
Abstract
Background
Green mold disease on citrus caused by Penicillium digitatum is the most serious and destructive disease. It is causing 90% of production losses during post-harvest handling.
Results
In this study, the activity of seven yeast isolates from lemons against P. digitatum, a fungal pathogen that causes the green mold disease in lemons, was isolated and examined. In vitro experiments showed that isolate three significantly reduced pathogen growths and were later identified as Schwanniomyces vanrijiae. In addition, 3% ethanolic extracts of propolis (EEP) caused a strong mycelial growth inhibition with inhibition halos of 1.4 cm. The use of S. vanrijiae treatments to protect lemon fruits from green mold has been reported (55%); however, reports describing the application of EEP are limited (40%). Thus, the effectiveness of the combination of S. vanrijiae and 3% EEP in an antagonistic mixture for protecting lemon fruits from P. digitatum was examined. EEP and S. vanrijiae treatments were applied alone and in combination in both in vitro and in vivo conditions. The combined application of 3% EEP + S. vanrijiae on lemon fruits significantly reduced the severity and incidence of green mold (80 and 93.7%, respectively) with much higher efficacy than either treatment alone. Lemon fruits treated with both S. vanrijiae and 3% EEP showed increased levels of antioxidants, peroxidase (POD), polyphenol oxidase (PPO), and phenol than the untreated control.
Conclusion
The results indicated that the combination of S. vanrijiae + 3% EEP can strongly protect lemon fruits from green mold compared with the sole application of either bioagent.
Similar content being viewed by others


Background
Citrus fruits are one of the most economically important fruits in Saudi Arabia and have additional worldwide importance, being a major source of vitamin C and carotenoids. The most serious postharvest diseases of lemons are the blue and green mold caused by Penicillium sp. (Wang et al. 2018), with the green mold disease caused by Penicillium digitatum being the most serious and destructive (Eckert and Ogawa 1988). It is responsible for 90% production losses during postharvest handling (Bagy et al. 2020). P. digitatum enters mature citrus fruits through wounds; this results in the appearance of white fungal mycelia exhibiting green-colored growths. This disease then spreads throughout the fruit, resulting in changes in color, texture, and taste, and the development of a large olive-green surface surrounded by mycelial growth. Synthetic fungicides are the most common method for controlling these types of plant diseases due to their speed and efficacy (Palou et al. 2008). However, post-harvest disease control with synthetic fungicides has resulted in the evolution of resistant strains, environmental pollution, and harmful effects on human health. Therefore, other methods for disease control besides synthetic fungicides must be developed.
The best current alternative to synthetic fungicides is bioagents application (Parafati et al. 2015). Yeast antagonists, including Pichia guilliermondii, Clavispora lusitaniae, and Cryptococcus laurentii (Zhang et al. 2020), have been shown to repress the growth of P. digitatum and P. italicum that affect citrus fruits. However, the biocontrol performance of antagonistic microorganisms is less effective than conventional fungicides under semi commercial circumstances (Liu et al. 2013). The efficacy of such antagonists in controlling postharvest diseases should thus be enhanced (Mahunu et al. 2016).
Propolis is a natural product from honeybees that exhibits multiple biological activities and has recently been described as a health supplement suitable for consumers. Water and ethanolic extracts of propolis (EEP) showed antifungal, antibacterial, and antioxidant behavior (Abo-Elyousr et al. 2017). The biological activities exhibited by and chemical compounds contained in propolis depend on the collection time, geographical distribution, and nearby plant species (Hegazi et al. 2014). Propolis is composed of approximately 10% essential oils, 30% wax, 50% resin, 5% pollen, and 5% of other organic compounds (Falcao et al. 2010). Propolis has been used to delay fugal decay in different fruits and extend lifetimes of stored fruits (Ozdemir et al. 2010). In addition, 3% EEP was used to reduce the decay of oranges caused by Penicillium spp. (El-Badawy et al. 2012), and 10% EEP completely repressed Penicillium spp. mycelial growth on potato dextrose agar (PDA) (Ayhan et al. 2013).
The study on the combined application of yeast + propolis bioagents to control the green mold disease of lemon has not been conducted before. Thus, this study was undertaken to examine the in vitro antagonistic activity of S. vanrijiae and EEP alone and in combination against P. digitatum, the pathogen responsible for green mold of lemon fruits. Also, in vivo studies to test the efficacy of the combination on P. digitatum in lemon fruits were performed.
Methods
Isolation and characterization of endophytic yeasts and green mold
Samples of healthy and infected lemon fruits that showed typical symptoms of the green mold disease were collected from a storage house located in Jeddah, Saudi Arabia, and seven isolates of endophytic yeasts and P. digitatum were obtained. Green mold pathogens were morphologically identified based on mycelial growth, colony color, and spore structure as described by Kurtzman et al. (2000), with identities confirmed by ITS region sequencing.
In vitro inhibition of P. digitatum by yeast
The in vitro inhibition of P. digitatum by seven yeast isolates was tested. A loop containing 24-h-old yeast was horizontally streaked near the edges of PDA; dextrose, 20 g; agar, 15 g; potato, 200 g) plates. A 5-mm mycelial plug of a 3-day-old P. digitatum colony grown on PDA was placed upside down on each streaked plates 3 cm from each streak. PDA plates containing only P. digitatum plugs were used as controls. Plates were incubated at 25 °C for 5 days, and the reduction in P. digitatum growth was calculated using the following formula:
Where I% is the percent mycelial growth inhibition, A represents the growth diameter of the control, and B represents the growth diameter (cm) of the pathogen for each treatments. Each experiment was performed twice with four replicates per experiment. The yeast isolates that most significantly reduced P. digitatum mycelial growth were selected for additional experiments.
Preparation of EEP
EEP were prepared by grinding 200 g of frozen propolis that was placed for 3 h prior to grinding. For grinding, 69% ethanol was used with an extraction ratio of 3:1 ethanol/propolis, and mixtures were shaken for 2 days at 150 rpm. Extracts were centrifuged at 5000 rpm for 10 min and filtered using filter paper. The resulting supernatant was collected and kept at room temperature for 3days to allow for evaporation, and the remaining resin was collected for future use. To prepare 1, 2, and 3% EEP solutions, equivalent weights of propolis were dissolved in the required volume of 70% ethanol. Then, EEP solutions were incubated in the refrigerator for further use Abdel-Rahim and Abo-Elyousr (2017).
In vitro inhibitory activity of EEP
To evaluate the in vitro inhibitory activity of EEP, conidia were collected from 1-week-old P. digitatum cultures grown on PDA plates incubated at 25 °C. Briefly, spore suspensions were prepared by addling distilled water to growing colony plates to collect spores, the concentration of each spore suspension was adjusted to 104 spore/mL, and 1 mL spore suspension was spread on the surface of a fresh PDA plate. Next, 0.5-cm-diameter wells were prepared in each plate in which 50 μL of 1%, 2%, or 3% EEP were poured, with 100 μL SDW plus 3% ethanol poured into wells of the control plates. Plates were then incubated at 25 °C for 5 days in the dark, and the inhibition zone (cm) around each well was measured. Four replicates for each treatment were used, and the experiment was conducted twice.
Molecular identification of bioagents
The selected bioagents were identified by PCR analysis of their ITS regions, except for P. digitatum, was identified based on morphology. For PCR amplification, DNA was extracted from 24-h-old yeast cultures using a DNeasy plant extraction kit (Qiagen, CA, USA) according to the manufacturer’s instructions. The primer pair ITS1 F (GCATCGATGAAGAACGCAGC) and ITS4 R (TCC TCC GCT TAT TGA TATGC) was used to amplify partial ITS gene sequences (White et al. 1990). Standard PCR was performed with reaction mixtures containing 20 μL DNA template, 4 μL 10 mM dNTPs, 5 μL 5× PCR buffer, 2.5 mM MgCl2, 2.5 U Taq polymerase (Promega), 25 pmol of each primer, and sterilized distilled water up to a final volume of 50 μL. Reactions were performed in a thermal cycler using the following conditions: initial denaturation at 94 °C for 3 min, followed by 35 cycles consisting of 94 °C for 60 s, 55 °C for 60 s, and 72 °C for 60 s, followed by a final extension of 72 °C for 7 min. PCR products (10 μL aliquots) were then analyzed on 1% agarose gels run for 75 min in TAE buffer (Tris, 40 mM; sodium acetate, 20 mM; EDTA, 1 mM; pH 7.2), with bands detected using a UV illuminator. Ribosomal ITS PCR products were identified based on their relative amounts of electrophoretic migration, then eluted from the gel, and purified using a purification kit (SolGent, Daejeon, South Korea), following the manufacturer’s instructions. PCR products were confirmed by sequencing (SolGent). Partial sequences of PCR products were compared with the whole yeast genome sequence obtained from NCBI, and the isolated sequences were 100% identical to the sequence obtained from NCBI.
In vitro effects of S. vanrijiae PHYTSV1 and EEP
In vitro inhibitory activity
To examine the in vitro inhibition of P. digitatum by EEP and S. vanrijiae, spore suspensions of P. digitatum were prepared from 3-day-old colonies by adding 5 mL distilled water to Petri plates containing 104 spores/mL. The resulting spore suspension (200μL) was then spread over the surface of a PDA plate. After the suspension had been absorbed by the plate, 0.5-cm punches were made into the agar and then filled with a 50-μL mixture of S. vanrijiae PHYTSV1 (1 × 109 CFU mL−1) and 3% EEP (1:1). Next, plates were incubated at 27 °C for 5 days, and the reduction in mycelial growth (I%) was calculated. Four replicates were used per experiment, and each experiment was conducted twice.
Effects on spore germination
Effects of EEP and S. vanrijiae PHYTSV1 on spore germination were determined. Glass tubes were filled by potato dextrose broth (PDB), and 100 μL P. digitatum conidial suspension (1 × 106 conidia/mL) was transferred to tubes containing S. vanrijiae PHYTSV1, EEP, or both. Control treatment tubes were also prepared with only PDB. Tubes were then incubated at 25 °C on a rotary shaker at 100 rpm for 15 h. To observe the germination rate for each treatment, at least 200 spores per replicate were examined under a microscope, and spore germination was determined using the criteria described by Elsherbiny et al. (2021). Germination inhibition of spores was calculated using the equation presented above. Three replicates were performed for each condition, and the experiment was conducted twice.
In vivo activity of S. vanrijiae PHYTSV1 and EEP
Preparation of pathogen and antagonist inoculum
To prepare the inoculum, P. digitatum was transferred onto PDA plates and incubated at 25 °C for 7 days. After germination, 10 mL distilled water was added to each plate, and spores were scraped into solution using a sterilized bacterial rod. The spore suspension was then filtered through a layer of sterile muslin cloth. Using a hemocytometer, the concentration of the conidial suspension was counted and adjusted to 24 × 106 CFU/mL (Abdel-Rahim and Abo-Elyousr 2018). Erlenmeyer flasks were filled with 50 mL yeast malt broth medium, inoculated with S. vanrijiae PHYTSV1, and then incubated at 25 °C with shaking at 200×g for 48 h. Tubes containing the resulting broth culture were then centrifuged at 6000 rpm for 10 min. The density of the re-suspended cells was then adjusted to 2 × 107 cells/mL using a hemocytometer.
Effects of S. vanrijiae and EEP on stored lemons
Effects of S. vanrijiae PHYTSV1 and EEP on lemon fruits (Citrus limon L. Burm) were examined using protocols described by Sallam et al. (2012) with minor modifications. Briefly, healthy lemons of the seedless lime variety were purchased, and the surface was sterilized by dipping the fruits in 2% NaClO for 2 min. After surface sterilization, fruits were rinsed 3 times with sterilized distilled water and then placed on sterile filter paper to dry. Next, 2-mm-deep and 5-mm-wide wounds were made on the surface of the peel of each fruit in opposing directions. Each wound was covered with 50 μL of S. vanrijiae (1 × 109 CFU/mL), 3% EEP, or a 1:1 v/v mixture of each. For control samples, 20 μL of sterile distilled water was added to each wound. Fruits were then allowed to sit at for 1 h, and then, 20 μL P. digitatum conidial suspension (104 CFU/mL) was injected into each fruit. The fruits were then incubated at 25 °C and 90% humidity for 7 days in (25 × 30 cm) plastic boxes. The resulting lesion diameter on each fruit was measured according to Madbouly et al. (2020), and the disease incidence and severity determined as follows:
Disease severity was evaluated using the following rating scale: 0 = no infection observed; 1 = a quarter of the fruit decayed; 2 = half the fruit decayed; 3 = three-quarters of the fruit decayed; and 4 = the entire fruit decayed.
Four replicates were used to perform each treatment with 20 lemons per treatment.
Effects of yeast and propolis on peroxidase (POD) and polyphenol oxidase (PPO) activities in lemon tissues
Enzyme extraction
To extract enzymes from lemons, 2 g of tissue from each fruit was collected. Tissues were placed in 20 mM Tris-HCl buffer (pH 7.2) and homogenized using a homogenizer. The homogenate was poured into glass tubes and centrifuged at 10,000 rpm at 4 °C for 10 min. Next, the supernatant was designated as crude extract and stored at − 20 °C for later determination of PPO and POD activities.
POD activity
POD activity was measured according to Bereika et al. (2020) with some modifications. Each prepared reaction mixture was 1 mL in total and contained 0.25 mL (0.2 M) sodium acetate (pH 5.5), 8 μL (0.97) M H2O2, 0.08 mL (0.5 M) guaiacol, and least amount of enzyme preparation. The change in absorbance at 470 nm reflecting guaiacol oxidation was then followed for 1 min using a spectrophotometer. Under standard assay conditions, the amount of enzyme that caused an increase in absorbance of at 1.0 O.D per min was defined as one unit of enzyme. Extraction buffer alone served as a blank reference sample. Three replicates were used for each treatment.
PPO activity
PPO activity was determined, following protocols from Batra and Kuhn (1975) using catechol as a substrate. A catechol solution (20 mM) was prepared using 0.01 M Na3PO4 (pH 6.8); then, 0.2 mL extract was added as quickly as possible to 2.8 mL catechol. Using a spectrophotometer, the change in absorbance at 400 nm was recorded for 2 min. An enzyme activity unit was defined as the amount of enzyme that caused an absorbance change of 0.1 in 1 min under standard assay conditions. Extraction buffer alone served as a blank reference. Three replicates were measured for each treatment.
Non-enzymatic assays
Sample preparation
For flavonoid and phenolic compound extraction, 1.0 g sample was suspended in 10 mL 70% ethanol (v/v). The suspension was then placed at 30 °C on a shaker at 120 rpm for 2 h and then centrifuged at 1013×g for 5 min. The resulting supernatant was used for further analysis.
Total phenol content
The total phenol content of each sample was measured by the method established by Malik and Singh (1980). Methanol extracts (50 μL) of each sample or standard (gallic acid) were mixed with 850 μL methanol, 100 μL Folin-Ciocalteu reagent was added, and samples were left at room temperature for 5 min. Next, samples were mixed with 500 μL 20% Na2CO3 and incubated at room temperature for 30 min to react. The absorbance of each sample at 750 nm was then measured, and the total phenol content was calculated based on a standard curve. The standard curve was prepared using gallic acid, and the phenol content in each extract was expressed as mg of gallic acid/g of fresh weight. Phenol content measurements were performed twice with three replicates.
Total flavonoid content
Using the modified colorimetric assay reported by Zhishen et al. (1999), flavonoid concentrations in each sample were determined. A 250-μL aliquot of standard (catechin) or methanol extract was mixed with 75 μL 5% NaNO2 and 1.25 mL sterile distilled water. The reaction mixture was left for 6 min and then mixed with 150 μL 10% AlCl3, 0.5 mL (1 M) NaOH, and 275 μL sterile distilled water was added to solution. The absorbance of each solution was measured at 510 nm, and the total flavonoid content was calculated using a standard curve. Known concentrations of catechin were used to generate a standard curve, and the results were described as mg of catechin equivalent/g.
Statistical analysis
Experimental data were analyzed using the statistical package Statistix (ver. 8.1), and two-way ANOVA was used. The least significant difference test at P = 0.05 was performed for disease severity and disease incidence means to identify meaningful differences between the means of results of various treatments. Data were expressed as mean ± SE.
Results
Isolation of endophytic yeasts and the causal pathogen of green mold of lemons
Fungal pathogens of green mold were isolated from infected lemons, while seven isolates of endophytic yeasts isolates were collected from healthy lemons.
In vitro antagonistic activity of yeast against P. digitatum
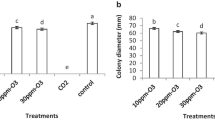
All isolates were able to inhibit the mycelial growth of P. digitatum pathogens with varying levels of efficacy. Isolate 3 reduced P. digitatum growth by a significantly higher level than other isolates, with mycelium growth reductions of over 75% than the control (Table 1). Thus, three isolates were selected for further experiments.
Molecular identification of bioagents and yeast species
Yeast isolate 3 was identified as S. vanrijiae based on BLAST searches of the NCBI Nucleotide Collection Database for similar ITS sequences. A phylogenetic tree was also assembled using ITS sequences from S. vanrijiae and close homologues (Fig. 1) by using MEGAX. The ITS sequences of the pathogenic and yeast isolates we obtained have been added to the GenBank database under accession no. MT523046.

Phylogenetic analysis of yeast isolate on ITS gene. This analysis was performed using the neighbor-joining method in BLAST pairwise alignments
In vitro inhibition of P. digitatum by EEP
The in vitro antimicrobial effect of 1, 2, and 3% EEP against P. digitatum was tested. The 3% EEP solution was the most effective one for inhibiting the mycelium growth of P. digitatum, with inhibition halos of 2.4 cm observed. Although lower, inhibitory activity was also observed for 1% and 2% EEP solutions, with inhibition halos of 1.3 cm and 1.4 cm, respectively.
In vitro inhibitory activity of S. vanrijiae PHYTSV1 and 3% EEP mixtures
The growth reduction of fungal pathogens was significantly higher when mixtures of S. vanrijiae and 3% EEP were used compared with the application of either component alone, with mixtures producing 3.3-cm inhibition halos.
Impact of the S. vanrijiae PHYTSV1 and 3% EEP mixture on spore germination
A mixture of S. vanrijiae and 3% EEP caused significantly higher inhibition of P. digitatum spore germination (P < 0.05) than other individual treatments. The combination of S. vanrijiae + 3% EEP reduced P. digitatum spore germination by 80.5%, while 3% EEP and S. vanrijiae caused 65 and 75% reductions in spore germination, respectively (Fig. 2).

Effect of S. vanrijiae, 3% EPE, and mixture on spore germination. Values in the column followed by different letters indicate significant differences among treatments according to lease significant differences test (P = 0.05)
In vivo activity of S. vanrijiae PHYTSV1 and 3% EEP
The activities of S. vanrijiae PHYTSV1 and 3% EEP were tested in vivo on lemon fruits. Artificial wounds were treated with S. vanrijiae PHYTSV1 and EEP either alone or as part of a mixture. After 1 h of incubation of wounds with S. vanrijiae PHYTSV1 and/or EEP, each wound was inoculated with a conidial suspension of P. digitatum. Results showed a significantly (P 0.05) greater reduction in disease incidence on fruits treated with S. vanrijiae PHYTSV1 and EEP mixture compared with fruits treated with either component individually. Furthermore, disease severity was also reduced in fruits receiving combined treatments than in untreated control (Table 2).
Effect of S. vanrijiae PHYTSV1 and EEP alone and in combination on levels of defense-related enzymes, phenolic compounds, and flavonoids in lemons
Effects on enzyme activity
During fruit storage, PO activity had significantly increased (P < 0.05) in all treated lemons compared with control lemons, 2 days post treatment. The maximum PO activity was reached 4 days post-treatment (Fig. 3). In addition, a similar pattern was observed for PPO activity, which began to decline 4 days post-treatment (Fig. 4).

Time course of peroxidase activity (U/min/g fw) in extracts from lemon cv seedless limes tissues treated or not with S. vanrijiae and 3% EPE in mixture. All the values are the means of four replicates ± SE

Time course of polyphenoloxidase activity (U/min/g fw) in extracts from lemon cv seedless limes tissues treated or not with S. vanrijiae and 3% EPE in mixture. All the values are the means of four replicates± SE
Total phenolic content
The total phenol content was determined at multiple times post treatment (Fig. 5). The three tested treatments increased the total phenol content at all times, except day 1. The total phenol content was highest in lemons treated with both S. vanrijiae PHYTSV1 and 3% EEP, followed by lemons treated with either control and with untreated lemons containing the lowest amounts. After 4 days post-treatment, total phenolic levels began to decline.

Time course of phenol content (phenols (mg/g fw) in extracts from lemon cv seedless limes tissues treated or not with S. vanrijiae and 3% EPE in mixture. All the values are the means of four replicates ± SE
Total flavonoid content
The total flavonoid content was measured up to 8 days post-treatment (Fig. 6). All treatments caused significant increases in the total flavonoid content than the controls. At 4 and 6 days post-treatment, flavonoid levels reached their maximum values, with flavonoid levels trending downward in lemons treated with a mixture of S. vanrijiae PHYTSV1 and EEP, EEP alone, and S. vanrijiae PHYTSV1 alone. After day 6, the flavonoid content began decreasing in all samples.

Time course of flavonoids (mg/g fw) in extracts from lemon cv seedless limes tissues treated or not with S. vanrijiae and 3% EPE in mixture. All the values are the means of four replicates ± SE
Discussion
In this study, seven yeast isolates from lemons had significant in vitro antifungal activity against P. digitatum, the causal pathogen of the green mold disease in citrus fruits. The antifungal potency of biocontrol agents likely arises due to the competition between antagonists and pathogens for nutrients, which can result in the production of antifungals, antibiotics, or mycoparasitism via cell wall hydrolysis (Madbouly et al. 2020). A previous study on the use of the yeast (P. guilliermondii) as treatment for green mold pathogens provides additional evidence that competition between pathogens and biocontrol antagonists for nutrients likely plays a key role in the biological control of P. digitatum (Winiewski et al. 1991). The production of metabolites by yeast may significantly affect the ability of other fungi to produce cell wall-degrading enzymes and may hydrolyze the conidia of competing fungi (Li et al. 2019).
In the present study, 3% EEP caused the highest reduction of mycelia growth of P. digitatum in vitro than 1 and 2% EEP, potentially due to the higher levels of antimicrobial and antioxidant activities of phenolic compounds resulting from increased EEP. Other researchers have reported a positive correlation between the antioxidant and antimicrobial characteristics of propolis and the polyphenols and flavonoid content of (Chaillou and Nazareno 2009). Moreover, previous work by Yang et al. (2009) showed that EEP contains more flavonoids and phenols than propolis extracts and exhibits more promising antifungal activity against P. italicum.
Spore germination and mycelial growth of P. digitatum was significantly reduced by S. vanrijiae and EEP. The discrepancy might be related to the levels of phenolic compounds in EEP or metabolites in S. vanrijiae (Agirman and Erten 2020).
Madbouly et al. (2020) reported that S. vanrijiae can be used to control disease in apple fruits under storage conditions. Using a separate metric for treatment success, Mattiuz et al. (2015) reported that propolis extracts can decrease the weight loss of citrus, apples, tomatoes, and cherries during storage. When fruits are treated with propolis extracts, their surfaces develop coatings of beeswax, which can minimize water loss, gas exchange, and microbial infestations; extend shelf life; and maintain tissue firmness. In addition, the postharvest treatment of citrus fruits with propolis extracts was found to protect against natural disease incidence. For example, treatment with 1000 mg/L propolis reduced fruit decay from 30 to 13% after 22 days of storage. Moreover, numerous additional studies have described the broad activity of propolis against yeast, bacteria, and fungi, including phytopathogenic fungi such as Colletotrichum gloeosporioides, Botrytis, and P. italicum.
PO has a key role in lignification and suberization of host cell walls, which restricts disease development (Bereika et al. 2020). PO also helps strengthen plant cell walls at attachment sites by promoting lignification or cross-linking of specific proteins (Abo-Elyousr et al. 2008). In the present experiments, PPO activity was higher in lemon fruits treated with yeast and EEP than in control fruits, with such activity increasing from 2 to 3 days after application.
In terms of total flavonoid and phenol content, this study demonstrated that a mixture of yeast and EEP caused a greater increase in phenol content and total flavonoids in lemons than application of either treatment on its own. These results are in agreement with previous studies that measured higher POD activity in apple fruits treated with Aureobasidium pullulans compared with control fruit (Youssef et al. 2020). S. vanrijiae and 3% EEP application either alone or in combination exhibited strong in vitro reduction of P. digitatum radial growth and in vivo reductions in disease development on lemon fruits. Moreover, these treatments showed promise in conidia germination inhibition. Furthermore, the enhancement of PO and PPO activities, as well as levels of flavonoids and phenolic compounds, may have a direct relationship with the process by which yeast and EEP induce resistance against green mold in lemon fruits. Intensification of natural host resistance significantly correlated with augmentation of antioxidant systems.
Availability of data and materials
Not applicable
Abbreviations
- EPE:
-
Ethanol propolis extracts
- POD:
-
Peroxidase activity
- PPO:
-
Polyphenol oxidase
- LSD:
-
Least significant difference
References
Abdel-Rahim IR, Abo-Elyousr KAM (2017) Using of endophytic Saccharomycopsis fibuligera and thyme oil for management of gray mold rot of guava fruits. Biol Cont 110:124–131. https://doi.org/10.1016/j.biocontrol.2017.04.014
Abdel-Rahim IR, Abo-Elyousr KAM (2018) Talaromyces pinophilus strain AUN-1 as a novel mycoparasite of Botrytis cinerea, the pathogen of onion scape and umbel blights. Microbiol Res 212-213:1–9. https://doi.org/10.1016/j.micres.2018.04.004
Abo-Elyousr KAM, Hussein MAM, Allam ADA, Hassan MHA (2008) Enhanced onion resistance against stemphylium leaf blight disease, caused by Stemphylium vesicarium, by di-potassium phosphate and benzothiadiazole treatments. Plant Pathol J 24:171–177. https://doi.org/10.5423/PPJ.2008.24.2.171
Abo-Elyousr KAM, Seleim MEA, El-Sharkawy RM, Bagy HMMK (2017) Effectiveness of Egyptian propolis on control of tomato bacterial wilt caused by Ralstonia solanacearum. J Plant Dise Protect 124(5):467–472. https://doi.org/10.1007/s41348-017-0120-x
Agirman B, Erten H (2020) Biocontrol ability and action mechanisms of Aureobasidium pullulans GE17 and Meyerozyma guilliermondii KL3 against Penicillium digitatum DSM2750 and Penicillium expansum DSM62841 causing postharvest diseases. Yeast 37(9-10):437–448. https://doi.org/10.1002/yea.3501
Ayhan T, Mumcu AS, Tuylu AO, Sorkun K, Salih B (2013) Antifungal activity of propolis samples collected from different geographical regions of turkey against two Food-related molds, Aspergillus versicolor and Penicillium aurantiogriseum. GIDA 38:135–142. https://doi.org/10.5505/gida.2013.10820
Bagy HMMK, Badawy FMI, Abou-Zaid E, A.A, Badawy SM, Sallam MAN (2020) Control of green mold disease using chitosan and its effect on orange properties during cold storage. Arch Phytopathol Plant Protect:1–16. https://doi.org/10.1080/03235408.2020.1847568
Batra G, Kuhn C (1975) Polyphenoloxidase and peroxidase activities associated with acquired resistance and its inhibition by 2-thiouracil in virus-infected soybean. Physiol Plant Pathol 5(3):239–248. https://doi.org/10.1016/0048-4059(75)90090-9
Bereika FFM, Sallam NMA, Alamri SAM, Abo-Elyousr KAM, Hashem M, Mostafa YS (2020) Approving the biocontrol strategy of potato wilt caused by Ralstonia solanacearum on field scale using Enterobacter cloacae PS14 and Trichoderma asperellum T34. Egypt J Biol Pest Cont 30:61. https://doi.org/10.1186/s41938-020-00262-9
Chaillou LL, Nazareno MA (2009) Bioactivity of propolis from Santiago del Estero, Argentina, related to their chemical composition. LWT Food Sci Technol 42(8):1422–1427. https://doi.org/10.1016/j.lwt.2009.03.002
Eckert JW, Ogawa JM (1988) The chemical control of postharvest diseases: deciduous fruits, berries, vegetables and root/tuber crops. Ann Revi Phytopathol 26(1):433–469. https://doi.org/10.1146/annurev.py.26.090188.002245
El-Badawy HE, Baiea MH, Eman AA (2012) Efficacy of propolis and wax coatings in improving fruit quality of Washington navel orange under cold storage. Res J Agric Biol Sci 8:420–428
Elsherbiny E, Dawood HD, Nesreen AS (2021) Antifungal action and induction of resistance by β-aminobutyric acid against Penicillium digitatum to control green mold in orange fruit. Pesticide Biochem Physiol. 171:104721. https://doi.org/10.1016/j.pestbp.2020.104721
Falcao SI, Vilas-Boas M, Estevinho LM, Barros C, Domingues MRM, Cardoso SM (2010) Phenolic characterization of Northeast Portuguese propolis: usual and unusual compounds. Anal Bioanal Chem 396(2):887–897. https://doi.org/10.1007/s00216-009-3232-8
Hegazi A, Abdou AM, Abd-Allah F (2014) Egyptian propolis 11: its antimicrobial activity with comparison with different localities. Int J Curr Microbiol App Sci 3:530–538
Kurtzman C, Fell JW, Boekhout T (2000) The yeasts: A taxonomic study. 14(3).
Li J, Li H, Ji S, Chen T, Tian S, Qin G (2019) Enhancement of biocontrol efficacy of Cryptococcus laurentii by cinnamic acid against Penicillium italicum in citrus fruit. Postharvest Biol Technol 149:42–49. https://doi.org/10.1016/j.postharvbio.2018.11.018
Liu J, Sui Y, Wisniewski M, Droby S, Liu Y (2013) Review: utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int J Food Microbiol 167(2):153–160. https://doi.org/10.1016/j.ijfoodmicro.2013.09.004
Madbouly A, Abo-Elyousr AM, Ismail MI (2020) Biocontrol of Monilinia fructigena the causal agent of brown rot of stored apple fruits using certain endophytic yeasts. Biol Cont 144C:104239. https://doi.org/10.1016/j.biocontrol.2020.104239
Mahunu GK, Hongyin Z, Qiya Y, Chaolan L, Xiangfeng Z (2016) Biological control of patulin by antagonistic yeast: a case study and possible model. Crit Rev Microbiol 42(4):643–655. https://doi.org/10.3109/1040841X.2015.1009823
Malik EP, Singh MB (1980) Plant Enzymology and Hittoenzymology, 1st edn. Kalyani Publishers, New Delhi, p 286
Mattiuz BH, Ducamp-Collin MN, Mattiuz CFM, Vigneault C, Marques KM, Sagoua W, Montet D (2015) Effect of propolis on postharvest control of anthracnose and qualityparameters of ‘Kent’ mango. Sci Hortic 184:160–168. https://doi.org/10.1016/j.scienta.2014.12.035
Ozdemir AE, Candir EE, Kaplankiran M, Soylu EM, Sahinler N, Gul A (2010) The effects of Ethanol-dissolved propolis on the storage of grapefruit cv. Star Ruby. Turk J Agric For 34:155–162. https://doi.org/10.3906/tar-0902-50
Palou L, Smilanick JL, Droby S (2008) Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Posthar Rev 4:1–16. https://doi.org/10.2212/spr.2008.2.2
Parafati L, Vitale A, Restuccia C, Cirvilleri G (2015) Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol 47:85–92. https://doi.org/10.1016/j.fm.2014.11.013
Sallam NMA, Badawy FM, Ibtesam A, Ibrahim R (2012) Biocontrol of green mold of orange using some yeasts strains and their effects on postharvest quality parameters. Int J Plant Pathol 3:14–24. https://doi.org/10.3923/ijpp.2012.14.24
Wang W, Liu S, Deng L, Ming J, Yao S, Zeng K (2018) Control of citrus post-harvest green molds, blue molds, and sour rot by the cecropin a-melittin hybrid peptide BP21. Front Microbiol 9:2455. https://doi.org/10.3389/fmicb.2018.02455
White TJ, Bruns T, Lee S, Tailor S (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innins MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols. A guide to methods and applications. Academic Press, Inc., San Diego, pp 315–322. https://doi.org/10.1016/B978-0-12-372180-8.50042-1
Winiewski M, Biles C, Droby S, McLaughlin R, Wilson C, Chalutz E (1991) Mode of action of the postharvest biocontrol yeast, Pichia guilliermondii: I. Characterization of attachment to Botrytis cinerea. Physiol Mol Plant Pathol 39:245–258. https://doi.org/10.1016/0885-5765(91)90033-E
Yang SZ, Peng LT, Pan SY, Yao XL, Gao YL (2009) Antifungal activity of ethyl acetate extract of propolis against Penicillium italicum and its stability. Food Sci 30:87–90
Youssef K, Roberto SR, Tiepo AN, Constantino LV, de Resende JTV, Abo-Elyousr KAM (2020) Salt solution treatments trigger antioxidant defense response against gray mold disease in table grapes. J Fungi 6(3):179. https://doi.org/10.3390/jof6030179
Zhang H, Zhenga X, Chengxin F, Xia Y (2020) Postharvest biological control of gray mold rot of pear with Cryptococcus laurentii. Postharvest Biol Technol 35:79–86. https://doi.org/10.1016/j.postharvbio.2004.03.011
Zhishen J, Mengcheng T, Jianming W (1999) The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem 64(4):555–559. https://doi.org/10.1016/S0308-8146(98)00102-2
Acknowledgements
This project was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah, Saudi Arabia under grant no (G:24-155-1441). The authors, therefore, acknowledge with thanks DSR for technical and financial support. Also, We would like to thank Muhammad Imran, PhD. at the Arid Land Agriculture Department, Faculty of Meteorology, Environment and Arid Land Agriculture, King Abdulaziz University, for his indispensable technical support.
Funding
This project was funded by the Deanship of Scientific Research (DSR) at King AbdulAziz University, Jeddah, Saudi Arabia under grant no (G:24-155-1441).
Author information
Authors and Affiliations
Contributions
All authors contributed equally in the manuscript. AKAM suggested the idea of the work and contributed to data curation and their validation as well as writing original draft. ADQ contributed to the formal analysis of the data. NMM contributed to the reviewing and editing the manuscript. All authors reviewed and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable. This manuscript is in accordance with the guide for authors available on the journal’s website. Also, this work has not been published previously and is approved by all authors and host authorities
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abo-Elyousr, K.A.M., Al-Qurashi, A.D. & Almasoudi, N.M. Evaluation of the synergy between Schwanniomyces vanrijiae and propolis in the control of Penicillium digitatum on lemons. Egypt J Biol Pest Control 31, 66 (2021). https://doi.org/10.1186/s41938-021-00415-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-021-00415-4