
Abstract
The genus Teucrium L. is a well-known medicinal plant belonging to Lamiaceae family and Ajugoideae subfamily. The present study aims to evaluate the taxonomic status of the Teucrium species using both morphological and flavonoid characteristics and to determine the flavonoid classes. A total of 64 accessions belonged to eight species, and five subspecies were collected from different Zagros regions, Iran. Twenty-nine quantitative and qualitative morphological characters were studied. Extraction of leaf flavonoid was accomplished using a rotary evaporator and MeOH 90%. Identification of the chemotaxonomic position and flavonoid class of Teucrium species was investigated by thin layer chromatography, column chromatography and High Performance Liquid Chromatography-Micromass Quattro micro Atmospheric Pressure Ionization Mass Spectrometer. All information was analyzed using NTSYS pc. 2, PAST v.3.18 and Cluster Vis v. 1.8.2. According to the findings, T. polium accessions significantly revealed the highest value of morphological and flavonoid variations. All subspecies of T. orientale were definitely separated, but some relationships were observed between its subspecies, including taylori and glabrescens. All members of two sections of Polium and Teucris were definitely recognized. A total of seven flavonoid classes were determined from which the highest amounts of abundance were attributed to flavones (18) and isoflavones (17). In conclusion, morphological and flavonoid markers were introduced as appropriate characteristics in the taxonomic relationships of Teucrium species. All the evidence of the present research was first described for Iran.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The genus Teucrium L. belongs to Lamiaceae family and Ajugoideae subfamily and is known as one of the large and complex genera (Rechinger 1982; Ecevit Gen et al. 2015). This genus is represented by 260 species throughout the world, and 19 species in Iran from which four endemic species were recognized for the country (Jamzad 2012; Salmaki et al. 2016). Its species are generally distributed in Europe, western Mediterranean regions, southeast Australia, northwest and south America, North Africa and temperate of Asia particularly in Irano-Turanian regions (Bukhari et al. 2015; Salmaki et al. 2016). Mediterranean regions are the main diversity centers for the genus Teucrium (Salmaki et al. 2016). The species grow as shrub, dwarf-shrub, biennial, perennial and annual forms in dry, open, sandy and rocky areas and hillsides (Rechinger 1982; Jaradat 2015; Ecevit Gen et al. 2015; Salmaki et al. 2016).
Teucrium species are traditionally used to treat different disorders. They have a rich source of chemical compounds with several and valuable biological activities and therapeutic effects, including antiseptic, antibacterial, anticancer, wound healing (Elmasri et al. 2015), hypoglycemic, hypolipidemic, hepatoprotective, antipyretic, anti-inflammatory, anti-ulcer, anti-tumor (El-Ashmawy 2018), anti-urogenital, anti-indigestion, antirheumatism (Sabet et al. 2013), antioxidant, anti-nociceptive and antitoxic (Stefkov et al. 2012). Almost its potent biological activities previously reported have been attributed to the presence of polyphenols involving radical-scavenging activities (Alwahsh et al. 2015).
From a taxonomical point of view, several classifications for the genus Teucrium were presented. However, its proposed classifications remain controversial. Bentham (1835) introduced nine sections (Teucrium Ging. ex Benth., Teucropsis Ging. ex Benth., Leucosceptrum (Sm.) Benth., Chamaedrys Benth., Polium Benth., Scordium (Mill.) Rchb., Pycnobotrys Benth., Scorodonia Benth., Stachyobotrys Benth.). Briquet (1897) adopted the Bentham’s classification, but considered two further sections (Spinularia Boiss., Isotriodon Boiss.). Rechinger (1982) classified Teucrium as seven sections (Polium, Chamaedrys, Isotriodon, Scordium, Scorodonia, Stachyobotrys, Teucris Benth.). Yuzepchuk (1954) considered five sections (Teucris, Stachyobotrys, Scordium, Chamaedrys, Polium) for this genus. Kastner (1989) also divided the genus Teucrium into five sections and 13 subsections. He also omitted the sections Polium, Spinularia and Pycnobotrys. Additionally, Navarro and El Oualidi (2000) proposed nine sections and four subsections (sub-sect. Polium, Simplicipilosa S. Puech., Rotundifolia Cohen ex Valdes Berm & Sanchez- Crespo, Pumilum Rivas- Mart.). However, Jamzad (2012) has not considered any classification for this genus. Ranjbar et al. (2017) taxonomically reviewed the section Teucrium. A few taxonomic changes were achieved in this section. According to different classifications, the taxonomy of Teucrium was disputed. Consequently, this genus has complexities mostly resulting from the presence of wide distribution, high morphological variability, different ploidy levels and natural hybridization (Salmaki et al. 2016).
Based on the morphological literature, different reports are currently available in Teucrium species. Bukhari et al. (2015) investigated some morphological characteristics of leaves and flowers in T. polium L. from Saudi Arabia. Pavlova and Vasileva (2010) studied different serpentine ecotypes of T. polium from Bulgaria. Navarro and El Oualidi (2000), Jurisic Grubesi et al. (2007), Eshratifar et al. (2011), Ecevit Genc et al. (2015) and Marzouk et al. (2017) provided further reports of micromorphological studies, including trichome, pollen and nutlet of Teucrium species in Mediterranean, Croatia, Iran, Turkey, and Lybia. The results provided by these data sources confirmed that these characters had taxonomical value at sectional and specific levels.
Morphological variations have been the subject of few researches. Some reports in T. polium characterized that seasonal periods and edaphic conditions affected the density, thickness and height of trichomes and the height of stem and leaf (Pavlova and Vasileva 2010; Bosabalidis 2013). On the contrary, Bukhari et al. (2015) indicated no correlations between environmental conditions and trichomes.
In the context of published works in chemical compounds, several researches into Teucrium species were presented. The major chemical constituents were ascribed to its species, including phenolic acids, polyphenols, flavonoids (Valant-Vetschera et al. 2003; Stefkov et al. 2012; Jurisic Grubesic et al. 2012; Mitreskia et al. 2014; Alwahsh et al. 2015; Boghrati et al. 2016; Venditti et al. 2017; Gecibesler et al. 2019), essential oils as monoterpenoids, diterpenoids, triterpenoids (Ahmad et al. 2014; Jaradat 2015; Elmasri et al. 2015; Semiz et al. 2016; Rahimi et al. 2019), iridoids, phenylethanol glycosides (Elmasri et al. 2015), alkaloids, anthraquinones, tannins, (Shah and Shah 2015), benzene ester (Ahmad et al. 2014), and β-sitosterol (Jurisic Grubesi et al. 2012).
From a chemotaxonomic point of view, there is limited evidence of flavonoid compounds used in the genus Teucrium. Some authentic reports have demonstrated the isolation of essential oils. Chemotaxonomic status of T. arundini L. (Kremer et al. 2012), T. scordium L. (Radulovic et al. 2010), T. pumilum Loefl. ex L. (Perez et al. 2000) and T. stocksianum Boiss. (Sonboli et al. 2013) were previously investigated using essential oils. Sonboli et al. (2013), Venditti et al. (2017) and Rahimi et al. (2019) specified the variation in chemical compounds in T. polium, T. hircanum L. and T. stocksianum as chemo type.
According to the literature survey, there are no reports of morphometric and chemotaxonomy of flavonoid compounds in Teucrium species in Iran. Moreover, owing to the presence of valuable genetic resources in the Zagros region, the present study focuses on the following aims: (1) comprehensive morphometric studies of quantitative and qualitative morphological characteristics, (2) investigation of the morphological variations at intraspecific levels, (3) chemotaxonomic study of flavonoid profiles at specific and intraspecific levels and 4) determination of the flavonoid classes of each species from the Zagros region. All data were first described for Iran.
2 Materials and Methods
2.1 Morphology Section
In this section, 63 accessions from eight species including five subspecies from section Polium (T. polium L., T. capitatum L., T. gnaphalodes L.’ Her), section Teucris (T. orientale subsp. orientale (recently named under T. orientale L.), T. orientale subsp. taylori (Boiss.) Rech.f., T. orientale subsp. glabrescens (Hausskn. ex Bornm.) Rech. f., T. orientale subsp. gloeotrichum Rech. f., T. oliverianum Ging. ex Benth., T. parviflorum Schreb.) and section Scordium (T. melissoides Boiss. ex Hausskn., T. scordium subsp. scordioides (Schreb.) Arcang.) were collected and characterized from the north, west, southwest, center and south of Zagros regions (Table 3; “Appendix”). All specimens were collected from various natural habitats from March to September from 2018 to 2019. The voucher specimens were deposited in the Herbarium of Shahrekord University. The identification process of all collected specimens was accomplished by Flora Iranica and Flora of Iran (Rechinger 1982; Jamzad 2012).
To study the morphometric, morphological variations and taxonomic status in Teucrium species, 11 quantitative and 18 qualitative morphological variables were selected and measured in each species as provided in Table 1. All morphological characteristics were measured using stereomicroscope Olympus SZX-ZB12. Data matrix was subjected to a cluster analysis followed by simple matching coefficient and UPGMA (Unweighted Pair Group Method with Arithmetic Mean) method with NTSYS Pc. 2.0 (Rohlf 2000). An analysis of variance (ANOVA) and Kruskal–Wallis test also assessed the quantitative and qualitative morphological variables among species by the IBM SPSS Statistics for Windows, version 20 (IBM Corp., Armonk, N.Y., USA) software.
2.2 Phytochemical and Chemotaxonomy Study
Extraction of flavonoids was accomplished using the procedure suggested by Rahman (2005). The total flavonoid of air-dried leaves (10 gr) from eleven taxa was isolated using 90% MeOH at 60 °C. The flavonoid extraction was accomplished using a rotary evaporator at 70 °C. Flavonoid purification was conducted by n-BuOH and consecutively screened through silica gel 60F 254 (16 mg, 70 ml H2O) thin layer chromatography (TLC; 5 µM, 20 × 20 cm). The chromatogram was treated in different solvent systems such as CHCl3–MeOH (60:40; 70:30; 80:20), CH2O2–CH3COOH–H2O (40:40:20; 30:30:40) and C4H8O2–MeOH–H2O (98:1.5:0.5) (Cakir et al. 2006; Venditti et al. 2017; Gecibesler et al. 2019). Spot detection with natural product reagents (H2SO4 5% in MeOH) was completed by ultraviolet-366 nm (Rahman 2005). The flavonoids were separated by column chromatography (50 × 4 cm), followed by Sephadex LH20 Sigma- Aldrich (Sephadex and MeOH 20% mixture) in 100 mL CHCl3–MeOH 60:40 to yield several fractions. Identification of flavonoid classes was achieved on ultraviolet spectrum (200–400 nm), MeOH solution and shift reagents such as AlCl3, AlCl3/HCl, NaOAc, NaOAc/H3BO3 and MeOH. All fractions acquired were analyzed using HPLC-Micromass Quattro micro API (Atmospheric Pressure Ionization) Mass Spectrometer to provide the mass to charge (m/z) value in each species. Chromatography condition was prepared at Atlantis T3-C18 column (3 µ, 2.1 × 100 mm, flow rate of 0.25 ml/min) at 30 °C. The mobile phase includes acetonitrile, methanol and formic acid (98%). Moreover, flavonoid standard from SIGMA Aldrich Co. includes apigenin with 98% purity (Hossain et al. 2010; Aghakhani et al. 2018).
To explore chemotaxonomy and flavonoid variations concerning 63 accessions, a cluster analysis followed by the UPGMA method and Dice coefficient and principle component analysis (PCA) using the PAST v.3.18 (Hammer et al. 2001) and the Cluster Vis v. 1.8.2 (Raden 2012) software were assessed. The presence (1) and absence (0) of color spots were scored in this manner. In addition, the spots retention factor (Rf) in each sample is provided.
3 Results and Discussion
3.1 Morphology Results
The present work demonstrates the existence of some morphological variability at intraspecific levels. Most measurements of qualitative morphological variables were indicated for the indumentum of stem, leaf, inflorescence, bract, pedicel, calyx, corolla tube, corolla lips, filament and style (Table 1). Moreover, the highest value of quantitative morphological variations was mainly found in the length of leaf, inflorescence axis, bract, calyx and filament, and width of leaf, bract and calyx. Among quantitative morphological characters, some features including the length of stem (T. melissoides), length and width of leaf and length of inflorescence axis (T. parviflorum), length of bract (T. polium), width of bract (T. oliverianum), length of corolla lip (T. orienale subsp. taylori), length of calyx (T. orienale subsp. orientale), and length of filament and width of calyx (T. orienale subsp. glabrescens) were found as diagnostic characters in Teucrium species.
It is important to state that several unique qualitative morphological characteristics were presented as diagnostic characteristics in four subspecies of T. orientale. Identified characters included indumentum of stem surface (subsp. taylori, subsp. gloeotrichum, and subsp. glabrescens) followed by indumentum of leaf (subsp. glabrescens), indumentum of petiole (subsp. glabrescens), indumentum of inflorescence axis (in subsp. taylori and in subsp. glabrescens), indumentum of pedicel (subsp. taylori, subsp. gloeotrichum, and subsp. glabrescens), indumentum of corolla lips (subsp. taylori), indumentum of calyx (subsp. taylori, subsp. gloeotrichum, and subsp. glabrescens) and indumentum of filament (subsp. glabrescens) (Table 1). It is well known that T. orientale subsp. orientale is differed based on hirtellous indumentum on the surface of pedicel.
Some morphological characteristics of T. polium complex were also elucidated as taxonomic characters based on the following evidence: indumentum of leaf (T. polium), indumentum of stem (T. polium), indumentum of inflorescence axis (T. polium and T. capitatum), indumentum of bract (T. polium, and T. capitatum), indumentum of bracteole (T. polium and T. gnaphalodes), indumentum of pedicel (T. polium, and T. capitatum), indumentum of calyx (T. polium and T. capitatum), indumentum of corolla tube (T. capitatum), indumentum of corolla lips (T. polium), indumentum of style (T. polium) and indumentum of filament (T. polium and T. capitatum) (Table 1).
Among all taxa under investigation, some diagnostic morphological characters were identified (Figs. 1, 2, 3). T. capitatum revealed indumentum of stellate at leaf, bract and inflorescence, pilose at corolla tube and emarginate apex of leaf. Moreover, T. polium was differentiated by the presence of hirsute and barbate indumentum at stem, floccose at inflorescence, stellate and glandular at bracteole, glandular at corolla lip, hirsute, velutinous and lanate at calyx, hirsute at pedicel and barbate and hirsute at filament. The presence of pannose indumentum at bracteole was characterized in T. gnaphalodes.

Representative indumentum of T. capitatum, T. polium, T. gnaphalodes and T. scordium subsp. scordioides. a, b stellate at inflorescence and stem (T. capitatum), c, d lanate and velutinous at calyx (T. polium), e pannose at bracteole (T. gnaphalodes), f lanuginose at inflorescence (T. scordium subsp. scordioides)

Representative indumentum of T. orientale subspecies. a hirtellous at pedicel (T. orientale subsp. orientale), b, c glandular at stem and calyx (T. orientale subsp. gloeotrichum), d glabrous at pedicel and calyx (T. orientale subsp. glabrescens), e, f villous and tomentose at inflorescence and stem (T. orientale subsp. taylori)

Representative indumentum of T. polium, T. oliverianum, T. melissoides, and T. parviflorum. a villous at leaf (T. polium), b villous at corolla lip (T. oliverianum), c hirtellous at calyx (T. melissoides), d pilose at corolla tube (T. melissoides), e barbate at corolla lip (T. parviflorum), f hirsute at pedicel (T. parviflorum)
In section Scordium, morphological characters including sub-cordate base of leaf and obtuse apex of bract, indumentum of lanuginose at stem, inflorescence and bract were observed in T. scordium subsp. scordioides. Moreover, ovate-elliptic form of bract was found as diagnostic character in T. melissoides.
In section Teucris, T. orientale subsp. glabrescens was discriminated by the presence of hirsute and tomentose indumentum at petiole. The indumentum of glabrous and sub-glabrous was characterized at stem, inflorescence, bract, pedicel, calyx and filament, whereas the existence of villous indumentum at inflorescence, and articulate at pedicel and bract corresponded to T. orientale subsp. subsp. taylori. The indumentum of glandular at stem, pedicel and calyx was also attributed to T. orientale subsp. gloeotrichum. Moreover, both species T. oliverianum and T. parviflorum were differentiated by indumentum of corolla lip (barbate, pannose) and form of leaf apex.
The ANOVA test was assessed for quantitative morphological characteristics. Significant differences (*p < 0.05; F-value = 2.07–5.7) were provided in Teucrium species. They were assigned in length of leaf, stem, inflorescence, petiole, bract, bracteole, corolla tube and calyx. The Kruskal–Wallis test statistically approved the significant difference (*p < 0.01) for qualitative characteristics, including indumentum of inflorescence, bracteole, calyx, corolla lips, margin, base and apex of bracteole.
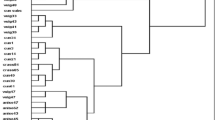
Based on the data presented in Fig. 4, the UPGMA dendrogram of morphological information showed two distinct groups. The first group is comprised of the members of section Teucris; T. orientale subsp. taylori, subsp. gloeotrichum, subsp. glabrescens, subsp. orientale, T. oliverianum and T. parviflorum. The second group was also formed by the members of the section Polium; T. polium, T. capitatum, T. gnaphalodes and the section Scordium; T. scordium subsp. scordioides and T. melissoides. According to the information, different groups were recognized, including T. polium with eleven groups, T. orientale subsp. taylori with seven groups, T. capitatum and T. gnaphalodes with four groups, T. scordium subsp. scordioides, T. gloeotrichum, T. oliverianum, T. orientale subsp. orientale, and T. parviflorum with two groups, and T. melissoides with one group. The main value of morphological variation was observed in the accessions of T. polium and T. orientale subsp. taylori. It has been found that in section Teucris; three subspecies of T. orientale were precisely separated. Nevertheless, it seems that the subsp. taylori with accession numbers 2, 6, 18 and 23 revealed few relations with subsp. gloeotrichum, subsp. glabrescens and subsp. orientale. It is important to point out that some accessions of these subspecies grow as sympatric distribution in Zagros regions. The presence of the specific indumentum of subsp. taylori led to display different groups such as accession no. 2 with hirtellous at stem and villous at inflorescence, accession no.18 with branched trichome at stem, tuberculate at petiole, style and leaf, accession no. 23 with pilose at bract and petiole and accession no. 6 with hirtellous at filament. In section Scordium, two accessions of T. scordium (nos. 40 and 45) were distinctly separated. It excludes variations caused by ecological and geographical conditions in Zagros regions as a possible reason. The morphological characteristics of T. scordium with accession no. 45 were differed from its accession no. 40, namely as oblong leaf, indumentum of pilose, tuberculate and lanuginose at stem, indumentum of lanate, pannose, glandular, tomentose and lanuginose at bract, and indumentum of tomentose and barbate at pedicel. Moreover, in section Polium, T. capitatum accessions were distinctly grouped, but T. gnaphalodes with accession number 51 was grouped with two T. capitatum accessions studied. Different characteristics, including indumentum of pilose at style and bracteole, led to this differentiation. It is remarked that morphological characteristics strongly resolve the complexity of sections Polium and Teucris.

Dendodgram of Teucrium species and their accessions using morphological characteristics. Pol: T. polium, cap: T. capitatum, gna: T. gnaphalodes, scor: T. scordium subsp. scordioides, melis: T. melissoides, gloeo: T. orientale subsp. gloeotrichum, tayl: T. orientale subsp. taylori, ori: T. orientale subsp. orientale, glab: T. orientale subsp. glabrescens, oliv: T. oliverianum, parv: T. parviflorum.1: section Polium, 2: section Scordium, 3: section Teucris
3.2 Chemotaxonomy Results
The flavonoid profiles of thin layer chromatography in Teucrium species showed an appropriate solvent system as CHCl3–MeOH (60:40). The extracts analysis in the present study revealed the existence of 353 color spots in Teucrium accessions. The variation in color spots was frequently provided as blue, fluorescent blue, green, dark yellow, yellow and fluorescent yellow. The color spots were presented under natural product identifiers, including blue, fluorescent blue, violet, yellow and fluorescent yellow. The retention factor (Rf) and m/z (mass to charge) were also estimated for each spot ranging from 0.49 to 0.59 and from 221 to 801 (Table 2).
Based on the following evidence on UPGMA dendrogram of flavonoid data (Fig. 5a), two main clusters were revealed. According to these findings, different groups were recognized. Nevertheless, the highest value of variability was observed in section Teucris, including T. orientale subsp. glabrescens with ten groups, T. orientale subsp. taylori with seven groups, and in section Polium, including T. polium with five groups. The lowest value of variability was provided with one group in section Teucris, including T. orientale subsp. orientale, T. parviflorum and T. oliverianum and in section Scordium, including T. scordium subsp. scordioides and T. melissoides. In section Teucris, further variations belonging to T. orientale subspecies taylori. T. orientale subsp. taylori with accession numbers 53 and 65 were significantly different from its accessions. It was identified that its accession no. 2 was clustered with T. orientale subsp. glabrescens with accession no. 11. This document was proved to be a factor showing the correlation and diversity of flavonoid at sub-specific levels. Moreover, there was a relation between T. orientale subsp. orientale with accession number 37 and T. orientale subsp. taylori with accession number 23 (Fig. 5a). However, they comprised of independent clusters. It is recognized that T. orientale subsp. orientale was discriminated by color spot (green).

a Dendrogram of Teucrium species and their accessions using flavonoid data. Pol: T. polium, cap: T. capitatum, gna: T. gnaphalodes, scor: T. scordium subsp. scordioides, melis: T. melissoides, gloeo: T. orientale subsp. gloeotrichum, tayl: T. orientale subsp. taylori, orien: T. orientale subsp. orientale, glab: T. orientale subsp. glabrescens, oliv: T. oliverianum, parv: T. parviflorum. b PCA analysis of Teucrium species and flavonoid data. orgo: T. orientale subsp. gloeotrichum, ortl: T. orientale subsp. taylori, or: T. orientale subsp. orientale, orgl: T. orientale subsp. glabrescens. c Circular dendrogram from the members of section Polium and d section Teucris using flavonoid data
Despite morphological similarity between the members of section Polium, the two species T. polium and T. gnaphalodes exhibited a clear-cut separation. The present evidence was observed exactly in T. polium and T. capitatum. T. capitatum with accession number 7 was accurately different from its other accessions. Flavonoid variation of T. polium accessions was significant, and in some cases, they were grouped with some T. capitatum accessions (Fig. 5a).
Principle component analysis (PCA) was also consistent with cluster analysis (Fig. 5b). Following this approach, in section Polium, T. polium accessions were strongly categorized from the two species T. capitatum and T. gnaphalodes. Moreover, in section Teucris, T. orientale subsp. glabrescens and T. orientale subsp. taylori exhibited high variations, but two subspecies were exactly classified using dark yellow and fluorescent blue spots, and the type of flavonoid classes.
To better characterize the exact position of the section Teucris members (T. orientale and its subspecies) and section Polium (T. polium, T. capitatum and T. gnaphalodes), cluster analysis was separately performed. The subspecies of T. orientale were definitely clustered. However, some relations exist between T. orientale subsp. taylori, T. orientale subsp. orientale and T. orientale subsp. glabrescens. Similarly, T. polium accessions were exactly distinguished even though some affinities were provided with T. capitatum (Fig. 5c, d).
3.3 Flavonoid Classes
According to flavonoid classes, seven groups were mostly recognized in the Teucrium species with the presence of flavone (18), isoflavone (17), flavonol (13), flavanone (3), chalcone (3), dihydroflavonol (2) and aurone (1) (Table 2). It is specifically found diagnostic flavonoid classes in the members of three sections. All sub-specific levels of T. orientale were discriminated with different flavonoid classes, including flavone nos. 6-11, isoflavone nos. 6-9 and flavonol nos. 1, 2 in subsp. glabrescens, flavone no. 18, flavonol nos. 11, 12 and chalcone no. 3 in subsp. taylori and isoflavone no. 13 in subsp. gloeotrichum. In addition, flavone no. 15, isoflavone nos. 3, 16 and flavanone no. 2 were observed in T. oliverianum. However, T. orientale subsp. orientale and T. parviflorum were identified by green and fluorescent yellow spots, respectively (Table 2). In addition, further flavonoid classes were ascribed to flavone nos. 4, 5 in T. capitatum, flavone no. 12, flavonol nos. 3-6 and flavanone no. 1 in T. gnaphalodes, and isoflavone no. 17, flavone no. 16 and flavonol nos. 18, 19 in T. polium (Table 2). Flavone no. 17 and flavonol nos. 10, 13 were assigned to T. scordium subsp. scordioides, and flavone no. 14 and isoflavone nos. 14, 15 were recognized in T. melissoides.
It was recognized that flavone 3 was the most common flavonoid class in all Teucrium species. The UV-spectral shift reagent for flavonoid classes ranged from 1 to 88 nm, and the λ max absorbance ranged from 224 to 377 nm (Table 2).
Further cluster analysis was performed using both morphological characters and flavonoid data (Fig. 6). This analysis completely separated the members of three sections through two different groups. Consequently, this analysis more thoroughly supports the taxonomic status of Teucrium species. Moreover, high diversity was observed in Teucrium accessions including T. orientale subsp. glabrescens (seven groups), T. polium (seven groups), T. orientale subsp. taylori (six groups), T. gnaphalodes (four groups) and T. capitatum (three groups). Other species consist of one group. As mentioned above, some relationships exist in a few accessions, e.g., T. orientale subsp. taylori, T. orientale subsp. orientale and T. orientale subsp. glabrescens, and T. polium and T. capitatum.

Dendrogram of Teucrium species and their accessions using both morphological characters and flavonoid data. Pol: T. polium, cap: T. capitatum, gna: T. gnaphalodes, scor: T. scordium subsp. scordioides, melis: T. melissoides, gloeo: T. orientale subsp. gloeotrichum, tayl: T. orientale subsp. taylori, orien: T. orientale subsp. orientale, glab: T. orientale subsp. glabrescens, oliv: T. oliverianum, parv: T. parviflorum
With the purpose of discrimination, two morphological and flavonoid keys were provided for Teucrium species and its subspecies.
Morphological key to Teucrium species
- 1a—Leaf pinnatifid or lobed in apex, inflorescence panicle:
-
2
- b—Leaf simple, inflorescence terminal racem or racem with two or five flower:
-
4
- 2a—Leaf apex with 3–5 lobed:
-
T. oliverianum
- b—Leaf deeply pinnatifid:
-
3
- 3a—Leaf 34-44 mm, corolla lip in upper surface 1–2.1 mm, filament 3–3.5 mm:
-
T. parviflorum
- b—Leaf 7.1–30 mm, corolla lip in upper surface 5–12 mm, filament 7–18 mm:
-
T. orientale
- 4a—Leaf linear, 7.1–19.3 mm:
-
5
- b—Leaf oblong and ovate-elliptic, 21–35 mm:
-
7
- 5a—Bract up to 17.6 mm, calyx velutinous or lanate:
-
T. polium
- b—Bract less than 17.6 mm, calyx non-velutinous or non-lanate:
-
6
- 6a—Indumentum of whole plant short, grayish:
-
T. capitatum
- b—Indumentum of whole plant long, white:
-
T. gnaphalodes
- 7a—Stem 60–80 cm, inflorescence 33–53 cm, upper corolla lip 2–3 mm, corolla white, leaf base acute:
-
T. melissoides
- b—Stem 30–46 cm, inflorescence 5–26 cm, upper corolla lip 1.2 mm, corolla pink, leaf base sub-cordate:
-
T. scordium subsp. scordioides
Morphological key to the subspecies of T. oreintale
- 1a—Stem non-glabrous:
-
2
- b—Stem glabrous:
-
subsp. glabrescens
- 2a—Plant glandular trichome:
-
subsp. gloeotrichum
- b—Plant non-glandular trichome:
-
3
- 3a—Stem non-dense indumentum, inflorescence up to 35 mm:
-
subsp. orientale
- b—Stem mostly dense indumentum, inflorescence up to 29 mm:
-
subsp. taylori
Flavonoid key to Teucrium species
- 1a—Flavonoid classes include flavone (15, 18) and isoflavone (3, 13, 16):
-
2
- b—Flavonoid classes include flavone (4–5, 12, 14, 16–17) and isoflavone (11, 14–15, 17):
-
4
- 2a-—Flavonid classes include flavone (18) and isoflavone (13):
-
T. orientale
- b—Flavonoid classes non-flavone (18) and non-isoflavone (13):
-
3
- 3a—Flavonid with flavone (15), flavanone (2) and isoflavone (3, 16), non-fluorescent yellow spot:
-
T. oliverianum
- b—Non-flavone (15), flavanone (2) and isoflavone (3, 16), with fluorescent yellow spot:
-
T. parviflorum
- 4a—Flavonoid classes include flavone (4–5, 12, 16) and isoflavone (11, 17):
-
5
- b—Flavonoid classes include flavone (14, 17), isoflavone (14–15) and flavonol (10, 13):
-
6
- 5a—Flavonoid with flavone (4–5), without specific isoflavone and flavonol:
-
T. capitatum
- b—Flavonoid with flavone (12, 16), isoflavone (11) and flavonol (3–6, 8–9):
-
7
- 6a—Flavonoid classes include flavone (14), isoflavone (14–15) and non-flavonol:
-
T. melissoides
- b—Flavonoid classes include flavone (17), non-isoflavone and flavonol (10, 13):
-
T. scordium subsp. scordioides
- 7a—Flavonoid classes include flavone (16), isoflavone (17) and flavonol (8–9):
-
T. polium
- b—Flavonoid classes include flavone (12), flavanol (3–6) and flavanone (1):
-
T. gnaphalodes
Flavonoid key to the subspecies of T. orientale
- 1a—Flavonoid with green spots:
-
subsp. orientale
- b—Flavonoid non-green spots:
-
2
- 2a—Flavonoid with chalcone (3), flavone (18) and flavonol (11–12):
-
subsp. taylori
- b—Flavonoid non-chalcone (3), with isoflavone (6–9, 13) or flavone (6–11), and flavonol (1–2):
-
3
- 3a—Flavonoid class includes isoflavone (13):
-
subsp. gloeotrichum
- b—Flavonoid classes include isoflavone (6–9), flavone (6–11) and flavonol (1–2):
-
subsp. glabrescens
Regarding the literature, Teucrium species comprise of complex groups, which need to be more investigated (Salmaki et al. 2016; Ranjbar et al. 2017). There is a lack of chemotaxonomic study in the Teucrium species in worldwide. Therefore, other recent studies provided in Teucrium were discussed here.
Regarding the flavonoid and morphological information of the section Teucris, the subspecies of T. orientale were certainly separated. Eshratifar et al. (2009) specified that the transection of midrib and leaf was different in the subspecies of T. orientale. According to the data presented in the UPGMA dendrogram of flavonoid profiles and quantitative and qualitative morphological characteristics (Table 1; Figs. 4,5a), three subspecies were definitely separated. It was noted that flavonoid profiles in cluster and PCA analyses presented high potential to discriminate four subspecies. The chemotaxonomic status of T. orientale subsp. orientale and T. orientale subsp. taylori displayed distinctly different specification. Cakir et al. (2006) also reported different amounts of flavonoid and iridoids in T. orientale var. orientale, confirming our evidence.
This was interpreted that there was a relationship between T. orientale subsp. taylori and subsp. glabrescens. In some cases of flavonoid profiles clustering, a group was found in T. orientale subsp. taylori with accession number 2 and subsp. glabrescens with accession number 11. Similarly, a relationship was observed between T. orientale subsp. taylori with accession numbers 58 and 63 and subsp. glabrescens with accession number 36. Since there was a complexity in section Teucris members, in the present research, a separated analysis using flavonoid information provided for the subspecies of T. orientale. It is elucidated that its subspecies were distinctly classified. Morphological data were consistent with those earlier reported that the foliar trichome type (branched and dense trichome) was similar in T. orientale subsp. taylori and subsp. glabrescens (Ecevit Gen et al. 2015). However, the intraspecific levels of T. orientale included different attributes (Ecevit Gen et al. 2015). This was proposed that T. orientale subspecies could be introduced as specific levels, but further knowledge is required to obtain the current concept. It is shown that T. orientale subsp. taylori illustrates high polymorphism (Rechinger 1982). Moreover, considerable geographical races were proved in Iran and India, which involved in gradual variability of trichomes (Rechinger 1982). The evidence can possibly ascertain the presence of intermediate species.
In formerly published works, T. orientale subsp. glabrescens and subsp. gloeotrichum exhibited similarity in terms of the thickness of upper parenchyma, collenchyma cell, upper palisade parenchyma and rows of spongious parenchyma (Eshratifar et al. 2009). Moreover, subsp. orientale and subsp. glabrescens have the same unbranched multicellular trichomes (Eshratifar et al. 2009; Ecevit Gen et al. 2015). In morphological findings, few relations were also observed between subsp. taylori (accession nos. 18, 2 and 23), subsp. glabrescens and subsp. gloeotrichum. In their natural habitats, some accessions of sub-specific levels grow in the same ecological regions of Zagros such as Lurestan and Isfahan provinces. Eshratifar et al. (2011, 2009) presented the similarities of these taxa in terms of shape and cell wall of nutlet. It is of note that both morphological and flavonoid data mainly represent distinct groups of T. orientale subspecies.
Ecevit Gen et al. (2015) have stated that there was similarity of trichome between T. orientale and T. parviflorum. In our morphological and both morphological and flavonoid analysis, a few similarities were found. In flavonoid evaluations, there was no similarity between two species. It was previously considered that a high complexity was occurred between T. orientale, T. oliverianum and T. parviflorum, and taxonomic delimitation was not resolved (Rechinger 1982).
Following presented results, T. parviflorum and T. oliverianum showed different groups. In previously published reports, the two species were different in the type of trichomes including multicellular and unicellular, the presence of micropapilate trichomes, and the shape of midrib (Eshratifar et al. 2009, 2011). In some cases, the similarities of two species were previously described in terms of the thickness of nutlet, the presence of one celled and 2–5 celled trichomes (Eshratifar et al. 2009, 2011). Apparently, the nutlet characteristics were found to be similar in both species. However, flavonoid profiles and morphological results (qualitative and quantitative characters) led to consider differentiation of the two mentioned species (Tables 1, 2).
In section Scordium, T. mellissoides was discriminated from T. scordium subsp. scordioides in terms of cluster and PCA analyses using flavonoid and morphological characteristics. Two species were previously different in type of epidermal cell, parenchyma, leaf trichomes and nutlet surface (Eshratifar et al. 2009, 2011). Controversially, Eshratifar et al. (2011) evidenced some resemblances such as long clavate glandular trichome of leaf between T. melissoides and T. scordium subsp. scordioides. Discrimination was also found in the quantitative and qualitative morphological characteristics studied (Table 1). Morphologically, the differentiation of the two studied accessions of T. scordium was presented, including Isfahan and Kurdistan provinces. To support the present flavonoid results, no chemical variations were previously provided in T. scordium subsp. scordioides (Jurisic Grubesic et al. 2012).
In section Polium, clustering and PCA analysis of both flavonoid and morphological markers were performed. T. polium accessions were mostly isolated from other species in this section. Three members of this section were precisely identified, which were obtained from morphology (quantitative and qualitative characters) and flavonoid information (Tables 1, 2). It was recognized that the indumentum of calyx was known as the specific taxonomic value. From previous published reports, different indumentum of calyx were identified between T. polium and T. capitatum (Rechinger 1982), confirming the present findings. Some T. polium with accession numbers 1 and 8 were grouped with T. capitatum with accession numbers 10 and 27. There was also a correlation between the two studied species responsible for flavonoid profiles. In the previous results, T. polium was introduced as a non-independent taxon, which needs to be further revised (Rechinger 1982). Based on the previous information of indumentum characteristics, two different groups were recognized in T. polium (Rechinger 1982). Moreover, high variation in leaf form and its indumentum led to the present imbricate characteristics, which did not certainly separate them (Jamzad 2012). According to our flavonoid and morphological evidence, several groups were recognized in T. polium. Bosabalidis (2013) reported the seasonally dimorphic of T. polium in Greece. It affects the type of trichome on the surface of leaf. Moreover, biodiversity characteristic was described in T. polium of Saudi Arabia, but environmental conditions did not affect the trichome structure (Bukhari et al. 2015). Furthermore, Pavlova and Vasileva (2010) investigated the effect of edaphic conditions on variability of vegetative characteristics in T. polium in Bulgaria. Jurisic Grubesic et al. (2012) and Venditti et al. (2017) also stated that variability of metabolites was found in T. polium.
According to the findings, similar or different accessions of Teucrium species were clustered together or separated including T. polium, T. scordium subsp. scordioides, and T. orientale subsp. taylori. The differentiation of T. polium accessions was related to the length of stem, leaf, petiole, bract, bracteole, calyx and indumentum of stem, corolla lip, pedicel and form of leaf apex. The similarities of its accessions were assigned to the length of inflorescence, corolla tube, filament and indumentum of leaf, inflorescence, bract, bracteole, calyx, corolla tube and style, and form of leaf, leaf base, leaf margin and bracteole. Evidence from the present research displays the variability of T. polium confirmed by published reports (Rechinger 1982; Jurisic Grubesic et al. 2007; Pavlova and Vasileva 2010; Jamzad 2012; Bosabalidis 2013). Both T. scordium subsp. scordioides accessions were also differed by the length of stem, leaf, petiole, bract, bracteole and indumentum of stem, bract, pedicel and leaf form. In T. orientale subsp. taylori, the differentiation of some accessions was assigned to the length of stem, leaf, petiole, inflorescence, bract, bracteole and calyx, and indumentum of stem, leaf, petiole, inflorescence, bract, filament and style. Their similarities were also assigned to the indumentum of calyx, corolla lip, pedicel and form of leaf and bract.
According to the results obtained by Valant-Vetschera et al. (2003), some chemo diversities were attributed to Teucrium species. In fact, some flavonoid compounds in this genus were attributed to sectional and intraspecific levels.
Ranjbar et al. (2017) referred to some morphological characteristics in section Polium; T. polium including glandular and unbranched trichome at stem, tomentose and branched trichome at calyx, lanceolate, oblong-lanceolate of leaf form, sinuate–undulate of leaf margin, and length of inflorescence axis ranged from 3 to 15 cm. Moreover, T. gnaphalodes includes eglandular and glandular trichome on stem surface, tomentose and glandular trichome at calyx, triangular-oblong of leaf form, and dentate, crenate and lobed of leaf margin. It should be noted that our reports in both species were completely different.
It was identified that flavonoid classes certainly resolved the taxonomic complexity of the species in sections Teucris and Polium. The identified flavonoid classes in the Teucrium species included 18 flavones, 17 isoflavones, 13 flavonols, 3 flavanones, 3 chalcones, 2 dihydroflavonols and one aurone.
In previous reports, the flavone derivatives were identified, including apigenin-glucoside in T. zanonii Pamp. (Abdelshafeek et al. 2006; Jaradat 2015), mono-hydroxy-methoxylated flavone in T. barbeyanum Aschers. (Alwahsh et al. 2015), luteolin glucoside and glucopyranoside in T. polium and T. chamaedrys subsp. sinuatum (Celak.) Rech.f. (Cakir et al. 2006; Galstyan 2014; Ozer et al. 2018; Gecibesler et al. 2019), and diosmetin in different Teucrium species (Mitreskia et al. 2014; Venditii et al. 2017). The obtained information was identical to that of the UV-spectral properties of flavone nos. 2 (λmax 257, 349 nm), 5 (λmax 344 nm), 9 (λmax 340 nm), 12 (λmax 333 nm), and 17 (λmax 242, 349 nm).
Flavonols derivatives were previously reported, including patuletin and fisetin in T. chamaedrys L. (Vlase et al. 2014), penduletin, quercitrin, quercetagetin-methyl ether, in T. polium (Ozer et al. 2018), quercetin in T. sandrasicum O. Schwarz (Kaska and Mammadov 2019), and quercetin and kaempferol glucoside derivatives in the Teucrium species (Mitreskia et al. 2014) supported the UV-spectral properties of flavonols no. 1 (λmax 362 nm), 2 (λmax 333 nm), 3 (λmax 371 nm), 7 (λmax 350 nm), 9 (λmax 254, 340 nm), 12 (λmax 244, 350 nm) and 13 (λmax 271, 340 nm).
According to the former literature, formononetin was isolated in T. polium as an isoflavonoid class (Mirdeilami et al. 2011). Regarding our findings, isoflavone no. 15 with λmax 301 nm illustrates consistency with its derivatives.
It is has been confirmed that flavonoid classes such as chalcone nos. 1 (λmax 344 nm) and 2 (λmax 318 nm), isoflavone nos. 4 (λmax 353 nm) and 6 (λmax 328 nm), flavone nos. 1 (λmax 323 nm), 3 (λmax 364 nm), 4 (λmax 350 nm), 8 (λmax 313 nm), 10 (λmax 333 nm), and 13 (λmax 307 nm), flavonol nos. 4 (λmax 370 nm), and 10 (λmax 377 nm), flavanone nos. 1 (λmax 312 nm), and 2 (λmax 326 nm), and dihydroflavonol no. 2 (λmax 308 nm) accorded with other genera of Lamiaceae such as Salvia L., Scutellaria L., Otostegia Benth., and Phlomis L. species (Mamadalieva et al. 2011; Sadeghi et al. 2014; Kharazian 2014; Uritu et al. 2018; Sajjadi et al. 2018; Jafari Dehkordi and Kharazian 2019; Hajialyani et al. 2019). It was identified that flavonoid classes such as flavone no. 6, 7, 10, 11, 14–16, 18, 19, isoflavone no. 1, 3, 5, 7–14, 16, flavonol no. 5, 6, 8, 11, flavanone no. 3, aurone no. 1, and dihydroflavonol no. 1 were first described for the Teucrium species.
4 Concluding Remarks
In the present work, flavonoid and morphological markers were ascertained as valuable indicators to detect the taxonomic relationships of the Teucrium species. Moreover, the flavonoid information provided some intraspecific relations in T. polium and T. capitatum, and T. orientale subspecies. Thus, the kind of flavonoid classes can be designated as a powerful marker to identify the boundaries of sub-specific levels. Nevertheless, the more detailed flavonoid compound identification of Teucrium species is in progress.
Due to the presence of morphological and chemical diversities in the genus Teucrium from Zagros region, it is required to protect this valuable germplasm and its endemic species for saving natural vegetation in this area.
References
Abdelshafeek KA, Ismail IA, Alwahsh MA (2006) Flavonoids and insecticidal activity of Teucrium zanonii. Planta Med 72:012. https://doi.org/10.1055/s-2006-949812
Aghakhani F, Kharazian N, Lori Gooini Z (2018) Flavonoid constituents of Phlomis (Lamiaceae) species using liquid chromatography mass spectrometry. Phytochem Anal 29:180–195
Ahmad Sh, Ullah R, Abdelsalam NM, Fouad H, Bibi A, Tariq Jan M, Ali Shad A, Arfan M (2014) One new royleanumoate from Teucrium royleanum Wall. ex Benth. Sci World J 2014:1–3
Alwahsh MAA, Melati Kh, Wong KCh (2015) Chemical constituents and antioxidant activity of Teucrium barbeyanum Aschers. Rec Nat Prod 9:159–163
Bentham G (1835) Labiatarum genera et specie. Teucrium. James Ridgway and Sons, London
Boghrati Z, Naseri M, Rezaie M, Pham N, Quinn RJ, Tayarani-Najaran Z, Iranshahi M (2016) Tyrosinase inhibitory properties of phenylpropanoid glycosides and flavonoids from Teucrium polium L. var. gnaphalodes. Iran J Basic Med Sci 19:804–811
Bosabalidis AM (2013) Glandular and non-glandular hairs in the winter and summer leaves of the seasonally dimorphic Teucrium polium (Lamiaceae). Biharean Biol 7:80–85
Briquet J (1897) Verbenaceae. In: Engler A, Prantl K (eds) Die natürlichen pflanzenfamilien. Engelmann, Leipzig, pp 132–182
Bukhari NA, Al-Otaibi RA, Ibhrahim MM (2015) Biodiversity characteristics of Teucrium polium species in Saudi Arabia. Saudi J Biol Sci 22:181–185
Cakir A, Mavi A, Kazaz C, Yildirim A, Kufrevioglu IO (2006) Antioxidant activities of the extracts and components of Teucrium orientale L. var. orientale. Turk J Chem 30:483–494
Ecevit Gen G, Ozcan T, Dirmenci T (2015) Micromorphological characters on nutlet and leaf indumentum of Teucrium sect. Teucrium (Lamiaceae) in Turkey. Turk J Bot 39:439–448
El-Ashmawy IM (2018) Anti-inflamatory and phytoconstituents of Teucrium oliverianum Ging. ex. Benth. Eur J Pharm Med Res 5:406–411
Elmasri WA, Yang T, Tran Ph, Hegazy MEF, Hamood AN, Mechref Y, Pare PW (2015) Teucrium polium phenylethanol and iridoid glycoside characterization and flavonoid inhibition of biofilm-forming Staphylococcus aureus. J Nat Prod 78:2–9
Eshratifar M, Attar F, Mahdigholi K (2009) Leaf anatomical study of the genus Teucrium L. (Lamiaceae) in Iran. Iran J Bot 1:196–204
Eshratifar M, Attar F, Mahdigholi K (2011) Micromorphological studies on nutlet and leaf indumentum of genus Teucrium L. (Lamiaceae) in Iran. Turk J Bot 35:25–35
Galstyan HM (2014) Standardization of felty germander (Teucrium polium L.) by natural phytoestrogens: phenylpropanoid and flavonoid glycosides. New Armen Med J 8:53–58
Gecibesler IH, Demirtas I, Koldas S, Behcet L, Gul F, Altun M (2019) Bioactivity-guided isolation of compounds with antiproliferative activity from Teucrium chamaedrys L. subsp. sinuatum (Celak.) Rech. f. Progr Nutr 21:458–470
Hajialyani M, Farzaei MH, Echeverria J, Nabavi SM, Uriarte E, Sobarzo-Sanchez E (2019) Hesperidin as a neuroprotective agent: a review of animal and clinical evidence. Molecule 24:648–666
Hammer O, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:9
Hossain MB, Rai DK, Brunton NP, Martin-Diana AB, Barry-Ryan C (2010) Characterization of phenolic composition in Lamiaceae spices by LC-ESI-MS/MS. J Agric Food Chem 58:10576–10581
Jafari Dehkordi F, Kharazian N (2019) Chemotaxonomy, morphology and chemo diversity of Scutellaria (Lamiaceae) species in Zagros, Iran. J Sci Islam Republ Iran 30:211–226
Jamzad Z (2012) Lamiaceae. In: Asadi M, Masoumi AA, Mozafarian V (eds) Flora Iran. Research Institute of Forest and Rangelands, Tehran, pp 43–85
Jaradat NA (2015) Review of the taxonomy, ethno botany, photochemistry, phytotherapy and phytotoxicity of germander plant (Teucrium polium L.). Asian J Pharm Clin Res 8:13–19
Jurisic Grubesic R, Vladimir-Knezevic S, Kremer D, Kalođera Z, Vukovi J (2007) Trichome micromorphology in Teucrium (Lamiaceae) species growing in Croatia. Biologia 62:148–156
Jurisic Grubesic R, Kremer D, Vladimir-Knezevic S, Vukovic Rodriguez J (2012) Analysis of polyphenols, phytosterols, and bitter principles in Teucrium L. species. Cent Eur J Biol 7:542–550
Kaska A, Mammadov R (2019) Antioxidant properties, proximate analysis, phenolic compounds, anthelmintic and cytotoxic screening of Teucrium sandraasicum; an endemic plant for Turkey. Ital J Food Sci 31:332–346
Kastner A (1989) Ubersicht zur systemstischen gliederung der gattung Teucrium L. Biocosme Mesogeen 6:63–78
Kharazian N (2014) Flavonoid constituents in some species of Salvia L. (Lamiaceae) in Iran. J Sci Islam Republ Iran 25:219–227
Kremer D, Stabentheiner E, Jurisic Grubesic R, Oberlander A, Vladimir-Knezevic S, Kosalec I, Ballian D (2012) A morphological and chemotaxonomic study of Teucrium arduini L. in Croatia, and Bosnia and Herzegovina. Plant Biosyst 146:402–412
Mamadalieva NZ, Herrmann F, El-Readi MZ, Tahrani A, Hamoud R, Egamberdieva DR, Azimova SS, Wink M (2011) Flavonoids in Scutellaria immaculata and S. ramosissima (Lamiaceae) and their biological activity. J Pharm Pharmacol 63:1346–1357
Marzouk R, El-Darier SM, Askar AB (2017) Pollen morphology of Teucrium L. (Lamiaceae, Ajugoideae) in Libya. Bangladesh J Plant Taxon 24:219–226
Mirdeilami SZ, Barani H, Mazandarani M, Heshmati GA (2011) Ethno pharmacological survey of medicinal plants in Maraveh Tappeh region, North of Iran. Iran J Plant Physiol 2:327–338
Mitreskia I, Petreska Stanoevaa J, Stefovaa M, Stefkov G, Kulevanova S (2014) Polyphenols in representative Teucrium species in the flora of R. Macedonia: LC/DAD/ESI-MSn profile and content. Nat Prod Commun 9:175–180
Navarro T, El Oualidi J (2000) Trichome morphology in Teucrium L. (Labiatae) a taxonomic review. Anal Jard Bot De Madrid 52:277–297
Ozer Z, Kilic T, Carikci S, Yilmaz H (2018) Investigation of phenolic compounds and antioxidant activity of Teucrium polium L. decoction and infusion. J Baun Inst Sci Technol 20:212–218
Pavlova DK, Vasileva MV (2010) Variation in morphology of Teucrium polium aggr. population in Bulgaria. Cent Eur J Biol 5:880–887
Perez I, Blazquez MA, Boira H (2000) Chemotaxonomic value of essential oil compounds in species of Teucrium pumilum aggregate. Phytochemistry 55:397–401
Raden K (2012) Cluster Vis Software for analysis and visualization of phylogenetic data. SequentiX - Digital DNA Processing, Klein Raden
Radulovic N, Dekic M, Joksovic M, Vukicevic R (2010) Chemotaxonomy of Serbian Teucrium sprcies inferred from essential oil chemical composition: the case of Teucrium scordium L. ssp. scordioides. Chem Biodivers 9:106–122
Rahimi MA, Nazeri V, Andi SA, Sefidkon F (2019) Variation in essential oil composition of Teucrium hircanicum L. from Iran-A rich source of (E)-α-bergamotene. Nat Prod Res 33:1227–1232
Rahman A (2005) Studies in natural products chemistry, bioactive natural products. Elsevier, Chicago
Ranjbar M, Mahmoudi Ch, Nazari H (2017) A contribution to Teucrium sect. Polium (Lamiaceae) in Flora Iranica. Novon 25:302–315
Rechinger KH (1982) Teucrium L. In: Rechinger KH (ed) Flora Iranica. Akademische Druck-U, Verlagsanstalt-Graz, pp 23–44
Rohlf FJ (2000) NTSYS 2.1: numerical taxonomic and multivariate analysis system. Exeter Software, New York
Sabet Z, Roghani M, Najafi M, Maghsoudi Z (2013) Antidiabetic effect of Teucrium polium aqueous in multiple low-dose streptozotocin-induced model of type1 diabetes in rat. Basic Clin Pathophysiol 1:34–38
Sadeghi Z, Akaberi M, Valizadeh J (2014) Otostegia persica (Lamiaceae): a review on its ethnopharmacology, phytochemistry, and pharmacology. Avicenna J Phytochem 4:79–88
Sajjadi SE, Delazari Z, Aghaei M, Ghannadian M (2018) Flavone constituents of Phlomis bruguieri Desf. with cytotoxic activity against MCF-7 breast cancer cells. Res Pharm Sci 13:422–429
Salmaki Y, Kattari S, Heubl G, Brauchler Ch (2016) Phylogeny of non-monophyletic Teucrium (Lamiaceae: Ajugoideae) implications for character evolution and taxonomy. Taxon 65:805–822
Semiz G, Celik G, Gonen E, Semiz A (2016) Essential oil composition, antioxidant activity and phenolic content of endemic Teucrium alyssifolium Staph. (Lamiaceae). Nat Prod Res 30:2225–2229
Shah M, Shah SMH (2015) Phytochemical, antioxidant, antinociceptive and anti-inflammatory potential of the aqueous extract of Teucrium stocksianum Boiss. BMC Complement Altern Med 15:351–358
Sonboli A, Bahadori MB, Dehghan H, Arabi L, Savehdroudi P, Nekuei M, Pournaghi N, Mirzania F (2013) Chemotaxonomic importance of the essential oil composition in two subspecies of Teucrium stocksianum Boiss. from Iran. Chem Biodivers 10:687–694
Stefkov G, Karapandzova M, Stefova M, Bankova V, Kulevanova S (2012) HPLC and UV-spectrophotometry analysis of flavonoids in spray-dried and freez-dried extracts of Teucrium polium L. (Lamiaceae). Maced Pharm Bull 58:39–44
Uritu CM, Mihai CT, Stanciu GD, Dodi G, Alexa-Stratulat T, Luca A, Leon-Constantin MM, Stefanescu R, Bild V, Melnic S, Tamba BI (2018) Medicinal plants of the family Lamiaceae in pain therapy. Pain Res Manag 2018:1–44
Valant-Vetschera KM, Roitman JN, Wollenweber E (2003) Chemodiversity of exudate flavonoids in some members of the Lamiaceae. Biochem Syst Ecol 31:1279–1289
Venditti A, Frezza C, Majd Zadeh SM, Foddai S, Serafini M, Bianco A (2017) Secondary metabolites from Teucrium polium L. collected in Southern Iran. Arab J Med Aromat Plants 3:108–123
Vlase L, Benedec D, Hanganu D, Damian G, Csillag I, Sevastre B, Mot AC, Silaghi-Dumitrescu R, Tilea I (2014) Evaluation of antioxidant and antimicrobial activities and phenolic profile for Hyssopus officinalis, Ocimum basilicum and Teucrium chamaedrys. Molecules 19:5490–5507
Yuzepchuk SV (1954) Teucrium L. In: Komarov VL (ed) Flora of the USSR. Academy of Sciences of the USSR, Moscow, pp 27–47
Acknowledgements
This project was supported by the Research Deputy of Shahrekord University. We are grateful to the Central Laboratory and Financial Affairs at Shahrekord University [Grant No. 96GRD1M1987].
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Table 3.
Rights and permissions
About this article

Cite this article
Bagheri Moghadam, H., Kharazian, N. Morphologic and Chemotaxonomic Studies of Some Teucrium L. (Lamiaceae) in Zagros Region, Iran. Iran J Sci Technol Trans Sci 44, 933–953 (2020). https://doi.org/10.1007/s40995-020-00908-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40995-020-00908-1