
Abstract
Four pigeons were studied under a resurgence procedure in which, in successive phases, 1) pecking the left response key (the target response) was reinforced while responding on the right response key (the alternative response) was not reinforced; 2) both responses were extinguished; 3) the alternative response was reinforced while the target response continued to be extinguished; and 4) the target response continued to be extinguished while unsignaled delays of reinforcement that progressively increased across several sessions were added to the alternative response key. As the delays progressively reduced alternative response rates and reinforcement rates, rates of pecking on the originally trained, but now extinguished, key-peck response recurred (resurged) in three of four subjects. The present findings suggest that resurgence is not restricted to conventional extinction where reinforcement is completely eliminated. They also suggest a means of controlling resurgence in the absence of access to the behavioral history that is the wellspring of that resurgence.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Resurgence typically is defined as the recurrence of a previously reinforced response when the currently reinforced response is extinguished. Resurgence studies often employ a four-phase procedure wherein: (1) a history of response A is established while a second recorded response, B, has no programmed consequence; (2) response A then is placed on extinction and extinction continues for response B; (3) response B is reinforced and extinction continues for response A; and (4) both responses are placed on extinction. Higher response A rates in the fourth relative to the third phase is evidence of resurgence. A three-phase variation of this procedure omits phase 2. Response A rates in the final phase are typically transient, with response rates increasing to a peak and then approaching zero across the sessions. Resurgence is theoretically interesting because of its implications for understanding the dynamics of extinction and behavioral history (Lieving and Lattal 2003), and of applied interest because of its role in the recurrence of problem behavior (e.g., Lieving et al. 2004).
Although the extinction of the recently reinforced response (response B) generally results in resurgence of the previously reinforced response (response A), little is known about how other variables operating on response B affect resurgence. Wilson and Hayes (1996) found that when response B was punished, response A increased, but Lieving and Lattal (2003) found only weak resurgence when the schedule of reinforcement of response B was changed from variable-interval (VI) 30 s to VI 360 s. Lieving and Lattal’s findings have since been replicated in both college students and a child with behavioral problems (Marstellar and St. Peter, 2012). By contrast, one manipulation that degrades responding but does not induce resurgence is eliminating the response-reinforcer dependency. When Lieving and Lattal changed the schedule of reinforcement for response B from a VI to an otherwise equivalent variable-time (VT) schedule in which the reinforcers were delivered independently of responding, there was no resurgence of response A.
Demonstrations of resurgence under conditions of degraded reinforcement and of punishment suggest that resurgence may be more general than originally proposed. Lieving and Lattal’s (2003) failure to observe resurgence when the response-reinforcer dependency of the schedule maintaining response B was eliminated, however, suggests limitations on the degraded reinforcement conditions under which resurgence occurs. Delay of reinforcement reduces responses similarly to response-independent reinforcement, and to almost the same levels at very long delays (Gleeson and Lattal 1987). Given the findings of Lieving and Lattal with respect to response-independent food delivery and resurgence, it was of interest to examine whether delay of reinforcement was a sufficient condition for developing resurgence.
Method
Subjects
Four White Carneau pigeons were maintained at 80 % of free-feeding weights. All had experimental histories on different reinforcement schedules. Water and health grit were freely available in the home cages, located in a vivarium where a 12 hr-12 hr light-dark cycle was maintained. All of the current procedures were in accordance with the guidelines established by the West Virginia University Institutional Animal Care and Use Committee.
Apparatus
A sound-attenuating operant conditioning chamber (31 cm wide, 29.5 cm long, and 32 cm high) contained an aluminum panel housing two 2.5 cm Gerbrands response keys. The keys were 4 cm apart and equidistant from the midline, with their lower edges 27 cm from the chamber floor. They could be transilluminated white. A rectangular aperture (4.5 cm wide by 6 cm high) was located on the midline of the panel, with its base 7.5 cm from the floor. The aperture provided 3-s access to mixed grain when a hopper was raised. A 28-V DC clear bulb illuminated the aperture. All other lights were dark during presentations of the hopper for reinforcer deliveries. White noise delivered through a speaker located above the chamber masked extraneous noise. Programming and data recording were controlled in an adjacent room by an IBM-compatible personal computer operating MED-PC® software and hardware (MED Associates, Inc. and Tatham 1991).
Procedure
Because the pigeons had extensive histories of responding on other schedules of reinforcement, pretraining was omitted. Sessions occurred 6-7 days a week at approximately the same time each day and, except in the response elimination phase, ended after either 30 reinforcers or 300 s without a key peck, whichever came first. There were four phases. The number of sessions for each pigeon in each phase are described below.
Reinforcement Phase
Pecking the left key (response A) resulted in a 3-s access to grain according to a VI 30-s schedule; all VI schedules consisted of 10 intervals generated as described by Fleshler and Hoffman (1962). Right-key (response B) pecks were without effect (extinction). This phase lasted minimally 15 sessions and until left-key pecking stabilized, i.e., when over the last six sessions the mean response rates of the first and last three sessions did not deviate from the grand mean by more than +/−4 % and without monotonic trends in those rates. This phase lasted 29 sessions for Pigeon 449, 18 sessions for Pigeon 545, 17 sessions for Pigeon 723, and 16 sessions for Pigeon 888.
Response Elimination Phase
Pecks to both keys were without effect (extinction). Unlike the other phases, which terminated after 30 reinforcers or a pause of 300 s, sessions in this phase lasted for a set duration equal to the duration of the final session of the previous phase. This duration was chosen in an effort to roughly equalize session duration across the phases and to allow sufficient time for extinction to occur. This phase continued until a session passed without a peck on either key. This phase lasted 9 sessions for Pigeon 449, 10 sessions for Pigeon 545, 7 sessions for Pigeon 723, and 9 sessions for Pigeon 888.
Alternative Reinforcement Phase
Pecks to the left key were without effect (extinction). Pecks to the right key were reinforced on a VI 30-s schedule. This phase continued for at least 13 sessions and ended when right-key response rates stabilized by the definition given above. This phase lasted 17 sessions for Pigeon 449, 13 sessions for Pigeon 545, 14 sessions for Pigeon 723, and 15 sessions for Pigeon 888.
Resurgence Phase
Progressively increasing unsignaled delays in reinforcement were arranged on the right key across successive sessions. Thus, pecks to the right key were reinforced under a tandem VI 30-s FT x-s schedule, where x increased across successive sessions until responding ceased completely for 300 s; at that point the resurgence testing phase was terminated. The delay sequence (s) was 0.5, 1, 5, 10, 20, 40, 80, 160, 320, and 640. Using this break-point criterion resulted in maximum delays of 640 s for 449, 320 s for 545, 40 s for 723, and 320 s for 888. Pecks to the left key were without effect (extinction).
Results
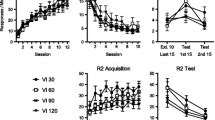
Figure 1 shows each pigeon’s response rates on each key through the phases. During the reinforcement phase, responding was almost exclusive to the left key, associated with the VI schedule. This responding decreased to near zero across sessions during the response elimination phase. During the alternative reinforcement phase, responding was almost exclusive to the right key. During the resurgence phase, responding on the right key systematically decreased as the delays increased for Pigeons 449, 545, and 888, and responding on the left key (resurgence) increased as a function of the decreasing responding on the right key. This delay-related increase in response rates on the target response key was most consistent for Pigeons 449 and 545. Pigeon 723’s response rates on the target response key during the progressive delay contingency actually increased for a few sessions before decreasing. Nonetheless, 723 also eventually showed resurgence of left-key responding.

Overall rates of responding for the left (A) and right (B) key during the final six sessions of the Reinforcement and Extinction phases, and for all sessions of the Alternative Reinforcement and Resurgence phases
Figure 2 shows the same effects described for Fig. 1, but with response rates during the resurgence phase expressed as a percentage of the mean of the last six sessions of the alternative reinforcement phase. Points above the horizontal dotted line (i.e., proportion of baseline > 1) indicate resurgence. Resurgence occurred during 7 of 9 sessions for 545, 5 of 6 sessions for 723, 3 of 9 sessions for 888, and 3 of 10 sessions for 449. Thus resurgence, as defined here, was most consistent for Pigeons 545 and 723. In general resurgence occurred more frequently at longer delays than shorter delays.

Response rates on both keys during the Resurgence phase transformed to a proportion of mean response rates for the last six sessions of the Alternative Reinforcement phase. Note: where the proportion of baseline was 0, data are plotted as 0.01
Discussion
Resurgence reliably occurred when reinforcement was delayed from the reinforced response for two (545 and 723) of the four pigeons tested here, and less reliably for a third pigeon (449). Although this effect was only reliably seen in two of the four pigeons, the present resurgence was more consistent and persistent than that reported by Lieving and Lattal (2003) and Marstellar and St. Peter (2012) when they decreased the rate of reinforcement of the alternative response. Their findings along with Wilson and Hayes’s (1996) demonstration of resurgence with punished alternative responding suggest that resurgence is not restricted to the conventional extinction of the alternative response. This said, however, the resulting resurgence under conditions of diminished reinforcement may be less than that obtained when reinforcement of the alternative response is eliminated altogether.
The reason delays of reinforcement resulted in resurgence in the current study, however, is somewhat unclear. The functional effect of progressively increasing delays of reinforcement was to progressively decrease response rates (except for Pigeon 723) and reinforcement rates (all pigeons). The relation between these two variables and resurgence was mixed, but in general, resurgence was more likely with longer delays, meaning lower response rates and reinforcement rates. Lieving and Lattal (2003, Experiment 3) found that response rate reductions alone were insufficient to produce consistent resurgence. Even though Lieving and Lattal (2003, Experiment 4) found that reducing reinforcement rates produced weak resurgence relative to that found when the alternative response was conventionally extinguished, resurgence was more likely during longer interreinforcer intervals (their Figure 8). As delays in reinforcement progressively lengthen, the periods of nonreinforcement defining the delay also lengthen, and it is those periods that seem to induce resurgence. Hence, the resurgence observed with decreased reinforcement rates may be a function of local periods of nonreinforcement, i.e., local extinction, cf. Lieving and Lattal (2003, Experiment 4). Previous studies, however, have controlled for these decreased reinforcement rates and still obtained systematic decreases in response rates. For example, in contrasting a variation of the present inter-session progression of successive delay durations with a condition wherein the rate of immediate reinforcement similarly increased across sessions, Jarmolowicz and Lattal (2013) found that decreases in the reinforcement rate did not systematically decrease response rates when the delay was not in effect. Although Jarmolowicz and Lattal examined response rate changes in relation to delays of reinforcement, rather than resurgence, the procedural similarities between the studies suggest a similar mechanism (i.e., decreases in response-reinforcer contiguity) underlying Jarmolowicz and Lattal’s response rate decreases and the present response rate decrease/resurgence. Hence, changes in the temporal interval between the reinforced response and the next reinforcer seemed largely responsible for the decreases in response rates observed with increasing delay durations, and thus to the resurgence obtained.
Generating resurgence via contemporary variables associated with the alternative response, such as punishment or delay of reinforcement, may stimulate research directly relevant to application. For example, practitioners intervening with behavior problems may have considerable control over the current environment, but with little access to the client’s history. Hence information regarding the contemporary circumstances under which resurgence will and will not occur may be particularly useful. For example, Lieving and Lattal’s (2003) finding that resurgence is related to local periods of nonreinforcement suggests that maintaining rich schedules of reinforcement may attenuate resurgence of problem behavior (e.g. Podlesnik et al. 2006; Lieving et al. 2004).
A recent review of schedule-thinning procedures used following functional communication training (Hagopian et al. 2011) suggested that the reemergence of problem behavior as schedules of alternative communication are thinned may be an instance of resurgence. As such, the present results suggest that because delays of reinforcement generate resurgence, they should be minimized or avoided during treatment regimens. This recommendation is consistent with Hanley, Iwata, and Thompson’s (2001) observation that the reemergence of problem behavior (e.g., self-injury) was more likely when schedules of appropriate communication were thinned by adding a delay of reinforcement than with other techniques (e.g., multiple schedules), and with Hagopian et al.’s (2011) observation that half of all published applications of delays of reinforcement for schedule-thinning have added additional contingencies (e.g., punishment; Hagopian et al. 1998) to maintain low rates during schedule-thinning.
References
Fleshler, M., & Hoffman, H. S. (1962). A progression for generating variable-interval schedules. Journal of the Experimental Analysis of Behavior, 5, 529–530.
Gleeson, S., & Lattal, K. A. (1987). Response–reinforcer relations and the maintenance of behavior. Journal of the Experimental Analysis of Behavior, 48, 383–393.
Hagopian, L. P., Fisher, W. W., Sullivan, M. T., Acquisto, J., & LeBlanc, L. A. (1998). Effectiveness of functional communication training with and without extinction and punishment: A summary of 21 inpatient cases. Journal of Applied Behavior Analysis, 31, 221–235.
Hagopian, L. P., Boelter, E. W., & Jarmolowicz, D. P. (2011). Reinforcement schedule-thinning following functional communication training: Review and recommendations. Behavior Analysis in Practice, 4, 4–16.
Hanley, G. P., Iwata, B. A., & Thompson, R. H. (2001). Reinforcement schedule-thinning following treatment with functional communication training. Journal of Applied Behavior Analysis, 34, 17–38.
Jarmolowicz, D. P., & Lattal, K. A. (2013). Delayed reinforcement and fixed-ratio performance. Journal of the Experimental Analysis of Behavior, 100, 370–395.
Lieving, G. A., & Lattal, K. A. (2003). Recency, repeatability, and reinforcer retrenchment: an experimental analysis of resurgence. Journal of the Experimental Analysis of Behavior, 80, 217–233.
Lieving, G. A., Hagopian, L. G., Long, E. S., & O’Conner, J. (2004). Response-class hierarchies and resurgence of severe problem behavior. The Psychological Record, 54, 621–634.
Marstellar, T., & St. Peter, C. C. (2012). Resurgence during treatment challenges. Mexican Journal of Behavior Analysis, 38, 7–23.
MED Associates, Inc. & Tatham, T.A. (1991). MED-PC Medstate notation. East Fairfield: MED Associates, Inc.
Podlesnik, C. A., Jimenez-Gomez, C., & Shahan, T. A. (2006). Resurgence of alcohol seeking produced by discontinuing non-drug reinforcement as an animal model of drug relapse. Behavioural Pharmacology, 17, 369–374.
Wilson, K. G., & Hayes, S. C. (1996). Resurgence of derived stimulus relations. Journal of the Experimental Analysis of Behavior, 66, 267–281.
Acknowledgments
The authors thank Sally Huskinson for her help conducting this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jarmolowicz, D.P., Lattal, K.A. Resurgence Under Delayed Reinforcement. Psychol Rec 64, 189–193 (2014). https://doi.org/10.1007/s40732-014-0040-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40732-014-0040-0