
Abstract
Based on the assumption that substitution of functions or function transfer is a fundamental principle underlying all conditioning processes, we attempt to produce an account wherein both operant and respondent events are understood in terms of substitution. We contend that, if event interactions are described in a way that accounts for all the stimulus and response events involved in a contingency relation, and further, if we assume symmetry as a property of substitution of functions as excitatory backward conditioning findings seem to suggest, traditional distinctions between operant and respondent conditioning may be rendered unnecessary. We present a reconceptualization of respondent and operant processes and suggest alternatives for empirical research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Several research studies have shown that stimulus-stimulus (S-S) relations are sufficient to describe a variety of human and animal behaviors that have been traditionally explained by reinforcement contingencies (Delgado and Hayes 2011; Delgado and Medina 2011; Delgado et al. 2011; Leader et al. 1996; Rehfeldt and Hayes 1998; Tonneau and González 2004; Tonneau et al. 2006). In the first part of this paper, we refer to some of this evidence and briefly discuss its associated theoretical implications. Based on these arguments, we examine the generality of substitution of functions beyond its operation in psychological events involving S-S relations. We do so by showing that when descriptions of contingency relations include all of the response and stimulus functions involved, distinctions between respondent and operant relations may be an artifact of our investigative procedures. Following this, we turn to the specific applicability of the notion of substitution for the explanation of psychological events of the R-S type (operant contingencies), and we discuss the conditions under which substitution may occur in these cases. Finally, we conclude that bidirectional relations are sufficiently supported by empirical research and are conceptually consistent with the notion of function transfer or stimulus substitution as to constitute the underlying foundation of all learning processes (Delgado and Hayes 2013).
Current research in Pavlovian conditioning of human complex behavior has shown that, as Rescorla once affirmed, “Pavlovian conditioning is not what we think it is” (Rescorla 1988). More specifically, Pavlovian conditioning is no longer restricted to the study of the conditioned reflex or limited to the participation of a biologically relevant stimulus; rather, it may be better characterized as the study of S-S associationsFootnote 1, whereby given a history of contiguous and contingent presentations of stimuli, substitution of functions may be observed for responses to both of the stimulus objects involved (e.g., De Houwer et al. 2002; Denniston et al. 1996; Delgado and Hayes 2013; Delgado and Medina 2011; Delgado and Hayes 2013; Tonneau 2001; Tonneau and González 2004).
In addition, the signalizing function of the conditioned stimulus (CS) appears to be a causal interpretation of the relation between two events that are presented contingently, contiguously and in a specific order for a number of trials. In particular, research on evaluative conditioning (Walther 2002; Walther et al. 2005), cue competition effects in humans (De Houwer et al. 2002; Glautier 2002; Wilson and Alexander 2008), and respondent stimulus equivalence (Delgado and Medina 2011; Delgado et al. 2011; Leader and Barnes-Holmes 2000; Leader et al. 1996; Tonneau and González 2004) suggests that the notion of function transfer adequately describes the outcomes of these non-causal types of stimulus relations (Delgado and Hayes 2013).
In the operant literature, equivalence relations have been described as emergent or derived (Sidman 1994; Sidman and Tailby 1982), for lack of an explanation as to why these relations are observed in the absence of reinforcement (i.e., direct training). However, referring to these findings as “generalized relating,” as Relational Frame Theory (RFT) suggests (Hayes et al. 2001), leaves the first occurrence of such generalized relating unexplained (Tonneau and González 2004), and thereby does little more than give the occurrence of emergent relations a new name. Incidentally, Sidman (2000) makes this same critique with respect to the notion of function transfer. Nonetheless, accounts based on the notion of function transfer (Delgado and Hayes 2013; Hayes 1992; Tonneau 2001, 2002; Tonneau and González 2004), seem to offer more explanatory value than more mainstream operant approaches. Function transfer accounts have attempted to describe derived relations by way of substitution of stimulus functions occurring under particular circumstances of event correlations. By contrast, stating that relating is a generalized operant constitutes a theoretical assumption that has not been subjected to systematic empirical examination.
As pointed out by others (Tonneau 2001; Tonneau and Sokolwski 1997), outcomes analogous to those of transitive relations have been demonstrated by way of Pavlovian procedures such as sensory preconditioning and second order conditioning prior to the emergence of equivalence research in the operant literature. Still, explanations of derived symmetry based on stimulus correlations have been largely debated. While some authors have provided accounts of symmetry in terms of forward conditioning via mediated generalization (Hall 1996), others (Barnes-Holmes et al. 2001) have argued that since Pavlovian accounts of symmetrical associations involve backward conditioning, which, according to most, does not appear to produce strong excitatory conditioning effects (see Mackintosh 1974), the account is theoretically untenable.
Nonetheless, data from several studies contradict this claim. For example, Zentall et al. (1992) trained pigeons in conditional discriminations involving differential outcomes (i.e., forward training). Results from their three experiments showed transfer effects consistent with the presence of bidirectional associations (see also Sherbrune and Zentall 1995). Similarly, Hearst (1989), and Hemmes et al. (1994) found evidence for excitatory backward conditioning in pigeons using an autoshaping preparation.
A brief overview of the backward conditioning literature shows that far from disputing whether or not excitatory backward conditioning occurs, researchers are currently focusing on examining the conditions that facilitate bidirectional relations. For example, some studies on backward conditioning have suggested that one of the critical variables in producing excitatory (and not inhibitory) effects given unconditioned stimulus-conditioned stimulus (US-CS) associations is the number of training trials to which participants are exposed. These studies show that while a large number of pairings tend to produce inhibitory conditioning, fewer training trials are associated with excitatory conditioning (Barnet and Miller 1996; Chang et al. 2003; Cole and Miller 1999; Spetch et al. 1982). These findings have been inconsistent, however. Other studies have shown reliable backward associations even with a large number of conditioning trials (Hearst 1989; Hemmes et al. 1994).
Overall, the literature shows that bidirectional associations occur quite reliably (Silva et al. 1996). Backward associations in non-human animals have been observed in instrumental conditional discrimination procedures, autoshaping procedures and in traditional Pavlovian procedures. Possibly, as Spetch et al. (1981) assert, the general acceptance of the weakness of backward associations may be due to the fact that traditional models of Pavlovian conditioning do not predict transfer after backward pairings, and/or, to the rigidity of procedures and the selection of dependent measures used to study it (see also Hemmes et al. 1994; Silva et al. 1998).
It may be the case that the biological relevance of the correlated stimuli affects function substitutability. Some Pavlovian accounts have argued that substitution (in cue competition preparations particularly) may occur more or less readily, depending on whether or not the US is a biologically relevant stimulus (Denniston et al. 1996). In our view, bidirectional relations may be facilitated when stimuli are arbitrary or non-biologically relevant (Delgado and Hayes 2013).
This is the case of most experimental preparations with human subjects (respondent and operant) in which pairings of words are frequently involved, as in studies of social behavior and predictive learning (Olson and Fazio 2002; Staats and Staats 1958), or when arbitrary characters or symbols are used in the context of equivalence studies (e.g., Delgado and Medina 2011; Clayton and Hayes 2004; Rehfeldt et al. 1998; Tonneau and González 2004). Interestingly, symmetrical relations are typically demonstrated when these types of stimulus objects are used. However, as pointed out by Hayes (1992), more research is needed on the conditioning of the perceptual response associated with the CS in simple classical conditioning preparations. The development of procedures that produce indirect measures of perceptual behavior would be particularly useful in animal research.
Nonetheless, there seems to be enough evidence to believe that function transfer may be symmetrical in both humans and animals. Logically and empirically, it seems difficult to sustain the view that function transfer occurs for only one of the stimuli in the S-S contingency. Rather, we suggest that due to the contingent relation between stimulus events, some of the functions of each will come to be present in the other (Hayes 1992; Delgado and Hayes 2013).
On the basis of the findings thus far discussed, we agree with Tonneau (2001, 2002) that stimulus substitution or function transfer is the process operative in all S-S relations (Delgado and Hayes 2013). Because the term function transfer is also associated with operant descriptions of equivalence relations, we will use Kantor’s (1924, 1959) notion of substitution of functions throughout the remainder of this analysis.
Substitution of functions takes place when, by virtue of a history of contiguous presentations of A and B, each stimulus acquires some of the functional properties of the other. The outcome is that when only one of the stimuli is present, it acts as a substitute for the absent stimulus. That is to say, because some of the functional properties of the absent stimulus are present in the other stimulus, the organism responds to the latter as if in the presence of the former. The organism is responding to the functional properties of the absent stimuli inhering in the present stimulus.
A lack of attention to Pavlovian conditioning accounts of behavior has hindered the evolution of important areas of psychological research that have also been described in terms of S-S relations and/or in terms of substitution processes in general (Tonneau 2002). This lack of interest in classical conditioning accounts is probably due to the historical understanding of classical and operant conditioning as processes operating upon different types of phenomena.
Nonetheless, in acknowledging the generality and scope of Pavlovian relations, two theoretical implications can be drawn. It could be argued that most psychological events (or at least more than initially thought) may be accounted for in terms of the classical conditioning paradigm. This conclusion is unlikely to be accepted after decades of understanding behavior as partitioned into two distinct categories, however. Possibly, a more constructive way to understand this implication is to say that whether we call behavior operant or respondent, the notion of substitution of functions may be sufficient to describe behavior as a result of its interdependent relation with a stimulating environment (Delgado and Hayes 2013).
Still, even if it is assumed that substitution of functions is the process that accounts for both respondent and operant relations, reasons to classify behavior into one type or the other may still be supported. For instance, the distinction between operant and respondent behavior could be useful in differentiating the conditioning of reflexive from other types of responses. In defense of this argument, one could refer to respondent relations as those in which a biological response is involved in the contingency. This distinction between reflexive/non-reflexive processes would prove warranted if it were demonstrated that a particular set of behavioral principles was applicable to the description of conditioned reflexes exclusively. At present, though, further evidence from comparative research is needed to support this possibility.
It could also be argued that in contrast to classical or respondent conditioning, operant conditioning describes a circumstance in which the reinforcement contingency plays a critical role in the acquisition and maintenance of behavior. However, we will re-interpret this view by attempting to show that substitution processes may also account for reinforcement contingencies. If this analysis is successful, the distinction between operant and classical processes will be rendered conceptually unnecessary. We now turn to examine the nature of psychological events and the distinguishing properties of what have been conceived to be different types of contingency relations.
The Psychological Event as an Outcome of Conditioning
As Kantor (1924, 1959) asserts, the possibility of one stimulus being partially substituted for another that is not physically present allows for the consideration of a wide range of psychological activities that could otherwise not occur (see also Pawlik 1997). Kantor refers to these events as instances of implicit behavior fields and explains that such behaviors are as subject to naturalistic investigation as those occurring directly with respect to immediately present stimulus objects (Kantor 1959; Parrott 1986; Midgley and Morris 2006; Hayes and Fryling 2010).
In our view, most of the phenomena that we call psychological include implicit responses to substitute stimulation (Hayes and Delgado 2005, 2006). Because implicit responses are independent of the original stimulating conditions in which they developed, there is a greater degree of variation in responses with respect to substitute stimuli than in responses with respect to original stimulus objects. For example, in the case of human behavior, perceptual responses may reflect variations in some of the physical dimensions of the original stimulus as it occurs in remembering (Fryling and Hayes 2010) or dreaming (Dixon and Hayes 1999). This process may entail symbolic activity or substitution of conventional functions of stimuli (Kantor 1924, 1959; Mahan 1968). Without question, the notion of substitution has been fundamental both to the understanding of S-S relations and to the study of complex psychological phenomena such as memory, perception and symbolic behavior.
Taking further the generality of this process, we argue that if substitution of functions accounts for reflexive, as well as for a wide range of other non-reflexive behavior involving S-S relations, then it may also be fundamental in relations other than S-S; namely S-R, R-S or R-R (behavior-behavior relations). In the next part of this paper, we argue that the process of substitution of functions applies to all interactions of stimulating and responding irrespective of whether the elements of those interactions are conceptualized as stimuli, responses, or more adequately, as event units of stimulating and responding.
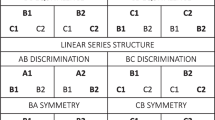
Traditionally, behavior analysts have studied four types of stimulus–response relations corresponding to all possible combinations of these elements. They have also categorized each type of relation as having its own set of principles, and hence have grouped them into different types of learning processes. Table 1 below summarizes the traditional behavior analytic view of these four possible stimulus–response interactions.
Notice how these relations, whether conceptualized as operant or respondent, always follow a linear or sequential order, where the first element of the relationship controls or produces the other in an if-then manner. Probably, it is because it has been assumed that operant processes control a greater variety of psychological events that S-S relations have been given a marginal role in behavior analysis. Notice also that all relations except for S → S fall under the umbrella of operant contingencies, where S-R represents the control of responses by discriminative stimuli, R-S represents the response-reinforcer contingency and R-R, represents behavior-behavior relations. Examples of R-R relations may include Premack’s (1965) and Timberlake and Allison’s (1974) views on reinforcement, where the relation emphasized is not between a response and a reinforcing stimulus, but between a response and a reinforcing response. Behavioral chains and relations between private events (e.g., self rules) and overt behavior could also be construed as examples of these relations (see S. Hayes and Brownstein 1986).
Extrapolating from the evidence obtained from the study of S-S relations, we considered it likely that substitutive processes could also account for all of these types of psychological interactions. What makes this seem a problematic extrapolation is the imprecise manner in which psychological events are depicted in Table 1. The insufficiency of such descriptions lies in failing to acknowledge that the notions of stimulus and response, necessarily and by definition, imply their stimulus or response counterparts (Kantor 1924, 1959; Parrott 1987). Consequently, a corrected version of these relations requires completing the omitted elements in each of the four interactions depicted in Table 1 (see Fig. 1).

Corrected description of behavioral interactions. The figure shows associations between sets of events (1 and 2). The top row of S and R events corresponds to pairings of stimuli and/or responses that apparently constitute different types of contingencies. In the bottom row we have added the corresponding stimulus and response elements that complete each event in the pair
As illustrated in Fig. 1, each stimulus or response involved in an association is itself part of an S ⇔ R interaction that may or may not be reflexive. A reflexive interaction per se is not a psychological event however. Interactions pertaining to the psychological domain include those in which responses, though still biologically determined, occur in the absence of the original eliciting stimulus. As a result of the conditioning of reflexive responses, for example, the conditional stimulus affects the organism from a distance (cited by Pawlik 1997); that is to say, without direct physical contact with the conditional response. This is how perceptual activity (i.e., perception of color, form, textures, etc.) may produce responses that bear no direct relation to the physiological effect of the unconditional stimulus (Pawlik 1997).
As early as 1904, Pavlov described conditioning as a process that, being psychological, is not localized inside the organism. His views on what are considered to be psychological events are consistent with Kantor’s, who asserts that learning does not occur in either the organism or the environment (Kantor 1924, 1959). Learning is a process and an outcome of an interaction between the behavior of the organism and conditions of stimulation. Thus, a learning event is best characterized as a coordination of stimulus and response functions (Kantor 1924, 1959; Midgley and Morris 2006).
However, as the term associative learning implies, any learning event entails more than a coordination of stimulus and response functions. As illustrated in Fig. 1, conditioning requires contingency relations between at least two pairs of interactions between functions of stimuli and responses. Said more precisely, conditioning occurs as an outcome of the process of substitution of functions between two or more instances of stimulating and responding. Thus, generally speaking, we would define a psychological event as an outcome of a conditioning process in which an organism has been exposed to a contingency relation between two or more stimulus–response interactions.
More complex psychological interactions involve contingency relations among stimulus–response events that are less dependent on the biological constraints of responses as reactions to the physical properties of stimulus objects (Hayes and Delgado 2005). Although generally we speak of associations between two sets of stimuli and responses, complex behavior often involves networks of associations where three or more S-R events may be involved. For example, responding with respect to classes of stimuli as observed in equivalence studies entails responding in a substitutive manner with respect to all members of the same class of stimuli (e.g., a1, b1 and c1). In this circumstance, it is likely that behaving with respect to each event as a class member entails responding with respect to the relational functions shared by all of them (Delgado and Hayes 2007).
A fundamental property of psychological events, though frequently overlooked, is the interdependence between the events involved in a contingency relation. This interdependence is present between the stimulus and response functions of each of the events comprising the contingency. The double-headed vertical arrows in Fig. 1 emphasize such interdependence between all the involved stimulus and response elements.
In some cases, as in withdrawing a hand from a hot surface, responses seem to follow stimuli, thus producing the illusion of sequentially organized occurrences. In other cases, conditions of stimulation and responding are less distinguishable as events occurring within discrete and successive segments of time (Parrott 1987). Seeing a cup or screaming upon encountering painful stimulation are examples of the latter case. Behavior-environment interactions may be better understood in terms of a set of events (physical or non-physical, organismic or environmental), which affect the probability of occurrence of a target behavior, whether they occur before, after or simultaneously with it (Parrott 1987). The notion of participating factors as opposed to cause and effect relations is even clearer in the analysis of behaviors such as running, grooming or washing. In these cases, organisms behave with respect to a variety of stimulating conditions, all of which involve perceptual events and all of which are subject to substitution.
It follows from these arguments that the four types of psychological events presented in Fig. 1 are not as different from one another as they appeared to be in Table 1. An argument against grouping these relations as the same type of psychological event could be made if substitution were to occur among different elements of the S-R pairs in each type of psychological event. For example, it might be said that, in S-S associations, substitution occurs between the stimulus functions of each interaction, while in R-R associations, it is the response functions that are substituted. This would be an error though—one that results from conceptualizing stimuli and responses as separable and independent units. In any learning outcome, what are partially substituted are always the circumstances of both stimulating and responding comprising the absent event.
When contingency relations are viewed this way, the logical conclusion is that substitution of functions is not peculiar to S-S associations, but rather is the foundation of conditioning of all types of psychological events. Based on this assumption, we will examine the extent to which some characteristics of S-S relations (i.e., symmetrical relations), and the conditionability of non-biological stimuli, favor substitution as an overarching process and not classically conditioned behaviors exclusively. In the following section, we focus on how substitution of functions may account for operant as well as for respondent contingencies.
Operant Conditioning
Most behavior analysts since Skinner’s time hold reinforcement to be the most important psychological principle, as it is said to explain the acquisition of a behavioral repertoire (O’Donohue and Ferguson 2001; Schoenfeld 1995). However, a cogent account of complex human behaviors such as verbal behavior, remembering, imagining or any behavior that entails responses with respect to non-physical sources of stimulation has yet to be achieved from an operant perspective. Some argue that such complex behaviors simply cannot be explained by a theory of operant behavior (Timberlake 2004). Others have shown that while research in behavior analysis is based almost exclusively on operant contingencies, for decades most treatments for behavioral disorders have involved classical conditioning processes (O’Donohue and Ferguson 2001; Savastano and Miller 2004).
Further, as shown above, current research makes it increasingly difficult to justify the operant-respondent dichotomy (Tonneau et al. 2006; see also Delgado and Hayes 2013). Finding that large classes of human behavior (e.g., the acquisition of behavior with respect to stimulus classes, and the attribution of causal or social judgments) may be acquired in the absence of operant reinforcers, raises the question as to whether or not other behaviors may be acquired in the same way. Based on the research findings thus far discussed, we can assert with some degree of confidence that reinforcement may not be necessary for the acquisition of responses with respect to arbitrary and/or conventional stimuli (Delgado and Medina 2011; Delgado et al. 2011). Moreover, it seems likely that when an organism is exposed to an S-S contingency, which is inherent in conditional discriminations of any sort, learning occurs as in any case of responding with respect to stimulus classes.
Applying the same logic of recent findings in the classical conditioning literature to the operant case, we may speculate that the principle of reinforcement may not be sufficient to account for the acquisition of behavior in general. In the following section, we attempt to further defend this statement by proposing an interpretation of operant relations in terms of substitution of functions.
Re-examining Operant Processes
From our perspective, there is nothing about the R-S association that makes it substantially different from events designated as S-S, R-R or S-R. To be precise, we argue that the reinforcement contingency could be adequately explained in terms of substitution of functions between coordinations of stimulus and response events. We will try to defend this claim by examining the arguments in favor of conceptualizing reinforcement as a separate process while comparing it to S-S relations as a reference point.
Having acknowledged the generality of substitutive activity in the psychological event, the following analysis will determine if there are any particularities about events of the R-S type that warrant its conceptualization as a different type of learning process, or if, as we suspect, all circumstances of substitution encompass the notion of reinforcement. We will first present some general arguments highlighting the similarities between respondent and operant learning. Then we will proceed to analyze the apparent differences between these learning processes, examine the reasons for these differences and finally, demonstrate that a more comprehensive view of behavioral events leads to a unified account of conditioning.
The defining feature of reinforcement is that it increases behavior probability. This increment is measured against the probability of responding during baseline conditions. Let us consider if a similar outcome is observed in S-S associations. As a result of a history of CS-US pairings, the probability of responses originally coordinated with the US increase above baseline levels. For example, while prior to conditioning, salivating occurs with respect to the US exclusively; after conditioning it occurs not only when the US or other formally similar stimuli are present, but also when a bell or stimuli physically similar to a bell are present.
In the respondent case, salivating to the food is measured by the amount of saliva produced. In both cases, however, the probability of responding is always 0 or 1. In addition, while the food powder is presented in discrete trials, the lever is permanently present. If the lever were also presented in discrete trials, conditioning could be measured in terms of the time it takes the animal to press the lever after it is made available.
Hence, the difference between elicitation and emission appears to be either procedural or limited to the distinction between reflexive and non-reflexive behavior. However, we question the extent to which this distinction continues to be useful. Some current respondent research involving non-biologically relevant stimuli employs methods similar to those used in operant research (Tonneau and González 2004; Tonneau et al. 2006), and a wide range of applied operant research and practice is based on a discrete trial model.
In respondent learning, the conditioning of reflexive responses introduced particular measurements and methods not used in operant learning. In fact, most of the differences attributed to these two processes are the result of confusing processes with procedures, or defining the former in terms of the latter (Pear and Eldridge 1984; Rehfeldt and Hayes 1998). As we have seen, though, behavior cannot be separated into dichotomous and discrete categories of reflexive and non-reflexive (Timberlake 2004; Wetherington 1982). It is rather a continuum that extends from behaviors that are more biologically determined to behaviors that are more biologically detached (Kantor 1924, 1959).
As shown in Fig. 1, all varieties of S-R pairings seem to constitute the same phenomenon. However, a more detailed comparison between respondent learning and learning by reinforcement contingencies may be achieved by examining the outcomes in each case. For illustrative purposes, in Fig. 2, each learning type is illustrated by its most representative experimental procedure.

Comparison between respondent and operant learning. Stimulus and response functions comprising each of the events paired in a respondent (left) and operant (right) contingency
At first glance, we observe notable differences in terms of outcomes, that is, different events seem to be changed in each type of learning as a result of the conditioning process. While in operant learning, event 1 increases in probability of occurrence as a result of systematic event 1–event 2 pairings; in respondent learning, it is the magnitude (and likelihood) of event 2 that increases. Furthermore, in respondent learning, the response factor of event 2 (salivation) is likely to be observed in the presence of the stimulus factor of event 1 (bell); while in operant learning, the response factor of event 1 (pressing) does not occur in the presence of the stimulus factor of event 2 (food), but rather, in its absence. In sum, it appears as though there are two main points of divergence: one pertains to the event that increases in probability (or the event that is changed) as a result of conditioning and the other pertains to the presence or absence of the unchanged event as a requirement for the occurrence of the other event.
The first of these distinctions may be the outcome of an experimental bias. The event showing an increase in probability in each of these learning types is the event that is either more conspicuous, more available for measurement, or simply the one that is purposefully programmed for change.
In respondent learning, evidence of conditioning is provided by salivation to the bell when the organism is food deprived. However, if the experimenter were to not ring the bell, one could expect behavior with respect to the bell to be observed (as in autoshaping experiments). It may also be speculated that in the absence of the CS, a deprived organism may behave perceptually with respect to it as in hearing it or seeing it in its absence (Hayes 1992); and further, that the probability of occurrence of these behaviors will be higher compared to baseline levels. However, because in contrast to reflexive responses, perceptual responses are difficult to measure, their occurrence has remained largely unexplored.
Similarly, because some of the functions of food/eating will inhere in lever pressing as a result of the operant contingency, it is likely that some response functions related to the food are present in lever pressing (i.e., salivation). Thus, in operant conditioning, behavior with respect to the food is also changed. It now occurs with respect to the stimulus object food and with respect to the food functions now present in the lever. In sum, most likely in operant as well as in respondent learning, both events involved in the contingency will be changed as a result of the conditioning process. Responses will be likely to occur with respect to their corresponding stimulus object and with respect to another stimulus object now having some of the functions of the original stimulus object.
Further, the point regarding the presence or absence of the apparently unchanged event results from confusing stimulus objects with their stimulating functions (Kantor 1924, 1959). This confusion is caused by focusing on the formal characteristics of psychological events exclusively (physical dimensions of stimuli, and response topographies), hence compromising a functional analysis of behavior.
Any behavioral event that has not undergone conditioning will occur only with respect to its original source(s) of stimulation. Lever pressing behavior will occur with respect to a lever; eating, salivating and all food-related behaviors will occur with respect to food, and hearing a bell or seeing a bell will occur with respect to a bell. After conditioning, though, responding with respect to each stimulus object constitutes a transformed event having functions additional to those related to the physical properties of stimulus objects.
Thus, in both respondent and operant conditioning examples, the measured response occurs in the presence of the other event. In respondent conditioning, salivation occurs in the presence of the bell, which is physically present; and in operant conditioning, lever pressing occurs in the presence of the food-eating functions that are now partially present in lever. The topography of lever pressing occurs both as exploratory behavior and, after conditioning, as food-related behavior. Similarly, salivation occurs in the presence of the food and in the presence of the bell, and in both cases it is also food related behavior; that is to say, both lever pressing and hearing the bell become part of the food-eating event.
Pavlov introduced the concept of reinforcement to the study of behavioral events (Schoenfeld 1995). In Pavlov’s interpretation, the US is considered a reinforcer to the extent that it increases and sustains the power of the CS to act as its substitute (Pawlik 1997). Notice that in Fig. 2, event 2 is the reinforcer for event 1 in both operant and respondent learning. In operant learning, the reinforcer increases lever pressing as a food-related response; and in respondent learning, the reinforcer may increase the probability of responding with respect to the bell, also a food-related response. In both cases the reinforcer alters the function of event 1 (lever pressing and hearing the bell) by way of substitution of functions, so as to turn the latter into a food-related response.
By this means, a new event is established having the topography of lever pressing and some of the functions of food-eating. In operant learning, although the topography that we see increase is lever pressing, ultimately it is food-related behavior that is increased due to deprivation. Hence, at any point at which the food-eating event is not accessible, the organism will behave with respect to the available (substitute) event having functional food-eating properties, in this case, lever-pressing. Lever pressing, just as approaching the food hopper, poking the head inside the hopper or foraging in the natural environment, constitutes behavior with respect to food. In other words, changes in response frequency with respect to the lever (event 1) reflect increments in the probability of occurrence of food-related behavior (event 2) in the sense that lever pressing occurs with respect to the food functions now inherent in the lever as a stimulus object.
In respondent learning, given states of food deprivation, a similar increment in responding to the bell may be observed in the absence of the food. If the animal were exposed to environmental conditions related to the bell-food contingency, it would be reasonable to speculate that the animal will hear the bell, or respond otherwise perceptually with respect to it, in its absence (Hayes 1992). It may also act to produce the sound of a bell if given the opportunity, as it is evidenced in autoshaping preparations (Brown and Jenkins 1968). Observing responses are a similar example of responding with respect to stimuli that have acquired properties of other events (see Dinsmoor 1983). The increase in the response strength of an observing response is due to the fact that the observing stimulus has acquired some of the properties of the reinforcer. Similarly, because the relevant function of the bell is related to the food-eating event, it is the food-eating event that changes in probability of occurrence; nonetheless, this increment will be observed with respect to the available event having food-eating properties.
To summarize, in both conditioning types, given a circumstance of deprivation of a biologically relevant stimulus and a contingency relation between two events, the second event will produce a change in the likelihood of responding to the first. Also in both cases, the experimental procedure consists of presenting the first event to the organism (the bell and the lever) and measuring food-related behavior in their presence. Distinctions between conditioning types result from focusing on the topographies observed and measured in respondent and operant conditioning as evidence of these conditioning processes. While classical conditioning measures changes in response topographies with respect to stimulus event 2 (food) in the presence of stimulus event 1 (neutral prior to conditioning), operant learning measures changes in the response topography with respect to stimulus event 1 (neutral prior to conditioning) due to its association with the food/eating event. Both experimental procedures evidence the acquisition of food/eating functions by the initially neutral event. However, this demonstration may be achieved in two different ways for the two learning types. In the presence of the initially neutral stimulus object (bell or lever), one could measure: a) a parameter of change in its correspondent response topography in relation to baseline levels (pressing the lever, hearing/orienting to the bell); and/or b) the occurrence of topographies corresponding to the physically absent stimulus object (food) (e.g., biting the lever, salivating in the presence of the bell).
Therefore, both conditioning processes constitute forms of demonstration of transfer of functions given a contingent relation between two events wherein behavior with respect to each present stimulus object may also include behavior related to the physically absent stimulus. Traditionally, operant conditioning has focused exclusively on procedure (a) and respondent conditioning on procedure (b). Thus, if it were necessary to make a distinction between conditioning types, it would be merely a procedural one. Basically the difference between these two cases would amount to the behavioral topography selected for measurement as evidence of behavior change.
It is likely that the sources of stimulation of the present stimulus object compete with the functional properties of the physically absent stimulus (Hayes and Delgado 2005). If so, factors such as deprivation levels, the biological significance of the stimulus, and the involvement of additional contextual functions may determine which behaviors will actually occur. However, much empirical support is required to test this formulation. Recent findings on Pavlovian models of cue competition may contribute to future research in this area.
The Issue of Symmetry
Bi-directionality of function transfer has been demonstrated by backward conditioning and respondent equivalence studies. In the present case, bi-directionality would be demonstrated by showing that the food/eating event also acquires some of the functions of the initially neutral stimulus after being exposed to a CS-US contingency or to a response-reinforcer contingency. As in forward transfer, this would be demonstrated in two ways: a) by detecting a change in some parameter of the food/eating topography; and/or b) by observing behavioral topographies corresponding to the formal properties of the initially neutral event, occurring in the presence of the food/eating event.
However, in both operant and respondent conditioning, the changes typically targeted for observation and measurement are those relative to behavior with respect to a biologically significant stimulus of which organisms are deprived. The fact that in traditional animal experiments one of the stimuli is typically a biologically significant stimulus (of which the organism is deprived) may obscure the demonstration of symmetrical transfer, as well as bias interpretations of processes and outcomes in favor of a one-way, predictive analysis of contingent relations.
The significance of investigating bidirectional relations is made apparent in classical conditioning research with humans. In these studies, the deprivation requirement is replaced by an explicit request to make a selection or produce a response. For example, research in respondent-type equivalence has shown repeatedly that symmetrical relations can be obtained when participants are exposed to sets of stimulus pairs (Clayton and Hayes 2004; Delgado and Medina 2011; Leader et al. 1996; Smeets et al. 1997; Tonneau and González 2004).
On the basis of the available empirical data on backward conditioning and respondent equivalence, we argue that bi-directional relations could be present (at least to some extent) in all cases of conditioning. Imagine a scenario where an animal is introduced into an operant chamber after exposure to an FR1 schedule. If the feeder drops a food pellet upon entrance to the chamber, the most likely behavior to follow the consumption of the pellet will be approaching and pressing the lever. If food deprived, even in the physical absence of the lever the organism is likely to engage in exploratory behavior, including approaching the place where the lever was, or simply engaging in perceptual behavior with respect to the lever in its absence. This could be construed as the occurrence of behavioral topographies with respect to the lever in the presence (not necessarily physical) of the food. The symmetrical substitutive relation is likely in the respondent example as well. After repeated occurrences of bell-food pairings, perceptual responses such as hearing or seeing the bell in its absence, or orienting towards the bell, are likely in the presence of the food (Hayes 1992).
Another indication of responding to the first stimulus event in the presence of the food/eating event corresponds to prompting appropriate behavior in the presence of the reinforcer. The reasoning behind this practice is that the presence of the reinforcer constitutes the occasion in which the occurrence of the target behavior is followed by access to the reinforcer. More precisely said, the target behavior is likely to occur because some of its functions are actualized through the presence of the reinforcing event.
Further, human research in respondent conditioning has shown that under certain circumstances, second order conditioning effects are observed even when both CSs (e.g., names of foods) are presented simultaneously (and not successively) in the second phase of the procedure; that is, when the US (e.g., an illness) is absent (Karazinov and Boakes 2007). In the natural environment, learning rarely occurs in discrete segments of time or always in the same order of event presentation whereby a one-way predictive relation would be established. Therefore, it is more likely that substitution effects are bi-directional and that they have a measurable effect on both events involved in the contingency. As mentioned, several studies in backward conditioning have undertaken the task to investigate the conditions under which these effects are manifested.
Evidently, additional research focusing on the conditions that facilitate bidirectional transfer is needed. Other noteworthy steps in this direction are the demonstrations of symmetrical responding by pigeons using conditional discrimination procedures reported by Zentall et al. (1992) and Sherburne and Zentall (1995), and the experiments on excitatory backward conditioning (Barnet and Miller 1996; Chang et al. 2003; Cole and Miller 1999; Hearst 1989; Hemmes et al. 1994; Spetch et al. 1981, 1982). Testing for a higher likelihood of either of the associated events, given equal conditions of deprivation and a comparable biological relevance, is also a possibility for future research. As we have noted, although only one event is traditionally measured in any instance of conditioning, it is unlikely that substitution of functions occurs exclusively from the second event to the first.
Summary and Conclusion: A Single Conditioning Process
Function transfer was defined by Sidman (2000) as the emergence of relations that had not been trained, or to which the organism had not been previously exposed (see also Sidman 2009). As such, it has been described as a property of stimulus equivalence and explained in terms of reinforcement contingencies (Sidman 2000). However, the notion of function transfer has been pivotal in classical conditioning research since the early work of Pavlov on the conditioned reflex and all throughout the development of classical conditioning accounts of complex human behavior (Glautier 2002; Karazinov and Boakes 2007; Rescorla 1988; Shanks 2010; Walther 2002).
By briefly describing some of the theoretical and empirical findings in the classical conditioning arena, we have shown that what the operant literature has heralded as a novel and difficult to explain phenomenon (Sidman 2009) has been systematically researched in the Pavlovian literature across different species and with different procedures, albeit with interpretations based on a different conceptual repertoire. These findings seem to suggest that bidirectional and transitive relations are not a product of reinforcement contingencies as the operant literature asserts (Tonneau 2001; Tonneau and González 2004; Tonneau and Sokolowski 1997).
Particularly relevant to this argument is the discussion of backward conditioning findings, as they are directly related to the issue of bidirectionality of event relations. Barnes-Holmes et al. (2001), and Tonneau (2001, 2002) have debated the significance of classical conditioning models, and particularly of findings from backward conditioning studies as support (or not) for an S-S account of derived relations. Following up on Tonneau’s efforts, we presented arguments and research findings in support of his views, and as the basis upon which a single process approach may be considered. In doing so, we attempted to develop a conceptual logic that unfolds into an integration of learning paradigms. It is founded on two main formulations.
The first one specifies that when considering the stimulus and response functions of each of the events involved in a contingency (whether conceptualized as operant or respondent), we are left with contingent associations between pairs (or sets) of events that are not as different as the incomplete descriptions of these relations seem to suggest (i.e., S-S, R-S, R-R, or even S.R-S).
In the second formulation, we argue that the outcomes of all conditioning processes can be explained by substitution of stimulus functions and we discuss the circumstances in which this process may act bi-directionally. In our view, due to substitution of functions, the probability of responding with respect to any of the events involved in a conditioning process increases with the presence of one or more members of that contingency. In other words, the outcome does not leave any of the stimulus–response events unchanged; and hence, the assumption of a particular directionality to the substitutive process cannot be sustained.
The description of the circumstances under which a particular behavior occurs is significantly broadened by considering that the following properties of conditioning are applicable to all events in the contingency: first, the occurrence of behavior is not limited by the physical presence of stimulus events; and second, a behavioral topography may occur with respect to the stimulus functions of an absent stimulus object inhering in a present stimulus object.
These characteristics apply to both operant and respondent learning. In fact, there don’t seem to be any features characteristic of respondent outcomes that are not also present in operant outcomes and vice-versa. On the basis of this assumption we propose a single-event perspective wherein substitution of functions is understood to be the fundamental process involved in all cases of conditioning.
As we see it, the only distinction between operant and respondent learning is that respondent learning includes—but is not limited to—the conditioning of reflexive responses. However, several studies have shown empirical evidence of reflexive behaviors being operantly conditioned (e.g., Lukowiak et al. 1996; Levene et al. 1968; Ley 1999).
The more biologically dependent responses are, the more limited their fields of interaction, and thereby, their range of variation. Because measurements and procedures are typically adjusted to the nature of the behavior of interest, procedural differences regarding the examination of simple and complex responses may be warranted. As events become more complex or less biologically determined, however, differences in procedures and measurements become less plausible as reasons for distinguishing among conditioning types. We argue that the use of different methods of observation does not justify classifying the objects of observation as different types of phenomena. Neither are different methods of observation indicative of separate types of basic processes (Pear and Eldridge 1984; Timberlake 2004; Wetherington 1982).
In many cases, experimental data have been difficult to classify as respondent or operant. Advocates of each learning process have conducted experiments that demonstrate phenomena as belonging to one category or the other. For example, little consensus has been reached with respect to the classification of autoshaping, conditioned avoidance, conditioned suppression, schedule-induced behavior, observing behavior, concept formation, and more recently, stimulus equivalence (Clayton and Hayes 2004; Fraley 2003; Rehfeldt and Hayes 1998).
These confusions have generated a number of reactions. The most common is the conciliatory view that respondent and operant processes are intertwined (Skinner 1953; Azrin et al. 1967). Other possibilities include reducing one category to the other, resorting to mentalistic explanations of the phenomena, proposing to relax the boundaries and conditions of each paradigm in order to fit all those events that do not seem to conform to the established requisites of each (Pear and Eldridge 1984), suggesting a third type of behavior/process (e.g., schedule induced behavior and equivalence), or as a last resort, maintaining traditional assumptions and neglecting the challenges imposed upon them by more recent empirical data.
More importantly, no major differences between the two types of learning with regard to a variety of principles and concepts have been demonstrated. To illustrate, pivotal principles of behavior science such as extinction, spontaneous recovery, stimulus discrimination, stimulus generalization and notions of primary and secondary reinforcers are described in both the respondent and the operant literature. The notion of reinforcer as an “after the fact” incontrovertible concept, or as Schoenfeld (1995) put it, “a reinforcer is as a reinforcer does” is virtually equivalent in operant and respondent process descriptions. The fact that the same set of fundamental laws are present in what are still described as differentiable types of learning processes constitutes additional support for the necessity of formulating an integrative theory of conditioning.
With respect to operant reinforcement, we contend that understanding the nature of substitutive operations may add to the possibilities of designing effective reinforcement-based procedures, provided that reinforcement is understood as a particular arrangement of experimental operations (Schoenfeld 1995), and not as a unique type of learning process.
In sum, we propose substitution of functions as a fundamental psychological principle by which the outcomes of contingency relations may be explained. These outcomes may be understood in terms of bidirectional relations between two or more stimulus–response interactions. As a result of such relations, it could be assumed that all participating events may change in two possible ways when in the presence of a particular stimulus object. One of these changes pertains to the frequency of the behavioral topography originally corresponding to a particular stimulus source which, after conditioning, will occur as a response to some of the properties of the absent event inhering in the present event. The second possible change corresponds to the occurrence of response topographies originally corresponding to the absent stimulus object. As a result of this reasoning, we conclude that substitution of functions constitutes a psychological principle that multiplies the functional properties of stimulus objects and response topographies.
The advancement of a science of human behavior may be enhanced by the development of a theory capable of accounting for all of the relevant phenomena (Kantor 1970). In our view, understanding psychological events in terms of substitution of functions may be a first step in the construction of a unified theory of conditioning. If substitution is as fundamental a process as we take it to be, the principles of Pavlovian conditioning should apply to all relations of the psychological variety. In other words, sensory pre-conditioning, second order conditioning and cue competition effects, for example, may also be relevant to reinforcement contingencies. Whether empirical data support or invalidate our formulations, we hope that they will offer research alternatives that may foster progress towards a more comprehensive account of human behavior.
Notes
Although Pavlovian studies often use meditational constructs in their models and interpretations of outcomes, S-S relations can be approached from a strictly naturalistic standpoint, that is to say, without appealing to mental processes. We refer to S-S associations from this latter standpoint. More specifically, we refer to conditions of association as they pertain to spatio-temporal relations among events in an integrated field (Kantor 1924).
References
Azrin, N. H., Hutchinson, R. R., & Hake, D. F. (1967). Attack, avoidance and escape reactions to aversive shock. Journal of the Experimental Analysis of Behavior, 10, 131–148.
Barnes-Holmes, D., Hayes, S. C., & Roche, B. (2001). The (not so) strange death of stimulus equivalence. European Journal of Behavior Analysis, 2, 35–41.
Barnet, R. C., & Miller, R. R. (1996). Second-order excitation mediated by a backward conditioned inhibitor. Journal of Experimental Psychology: Animal Behavior Processes, 22, 279–296. doi:10.1037//0097-7403.22.3.279.
Brown, P. L., & Jenkins, H. M. (1968). Auto-shaping of the pigeon’s key-peck. Journal of the Experimental Analysis of Behavior, 2, 1–8.
Chang, R., Blaisdell, A. P., & Miller, R. R. (2003). Backward conditioning: Mediation by the context. Journal of Experimental Psychology: Animal Behavior Processes, 29, 171–183. doi:10.1037/0097-7403.29.3.171.
Clayton, M. C., & Hayes, L. J. (2004). A comparison of match-to-sample and respondent-type training of equivalence classes. The Psychological Record, 54, 579–602.
Cole, R. P., & Miller, R. R. (1999). Conditioned excitation and conditioned inhibition acquired through backward conditioning. Learning and Motivation, 30, 129–156. doi:10.1006/lmot.1998.1027.
De Houwer, J., Beckers, T., & Glautier, S. (2002). Outcome and cue properties modulate blocking. Quarterly Journal of Experimental Psychology, 55, 965–985. doi:10.1080/02724980143000578.
Delgado, D., & Hayes, L. J. (2007). The acquisition of a conceptual repertoire: An analysis in terms of substitution of functions. The Behavior Analyst Today, 8, 307–316.
Delgado, D., & Hayes, L. (2011). Using the operant MTS procedure as a masking task for respondent acquisition of stimulus classes. Revista Latinoamericana de Psicología, 43, 13–25.
Delgado, D., & Hayes, L. J. (2013). The integration of learning paradigms by way of a non-causal analysis of behavioral events. Conductual, 1(2), 39–54.
Delgado, D., & Medina, I. F. (2011). Efectos de dos tipos de entrenamiento respondiente sobre la formación de clases de equivalencia. Revista Mexicana de Análisis de la Conducta, 37, 33–50. doi:10.5514/rmac.v37.i1.19475.
Delgado, D., Medina, I. F., & Soto, J. S. (2011). El lenguaje como mediador de la transferencia de funciones: ¿Es necesario nominar para relacionar? Revista Mexicana de Análisis de la Conducta, 37, 31–52. doi:10.5514/rmac.v37.i2.26138.
Denniston, J. C., Miller, R. R., & Matute, H. (1996). Biological significance as a determinant of cue competition. Psychological Science, 7, 325–331. doi:10.1111/j.1467-9280.1996.tb00383.x.
Dinsmoor, J. A. (1983). Observing and conditioned reinforcement. Behavioral and Brain Sciences, 6, 693–728.
Dixon, M. R., & Hayes, L. J. (1999). A behavioral analysis of dreaming. The Psychological Record, 49, 613–627.
Fraley, L. E. (2003). A behaviorological analysis of adjunctive behavior. Behaviorology Today, 6, 15–29.
Fryling, M. J., & Hayes, L. J. (2010). An interbehavioral analysis of remembering. European Journal of Behavior Analysis, 11, 53–68.
Glautier, S. (2002). Spatial separation of target and competitor cues enhances blocking of human causality judgments. The Quarterly Journal of Experimental Psychology, 55B, 121–135. doi:10.1080/02724990143000207.
Hall, G. (1996). Learning about associatively activated stimulus representations: Implications for acquired equivalence and perceptual learning. Animal Learning and Behavior, 24, 233–255.
Hayes, L. J. (1992). Equivalence as a process. In S. C. Hayes & L. J. Hayes (Eds.), Understanding Verbal Relations (pp. 97–108). Reno: Context Press.
Hayes, S. C., & Brownstein, A. J. (1986). Mentalism, behavior-behavior relations, and a behavior analytic view of the purposes of science. The Behavior Analyst, 9, 175–190.
Hayes, L. J., & Delgado, D. (2005). Transgenic and knockout models: The problem of language. In G. Fisch & J. Flint (Eds.), Transgenic and knockout models of neuropsychiatric disorders (pp. 45–67). Towanta, NJ: Humana.
Hayes, L. J., & Delgado, D. (2006). Animal models of non-conventional human behavior. Behavior Genetics, 37(1), 11–17.
Hayes, L. J., & Fryling, M. J. (2010). Overcoming the pseudo-problem of private events in the analysis of behavior. Behavior and Philosophy, 37, 39–57.
Hayes, S. C., Barnes-Holmes, D., & Roche, B. (2001). Relational frame theory: A post-Skinnerian account of human language and cognition. New York: Kluwer Academic/Plenum.
Hearst, E. (1989). Backward associations: Differential learning about stimuli that follow the presence versus the absence of food in pigeons. Animal Learning & Behavior, 19, 280–290. doi:10.3758/BF03209801.
Hemmes, N. S., Brown, B. L., & Cabeza de Vaca, S. (1994). Appetitive backward conditioning in pigeons. The Psychological Record, 44, 221–237.
Kantor, J. R. (1924). Principles of Psychology (Vol. 1). New York: Knopf.
Kantor, J. R. (1959). Interbehavioral Psychology: A Sample of Scientific System Construction. Bloomington, IN: Principia Press.
Kantor, J. R. (1970). An analysis of the Experimental Analysis of Behavior (TEAB). Journal of the Experimental Analysis of Behavior, 13, 101–108.
Karazinov, D. M., & Boakes, R. A. (2007). Second order conditioning in human predictive judgments when there is little time to think. Quarterly Journal of Experimental Psychology, 60, 448–460. doi:10.1080/17470210601002488.
Leader, G., & Barnes-Holmes, D. (2000). Establishing equivalence relations using a respondent type training procedure III. Psychological Record, 50, 63–78.
Leader, G., Barnes, D., & Smeets, P. M. (1996). Establishing equivalence relations using a respondent-type training procedure. The Psychological Record, 46, 685–706.
Levene, H. I., Engel, B. T., & Pearson, J. A. (1968). Differential operant conditioning of heart rate. Psychosomatic Medicine, 30(6), 837–845.
Ley, R. (1999). The modification of breathing behavior: Pavlovian and operant control in emotion and cognition. Behavior Modification, 23(3), 441–479.
Lukowiak, K., Ringseis, E., Spencer, G., Wildering, W., & Syed, N. (1996). Operant conditioning of aerial respiratory behavior in Lymnaea Stagnalis. The Journal of Experimental Biology, 199, 683–691.
Mackintosh, N. J. (1974). The psychology of animal learning. New York: Academic Press.
Mahan, H. C. (1968). The Interactional Psychology of J. R. Kantor: An Introduction. San Marcos, CA: Project Socrates Press.
Midgley, B. D., & Morris, E. K. (Eds.). (2006). Modern perspectives on J. R. Kantor and interbehaviorism. Reno, NV: Context Press.
O’Donohue, W., & Ferguson, K. E. (2001). The Psychology of B. F. Skinner. Thousand Oaks, CA: Sage Publications.
Olson, M. A., & Fazio, R. H. (2002). Implicit acquisition and manifestation of classically conditioned attitudes. Social Cognition, 20, 89–103. doi:10.1521/soco.20.2.89.20992.
Parrott, L. J. (1986). The role of postulation in the analysis of unapparent events. In H. W. Reese & L. J. Hayes (Eds.), Behavior science: Philosophical, methodological and empirical advances (pp. 35–60). Hillsdale, NJ: Erlbaum.
Parrott, L. J. (1987). On the distinction between setting events and stimuli. Experimental Analysis of Human Behavior Bulletin, 5(1), 6–11.
Pawlik, K. (1997). To the memory of Ivan Petrovich Pavlov (1849-1936). European Psychologist, 2, 91–96. doi:10.1027/1016-9040.2.2.91.
Pear, J. J., & Elridge, G. D. (1984). The operant-respondent distinction: Future directions. Journal of the Experimental Analysis of Behavior, 42, 453–467. doi:10.1901/jeab.1984.42-453.
Premack, D. (1965). Reinforcement Theory. In D. Levine (Ed.), Nebraska symposium on motivation. University of Nebraska Press: Lincoln.
Rehfeldt, R. A., & Hayes, L. J. (1998). The operant-respondent extinction revisited: Toward an understanding of stimulus equivalence. The Psychological Record, 48, 187.
Rehfeldt, R. A., Dixon, M. R., Hayes, L. J., & Steele, A. (1998). Stimulus equivalence and the blocking effect. The Psychological Record, 48, 647–664.
Rescorla, R. A. (1988). Pavlovian conditioning is not what you think it is. American Psychologist, 43, 151–160.
Savastano, H. I., & Miller, R. R. (2004). Behavioral momentum and Pavlovian conditioning. Behavioral and Brain Sciences, 27, 694–695. doi:10.1017/S0140525X04230163.
Schoenfeld, W. N. (1995). Encore. “Reinforcement” in Behavior Theory. The Behavior Analyst, 18, 173–185.
Shanks, D. R. (2010). Learning: From Association to Cognition. Annual Review of Psychology, 61, 273–301. doi:10.1146/annurev.psych.093008.100519.
Sherburne, L. M., & Zentall, T. R. (1995). Delayed matching in pigeons with food and no-food samples: Further examination of backward associations. Animal Learning & Behavior, 23(2), 177–181. doi:10.3758/BF03199932.
Sidman, M. (1994). Equivalence relations and behavior: A research story. Boston: Authors Cooperative.
Sidman, M. (2000). Equivalence relations and the reinforcement contingency. Journal of the Experimental Analysis of Behavior, 74, 127–146. doi:10.1901/jeab.2000.74-127.
Sidman, M. (2009). Equivalence relations: An introductory tutorial. The Analysis of Verbal Behavior, 25(1), 5–17.
Sidman, M., & Tailby, W. (1982). Conditional discrimination vs. matching to sample: An expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior, 37, 5–22.
Silva, F. J., Timberlake, W., & Koehler, T. L. (1996). A behavior systems approach to bidirectional excitatory conditioning. Learning and Motivation, 27, 130–150. doi:10.1006/lmot.1996.0008.
Silva, F. J., Timberlake, W., & Cevik, M. O. (1998). A behavior systems approach to the expression of backward associations. Learning and Motivation, 29, 1–22. doi:10.1006/lmot.1997.0987.
Skinner, B. F. (1953). Science and Human Behavior. New York: Macmillan.
Smeets, P. M., Leader, G., & Barnes, D. (1997). Establishing stimulus classes in adults and children using a respondent-type training procedure: A follow-up study. The Psychological Record, 47, 285–308.
Spetch, M. L., Wilkie, D. M., & Pinel, J. P. (1981). Backward conditioning: A reevaluation of the empirical evidence. Psychological Bulletin, 89(1), 163–175. doi:10.1037/0033-2909.89.1.163.
Spetch, M. L., Terlecki, L. J., Pinel, J. P. J., Wilkie, D. M., & Treit, D. (1982). Excitatory backward conditioning of defensive burying in rats. Bulletin of the Psychonomic Society, 19, 111–114. doi:10.3758/BF03330056.
Staats, A. W., & Staats, C. K. (1958). Attitudes established by classical conditioning. Journal of Abnormal & Social Psychology, 11, 187–192. doi:10.1037/h0042782.
Timberlake, W. (2004). Trends in the study of Pavlovian conditioning. International Journal of Comparative Psychology, 17, 119–130.
Timberlake, W., & Allison, J. (1974). Response deprivation: An empirical approach to instrumental performance. Psychological Review, 81, 146–164.
Tonneau, F. (2001). Equivalence relations: A critical analysis. European Journal of Behavior Analysis, 2, 1–128.
Tonneau, F. (2002). Who can understand Relational Frame Theory? A reply to Barnes-Holmes and Hayes. European Journal of Behavior Analysis, 3, 95–102.
Tonneau, F., & González, C. (2004). Function transfer in human operant experiments: The role of stimulus pairings. Journal of the Experimental Analysis of Behavior, 81, 239–255. doi:10.1901/jeab.2004.81-239.
Tonneau, F., & Sokolowski, M. B. C. (1997). Standard principles, nonstandard data, and unsolved issues. Journal of the Experimental Analysis of Behavior, 68, 266–270. doi:10.1901/jeab.1997.68-266.
Tonneeau, F., Arreola, F., & Martínez, A. G. (2006). Function transformation without reinforcement. Journal of the Experimental Analysis of Behavior, 85, 393–405. doi:10.1901/jeab.2006.49-05.
Walther, E. (2002). Guilty by mere association: Evaluative conditioning and the spreading attitude effect. Journal of Personality and Social Psychology, 82, 919–934.
Walther, E., Nagengast, B., & Trasselli, C. (2005). Evaluative conditioning in social psychology: facts and speculations. Cognition and Emotion, 19(2), 175–196. doi:10.1080/02699930441000274.
Wetherington, C. L. (1982). Is adjunctive behavior a third class of behavior? Neuroscience and Biobehavioral Reviews, 6, 329–350. doi:10.1016/0149-7634(82)90045-8.
Wilson, P. N., & Alexander, T. (2008). Blocking of spatial learning between enclosure geometry and a local landmark. Journal of Experimental Psychology: Learning, Memory & Cognition, 34, 1369–1376. doi:10.1037/a0013011.
Zentall, T. R., Sherburne, L. M., & Steirn, J. N. (1992). Development of excitatory backward associations during the establishment of forward associations in a delayed conditional discrimination by pigeons. Animal Learning & Behavior, 20, 199–206. doi:10.3758/BF03213373.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Delgado, D., Hayes, L.J. An Integrative Approach to Learning Processes: Revisiting Substitution of Functions. Psychol Rec 64, 625–637 (2014). https://doi.org/10.1007/s40732-014-0071-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40732-014-0071-6