movement. Clark et al. (2011) found such a role for ATP in A. thaliana by showing that both exogenous ATP and ATPγS promote stomatal movement. Interestingly, both stimuli that promote stomatal opening (light) and stimuli that promote stomatal closing (ABA) lead to eATP increase and apyrase accumulation in the outer space of guard cells, indicating a role for eATP as a signal transducer in these processes. It was also reported that ATP, ADP, GTP, and weakly hydrolyzable ATP analogs (ATPγS, Bz-ATP, and 2meATP) promoted stomatal opening of A. thaliana (Hao et al. 2012) and Vicia faba (Wang et al. 2014) in light and darkness, further proving the involvement of purine signals in stomatal movement.
2.5 Root Gravitropism
Tang et al. (2003) reported that addition of 1–2 mM ATP strongly blocked gravitropic growth of Arabidopsis root, leading to horizontal growth of roots that normally grow vertically. Higher concentration of ATP (3 mM) even led to root curling. Later investigation of the mechanisms of eATP-inhibited root gravitropism revealed that eATP disturbs auxin distribution in root cells by inhibiting auxin export from root tip cells (Liu et al. 2012). Root growth orientation is sensitive to environmental stimuli, including gravity and light. The inhibitory effect of ATP on root gravitropism can be defined as eATP-regulated root growth reorientation. The physiological significance of this process remains to be clarified.
2.6 Nodulation
Etzler et al. (1999) and Day et al. (2000) reported that eATP levels affect root nodulation of leguminous plants. Nod factors released by rhizobia promote expression and secretion of apyrase. Apyrase binds with nod factors, decomposes eATP, and promotes nodulation of Dolichos biflorus and Glycine soja roots. Consistent with this, function-blocking antibodies against apyrase markedly inhibit nodulation. McAlvin and Stacey (2005) also found that overexpression of apyrase genes in Lotus japonica led to a remarkable increase in root nodules, while suppression of apyrase expression resulted in decreased nodulation. These results indicate that eATP may be involved in the communication between plant cell and outer microorganisms.
2.7 Stress Responses
Several studies have shown that eATP leads to reduced plant resistance to antibiotics or herbicides (Thomas et al. 2000; Windsor et al. 2003; Chivasa et al. 2009). Specifically, Thomas et al. (2000) reported that ATP markedly increased the sensitivity of yeast and Arabidopsis cells to cycloheximide. Likewise, apyrase decreased eATP level and enhanced resistance of these cells to cycloheximide. Windsor et al. (2003) also reported that overexpression of apyrase in Arabidopsis markedly promoted the resistance of plants to herbicides. Cells of apyrase overexpression lines excreted more herbicides than wild-type, which weakened the effects of herbicides on plant cells. Results from Chivasa et al. (2009) showed that exogenous glucose–hexokinase or AMP-PCP led to death of tobacco leaf cells, though leaf cells grown in darkness were far less sensitive to glucose–hexokinase and AMP-PCP than leaf cells grown in light. eATP depletion resulted in increased expression of pathogenesis-related genes and enhanced resistance to tobacco mosaic virus. Under low-light conditions, AMP-PCP treatment induced expression of three pathogenesis-related genes: PR-1, PR-2, and PR-5. In tobacco leaves pretreated with glucose–hexokinase or AMP-PCP, growth of inoculated tobacco mosaic virus was significantly suppressed. Glucose–hexokinase or AMP-PCP pretreatment also promoted leaf cells’ resistance to the bacteria Pseudomonas syringae pv. tabaci. These results indicate that eATP effectively suppresses plant pathogen resistance (PR). Negative regulation of PR responses by eATP may have important physiological roles. Fast-growing young tissues, which can respond to pathogens rapidly and effectively, have high eATP levels. Chivasa et al. (2009) suggest that these eATP may switch off unnecessary defensive responses to let cells grow and develop intensively. In older senescent cells, which are unable to respond efficiently to pathogens, eATP levels decline and expression of genes related to PR will be switched on, preparing for possible attacks.
Most recently, eATP was reported to be involved in cold tolerance of P. euphratica (Deng et al. 2015). Cold stress stimulated the release of ATP. Addition of low concentrations of ATP promoted cold tolerance, while addition of high concentrations of ATP inhibited it. Furthermore, low temperature upregulated the expression of the apyrase PeAPY2 in callus cells. Ectopic expression of PeAPY2 in A. thaliana strongly enhanced the cold tolerance of transgenic seedlings. While cold stimulation led to injury of the PM and leakage of intracellular solutes in wild-type plants, metabolite leakage was reduced in PeAPY2 transgenic plants. Transgenic plants also showed increased vesicular trafficking (e.g., endocytosis and exocytosis) during cold treatment and recovery in normal temperature. Exogenous ATP at low concentrations also accelerated vesicular trafficking, while ATP at high concentrations suppressed it. Based on these results, it was concluded that cold stress-induced eATP elevation may stimulate vesicular trafficking and efficient PM repair. However, continuous accumulation of eATP hurts cell viability, so apyrase is needed to drop eATP concentration to a safe level. Under cold stress, an accurate regulatory mechanism controlling ATP release and turnover is required for modulating cellular responses related to cold tolerance.
Kim et al. (2009) found that eATP had a positive effect on hypertonic stress responses of Arabidopsis seedlings. ATP release and apyrase accumulation in the apoplast were stimulated in high concentration NaCl, Mg(NO3)2, or MgCl2 solutions. Accumulated eATP enhanced cell resistance to hypertonic stress. Apyrase-mediated eATP degradation, which occurs after eATP triggers downstream signal transduction cascades, may also be a necessary step in hypertonic tolerance. Addition of AMP-PCP to competitively block eATP signaling resulted in hypertonic stress and cell death.
eATP is also involved in salt tolerance of suspension-cultured P. euphratica cells (Sun et al. 2012a, b). Addition of NaCl induced ATP release and stimulation of salt-tolerance responses. Depletion of ATP by exogenous ATP hydrolase or blockade of eATP signaling by PPADS (pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid) and suramin resulted in a significant decrease in the viability of NaCl treated cells. These results indicate that eATP is needed for maintaining cell viability under salt stress.
The physiological roles of eATP in stress tolerance have been discussed by Choi et al. (2014b). When plant or animal cells are damaged by physical, chemical, or biotic stimuli, intracellular molecules known as damage-associated molecular patterns (DAMPs) are released through the wound to initiate defense responses. In animals, eATP has been considered a DAMP. Since in plants, eATP is released to extracellular spaces when cells are injured by biotic or abiotic stresses, and triggers signal cascades to induce resistant responses, it is reasonable to consider it a DAMP in plants.
Different labs have demonstrated seemingly contradictory effects of eATP on plant growth, development, and stress resistance. Relatively low concentrations of eATP are necessary for plant growth, development, and stress resistance, while high concentrations of eATP lead to growth inhibition or even PCD. Choi et al. (2014b) suggested a model that illustrates the mechanism by which eATP may perform such seemingly contradictory effects. As shown in Fig. 5, eATP concentration is normally maintained at a certain level by the balance of ATP secretion and hydrolysis. This level is beneficial for plant cell growth and development. Small or moderate increases or decreases in eATP level will lead to defense responses, including inhibited plant cell growth rate and special metabolite accumulation. However, very low or very high eATP levels can lead to PCD. This model explains the effects of exogenous ATP or apyrase on plant cell growth, development, and stress resistance. Nevertheless, the mechanism by which low and high levels of eATP can stimulate similar responses remains to be verified.

A model of the effect of eATP concentration on plant growth and immunity. When eATP levels deviate from the optimal concentration for plant growth, ATP serves as a danger signal to inform cells of their abnormal status. Those concentrations that deviate slightly from optimal extracellular ATP levels appear to trigger plant defense responses. In contrast, extreme depletion or excess of extracellular ATP is correlated with cell death (Choi et al. 2014b)
3 The Existence and Secretion of eATP
Although application of exogenous ATP or apyrase shows remarkable effects in various experiments, the existence of endogenous eATP must be verified before it can be defined as an apoplastic messenger. To measure eATP level in the ECM, ATP was extracted and its concentration in extracted fluid was measured to estimate the eATP level in cell wall. Estimates of the resting eATP concentration in the ECM range from dozens of nM to dozens of μM (Thomas et al. 2000; Song et al. 2006; Wu et al. 2008; Clark et al. 2010; Weerasinghe et al. 2009). Nevertheless, methods which can in situ accurately measure the eATP level in the apoplast are still absent.
In most experiments, ATP concentration was calibrated by using the bioluminescent indicator luciferin. Luciferase catalyzes the oxidation of luciferin in an ATP-dependent manner, generating a fluorescent signal that can be detected by illuminance-detecting instruments. Illuminant intensity is correlated to ATP concentration. To detect eATP localization and dynamics in situ, Kim et al. (2006) fused firefly luciferase with a peptide containing cellulose binding domain (CBD). The CBD peptide ensures that the fused proteins can be anchored to the cell wall. The localization and density of bioluminescence reflects the localization and relative concentration of ATP in the apoplast. Using this indicator, they visualized eATP in the cell wall of A. thaliana, alfalfa (Medicago truncatula), wheat (Triticum aestivum), and lotus (Lotus japonicus). Development of this method prompted investigation of eATP localization, secretion, turnover, and dynamics in a number of plant systems.
Most recently, Vanegas et al. (2015) developed a self-referencing electrochemical biosensor for direct ATP flux measurement. They coated a grapheme-functionalized platinum microelectrode with a laponite sol gel containing two enzymes: glycerol kinase and glycerol-3-phosphate oxidase. By detecting electro-potential dynamics driven by enzyme-catalyzed ATP degradation, the dynamics of ATP concentration in solution could be sensitively detected. This electrochemical biosensor has been used to rapidly and sensitively detect touch- and wound-stimulated ATP efflux from maize (Zea mays) root cells and light-induced ATP release from developing Ceratopteris spores. This biochemical biosensor has proven to be a useful tool for non-invasive measurement of eATP secretion from living cells.
In animal cells, ATP is released through secretory vesicles, specific transporters, or PM wound. Studies using the methods described above revealed that similar mechanisms of secretion might also exist in plants. Secretory vesicles originating from the Golgi apparatus are major resources of eATP. Kim et al. (2006) reported that eATP concentration is correlated with extracellular Ca2+. The high eATP level at the tip of root hairs was decreased by the Ca2+ chelator EGTA or the Ca2+ channel blocker (Gd3+), and increased by addition of exogenous Ca2+, indicating that Ca2+ influx is necessary for eATP secretion. Wu et al. (2008) also found that EGTA and the Ca2+ channel blocker La3+ strongly inhibited ATP release from cultured hairy roots of Salvia miltiorrhiza. Since cytosolic Ca2+ is an essential regulator of secretory vesicle transport and fusion with PM, these results suggest that eATP may be secreted through vesicles. Further support for the role of these vesicles in eATP secretion comes from the finding that Brefeldin A, a toxin that inhibits vesicle transport, significantly decreased eATP accumulation in root hair tip (Kim et al. 2006).
ATP Binding Cassette (ABC) transporters play crucial roles in ATP secretion in animal cells. Although similar transporters have been detected in plant cells (Crouzet et al. 2006), their role in eATP secretion remains uncertain. Thomas et al. (2000) reported that overexpression of some members of the plant ABC transporter family resulted in accelerated ATP secretion in yeast and A. thaliana.
In animals, mechanical touch and stress stimulate ATP release (Bodin and Burnstock 2001). Similar mechanisms have been found in plants. Touch stimulation and osmotic stress increased eATP level in Arabidopsis seedlings (Jeter et al. 2004). Mechanical touch also stimulated ATP release from Arabidopsis root tip cells (Weerasinghe et al. 2009). Kim et al. (2006) reported that treatment with a chitin mixture, to mimic a pathogen attack, promoted ATP secretion. In addition, Wu et al. (2008) reported that polysaccharide elicitors in yeast extract promoted ATP release from cultured hairy roots of S. miltiorrhiza. Hypertonic stress also stimulated ATP secretion and promoted a cell resistance response (Kim et al. 2009). Osmotic stress (high concentration of sorbitol), ABA, and glutamate also induced ATP release from A. thaliana roots (Dark et al. 2011). Finally, wounding-induced eATP accumulation played crucial roles in inducing protective responses (Song et al. 2006). In stress-induced ATP release, ATP efflux may occur primarily through PM wound. These eATP molecules may be required for triggering cytoplasmic reactions to enhance tolerance of plant cells to related stresses.
4 The Signal Transduction of eATP
In animal systems, eATP participates in cell signaling in two ways: (1) as a substrate to regulate protein activity through phosphorylation, and (2) as a primary messenger to stimulate signaling cascades through binding of eATP receptors in the PM. eATP may play similar roles in plant systems. Using proteomic analysis, Chivasa et al. (2002) identified phosphorylated proteins in the ECM of plant cells. Although there is no direct evidence that eATP participates in protein phosphorylation outside the cell, the possibility cannot be excluded.
Research concepts and methods used widely in animal eATP signaling research have been adopted in most studies of plant eATP signal transduction. Although these methods may be applicable in plant research based on the similarities between plant and animal cells, the distinct characteristics of plant cells may require additional specific research ideas and methods.
4.1 eATP Receptors
In early studies, animal eATP receptor antagonists or inhibitors were widely used in research on plant eATP signaling. Three such commonly used antagonists, PPADS, suramin, and reactive blue (RB), effectively inhibited or blocked eATP-induced physiological effects in most plant studies in which they were used (Demidchik et al. 2003; Chivasa et al. 2005; Song et al. 2006; Sun et al. 2012a). However, genomic sequence surveys revealed that the animal P2-like receptor, which is sensitive to these antagonists, exists only in green algae (however, it is proved to be exist intracellularly rather than in PM) (Fountain et al. 2008) and not in higher plants. The target of these antagonists in higher plant cells needs to be verified before any conclusions can be drawn about the roles of eATP in plant cells.
In animal cells, eATP binding with P2X receptors, which are ligand-gated ion channels, triggers Ca2+ influx into cytoplasm. eATP-triggered Ca2+ influx has been detected in several plant species. Furthermore, Ca2+ influx channel was identified using electrophysiological methods (Demidchik et al. 2009; Wang et al. 2014). Based on these data, it is reasonable to propose that eATP receptor-like Ca2+ channels may exist in the PM of plant cells.
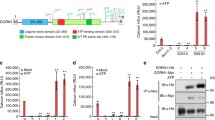
Recently, an eATP receptor was identified in A. thaliana (Choi et al. 2014a). By screening ethyl methanesulfonate (EMS)-mutagenized seedlings, 2 mutants that were unresponsive to ATP stimulation were identified. The mutants, named dorn1, had impairments in ATP-induced [Ca2+]cyt increase, MAPK activation, and gene expression, all of which are typical ATP-inducible responses in wild-type plants. DORN1 encodes a lectin receptor kinase (LecRK-1.9) that consists of three domains: (1) extracellular legume-type lectin domain, (2) single transmembrane domain, and (3) intracellular serine/threonine kinase domain. T-DNA insertion mutagenesis revealed that point mutation of each domain caused complete impairment of eATP-induced cellular responses, indicating that all three domains are necessary for eATP signaling (Choi et al. 2014b). Reduced LecRK-1.9 gene expression resulted in suppressed eATP responses, while its overexpression resulted in strengthened ATP responses. Likewise, most wounding- or eATP-induced gene expression was suppressed in dorn1 and significantly promoted in DORN1 overexpression lines. These findings indicate that ATP released upon wounding binds to DORN1, stimulating signaling that triggers functional gene expression. Since DORN1 possesses protein kinase activity, it was later named P2K receptor to distinguish it from P2X and P2Y receptors in animal cells. Ectopic expression of the Arabidopsis LecRK1.9 (DORN1) gene in tobacco and potato plants significantly enhanced their resistance to Phytophthora infestans (Bouwmeester et al. 2013), indicating the involvement of P2K-related ATP signaling in pathogen attack responses.
4.2 Transmembrane Signal Transduction and Generation of Second Messengers
4.2.1 Heterotrimeric G Protein
Data suggest that heterotrimeric G proteins in PM may be involved in eATP signaling in plant systems. Weerasinghe et al. (2009) reported that heterotrimeric G protein is involved in the eATP-regulated obstacle-avoidance response of A. thaliana roots. Roots of Gα/Gβ double null mutants do not respond to touch stimulation and change root growth direction more slowly than wild-type roots. In addition, the refractory period after touch-induced ATP release is significantly attenuated in the Gα/Gβ double null mutant compared to wild-type. Based on these results, the authors suggest that touch-induced ATP release and the fine-tuning of eATP level that follows, both of which are controlled primarily by heterotrimeric G proteins, may be key signaling events in the obstacle-avoidance response. Tanaka et al. (2010b) suggest Gα and Gβ subunits participate in ATP-stimulated Ca2+ increase to different degrees. They found that in Gα null mutants, eATP-stimulated Ca2+ increase was similar to that of wild-type, while in Gβ null mutants, eATP-stimulated Ca2+ increase was enhanced, indicating that Gβ may be a negative regulator of eATP-induced cytoplasmic Ca2+ ([Ca2+]cyt) increase. Hao et al. (2012) reported that eATP promoted stomatal opening and generation of secondary messengers (Ca2+, ROS) in A. thaliana, but that these processes were impaired in two Gα null mutants: gpa1-1 and gpa1-2. These findings indicate that heterotrimeric G protein may participate in eATP signaling in guard cells. The role of heterotrimeric G protein in transmembrane signal transduction of eATP needs further verification.
4.2.2 Ca2+
Ca2+ was the first agent studied for its relation to plant eATP signaling, and remains the most widely investigated. Exogenous ATP stimulates transient Ca2+ elevation or Ca2+ oscillation and is thought to be involved in eATP-regulated physiological functions in plant cells.
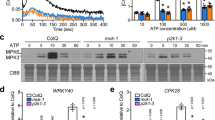
Pharmacological manipulation of Ca2+ channels has revealed distinct sources of eATP-induced Ca2+. Exogenous ATP induces a transient two-peak elevation curve of cytosolic Ca2+ concentration ([Ca2+]cyt) in A. thaliana root cells (Demidchik et al. 2003; Tanaka et al. 2010b). eATP-induced Ca2+ dynamics were inhibited by a Ca2+ chelator (EGTA) and Ca2+ channel blockers (Demidchik et al. 2003; Jeter et al. 2004; Hao et al. 2012). However, while ATP-induced gene expression was strongly inhibited by the Ca2+ channel blockers La3+ and Gd3+, it was unaffected by the Ca2+ release inhibitor U73122, indicating that Ca2+ influx, rather than Ca2+ release, may be the main source of eATP-induced Ca2+ (Jeter et al. 2004). A key mediator of eATP-induced Ca2+ influx is a voltage-dependent Ca2+ channel in PM that is activated by hyperpolarization of the PM potential (Demidchik et al. 2009; Wang et al. 2014). Tanaka et al. (2010b) suggested that in the ATP-induced two-peak Ca2+ increase, the first peak may result from Ca2+ influx through Gd3+ sensitive channels in the PM, and the second peak may result from both Ca2+ influx from ECM and Ca2+ release from intracellular Ca2+ stores.
A surprising result from Tanaka et al. (2010b) is that eATP induces a 4–6 peak [Ca2+]cyt oscillation when the apyrase inhibitor NGXT191 is present in the apoplast of Arabidopsis cells. This oscillation can be blocked by Gd3+ or U73122, suggesting it is mediated by both Ca2+ influx and Ca2+ release. eATP did not induce [Ca2+]cyt oscillations in the absence of NGXT191, indicating that suppression of apyrase activity is necessary for its occurrence. Brefeldin A also blocked eATP-induced [Ca2+]cyt oscillations, indicating that when apyrase is inhibited by NGXT191, increased eATP levels may stimulate further ATP release via secretory vesicles. Released ATP may stimulate Ca2+ influx and release, eventually initiating [Ca2+]cyt oscillation. The authors propose that the ATP-induced ATP release and [Ca2+]cyt oscillation under conditions of apyrase suppression may be critical for several physiological responses. Since decreased apyrase gene expression results in significant changes in plant cell growth and responses to stimuli, it can be deduced that constantly accumulated eATP may regulate physiological reactions through cytosolic Ca2+ oscillations.
4.2.3 Reactive Oxygen Species
In plant cells, ROS concentration in the cytoplasm is correlated with eATP level. More specifically, exogenous ATP induced accumulation of ROS (including superoxide anions, hydroxyl radicals, and H2O2), whereas exogenous apyrase induced a decrease in ROS (Kim et al. 2006; Song et al. 2006; Demidchik et al. 2009; Tonón et al. 2010; Hao et al. 2012; Sun et al. 2012b; Wang et al. 2014; Lim et al. 2014).
NADPH oxidase in the PM is a key component of eATP-induced ROS generation. The NADPH oxidase inhibitor diphenylene iodonium (DPI) effectively inhibited eATP-induced ROS accumulation. In addition, eATP-induced ROS generation was impaired in the null mutants of NADPH oxidase, rhd-2, and atrbohD/F (Song et al. 2006; Demidchik et al. 2009; Tonón et al. 2010; Hao et al. 2012; Wang et al. 2014). It has also been revealed that heterotrimeric G protein in PM (Hao et al. 2012), cytoplasmic Ca2+, and calmodulin (Song et al. 2006) may be involved in eATP-stimulated NADPH oxidase activation.
eATP-induced ROS is involved in a number of functions related to the regulation of plant cell growth, development, and stress tolerance. These include root hair growth (Kim et al. 2006), defense response (Kim et al. 2006; Song et al. 2006), stomatal movement (Hao et al. 2012; Wang et al. 2014), cell wall construction (Lim et al. 2014), hypocotyl growth of etiolated seedlings (Tonón et al. 2010), and PCD (Sun et al. 2012a, b). Though little is known about ROS-regulated downstream responses, ROS may stimulate ion transport (Demidchik et al. 2009; Hao et al. 2012; Wang et al. 2014) and expression of several defense genes (Song et al. 2006).
4.2.4 Nitrogen Monoxide
NO has been detected in eATP-treated plant cells and is an important component of eATP signaling (Foresi et al. 2007; Wu and Wu 2008; Reichler et al. 2009; Tonón et al. 2010; Clark et al. 2011). NO is involved in eATP-regulated cell growth (Foresi et al. 2007; Wu and Wu 2008), pollen germination (Reichler et al. 2009), hypocotyl growth (Tonón et al. 2010), and stomatal movement (Clark et al. 2011).
Wu and Wu (2008) reported that in S. miltiorrhiza hairy root cells, NO production could be detected 30 min after ATP treatment. NO level was increased by low doses of ATP (10–100 μM) and decreased by higher doses of ATP (>100 μM). Results of pharmacological experiments suggest that NO is generated from reactions catalyzed by NO synthase and/or nitrate reductase (Wu and Wu 2008; Tonón et al. 2010), which rely on Ca2+ and calmodulin. The key role of these enzymes in eATP-triggered signaling and physiological responses is supported by findings in null mutants of nitrate reductase (nia1nia2) or NO synthase, in which eATP-stimulated NO accumulation and physiological reactions were impaired (Reichler et al. 2009; Clark et al. 2011).
4.3 eATP-Regulated Gene Expression
Investigation of gene expression after eATP or apyrase treatment is necessary to clearly understand the role and working mechanism of eATP. eATP-regulated gene expression was investigated in several plant species using cDNA microarray or proteomic analysis methods.
Jeter et al. (2004) reported that wound, exogenous eATP, and osmotic stress all led to increased expression of MAPK signaling components. eATP-induced expression of ethylene biosynthetic enzymes and several ethylene responsive factors (ERF). So, eATP may trigger downstream reactions related to stress tolerance through ethylene signaling and MAPK cascades.
Chivasa et al. (2010) found that ATP upregulated or downregulated many functional genes in tobacco cells. The downregulated proteins include photosynthetic proteins, mitochondrial ATP synthetic proteins, redox proteins, and some defense proteins. Consistent with their downregulation by ATP, expression of defense proteins increased remarkably when eATP signaling was blocked by AMP-PCP. These changes in the expression levels of defense proteins led to a corresponding increase in the resistance of tobacco leaf cells to pathogens after AMP-PCP treatment and a corresponding decrease after ATP treatment. In a 2011 study, Chivasa et al. (2011) pretreated suspension-cultured Arabidopsis cells with FB1 to deplete endogenous eATP, then added exogenous ATP into culture media and analyzed changes in protein levels in cells. This analysis led to identification of 26 proteins controlled by eATP, including molecular chaperones, cellular redox enzymes, glycolytic enzymes, and cellular protein degradation machinery components. Although the mechanism of action of FB1 is unclear, FB1 treatment led to significant downregulation of 8 ATP synthase β-subunits, while subsequent ATP treatment restored the expression level of these proteins. As mentioned above, FB1 caused cell death of Arabidopsis leaves, but addition of ATP rescued cells from FB1 injury. In ATP synthase β-subunit knockout mutants, FB1-induced cell death was effectively blocked, indicating ATP synthase β-subunit might be a target of FB1. Based on these data, they suggest that FB1 inhibits ATP synthase by suppressing its expression or by directly binding and inhibiting its activity. The consequent disruption of oxidative phosphorylation may be the main reason for FB1-induced cell death. Exogenous ATP restored the expression of these proteins, rescued cellular energy metabolism, and increased cell viability. These findings may explain why eATP-treated cells are resistant to FB1 treatment.
Lim et al. (2014) reported that decreased apyrase expression led to an increase in eATP levels and suppression of plant cell growth rate. In apyrase knockdown lines, upregulation of genes related to systemic acquired resistance, defense response to fungus or bacteria, regulation of H2O2 metabolism, regulation of hypersensitive response, and some signal transducer genes (e.g., MAP kinase) was observed, and downregulation of genes related to metal ion and nitrate transport, root hair differentiation, UDP-glycosyltransferase, and sterol biosynthetic process was observed. Further study is necessary to verify the relationship between changes in gene expression and physiological functions, such as decreased growth rate of root cells and increased resistance to biotic attack.
4.4 Crosstalk Between eATP and Plant Hormones
Plant hormones, including auxin, ethylene, and salicylic acid, are involved in eATP-regulated physiological functions. eATP may regulate some physiological processes through crosstalk with these hormones.
Auxin is involved in eATP-regulated plant cell growth and development. Tang et al. (2003) reported that eATP inhibited root elongation and disturbed root gravitropic growth of Arabidopsis. In addition, ATP inhibited basipetal auxin transport in Arabidopsis and maize roots, resulting in auxin accumulation in root tip cells. Furthermore, inhibition of apyrase activity significantly changed auxin level and distribution by suppressing auxin polar transport in root and hypocotyl of Arabidopsis. This led to abnormal growth behavior of these organs. In root tip of a null mutant of apyrase, asymmetric auxin distribution induced by gravity stimulation was impaired, and the root tip did not show gravitropic growth (Liu et al. 2012).
Ethylene, a key component in stress responses, is also involved in cellular responses to eATP. Wound, eATP, and osmotic stress promoted expression of ACS6, which is a key enzyme for biosynthesis of the ethylene precursor ACC (1-aminocyclopropane-1-carboxylic acid). ATP also induced expression of several ERFs. These proteins are well-known participants in biotic or abiotic stress-induced plant defense responses. Based on these findings, it was thought that ethylene synthesis and ethylene-stimulated signal transduction could be involved in eATP-induced downstream reactions (Jeter et al. 2004). Consistent with this idea, ATP stimulated ethylene accumulation in cultured ovule cells. ACC strengthened cotton fiber growth stimulated by low doses of eATP, while the ethylene antagonist aminovinylglycine strongly blocked this effect (Clark et al. 2010).
Salicylic acid (SA) is also involved in eATP-induced intracellular responses. SA effectively depleted eATP and induced pathogen resistance responses in Arabidopsis. Exogenous ATP decreased intracellular SA level and suppressed pathogen resistance (Chivasa et al. 2009). These findings suggest that eATP may suppress pathogenesis-related responses by inhibiting intracellular SA signaling.
5 Conclusions and Perspectives
Based on current research findings, it can be concluded that ATP exists in the apoplast of plants, where it functions as an essential extracellular messenger to promote regulation of cell metabolism, organ growth and development, and plant resistance to biotic and abiotic stresses. The downstream signal transduction pathways mediated by eATP have been intensively studied in the hopes that they would reveal insights into the mechanisms of eATP-regulated physiological functions. So far, our knowledge of eATP is still insufficient to clearly understand its novel role as a signaling molecule. Future studies will need to undertake the difficult task of verifying the physiological significance of eATP. We need more research to answer the remaining questions of what underlies the purpose of ATP secretion in plant cells, and why ATP, rather than other metabolites, is secreted as a messenger. To clarify the mechanisms of eATP-regulated physiological responses, the characteristics of eATP receptors and transmembrane signal transducers need to be determined. We need to contribute to current knowledge about eATP-induced intracellular messengers by identifying their targets and downstream biochemical reactions in eATP-regulated physiological responses. We are also in urgent need of determining the reasons for why low and high eATP concentrations lead to the same physiological reactions. To accomplish these tasks, creative ideas and novel research methods are urgently required.
References
Bodin P, Burnstock G (2001) Purinergic signalling: ATP release. Neurochem Res 26:959–969
Bouwmeester K, Han M, Blanco-Portales R, Song W, Weide R, Guo L, van der Vossen E, Govers F (2013) The Arabidopsis lectin receptor kinase LecRK-1.9 enhances resistance to Phytophthora infestans in Solanaceous plants. Plant Biotechnol J 12:10–16
Burnstock G (2006) Historical review: ATP as a neurotransmitter. Trends Pharmacol Sci 27:166–176
Burnstock G (2007) Purine and pyrimidine receptors. Cell Mol Life Sci 64:1471–1483
Burnstock G, Arnett T, Orriss I (2013) Purinergic signalling in the musculoskeletal system. Purinergic Signal 9:541–572
Cao Y, Tanaka K, Nguyen C, Stacey G (2014) Extracellular ATP is a central signaling molecule in plant stress responses. Curr Opin Plant Biol 20:82–87
Cavaliere F, Donno C, Ambrosi N (2015) Purinergic signaling: a common pathway for neural and mesenchymal stem cell maintenance and differentiation. Front Cell Neurosci 9:211
Chivasa S, Slabas AR (2012) Plant extracellular ATP signalling: new insight from proteomics. Mol Biosyst 8:445–452
Chivasa S, Ndimba B, Simon W, Robertson D, Yu X, Knox J, Bolwell P, Slabas A (2002) Proteomic analysis of the Arabidopsis thaliana cell wall. Electrophoresis 23:1754–1765
Chivasa S, Ndimba BK, Simon WJ, Lindsey K, Slabas A (2005) Extracellular ATP functions as an endogenous external metabolite regulating plant cell viability. Plant Cell 17:3019–3034
Chivasa S, Simon WJ, Murphy AM, Lindsey K, Carr J, Slabas A (2010) The effects of extracellular adenosine 5′-triphosphate on the tobacco proteome. Proteomics 10:235–244
Chivasa S, Tome D, Hamilton J, Slabas A (2011) Proteomic analysis of extracellular ATP-regulated proteins identifies ATP synthase beta-subunit as a novel plant cell death regulator. Mol Cell Proteomics 10:M110.003905
Chivasa S, Murphy A, Hamilton J, Lindsey K, Carr J, Slabas A (2009) Extracellular ATP is a regulator of pathogen defence in plants. Plant J 60:436–448
Choi J, Tanaka K, Cao Y, Qi Y, Qiu J, Liang Y, Lee S, Stacey G (2014a) Identification of a plant receptor for extracellular ATP. Science 343:290–294
Choi J, Tanaka K, Liang Y, Cao Y, Lee S, Stacey G (2014b) Extracellular ATP, a danger signal, is recognized by DORN1 in Arabidopsis. Biochem J 463:429–437
Clark G, Roux SJ (2009) Extracellular nucleotides: ancient signaling molecules. Plant Sci 177:239–244
Clark G, Roux S (2011) Apyrases, extracellular ATP and the regulation of growth. Curr Opin Plant Biol 14:700–706
Clark G, Torres J, Finlayson S, Guan X, Handley LJ, Kays J, Chen Z, Roux S (2010) Apyrase (NTPDase) and extracellular nucleotides regulate cotton fiber elongation in cultured ovules. Plant Physiol 152:1073–1083
Clark G, Fraley D, Steinebrunner I, Cervantes A, Onyirimba J, Liu A, Torres J, Tang W, Kim J, Roux S (2011) Extracellular nucleotides and apyrases regulate stomatal aperture in Arabidopsis. Plant Physiol 156:1740–1753
Crouzet J, Trombik T, Fraysse AS, Boutry M (2006) Organization and function of the plant pleiotropic drug resistance ABC transporter family. FEBS Lett 580:1123–1130
Dark A, Demidchik V, Richards S, Shabala S, Davies J (2011) Release of extracellular purines from plant roots and effect on ion fluxes. Plant Signal Behav 6:1855–1857
Day R, McAlvin C, Loh J, Denny R, Wood T, Young N, Stacey G (2000) Differential expression of two soybean apyrases, one of which is an early nodulin. Mol Plant Microbe Interact 13:1053–1070
Demidchik V, Nichols C, Oliynyk M, Dark A, Glover B, Davies J (2003) Is ATP a signalling agent in plants? Plant Physiol 133:456–461
Demidchik V, Shang Z, Shin R, Thompson E, Rubio L, Laohavisit A, Mortimer J, Chivasa S, Slabas A, Glover B, Schachtman D, Shabala S, Davies J (2009) Plant extracellular ATP signaling by plasma membrane NADPH oxidase and Ca2+ channels. Plant J 58:903–913
Deng S, Sun J, Zhao R, Ding M, Zhang Y, Sun Y, Wang W, Tan Y, Liu D, Ma X, Hou P, Wang M, Lu C, Shen X, Chen S (2015) Populus euphratica APYRASE2 enhances cold tolerance by modulating vesicular trafficking and extracellular ATP in Arabidopsis plants. Plant Physiol 169:530–548
Drury AN, Szent-Györgyi A (1929) The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart. J Physiol 68:213–237
Etzler M, Kalsi G, Ewing N, Roberts N, Day R, Murphy J (1999) A nod factor binding lectin with apyrase activity from legume roots. Proc Natl Acad Sci U S A 96:5856–5861
Ferrari D, Vitiello L, Idzko M, la Sala A (2015) Purinergic signaling in atherosclerosis. Trends Mol Med 21:184–192
Foresi NP, Laxalt AM, Tonón CV, Casalongué C, Lamattina L (2007) Extracellular ATP induces nitric oxide production in tomato cell suspensions. Plant Physiol 145:589–592
Fountain S, Cao L, Yound M, North R (2008) Permeation properties of a P2X receptor in the green algae Ostreococcus tauri. J Biol Chem 283:15122–15126
Hao L, Wang W, Chen C, Wang Y, Liu T, Li X, Shang Z (2012) Extracellular ATP promotes stomatal opening of Arabidopsis thaliana through heterotrimeric G protein α subunit and reactive oxygen species. Mol Plant 5:852–864
Idzko M, Ferrari D, Riegel A, Eltzschig H (2014) Extracellular nucleotide and nucleoside signaling in vascular and blood disease. Blood 124:1029–1037
Jaffe M (1973) The role of ATP in mechanically stimulated rapid closure of the venus’s-flytrap. Plant Physiol 51:17–18
Jeter C, Tang W, Henaff E, Butterfield T, Roux S (2004) Evidence of a novel cell signalling role for extracellular adenosine triphosphates and diphosphates in Arabidopsis. Plant Cell 16:2652–2664
Kennedy C (2015) ATP as a cotransmitter in the autonomic nervous system. Auton Neurosci 191:2–15
Kim SY, Sivaguru M, Stacey G (2006) Extracellular ATP in plants. Visualization, localization, and analysis of physiological significance in growth and signaling. Plant Physiol 142:984–992
Kim S, Yang S, Kim T, Han J, Suh J (2009) Hypertonic stress increased extracellular ATP levels and the expression of stress responsive genes in Arabidopsis thaliana seedlings. Biosci Biotechnol Biochem 73:1252–1256
Lew RR, Dearnaley J (2000) Extracellular nucleotide effects on electrical properties of growing Arabidopsis thaliana root hairs. Plant Sci 153:1–6
Lim M, Wu J, Yao J, Gallardo I, Dugger J, Webb L, Huang J, Salmi M, Song J, Clark G, Roux S (2014) Apyrase suppression raises extracellular ATP levels and induces gene expression and cell wall changes characteristic of stress responses. Plant Physiol 164:2054–2067
Liu X, Wu J, Clark G, Lundy S, Lim M, Arnold D, Chan J, Tang W, Muday G, Gardner G, Roux S (2012) Role for apyrases in polar auxin transport in Arabidopsis. Plant Physiol 160:1985–1995
Lüttge U, Schöch EV, Ball E (1974) Can externally applied ATP supply energy to active ion uptake mechanisms of intact plant cells? Aust J Plant Physiol 1:211–220
McAlvin C, Stacey G (2005) Transgenic expression of the soybean apyrase in Lotus japonicus enhances nodulation. Plant Physiol 137:1456–1462
Nejidat A, Itai C, Roth-Bejerano N (1983) Stomatal response to ATP mediated by phytochrome. Physiol Plant 57:367–370
Raghavendra A (1981) Energy supply for stomatal opening in epidermal strips of Commelina benghalensis. Plant Physiol 67:385–387
Ralevic V, Burnstock G (1998) Receptors for purines and pyrimidines. Pharmacol Rev 50:415–492
Reichler S, Torres J, Rivera AL, Cintolesi V, Clark G, Roux S (2009) Intersection of two signaling pathways: extracellular nucleotides regulate pollen germination and pollen tube growth via nitric oxide. J Exp Bot 60:2129–2138
Riewe D, Grosman L, Fernie A, Wucke C, Geigenberger P (2008) The potato-specific apyrase is apoplastically localized and has influence on gene expression, growth, and development. Plant Physiol 147:1092–1109
Roux SJ, Steinebrunner I (2007) Extracellular ATP: an unexpected role as a signaler in plants. Trends Plant Sci 12:522–527
Schiller M, Massalski C, Kurth T, Steinebrunner I (2012) The Arabidopsis apyrase AtAPY1 is localized in the Golgi instead of the extracellular space. BMC Plant Biol 12:123
Song C, Steinebrunner I, Wang X, Stout S, Roux S (2006) Extracellular ATP induces the accumulation of superoxide via NADPH oxidases in Arabidopsis. Plant Physiol 140:1222–1232
Steinebrunner I, Wu J, Sun Y, Corbett A, Roux S (2003) Disruption of apyrases inhibits pollen germination in Arabidopsis. Plant Physiol 131:1638–1647
Sun J, Zhang C, Deng S, Lu C, Shen X, Zhou X, Zheng X, Hu Z, Chen S (2012a) An ATP signalling pathway in plant cells: extracellular ATP triggers programmed cell death in Populus euphraticapce. Plant Cell Environ 35:893–916
Sun J, Zhang X, Deng S, Zhang C, Wang M, Ding M, Zhao R, Shen X, Zhou X, Lu C, Chen S (2012b) Extracellular ATP signaling is mediated by H2O2 and cytosolic Ca2+ in the salt response of Populus euphratica cells. PLoS One 12, e53136
Tanaka K, Gilroy S, Jones AM, Stacey G (2010a) Extracellular ATP signaling in plants. Trends Cell Biol 20:601–608
Tanaka K, Swanson S, Gilroy S, Stacey G (2010b) Extracellular nucleotides elicit cytosolic free calcium oscillations in Arabidopsis. Plant Physiol 154:705–719
Tanaka K, Choi J, Cao Y, Gary S (2014) Extracellular ATP acts as a damage-associated molecular pattern (DAMP) signal in plants. Front Plant Sci 5, Article 446
Tang W, Brady S, Sun Y, Muday G, Roux S (2003) Extracellular ATP inhibits root gravitropism at concentrations that inhibit polar auxin transport. Plant Physiol 131:147–154
Thomas C, Sun Y, Naus K, Lloyd A, Roux S (1999) Apyrase functions in plant phosphate nutrition and mobilizes phosphate from extracellular ATP. Plant Physiol 119:543–551
Thomas C, Rajagopal A, Windosr B, Dudler R, Lloyd A, Roux S (2000) A role for ectophosphatase in xenobiotic resistance. Plant Cell 12:519–533
Tonón C, Terrile MC, Iglesias MJ, Lamattina L, Casalongué C (2010) Extracellular ATP, nitric oxide and superoxide act coordinately to regulate hypocotyl growth in etiolated Arabidopsis seedlings. J Plant Physiol 167:540–546
Udvardy J, Farkas GL (1973) ATP stimulates the formation of nucleases in excised Avena leaves. Z Pflanzenphysiol 69:394–401
Vanegas D, Clark G, Cannon A, Roux S, Chaturvedi P, McLamore E (2015) A self-referencing biosensor for real-time monitoring of physiological ATP transport in plant systems. Biosens Bioelectron 74:37–44
Wang F, Jia J, Wang Y, Wang W, Chen Y, Liu T, Shang Z (2014) Hyperpolization-activated Ca2+ channels in guard cell plasma membrane are involved in extracellular ATP-promoted stomatal opening in Vicia faba. J Plant Physiol 171:1241–1247
Weerasinghe R, Swanson S, Okada S, Garrett M, Kim S, Stacey G, Boucher R, Gilroy S, Jones A (2009) Touch induces ATP release in Arabidopsis roots that is modulated by the heterotrimeric G-protein complex. FEBS Lett 583:2521–2526
Windsor B, Roux SJ, Lloyd A (2003) Multiherbicide tolerance conferred by AtPgp1 and apyrase in Arabidopsis: a novel mechanism of herbicide resistance. Nat Biotechnol 21:428–433
Wolf C, Hennig M, Romanovicz D, Steinebrunner I (2007) Developmental defects and seedling lethality in apyrase AtAPY1 and AtAPY2 double knockout mutants. Plant Mol Biol 64:657–672
Wu S, Wu J (2008) Extracellular ATP-induced NO production and its dependence on membrane Ca2+ flux in Salvia miltiorrhiza hairy roots. J Exp Bot 59:4007–4016
Wu J, Steinebrunner I, Sun Y, Butterfield T, Torres J, Arnold D, Gonzalez A, Jacob F, Reichler S, Roux S (2007) Apyrases (NTPDases) play key role in growth control in Arabidopsis. Plant Physiol 144:961–975
Wu S, Liu Y, Wu J (2008) The signaling role of extracellular ATP and its dependence on Ca2+ flux in elicitation of Salvia miltiorrhiza hairy root cultures. Plant Cell Physiol 49:617–624
Acknowledgement
We thank National Natural Science Foundation of China (Grant No. 31370319) and the Natural Science Foundation of Hebei Province (Grant No. C2014205079) for supporting this work.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Liu, T., Shang, Z. (2016). Extracellular ATP: An Essential Apoplastic Messenger in Plants. In: Cánovas, F., Lüttge, U., Matyssek, R. (eds) Progress in Botany Vol. 78. Progress in Botany, vol 78. Springer, Cham. https://doi.org/10.1007/124_2016_9
Download citation
DOI: https://doi.org/10.1007/124_2016_9
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-49489-0
Online ISBN: 978-3-319-49490-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)