
Abstract
To attain sustainable agricultural crop protection, tools such as host plant resistance, enhanced ecosystem services (i.e. conserving natural enemies) and the deployment of companion plants should be promoted in pest management programmes. These agro system manipulations could be based on chemical ecology studies considering the interactions with natural enemies and pests, regarding specifically plant defence signalling. Further, new crop protection strategies might rise from widening the knowledge regarding how herbivore-induced plant volatiles can govern a multifaceted defence response including natural enemy recruitment, pest repellence or induced defence in neighbouring plants. It is crucial to use a multitrophic approach to understand better the interactions involving companion plants, herbivores and natural enemies in the field, increasing the knowledge to build more efficient and sustainable pest management strategies. In this review, we explore the perspectives of companion plants and their semiochemicals to promote conservation biological control according to the ‘smart plants’ concept. Further, we discuss the advantages and disadvantages of using companion plants and explore the application of companion plants in different agroecosystems using several case studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Plant Diversity and Biological Control
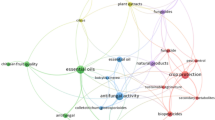
Increasing the diversity of plants in agroecosystems has been studied extensively in recent decades as a strategy for promoting conservation biological control of invertebrate (insect, mite) pests (e.g. Sheehan 1986; Andow 1991; Poveda et al. 2008; Letourneau et al. 2009; 2011; Isbell et al. 2017). Here, more suitable resources and conditions for pest natural enemies are provided (Venzon et al. 2019b), and the need for external inputs, e.g. pesticides, is reduced (Shields et al., 2019). However, the effectiveness of this strategy has met with variable levels of success (Letourneau et al. 2009; 2011). Although general positive effects on pest management have been observed in different systems, there are examples where increasing plant diversity in agroecosystems can lead to difficulties for natural enemies (mostly specialists) to locate their prey (Letourneau et al. 2011). For example, plant diversity can in some instances promote antagonistic interactions, such as intraguild predation or apparent competition, which can reduce the impact of natural enemies on pest populations (Venzon et al. 2001; Koss and Snyder 2005; Janssen et al. 2007). Plant diversity can also increase unfavourable conditions for natural enemies (Corbett and Plant 1993). Additionally, some plants may provide additional resources to phytophagous insects, increasing their attraction and damage to the main crop (Norris and Kogan 2000). Therefore, increasing plant diversity per se is sometimes insufficient to reduce pest populations, and there is a need to better understand the ecological interactions underlying this management strategy (Venzon et al. 2019a; 2019b) (Fig. 1).

General overview of ecological interactions in crops systems with increased plant diversity by introduction of companion plants (left) and botton-up and top-down effects on herbivorous populations. The relevance of identification of semiochemicals mediating these interactions to develop smart plants to strengthen bottom-up and top-down effects
Although ecological interactions that operate in natural ecosystems are also present in agroecosystems, interactions in the latter are often characterised by continuous disturbance due to agriculture intensification. Agroecosystems can have high species turnover, frequent introduction of germplasm with low genetic variability, disturbance of natural communities caused by synthetic pesticides and continuous exportation of biomass from the system through harvesting (Nicholls and Altieri 2004). To understand better the ecological interactions operating in agroecosystems, it is necessary to elucidate main factors that affect interactions within guilds and between trophic levels (Venzon et al. 2001). Most interactions related to pest management may involve cropped plants (e.g. main and secondary crops), natural vegetation, weeds, herbivorous pests, non-pest herbivores and natural enemies (Venzon et al. 2019b). In such food webs, bottom-up (a lower trophic level influences a higher trophic level) and top-down (a higher trophic level influences a lower trophic level) effects can directly and indirectly operate simultaneously upon populations of insect pests (Fig. 1).
Achieving a better understanding of specific interactions in food webs is an important first step for manipulating plant diversity and enhancing pest management (Begg et al. 2017). Moreover, it is known that semiochemicals (naturally occurring behaviour-modifying compounds, e.g. herbivore-induced plant volatiles (HIPVs)) play a central role in mediating the interactions between plants, pests and their natural enemies. Understanding the chemical ecology of plant-pest-natural enemy interactions, specifically the role of inducible or primed host semiochemicals in plant resistance, has advanced rapidly in recent years, thereby increasing the use of semiochemicals in pest management (Birkett and Pickett 2014) (Fig. 1). Therefore, improving our understanding of the synergies between the use of companion plants and the interactions mediated by semiochemicals could stimulate a new generation of conservation biological control strategies.
In this review, we seek to explore the use of companion plants and their associated semiochemicals to promote conservation biological control within the framework of ‘smart plants’, i.e. selected plant varieties that present traits allowing them to respond earlier to pest colonisation (Birkett and Pickett 2014). We believe that smart plant deployment is an important innovation for sustainable pest management, as it is based on the manipulation of ecological processes rather than relying predominantly on deployment of pesticides. We present (i) a theoretical discussion on the background related to companion plants, semiochemicals and smart plants, in order to find possible complementary effects between their use in agriculture, and (ii) successful case studies, with a primary focus on experiments carried out in Brazil. Finally, we provide a future perspective on opportunities for research in conservation biological control and possible routes for innovation in agricultural food production.
From a bottom-up effect perspective, phytophagous insects use a wide array of signals and stimuli from different distances and sources to locate and colonise a host plant (Bernays and Chapman 2007). At long distances, insects mostly use olfactory and visual signals to orient towards an appropriate habitat containing their host plants (Borges and Blassioli-Moraes; 2017; Prokopy et al. 1983), and then at a short distance, plant traits start to play an important role (Borges and Blassioli-Moraes 2017; Prokopy et al. 1983).
In a more complex environment with a variety of plants (i.e. other than the target crop), visual cues may be difficult for phytophagous insects to exploit in differentiating between host and non-host plants, and therefore may waste time and energy in finding a host (Dethier et al. 1960). Therefore, it is expected that tritrophic interactions that rely on chemical volatile information could be more advantageous in complex environments. However, a mixed blend of plant odours promoted by plant diversity in agroecosystems could also act as a repellent stimulus (Robinson et al. 2008; Ratnadass et al. 2012), thereby causing individuals to move to other habitats or patches. Additionally, plant volatiles from non-preferred or non-host plants can produce a masking effect of the cues from the host plant, thus producing an associational resistance effect (Thiery and Visser 19871986). This phenomenon is more restricted to polyphagous insects, where decision-making ability is less accurate than that of specialist insects, possibly because they need to deal with more complex and diverse information in the environment to benefit from the exploitation of different hosts (Bernays and Minkenberg 1997). There is a certain neural limitation to integrate information and make choice in complex environments over a simpler environment (Bernays 2001). According to Rosado et al. (1973) and their Resource Concentration Hypothesis, monoculture crops are easier to be found than polycultures. Phytophagous insects tend to colonise and be retained in such areas because they find resources that are more suitable and conditions ideal for survival and reproduction.
At shorter distances, for phytophagous insects to be retained in the habitat, chemical (including volatile and non-volatile chemicals), physical and visual stimuli can be used. Finch and Collier (2000) summarised the theoretical ecology involved in host plant selection at short distance. The choice of a host consists in accumulating several positive stimuli (e.g. olfactory, tactile and visual) when landing on different plant parts after arriving in a patch within the habitat. When an insect consistently gains more positive stimuli, when probing a host plant several times, it can decide to use the plant as a host, i.e. start egg laying. However, when non-preferred or non-host plants are present during the acceptance phase, an insect can deal with multiple negative stimuli, including barriers to movement and as well as mechanical plant defence against feeding. After balancing the negative and positive stimuli, when plant diversity is present, phytophagous insects tend to move to another area. Therefore, plant diversity in agroecosystems can affect host plant selection and insect retention.
Natural enemy top-down effects, in a given crop location, are engendered by species that can effectively colonise hosts and be retained within the crop field (Tscharntke et al. 2005a, b). Some habitats in the landscape such as natural vegetation areas (Rand et al. 2006), fallow areas (Sarthou et al. 2014; Togni et al. 2019a; 2019b), agroforestry systems (Harterreiten-Souza et al. 2014; 2021; Rezende et al. 2014) and field margins nearby the crop (Amaral et al. 2016) can act as sources of natural enemy species where they can spill over to the cropped area (Tylianakis et al. 2005; Tscharntke et al. 2012). Consequently, the effects of plant diversity on natural enemies can depend on spatial variables, often at the landscape level. As most of the strategies related to the increase of plant diversity are connected to farm scale level, we will therefore focus our discussion on the attraction and retention of natural enemies inside the crop area.
According to the natural enemies’ hypothesis, increasing plant diversity in agroecosystems raises the abundance of natural enemies, as there are more resources and conditions to harbour larger populations of different species in the area (Root 1973; Andow 1991). As a consequence, there is a higher mortality of phytophagous insects (Root 1973). However, some predictions created by this hypothesis are not always confirmed (e.g. Andow 1991; Letourneau et al. 2011; Rosenheim et al. 2016). Thus, a multitrophic approach should be adopted to increase the positive effects of plant diversification in agroecosystems. Focusing on how different natural enemy species can co-exist in the same area, and increase the population regulation of phytophagous insects, is a key factor for conservation biological control research.
The attraction of natural enemies to crops depends on cues related to the provision of suitable resources and conditions (Begg et al. 2017). The attractiveness of a crop can be increased by provision of edible resources (pollen and nectar), favourable conditions (shelter for eggs) and plant diversity (Togni et al. 2016; Venzon et al. 2019a). HIPVs can be used as a reliable signal to attract natural enemies (Dicke et al. 1990; Karban and Baldwin 1997). However, retention of natural enemies is not guaranteed. For example, some parasitoids can be attracted to a given crop by its volatiles, but they need first to gain energy from food sources, such as pollen and nectar, to go after their hosts. Nectar, for example, can be present in floral and extrafloral nectaries that can selectively attract natural enemy species (Rezende et al. 2014), due to differences in the nutritional quality and accessibility to different species (Patt et al. 1997). Thus, introducing plants with edible pollen and suitable nectar, including spontaneous vegetation, can increase the attraction, retention and efficiency of natural enemies in controlling pest populations (Simpson et al. 2011; Venzon et al. 2019b). As more species of natural enemies are attracted to a given habitat, multiple negative interactions, such as intraguild predation and apparent competition, are also prone to occur with negative effects on biological control (Janssen et al. 2007). Therefore, specific plant traits should be considered when introducing or managing plant diversity to avoid such interactions. Moreover, alternative prey, such as non-pest insects, can increase the retention, and survival, of natural enemies (Biondi et al. 2016). Besides the provision of food, plant diversity can improve the provision of shelter, and microclimatic conditions, to natural enemies (Gontijo 2019).
When resources are not limited to natural enemy species, there is no competition within, and between, species allowing their coexistence in the same habitat. Plant architecture, and shelters provided by increasing plant diversity, can also reduce the rate of encounter between individuals from the same guild and reduce intraguild predation due to changes in the habitat structure (Janssen et al. 2007; Gontijo 2019).
Moreover, plant diversity can increase the movement of phytophagous insects within and amongst plants in agroecosystems because the habitat has a more complex architecture and the presence of non-host species make the individuals move. Consequently, phytophagous insects tend to be more exposed and vulnerable to predation by different species (Straub et al. 2014). All the positive effects of plant diversity on natural enemies should depend on the knowledge about the phenology synchrony between populations of pests and natural enemies dispersing in different crops at a regional/landscape scale (Begg et al. 2017).
Development of the Smart Plant Concept for Agroecosystems
Besides physical and visual effects, plant semiochemicals are probably the most important factors mediating the interactions and processes described above, in particular HIPVs. The use of HIPVs and other semiochemicals can be an alternative, or complement, to increase diversity in agroecosystems. HIPVs have the potential to be exploited in biological control of agricultural insect pests, either as direct attractants for natural enemies, inducers of crop defence to increase their attractiveness to natural enemies, targets for breeding or genetic engineering of crop plants, or as targets for companion cropping (Turlings and Erb 2018). Attempts to use HIPVs for natural enemy recruitment have been increasingly explored in recent years with some success, e.g. companion cropping for management of pests on cereals (Pickett and Khan 2016).
The concept of using smart plants that can respond to insect damage by rapid HIPV emission and recruitment of natural enemies from natural habitats has been described previously (Birkett and Pickett 2014) (Fig. 1). A number of underpinning studies have given a platform for developing the smart plant concept, by providing an understanding of the role of plant defence signalling in plant/insect/natural enemy interactions, the smart defence response trait in crop genotypes, the deployment of companion plants alongside smart main crops for semiochemical-based crop protection and genetic engineering of a crop plant to produce a natural enemy recruitment cue (Pickett and Khan 2016; Khan et al. 2014; Birkett and Pickett 2014; Bruce et al. 2015; Tamiru et al. 2011). In a further development of the smart plant concept, HIPV emission can be exploited in the design of sensitive companion (‘sentinel’) plants for rapid, early and reliable detection of insects that are hard to detect before recognisable symptoms appear. Here, HIPV emission from sentinel plants is a target for airborne plant/plant communication, where the sentinel plant induces defence in smart crops that have been tailored or selected to respond rapidly to the emitted HIPVs, for the recruitment of natural enemies (Birkett and Pickett 2014).
For successful development of the smart plant concept in pest management, hypothesis-driven science, together with field experiments, is needed to meet the challenge of upscaling companion plant concepts into intensive agricultural systems and inform the next steps in development of smart plants for inclusion in the deployment of sentinel-based cropping systems. Furthermore, experimental farm platforms are needed to test the viability of smart crop/sentinel plant cropping systems in managing pest populations. Trap crops, nurse crops or cover crops, as well as flowering field margins, are required to investigate for sentinel plant effects. Research is needed to address key questions relating to (i) planting arrangements, density and proportion of sentinel plant vs main crops and (ii) the ability of the main crop to respond to sentinel plant signals under field conditions.
Semiochemicals and HIPVs have the potential to be exploited in biological control of agricultural pests and can also be considered direct indicators of early plant stress events (e.g. insect colonisation). The strategic value of developing an accurate early warning system to aid with crop pest management is of vital importance to the global economy. With current crop management practices, around 30–40% of crop yield worldwide is lost to pests (Savary et al. 2019). Having accurate and reliable information on when to apply pesticides will reduce environmental impact (Barzman et al. 2015) and allow target precision application of crop protection tools. The idea of exploiting plant response to stress volatile organic compounds (VOCs) fits well with the smart plant concept where known VOCs can be biomarkers for real-time sensor technologies (plant as biosensors) (Volkov and Ranatunga 2006).
Biotic and abiotic stress factors significantly impact crop production worldwide. Mitigation strategies work effectively when there is a known problem. However, the problem is not always visible. Cryptic pests, disease-infected plants and abiotic factors cannot be seen in crops until there are visual symptoms at which point it may be too late for effective mitigation. The exploitation of plant VOCs as indictors of stress will allow detection at earlier time points, as plants react to stresses which results in changes to VOC profiles (potential biomarkers) (Kollist et al. 2019).
Over the last three decades, volatile analysis technology has rapidly moved away from traditional laboratory-based GC and GCMS instrumentation to non-invasive real-time analysis systems, e.g. proton-transfer-reaction mass spectrometry and selective ion flow tube mass spectrometry (Niederbacher et al. 2015; Materić et al. 2015). These techniques have allowed real-time analysis of plant volatile emission, thus providing an understanding of diurnal cycles in VOC emission, and changes in VOC emissions as a result of biotic or abiotic stress conditions. However, a major limitation to these technologies is that they are laboratory-based systems. VOC gas analysers (based on electronic nose sensor devices), which have developed rapidly over the last 3 decades, offer a solution to this limitation, by enabling rapid monitoring of VOCs at the source of emission (Cui et al. 2019). Such analysers cannot discriminate between compounds within a headspace sample. Sensor designs can be optimised for particular groups of compounds but still cannot truly distinguish between compounds, as can be achieved with mass spectrometry-based systems. However, with advanced statistical models, the analysis of whole head space samples can be categorised and give indications of plants stress (cryptic herbivore infestation and disease status). This sensor technology offers the ability to quantify VOC changes rapidly on location, and, if needed, traditional MS methods can be used to identify the compounds of significance.
As with all sensor technologies, problems with sensitivity and efficient sample collection provide barriers to widespread acceptance in agricultural settings (Cellini et al. 2017). For successful deployment in pest management, hypothesis-driven science and proof-of-concept field testing are required to fully characterise crops and smart sentinel plants under different biotic and abiotic stressors. Next-generation advanced statistical models, artificial intelligence (AI) identification and discrimination between stressors, can be used to create digital libraries to help with farm management decisions on when and where to use crop protection tools.
Case Studies
Companion and Smart Plants to Manage Pest in Maize Crops
Maize, Zea mays L. (Poaceae), is one of the most important cultivated plant worldwide (Ranum et al. 2014). One of the biggest constraints to improving the yield of maize crops in smallholder and family-run farms is the high number of pests attacking maize, mainly herbivores and weeds. In Brazil, maize plants are attacked by several insect pests both aboveground and belowground (Moreira and Aragáo 2009), with the fall armyworm, Spodoptera frugiperda Smith, 1797 (Lepidoptera: Noctuidae), being considered the primary pest in Brazil (Cruz et al. 2010). Farmers have few alternatives to manage these pests other than intensive insecticide applications, which are usually not fully effective because of the development of insecticide resistance (Lira et al. 2020). Use of Bt maize in Brazil has significantly reduced lepidopteran populations and insecticide application (Farias et al. 2014). However, this technology is still not accessible to smallholder and family-run farms due to its high cost. In addition, failure of Bt maize to control fall armyworm populations has been reported (Farias et al. 2014; Bernardi et al. 2015).
The International Centre of Insect Physiology and Ecology (icipe) successfully developed the use of companion plants to protect maize crops in Sub-Saharan countries against lepidopteran pests, based on plant semiochemicals in a push–pull system. The target species to be controlled were the exotic stemborer Chilo partellus Swinhoe, 1885 (Lepidopetra: Crambidae), the indigineous Busseola fusca Fuller, 1901 (Lepidoptera: Noctuidae) and the weed Striga hermonthica Benth. (Orobanchaceae). Napier grass Pennisetum purpureum Schumach. (Poaceae) was selected as a trap plant from a group of natural hosts of the borers (Schulthess et al. 1997). Desmodium uncinatum Jacq.DC (Fabaceae) was used as the push plant, as it repels the herbivores and reduces emergence of S. hermonthica in maize fields. Chemical ecology studies showed that Napier grass and maize plants release a blend of green leaf volatiles (hexanal, (E)-2-hexenal, (Z)-3-hexen-1-ol, (Z)-3-hexen-1-yl acetate) that attracts the herbivores. However, Napier grass produces higher levels of these compounds in the first hour of nightfall, when lepidopterans actively forage for host plants. On the other hand, D. uncinatum produces a blend of repellent volatiles including (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT), (E)-ocimene, (E)-caryophyllene, α-humulene and α-terpinolene. In addition, studies reported that D. uncinatum flowers are attractive to the natural enemy, Cotesia sesamiae Cameron 1906 (Hymenoptera: Braconidae). Finally, the striga weed is controlled by a complex mechanism involving different isoflavonones released by the roots of D. uncinatum (Khan et al. 2008, 2007).
Another alternative to manage lepidopteran pests on maize could be to explore the genetic variability of different cultivars that can influence the secondary metabolites that are involved in insect-plant interactions. Michereff et al. (2019) showed that from six different neotropical maize genotypes, only one genotype, i.e. Sintetico Spodoptera (SS), produced a specific blend of HIPVs after S. frugiperda feeding damage. This HIPV blend attracts its natural enemy, the egg parasitoid Telenomus remus (Nixon, 1937) (Hymenoptera: Platygastridae). The study suggests that two factors could be involved in this natural enemy attraction, i.e. the higher concentration of the compounds in the SS genotype and the ratio between the compounds in the HIPV blend. The SS maize genotype appears to have a great potential as a sentinel and smart plant, since it produces higher level of HIPVs, and this genotype is primed by HIPVs released by neighbouring plants, triggering earlier and stronger defence against herbivory of S. frugiperda (Michereff et al. 2021).
Recently, several field studies have reported that the stink bugs D. melacanthus and D. furcatus move from harvested soybean to seedling maize. A potential trap crop for these pests is pigeon pea, Cajanus cajan (Millsp.) (Fabaceae). Nymphs of D. melacanthus are not able to develop when fed with pigeon pea pods (Borges and Blassioli-Moraes unpublished data). To test the potential effect of this trap crop and other companion plants (CP) as border plots on maize insect pests and natural enemy diversity, a field experiment with four different treatments was set up in small (25 m2) plots. The treatments were as follows: (1) maize as a monocrop, (2) maize with C. cajan, (3) maize with P. purpureum and (4) maize with Crotalaria spectabilis Roth. (Fabaceae). Plots with maize and C. spectabilis as the border crop showed a smaller number of plants with severe injury from S. frugiperda compared to monocrop maize, C. cajan maize plot and to P. purpureum/maize.
Other studies have shown a negative effect on S. frugiperda population in maize when cultivated with Crotalaria sp. The mechanisms underlying the effects of these CP with maize are still to be determined. Therefore, more laboratory studies are required to understand these interactions, and more field studies are necessary to establish the use of CPs in maize crops on smallholder and family-run farms in Brazil.
Companion and Smart Plants to Manage Stink Bugs in Soybean Crops
Stink bugs are oligophagous or polyphagous insects that use a wide variety of plants belonging to more than 10 families (Panizzi and Lucini 2017; Esquivel et al. 2018). Host plants, i.e. plants where the insects feed and can complete their development, are decisive for stink bug biology. It has been demonstrated that alternative hosts, i.e. plant species where the insects feed but do not complete full development, could have a complementary and fundamental role in stink bug biology. These alternative plants offer a substitute and complementary source of food that may positively influence their population dynamics (Panizzi and Slansky 1991; Panizzi and Saraiva 1993; Panizzi and Lucini 2017; Velasco and Walter 1993; Silva et al. 2018). As most of the plants used by stink bugs are either annual or semi-perennial herbs and bushes, these resources are not continuous in both space and time, so stink bug ecology is characterised by a continuous displacement between areas with cultivated plants, and areas with native or spontaneous vegetation (Panizzi 1997). This switching between more and less preferred plants has been exploited to develop trap crops as a stink bug management strategy. Trap cropping consists of the use of an attractive plant species to arrest the insects and reduce the colonisation of crop fields (Hokkanen 1991).
In the Nearctic region, trap crop systems have been successfully implemented for stink bug management in both organic and conventional cultures. Many plants in the Brassicaceae, e.g. white mustard, Sinapis alba (L.) and black mustard, Brassica nigra (L.), the Fabaceae e.g., pea, Psium sativum (L.), Vicia villosa Roth, crimson clover, Trifoliumin carnatum L. and soybean Glycine max (L.), the Poaceae eg sorghum, Sorghum bicolor (L.) and pearl millet, Pennisetum glaucum (L.) R.Br., sunflower, Helianthus annuus L. (Asteraceae) and buckwheat, Fagopyrum esculentum Moench (Polygonaceae) have been used successfully to manage Nezara viridula (Linnaeus, 1758), Euschistus servus (Say, 1832) and Chinavia hilaris (Say, 1832) stink bugs (Bundy and McPherson 2000; Rea et al. 2002; Tillman 2006; Mizell et al. 2008; Tillman and Cottrell 2012). However, despite these successful examples of stink bug management and the wide knowledge of host and alternative plants in the Neotropical region (Panizzi and Silva 2012; Panizzi and Lucini 2017), trap cropping has been not used with soybean until now in cultivated areas.
Egg parasitoids (Hymenoptera: Scelionidae) are the main natural enemies of stink bugs. In the Neotropical region, there is a high diversity of these native parasitoids (Cingolani 2012) which provides an opportunity to develop augmentative biological control programmes. However, because of the difficulty in rearing these insects and their high mobility and susceptibility to insecticides (Corrêa-Ferreira et al. 2010), conservation biological control using natural populations of parasitoids is likely a better option. A common approach to promote conservation biological control is to use attract-and-reward strategies, which allows the spill over of parasitoids attracted by semiochemicals to be managed. Here, additional resources such as shelter and food (nectar) via flowering plants are offered (Simpson et al. 2011). Some semiochemicals have shown their potential to attract stink bug parasitoids in the laboratory and field, e.g. plant volatiles (Blassioli-Moraes et al. 2005; 2008; Michereff et al. 2015; 2016), host sex pheromones (Borges et al. 1998) and host defensive compounds (Laumann et al. 2009). Plant volatiles are of particular interest because of their potential to underpin the development of smart plants with enhanced volatile production through breeding and genetic engineering. However, although the application of semiochemicals under field conditions increases the recruitment of egg parasitoids, it does not increase parasitism rate (Vieira et al. 2013; 2014), probably because attraction alone is insufficient to increase the performance of parasitoids. The use of flowering plants in combination with semiochemicals could be an efficient way to solve this problem. Foti et al. (2017) have shown the beneficial effect of some flowering plants, especially buckwheat and sweet basil, Ocimum basilicum (L.) (Lamiaceae), to increase the offspring of the egg parasitoid Trissolcus basalis (Wollaston, 1858) (Hymenoptera: Scelionidae). The authors also showed that flower volatiles of buckwheat are attractive to the parasitoid. In a 2-year experiment, it was demonstrated that strips of buckwheat cultivated in combination with tomato plants increase the parasitism by T. basalis in N. viridula eggs. In contrast, this plant appears to repel another egg parasitoid, Ooencyrtus telenomicida (Vassiliev, 1904) (Hymenoptera: Encyrtidae). This shows that flowering plants, specifically flower volatiles, could mediate intra guild competition favouring some species of egg parasitoids (Foti et al. 2019), an aspect that needs to be considered for the design of attract-and-reward strategies. Similar beneficial effects of buckwheat flowers have been observed for the Neotropical parasitoid Telenomus podisi. In laboratory experiments, it was demonstrated that feeding on buckwheat increases the survival and fecundity of the parasitoid and that buckwheat volatiles are attractive to T. podisi females. In preliminary field test plots of soybean with buckwheat as a border plant, higher parasitism indices were observed compared to control plots (without buckwheat), and no negative effect on the parasitoid community structure was observed (Aquino, Blassioli-Moraes, Borges and Laumann, unpublished data).
Companion Plants as Reservoirs for Natural Enemies in Organic Crops
The use of companion plants can enhance the efficiency and reliability of conservation biological control in organic farms. In Brazil, crops such as garlic, onion, lettuce, tomatoes, cabbage/collards, cucumber and sweet pepper have been grown in association with African marigold, Tagetes erecta (L.), pot marigold, Calendula sp. (Asteraceae), coriander, Coriandrum sp., dill, Anethum sp. and anise, Pimpinella sp. (Apiaceae), basil (Ocimum sp.), forage turnip, Raphanus sp. (Brassicaceae) and spontaneous plants (weeds) as companion plants.
In a crop system including onions and African marigold (Silveira et al. 2009), the proximal (until 5 m) and distant (30 m) effects of the companion plants were assessed. The results showed that phytophagous populations, mainly Thrips tabaci Lindeman, 1889 (Thysanoptera: Thripidae), were twofold greater in onion plants distant from the companion plant, and that entomophagous species were more abundant in onion plants close to the companion plants. In another experiment with lettuce as the main crop instead of onions, it was demonstrated that African marigold plants could contribute to natural enemy abundance in lettuce beds until distances ranged between five and 6 m (Zaché 2009).
In another experiment where African marigold (a single line 1.7 m long) were installed across a lettuce field, it was demonstrated that this arrangement was more suitable for spreading natural enemies across a whole field (Haro 2015; Haro et al. 2018). The proximity of the companion plants mediated shifts in the food webs, since they became stronger and more intricate when close to African marigold (Haro et al. 2018).
The effect of companion plants on arthropod food web composition and attractiveness of natural enemies appears to be influenced by volatiles from essential oils of plants. Haro (2015) found that oils of African marigold, extracted from leaves and flowers, resulted in positive olfactory responses of various natural enemies as the parasitoids Aphidius colemani Viereck, 1912 (Hymenoptera: Braconidae), Aphelinus abdominalis (Dalman, 1820) (Hymenoptera: Aphelinidae) and Encarsia formosa Gahan, 1924 (Hymenoptera: Aphelinidae). Also, the predators Adalia bipunctata (Linnaeus, 1758) (Coleoptera: Coccinellidae) and Orius laevigatus (Fieber, 1860) (Hemiptera: Anthocoridae) responded positively to the presence of these oils.
The association of African marigold with tomato also led to reduced herbivorous injury on tomato plants (Haro 2011). In a greenhouse experiment, enhanced tomato production (ca 30%) was observed when tomatoes were accompanied by African marigold. The production increase was likely mediated by various factors, but mainly by the reduction of important pests. Thrips, aphids and leaf miners did not reach pest threshold levels in the African marigold treatment, whereas for tomato monocultures, herbivore threshold levels were reached during experiments (Haro 2011).
These data clearly suggest that African marigold can be used as a companion plant in horticulture with benefits to natural enemies and conservative biological control. Other species also have potential to fulfil the same role, and when comparing the effects of African marigold with C. officinalis (also an Asteraceae species), and the Apiaceae species dill (A. graveolens) and coriander (C. sativum), incidence of the aphid Lipaphis erysimi was significantly lower in the kale-African marigold system when compared with all other treatments (Silva et al. 2016). One of the possible reasons for this was aphid predation and parasitism; the kale-African marigold system resulted in a higher incidence of predators in comparison with the other treatments, whilst the number of parasitized aphids was highest in the kale-African marigold and kale-calendula systems.
Members of the Apiaceae, i.e. coriander, dill and fennel (Foeniculum vulgare Mill.), were studied to determine the assemblage of predators associated with them (Resende et al. 2012). Fennel showed the higher diversity of species (Shannon H’), whilst coriander was more abundant in important species such as Orius insidiosus (Say, 1832) (Hemiptera: Anthocoridae), an important thrips predator.
Other species of vegetable pest predators were found in all Apiaceous species, especially adults and larvae of coccinellids, which are important aphid predators. Also, the flowers of Apiaceae species can serve as sources of pollen for predators as adults of Chrysoperla externa Hagen, 1861 (Neuroptera: Chrysopidae) and larvae and adults of Eriopis connexa (Germar, 1824) (Coleoptera:Coccinellidae) as shown in Resende et al. (2015, 2017). Larvae and adults of ladybeetles were shown to survive in the presence of only coriander flowers, whilst adult chrysopids may survive and reproduce up to the third generation by feeding only on resources provided by flowers of dill, coriander and fennel.
Haro et al. (2015) studied the parasitoids of important horticultural pests associated to these Apiaceae. The authors found higher richness and abundance in coriander and dill and also important parasitoids of pest species in fennel. Therefore, parasitoids and predators could contribute to the biological control of pests when Apiaceous species are used as companion plants.
Basil, a plant used for cooking and medicinal purposes, has also good potential to preserve natural enemies, especially parasitoids (Souza et al. 2019). Parasitoid diversity was studied in organic sweet pepper associated with basil and African marigold, and it was reported that sweet pepper monoculture, sweet pepper-basil intercropping and sweet pepper-marigold intercropping hosted 40, 98 and 130 individuals, respectively, with similar richness of species between them. Furthermore, the insects of greater abundance in basil and marigold were different to those collected in the monoculture, resulting in distinct groups of similarities amongst the treatments, as shown in the non-metric multidimensional scaling (NMDS) analysis and with significant similarity analysis (Anosim). Therefore, the number of parasitoids increased in the associations of sweet pepper with basil and marigold, providing advantages in the use of vegetable diversification for organic pepper production.
Companion Plants for Intercropping with Tomatoes in Organic Cropping Systems
In Costa Rica, Hilje and Stansly (2008) reported the reduction of the incidence and damage caused by the whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) on tomato crops when intercropped with coriander plants. The intercropping effects on pest incidence and damage were comparable to the use of synthetic insecticides, but with the advantage of returning an extra income to farmers for the sale of the coriander and ease of handling (Hilje and Stansly 2008).
In Brazil, this intercrop was adapted for managing whiteflies and tomato pinworm Tuta absluta (Meyrick, 1917) (Lepidoptera: Gelechiidae), mostly in organic tomato crops through bottom-up and top-down effects on these pest insects (Medeiros et al. 2009a; Togni et al. 2009). Coriander should be planted, intercropped with tomatoes, 15 days before the tomato seedlings are transplanted in the field. After 50–55 days, the coriander plants can be harvested, leaving about 6–10 plants to bloom between every pair of tomato plants. Coriander seeds are sown again so that, when the previous plants become senescent, new flowers will be available and the growers can harvest the coriander seeds (see details in Medeiros et al. 2009b; Togni et al. 2009).
Tuta absoluta oviposited approximately twice as many numbers of eggs on tomato plants in the conventional system than in the organic system, with no detectable effect from coriander plants in the conventional system (Medeiros et al. 2009a). In the organic system, the intercrop with coriander reduced the number of eggs of T. absoluta by 34% (Medeiros et al. 2009a) and the number of B. tabaci nymphs by 48% (Togni et al. 2009). Additionally, the abundance of natural enemies of both pest species was almost threefold higher in the organic system (Togni et al. 2009), and more species were found in the intercropped plots (Togni et al. 2010a).
These results suggest that bottom-up and top-down effects acted together in reducing the populations of B. tabaci and T. absoluta when coriander plants were added to the cropping system. However, each species responds differently to plant diversification and the cultural practices used. In a free-choice experiment in a greenhouse, T. absoluta exhibited a clear preference for laying eggs on tomato plants cultivated in soils from the conventional system (Medeiros et al. 2009c). For B. tabaci, coriander volatiles produce an odour masking effect on adults when presented together with tomato volatiles, making host choice more difficult and causing individuals to colonise areas with coriander plants (Togni et al. 2010b). Another cultural practice that can be integrated with the tomato and coriander intercrop system is the use of sprinkler irrigation, which causes mortality of T. absoluta eggs (Medeiros et al. 2011) as well as reduces B. tabaci colonisation and nymph settlement on plants in organic systems (Togni et al. 2018).
As the flowers of coriander are abundant with natural enemies able to access resources (pollen and nectar) easily (Patt et al. 1997), several natural enemy species can be attracted. Predators such as C. externa, Hippodamia convergens Guérin-Menéllive, 1842 (Coleoptera: Coccinellidae) and Cycloneda sanguinea (Linnaeus, 1763) (Coleoptera: Coccinellidae) can use the pollen of coriander plants as food sources, increasing their survival (Medeiros et al. 2010; Togni et al. 2016). Some predators such as C. sanguinea exhibit an innate attraction to coriander volatiles because these plants can be used as oviposition sites and shelter for the larvae (Togni et al. 2016). When coriander is intercropped with tomato plants, there is an increase in T. absoluta egg predation but no increase in the parasitism by Trichogramma spp. (Hymenoptera: Trichogrammatidae) was observed (Medeiros et al. 2011). Similarly, predation is a key mortality factor of B. tabaci populations and it is positively related with the abundance and richness of B. tabaci natural enemies (Togni et al. 2019c).
The tomato and coriander intercrop did not affect the yields of tomato plants in comparison with monocultured plots, even when it is irrigated by sprinklers (Togni et al. 2009; Marouelli et al. 2011). However, coriander plants can provide an extra income to the farmers and increase land use occupancy and efficiency (Hilje and Stansly 2008; Medeiros et al. 2009b). In conclusion, the increase in the vegetation diversity in organic tomato crops provided by coriander plants can favour positive interactions between natural enemies and the population regulation of pest insects and disrupt the host-plant interactions with direct benefits to farmers.
Companion Plants and Conservation Biological Control in Brassica Crops
Brassica spp. plants comprise the most economically important cruciferous group amongst the 51 genera in the tribe Brassicaceae (Gomez-Campo 1980). In general, brassicas are attacked by a variety of arthropod pests, especially aphids and lepidopterans.
Due to increasing demand for organic vegetables in Brazil (Oelofse et al. 2010) and the fewer options of insecticides registered for brassicas, more biological control studies have been warranted. In this sense, conservation biological control of brassica pests has been a continuous research effort in many parts of Brazil. Some related studies presented herein have been carried out in open fields, mostly at the Federal University of Viçosa, Campus Florestal.
In general, these studies have investigated how intercropping either flowering or non-flowering plants with brassicas can affect the conservation biological control of herbivorous pests such as aphids, thereby prompting applied and fundamental implications. The flowering species used in these studies has been alyssum Lobularia maritima (L. Desv.) (Brassicaceae), which forms a low-growing mat with high flower density, attracts several natural enemies and very few pests, and is unlikely to become a weed (Chaney 1998; Gontijo et al. 2013). In fact, this flower species has often been used as an insectary plant in the USA to enhance conservation biological control (Hogg et al. 2011; Gontijo et al. 2013; Brennan 2016).
Ribeiro and Gontijo (2017) investigated in Goias State how strip intercropping alyssum flowers with collards would influence the conservation biological control of multiple brassica pests, including aphids, diamondback moth larvae and whiteflies. This study showed that the attractiveness of alyssum flowers to generalist predators mediated a significant reduction of those pests, especially aphids. The abundance of generalist predators in this study was significantly higher in treatments associated with strips of alyssum, which likely served as a source of alternative non-prey food (i.e., pollen and nectar) and shelter. In a similar study, Silva et al. (2021) investigated whether the intraspecific variation of general morphology in a mixture of brassicas (i.e. collards + broccolis + cabbages) could influence lepidopteran and aphid infestation/suppression. Additionally, in some treatments, alyssum plants were added to the brassica mixture to assess if the interspecific variation of multiple plant traits (i.e. plant morphology and flower availability) could further enhance natural enemy activity and pest suppression. The results suggest that adding alyssum flowers to fields planted with either monoculture or mixture of brassicas was important to enhance aphid suppression. Moreover, whilst the results indicate that brassica mixture (i.e., variation of single plant trait) was enough to promote predator abundance, it was not able to drive pest suppression alone. In this specific study, collards associated with alyssum also had higher leaf fresh weight as well as higher number of marketable leaves per plant. Taken together, these results indicate promoting the interspecific variation of distinct plant traits can engender stronger pest suppression than the intraspecific variation of single plant traits in regard to promoting biological control.
Gontijo et al. (2018) studied how non-flowering parsley intercropped with collards would affect different guilds of natural enemies (diurnal and nocturnal) and consequently the conservation biological control of aphids. The results showed that the absence of flowers on the companion plants was not detrimental in attracting and sheltering generalist predators, which contributed significantly to a reduction in the aphid population. Furthermore, this suggests that non-flowering plants may still contribute to conservation biological control by providing natural enemies with adequate microclimate and alternative prey/host. Nonetheless, this study showed that companion plants forming the low stratum can hamper the movement of non-flying natural enemies in the short term (e.g. nocturnal predators), and thus impair their ability to encounter pests on the focal host plants at the high stratum. Using a similar experimental design where non-flowering parsley was intercropped with collards, Saldanha et al. (2019) investigated how companion planting could affect natural infestation by aphids, natural enemy activity and parasitoid emergence. The results indicate that non-flowering parsley is important to shelter generalist predators that will form the first line of defence against aphids early in the season. Additionally, the effect of companion planting covaried with the on-plant aphid density to influence the numerical response from specialist natural enemies, which was weaker in the intercropping treatment. Although not detrimental to conservation biological control in the short term, companion planting appeared to mediate an increase in intraguild predation of immature aphid parasitoids. These results were likely due to companion plants promoting the breakdown of the prey dilution effect and making the mummified aphids more prone to predation.
Taken together, all these studies indicate that companion planting can be an important strategy to promote conservation biological control of pests. Despite some negative interactions mediated by companion planting (e.g. intraguild predation, hampering non-flying predators), the overall benefits of its implementation to enhance conservation biological control still outweigh those potential short-term adverse effects. The results also suggest that companion planting and natural shelter could be more important for generalist natural enemies, which are more likely to arrive sooner in the agroecosystem after crop emergence. Therefore, implementing efficient companion planting or natural shelter may entail methods that will ensure their field establishment prior to pest arrival. In respect to improving brassica production, advancing the conservation biological control of cruciferous pests is paramount to enhancing quality and safety for this group of vegetables, which is widely consumed and vulnerable to excessive insecticide applications.
Companion Plants and Conservation Biological Control in Coffee Agroecosystems
Coffee, which is cultivated mainly in tropical developing countries, is an important cash crop. Brazil is the leading global producer of coffee, and this crop has predominantly been cultivated in monocultures under full sun in the country. However, environmental and social harms caused by conventional agricultural practices have encouraged the development of other cultivated systems that aim to recover the soil, reduce erosion, improve nutrient recycling, enhance pest control and increase food security and sovereignty (Souza et al. 2012; Rezende et al. 2014; Gomes et al. 2020).
Increasing plant diversity in coffee agroecosystems can reduce damage caused by the main coffee pests (Rezende et al. 2014; Venzon et al. 2019a). Amaral et al. (2010) investigated in two organic coffee production systems the effects of plant diversification on the population coffee leaf-miner Leucoptera coffeella (Guérin-Mèneville, 1842) (Lepidoptera: Lyonettidae), one of the key coffee pests in Brazil. One system consisted of coffee intercropped with banana trees, Musa spp. (Musaceae) (shaded system) and the other consisted of coffee intercropped with pigeon pea, Cajanus cajan (Fabaceae) (unshaded system). The increase in plant diversity on both systems was achieved via introduction of cover crops, i.e. perennial peanut, Arachis pintoi Krapov. and W.C. Gregory, sun hemp, Crotalaria juncea L., and Brazilian lucerne, Stylosanthes guianensis. A positive and significant relationship between plant diversity and coffee leaf miner predation by wasps on the unshaded coffee system (without banana trees) was observed, whereas a negative relationship on the shaded coffee system found the opposite effect.
Further studies were carried out in order to select the most suitable cover crops to introduce into coffee agroecosystems, with the aim of selectively increasing natural enemy performance via provision of plant provided food. Venzon et al. (2006) evaluated the suitability of leguminous cover crop pollen to the green lacewing Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae), a common predator species in coffee agroecosystems (Ecole et al. 2002; Ribeiro et al. 2014). Both adults and larvae of C. externa can feed on plant-material, whilst larvae can feed on a variety of soft-bodied arthropod prey including the coffee leaf miner and coffee berry borer (Ecole et al. 2002; Carvalho et al. 2020; Botti et al. 2021). The presence of alternative plant food sources for lacewings is especially important in times of prey scarcity. Pollen of pigeon pea and sun hemp were equally suitable for C. externa, especially when they were complemented with a carbohydrate source (indicate here the type of carbohydrate used). The results suggest that to successfully enhance predator effectiveness, coffee crops should be diversified in a manner of combining plants that provide pollen with plants that provide nectar. In fact, laboratory (Rosado 2007) and field experiments (Rosado et al. 2021) confirm that when sun hemp, a pollen source, was associated with buckwheat, Fagopyrum esculentum Moench (Poligonaceae), a nectar and pollen source, an increase in coffee leaf miner parasitism and predation rate was observed.
Besides increasing diversity by adding cover crops, the introduction of trees in coffee agroecosystems can also provide pest control services. Trees play an important role within coffee agroforestry systems, providing resources to natural enemies that enhance natural regulation of coffee pests (Railsback and Johnson 2014; Rezende et al. 2014). Many trees selected for intercropping with coffee, as those of the genus Inga Miller (Leguminosae), possess extrafloral nectaries (Soto-Pinto et al. 2001; Souza et al. 2010) that are accessible and available during all stages of plant growth, thereby extending food availability (Wäckers 2005). Plants bearing extrafloral nectaries are known to suffer less from herbivory than plants lacking these structures (Mathews et al. 2007).
Rezende et al. (2014) showed that extrafloral nectaries of associated trees can boost natural pest control in agroforestry systems. Whilst species richness had no significant effect, leaf miner parasitism increased significantly with the abundance of nectary visitors, and the proportion of mined leaves decreased significantly with this abundance. The proportion of bored fruits decreased with increasing abundance of visitors, but this trend was not significant. Inga trees host an important natural enemy of coffee berry borer, Hypothenemus hampei (Ferrari, 1867) (Coleoptera: Scolytidae), i.e. predatory thrips of the genus Trybomia (Thysanoptera: Phlaeothripidae) (Rezende et al. 2014; Pantoja 2018).
Research has been carried out to select more plant species to be used in coffee agroecosystems (Venzon et al. 2018; 2019a), with the aim of reducing coffee pest populations using diverse strategies such as the use of ecological corridors and agroforestry (according to the production system). Understanding about the ecosystem services provided by individual plant species will help in unravelling the mechanisms which enhance pest control in diversified systems, and can also help in the design of pest-suppressive coffee systems (Rezende et al. 2021).
Plant Diversification as a Tool for Smallholder Farmers: a Successful Case for Perennial and Annual Crops
The development of a productive, resilient and affordable agricultural system is essential to assure smallholder farmers sustainability as well as maximise the economical, environmental and social long-term benefits. As plant diversification can also help mitigating the impact of climate change, the Santa Catarina State Agricultural Research and Rural Extension Agency (Epagri) has been encouraging the provision of multifunctionality in agricultural landscapes, ecosystem services and biodiversity for the cultivation of perennial and annual crops in the Itajaí River Valley (Lichtenberg et al. 2017; Rusch et al. 2016). Smallholder farmers in Itajaí River Valley grow mainly vegetables such as lettuce, arugula, tomatoes, cabbages, spices and aromatic plants, which are supplied to all coastal cities in that region. Usually, an excessive number of pesticides are used to control pests and diseases, potentially damaging these crops (Ngowi et al. 2007). However, there is a growing concern about food safety and environmental contamination, due to the proximity of these cropping fields to water sources and urban areas (Huang et al. 2006).
As part of a response to those concerns, some researchers have suggested the intercropping of vegetables with African marigold, Tagetes erecta L. (Asteraceae), which has been shown in previous studies to enhance pest suppression without pesticides (Silveira et al. 2009; Haro et al. 2018). The conservation biological control provided by this specific intercropping successfully extinguished insecticide spraying for pests such as thrips as Frankliniella occidentalis (Pergande) and Frankliniella schultzei (Trybom) (Thysanoptera: Thripidae) on spring onion and lettuce; vegetable leafminer (Lyriomiza spp. (Diptera: Agromyzidae) on lettuce, spring onion and arugula; and Tetranychus urticae Koch (Acari: Tetranychidae).
In banana production systems, intercropping with companion plants could improve soil physical quality and structure, control nematode population and reduce Fusarium oxysporum incidence, the causal agent of Fusarium wilt (Pattison et al. 2014; Quaresma et al. 2015; Almeida et al. 2018). All these benefits can effectively reduce the need for external inputs as a result of using natural resources for long-term management of farmland, and thereby promote a sustainable farming system (Quaresma et al. 2015; Orr and Nelson, 2018). The main species used in the intercropping are Calopogonium mucunoides Desv. (Fabaceae), Raphanus sativus L. var. oleifera Metzg (Brassicaceae), Avena strigosa Schreb (Poaceae), Vicia sativa L. (Fabaceae) and Lolium multiflorum Lam (Poaceae). Additionally, producers are associating lower damages caused by banana rust and eruption thrips with intercropped farms. Previous studies indicate a higher diversity and abundance of insects in intercropped banana systems (Casaril et al. 2019), and such beneficial effect on thrips population control is currently being studied.
Therefore, there is an immediate need to implement companion plant technologies that have already been tested (e.g. African marigold as companion plants) mainly through the effort of extension agencies. Additionally, it is necessary to investigate techniques currently used by producers for other primary purposes (e.g. intercropping to improve soil quality and control plant diseases in banana crop system), which could have some potential to be used as a pest management tool. These integrated procedures can contribute for the optimisation and long-term sustainability of the agricultural environment, especially for smallholder farmers.
General Conclusions and Synthesis
Increasing plant diversity in agroecosystems has been linked to increased pest management; however, in some cases, increasing plant diversity is not directly related to reduction of pest populations. To better understand the underlying natural process and mechanisms regulating this effect, it is necessary to increase our scientific knowledge. The effects of plant diversity in pest populations are associated with ecological processes of the diversity components within guilds and between trophic levels, including host specificity and selection, species competition or displacements, intraguild predation and apparent competition. For insects and mites, most of these ecological processes are mediated by semiochemicals that drive and modulate insect/mite behaviour. Based on this framework, the concept of deploying smart plants that can respond rapidly to pest damage by releasing or responding to HIPVs has been developed. Within this strategy, smart plants could be used to repel herbivorous pests, attract natural enemies or even to promote plant-plant communication, i.e. plants emitting HIPV blends signalling imminent danger to neighbouring plants. In addition, with technological development, some of this knowledge could be applied to insect monitoring and early pest detection, especially for cryptic pests that are otherwise difficult to detect and/or manage.
The case studies presented herein show the potential of applying companion plants agriculture as tools for pest management. Moreover, these studies provide several examples of repellent or attractive plants, and the provision of resources by these plants, in various cropping systems, to different insects. From this position, the basic information for landscape management is now available for crops such as maize, fruits, vegetables and coffee. In addition, for some crops, as in the case of maize, fundamental knowledge of HIPVs and their effect on pests and their natural enemies can be applied to the development of pest management strategies based on the concept of smart plants. Companion plants, real-time detection of pests and injured plants using advanced technology such as electronic noses and image detectors bring a cleaner and efficient tool to a more sustainable agriculture. It is expected that identification and characterisation of semiochemicals in real time could help to better understand the role of companion plants and the behavioural mechanism of insects/mites mediated by these compounds. This can contribute to establish more efficient and profitable mixtures of crops (i.e. polycultures), and for new tactics for pest management based on insect interactions, plant defences, insect behavioural manipulation and biological control.
It is crucial to mention that some of the studies reported here are already implemented in the field and are part of the farmers’ management strategies in several regions in Brazil. Basic research was fundamental in developing effective and reliable strategies that can be used in conventional and organic systems. The challenge is to upscale these technologies to large cropped areas. Another important issue is that scientific cooperation and the interplay amongst research fields and researchers play a central role in developing such technologies.
With recent advances in fields such as ecology, conservation biology, agronomy and biotechnology, we can now look forward to new opportunities that we did not previously view. We are aware that the initial cost of implementing the smart plant technologies in complex environments with companion plants may be challenging in a first moment. However, this is one of the multiple ways to move from the perspective of pest control based on the use of pesticides to a truly sustainable pest management grounded in the use and manipulation of ecological interactions with a solid scientific background. In this sense, multidisciplinary studies and actions amongst several research areas, extension agencies, governmental and non-governmental entities, and the private sector allied with the farmers needs may play a pivotal role.
References
Almeida NO, Teixeira RA, Carneiro FA, Oliveira CM, Ribeiro VA, Lobo Júnior M, Rocha MR (2018) Occurrence and correlations of nematodes, Fusarium oxysporum and edaphic factors on banana plantations. J Phytopathol 166:265–272
Amaral DSSL, Venzon M, dos Santos HH, Sujii ER, Schmidt JM, Harwood JD (2016) Non-crop plant communities conserve spider populations in chili pepper agroecosystems. Biol Control 103:69–77
Amaral DSSL, Venzon M, Pallini A, Lima P, DeSouza O (2010) A Diversificação da vegetação reduz o ataque do bicho-mineiro-do-cafeeiro Leucoptera coffeella (Guérin-Mèneville) (Lepidoptera: Lyonetiidae)? Neotrop Entomol 39:543–548
Andow DA (1991) Vegetational diversiy and arthropod population response. Ann Rev Entomol 36:561–586
Barzman M, Bàrberi P, Birch ANE, Boonekamp P, Dachbrodt-Saaydeh S, Graf B, Hommel B, Jensen JE, Kiss J, Kudsk P, Lamichhane JR, Messéan A, Moonen AC, Ratnadass A, Ricci P, Sarah JL, Sattin M (2015) Eight principles of integrated pest management. Agron Sust Dev 35:1199–1215
Begg GS, Cook SM, Dye R, Ferrante M, Franck P, Lavigne C, Lövei GL, Mansion-Vaquie A, Pell JK, Petit S, Quesada N (2017) A functional overview of conservation biological control. Crop Prot 97:145–158
Bernardi D, Salmeron E, Horikoshi RJ, Bernardi O, Dourado PM, Carvalho RA et al (2015) Cross-resistance between Cry1 proteins in fall armyworm (Spodoptera frugiperda) may affect the durability of current pyramided Bt maize hydrids in Brazil. PLoS ONE 10:e0140130. https://doi.org/10.1371/journal.pone.0140130
Bernays EA (2001) Neural limitation in phytophagous insects: implications for diet breadth and evolution of host affiliation. Ann Rev Entomol 46:703–727
Bernays EA, Chapman RF (2007) Host-plant selection by phytophagous insects. Contemporary topics in entomology 2. Chapman & Hall, New York, NY. 326 pp
Bernays EA, Minkenberg OPJM (1997) Insect herbivores: different reasons for being a generalist. Ecology 78:1157–1169
Biondi A, Zappalà L, Di Mauro A, Garzia GT, Russo A, Desneux N, Siscaro G (2016) Can alternative host plant and prey affect phytophagy and biological control by the zoophytophagous mirid Nesidiocoris tenuis? Biocontrol 61:79–90
Birkett MA, Pickett JA (2014) Prospects of genetic engineering for robust insect resistance. Curr Opin Plant Biol 19:59–67. https://doi.org/10.1016/j.pbi.2014.03.009
Blassioli-Moraes MC, Laumann RA, Pires CSS, Borges M (2005) Induced volatiles in soybean and pigeon pea plants artificially infested with the Neotropical brown stink bug, Euschistus heros, and their effect on the egg parasitoid, Telenomus podisi. Entomol Exp Appl 115:227–237
Blassioli-Moraes MCB, Pareja M, Laumann RA, Hoffmann-Campos CB, Borges M (2008) Response of the parasitoid Telenomus podisi to induced volatiles from soybean damaged by stink bug herbivory and oviposition. J Plant Int 5:1–13
Borges, M , Blassioli-Moraes, MC (2017) The Semiochemistry of Pentatomidae. In: Miguel Borges and Andrej Colk. (Org.). Stinkbugs: Biorational Control Based on Communication Processes. 1st ed.Londres: CRC press, 2016, 1: 20–50
Borges M, Medeiros MA, Mori K, Zarbin PHG, Ferreira JTB (1998) Field Responses of stink bugs to the natural and synthetic pheromone of the Neotropical brown stink bug, Euschistus heros (Heteroptera: Pentatomidae). Phys Entomol 23:202–207
Botti JMC, Martins EF, Franzin ML, Venzon M (2021) Predation of coffee berry borer by a green lacewing. Neotrop Entomol. https://doi.org/10.1007/s13744-021-00884-0
Brennan EB (2016) Agronomic aspects of strip intercropping broccoli with alyssum for biological control of aphids. Biol Control 97:109–119
Bruce TJA, Aradottir GI, Smart LE, Martin JL, Caulfield JC, Doherty A, Sparks CA, Woodcock CM, Birkett MA, Napier JA, Jones HD, Pickett JA (2015) The first crop plant genetically engineered to release an insect pheromone for defence. Sci Rep 5:11183. https://doi.org/10.1038/srep11183
Bundy CS, McPherson RM (2000) Dynamics and seasonal abundance of stink bugs (Heteroptera: Pentatomidae) in a cotton–soybean ecosystem. J Econ Entomol 93:697–706
Carvalho CF, Carvalho SM, Souza B (2020) Coffee. In: Souza B, Vázquez LL, Maruccci RC (eds) Natural Enemies of Insect Pests in Neotropical Agroecosystems. Springer International Publishing, Springer Nature, Switzerland, pp 277–291
Casaril CE, Oliveira Filho LCI, Santos JCP, Rosa MG (2019) Fauna edáfica em sistemas de produção de banana no Sul de Santa Catarina. Rev Bras Ciên Agr 14:e5613
Cellini A, Blasioli A, Biondi E, Bertaccini A, Braschi I, Spinelli F (2017) Potential applications and limitations of electronic nose devices for plant disease diagnosis. 17:2596
Chaney WE (1998) Biological control of aphids in lettuce using in-field insectaries. In: Pickett CH, Bugg RL (eds) Enhancing biological control: habitat management to promote natural enemies of arthropod pests. University of California Press, Berkeley, pp 73–83
Cingolani MF (2012) Parasitismo de huevos de Piezodorus guildinii (Hemiptera: Pentatomidae) por Trissolcus basalis y Telenomus podisi (Hymenoptera: Scelionidae) en el noreste de la provincia de Buenos Aires. Ph.D. Thesis, Universidad Nacional de La Plata (Buenos Aires, Argentina)
Corbett A, Plant RE (1993) Role of movement in the response of natural enemies to agroecosystem diversification: a theoretical evaluation. Env Entomol 22:519–531
Corrêa-Ferreira BS, Alexandre TM, Pellizzaro EC, Moscard F, Bueno, AF (2010) Práticas de manejo de pragas utilizadas na soja e seu impacto sobre a cultura. Londrina: Embrapa Soja, 2010 (Embrapa Soja. Circular Técnica, 78) 16 p
Cui S, Alfaro Inocente EA, Acosta N, Keener HM, Zhu H, Ling PP (2019) Development of fast E-nose system for early stage diagnosis of aphid-stressed tomato plants. Sensors 19:3480
Cruz I, Figueiredo MLC, Silva RB (2010) Monitoramento de adultos de Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) e Diatraea saccharalis (Fabricius) (Lepidoptera: Pyralidae) em algumas regiões produtoras de milho no Brasil. Documentos/ Embrapa Milho e Sorgo n. 93, 42p
Dethier VG, Browne LB, Smith CN (1960) The designation of chemicals in terms of the responses they elicit from insects. J Econ Entomol 53:134–136
Dicke M, Sabelis MW, Takabayashi J, Bruin J, Posthumus MA (1990) Plant strategies of manipulating predator–prey interactions through allelochemicals: prospects for application in pest control. J Chem Ecol 16:3091–3118
Ecole CC, Silva RA, Louzada JNC, Moraes JC, Barbos LR, Ambrogi BG (2002) Predação de ovos, larvas e pupas do bicho-mineiro-do-cafeeiro, Leucoptera coffeella (Guérin-Menèville & Perrottet, 1842) (Lepidoptera: Lyonetiidae) por Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae). Ciênc Agrotecnol 26:318–324
Esquivel JF, Musolin DL, Jones WA, Rabitsch W, Greene JK, Toews MD, Schwertner CF, Grazia J, McPherson RM (2018) Nezara viridula (L.). In: McPherson JE (ed) Invasive stink bugs and related species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management. CRC Press, UBoca Raton, pp 351–423
Farias JR, Andow DA, Horikoshi RJ, Sorgatto RJ, Fresia P, Santos AC, Omoto C (2014) Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot 64:150–158
Finch S, Collier RH (2000) Host-plant selection by insects – a theory based on ‘appropriate ⁄ inappropriate landings’ by pest insects of cruciferous plants. Entom Exp Et Appl 96:91–102
Foti MC, Rostás M, Peri E, Park FC, Slimani T, Wratten SW, Colazza S (2017) Chemical ecology meets conservation biological control: identifying plant volatiles as predictors of floral resource suitability for an egg parasitoid of stink bugs. J Pest Sci 90:299–310
Foti MC,·Peri E, Wajnberg E, Colazza S, Rostás M (2019) Contrasting olfactory responses of two egg parasitoids to buckwheat floral scent are reflected in field parasitism rates. J Pest Sci 92:747-756
Gomes L, Bianchi FJJA, Cardoso IM, Fernandes RBA, Fernandes Filho EI, Schulte POR (2020) Agroforestry systems can mitigate the impacts of climate change on coffee production: a spatially explicit assessment in Brazil. Agric Ecos Env 294:106858
Gomez-Campo C (1980) Morphology and morpho-taxonomy of the tribe Brassiceae. In: Tsunoda S, Hinata K, Gomez-Campo C (eds) Brassica crops and wild allies, biology and breeding. Japan Scientific Societies Press, Tokyo, pp 3–31
Gontijo LM (2019) Engineering natural enemy shelters to enhance conservation biological control in field crops. Biol Control 130:155–163
Gontijo LM, Beers EH, Snyder WE (2013) Flowers promote aphid suppression in apple orchards. Biol Control 66:8–15
Gontijo LM, Saldanha AV, Souza DR, Viana RS, Bordin BC, Antonio AC (2018) Intercropping hampers the nocturnal biological control of aphids. Ann Appl Biol 172:148–159
Haro, MM (2011) Controle biológico conservativo de pragas em cultivo protegido de tomate orgânic. Lavras: UFLA, 88 p. Dissertação (Mestrado em Entomologia)
Haro MM (2015) Chemical, spatial and temporal characteristics of Tagetes erecta floral resources mediating agricultural food webs. Lavras: UFLA, 154 p. Tese (Doutorado em Entomologia)
Haro MM, Resende ALS, Silva VF, Souza B, Silveira LCP (2015) Parasitoids of horticultural pests associated to commercial development stages of cultivated Apiaceae. Entomotropica 30:174–180
Haro MM, Silveira LCP, Wilby A (2018) Stability lies in flowers: plant diversification mediating shifts in arthropod food webs. PlosOne 13(2):e0193045
Harterreiten-Souza ES, Togni PHB, Pires CSS, Sujii ER (2014) The role of integrating agroforestry and vegetable planting in structuring communities of herbivorous insects and their natural enemies in the Neotropical region. Agrofor Sys 88:205–219
Harterreiten-Souza ES, Togni PHB, Capellari RS, Bickel D, Pujol-Luz JR, Sujii ER (2021) Spatio temporal dynamics of active flying Diptera predators among different farmland habitats. Agr Forest Entomol 23:334–431
Hilje L, Stansly PA (2008) Living ground covers for management of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) and tomato yellow mottle virus in Costa Rica. Crop Protect 27:10–16
Hogg BN, Nelson EH, Mills NJ, Daane KM (2011) Floral resources enhance aphid suppression by a hoverfly. Entomol Exp Appl 141:138–144
Hokkanen HMT (1991) Trap cropping in pest management. Ann Rev Entomol 36:119–138
Huang B, Shi X, Yu D, Öborn I, Blombäck K, Pagella TF, Wang H, Sun W, Sinclair FL (2006) Environmental assessment of small-scale vegetable farming systems in peri-urban areas of the Yangtze River Delta Region, China. Agr Ecosyst Envir 112:391–402
Isbell F, Adler PR, Eisenhauer N, Fornara D, Kimmel K, Kremen C, Letourneau DK, Liebman M, Polley HW, Quijas S, Scherer-Lorenzen M (2017) Benefits of increasing plant diversity in sustainable agroecosystems. J Ecol 105:871–879
Janssen A, Sabelis MW, Magalhães S, Montserrat M, van der Hammen T (2007) Habitat Structure Affect Intraguild Predation. Ecol 88:2713–2719
Karban R, Baldwin IT (1997) Induced responses to herbivory. University of Chicago Press, Chicago
Khan ZR, Midega CAO, Wadhams LJ, Pickett JA, Mumuni A (2007) Evaluation of Napier grass (Pennisetum purpureum) varieties for use as trap plants for the management of African stemborer (Busseola fusca) in a ‘push –pull’ strategy. Entomol Exp Appl 124:201–211
Khan ZR, Pickett JA, Hassanali A, Hooper AM, Midega CAO (2008) Desmodium species and associated biochemical traits for controlling Striga species: present and future prospects. Weed Res 48:302–306
Khan ZR, Midega CAO, Pittchar JO, Murage AW, Birkett MA, Bruce TJA, Pickett JA (2014) Achieving food security for one million sub-Saharan African poor through push–pull innovation by 2020. Phil Trans R Soc B 369:1–10
Kollist H, Zandalinas SI, Sengupta S, Nuhkat M, Kangasjärvi J, Mittler R (2019) Rapid responses to abiotic stress: priming the landscape for the signal transduction network. Trends Pl Sci 24:25–37
Koss AM, Snyder WE (2005) Alternative prey disrupt biocontrol by a guild of generalist predators. Biol Control 32:243–251
Laumann RA, Aquino MFS, Blassioli Moraes MC, Pareja M, Borges M (2009) Response of the egg parasitoids Trissolcus basalis and Telenomus podisi to compounds from defensive secretions of stink bugs. J Chem Ecol 35:8–19
Letourneau DK, Jedlicka JA, Bothwell SG, Moreno CR (2009) Effects of natural enemy biodiversity on the suppression of arthropod herbivores in terrestrial ecosystems. Ann Rev Ecol Evol Syst 40:573–592
Letourneau DK, Ambrecht I, Rivera BS, Lerma JM, Carmona EJ, Daza MC, Escobar S, Galindo V, Gutiérrez C, López SD, Mejía JL, Rangel AMA, Rangel JH, Rivera L, Saavedra CA, Torres AM, Trujillo AR (2011) Does plant diversity benefit agroecosystems? A synthetic review. Ecol Appl 21:9–21
Lichtenberg, EM Kennedy CM, Kremen C, Batáry P, Berendse F, Bommarco R, Bosque-Pérez NA, Carvalheiro LG, Snyder WE, Williams NM, Winfree R, Klatt BK, Åström S, Benjamin F, Brittain C, Chaplin-Kramer R, Clough T, Danforth B, Diekötter T, Eigenbrode SD, Ekroos J, Elle E, Freitas Materić D, Lanza M, Sulzer P, Herbig J, Bruhn D, Turner C, Mason N, Gauci V (2015) Monoterpene separation by coupling proton transfer reaction time-of-flight mass spectrometry with fastGC. Anal Bioanal Chem 407:7757-7763.BM, Fukuda Y, Gaines-Day HR, Grab H, Gratton C, Holzschuh A, Isaacs R, Isaia M, Jha S, Jonason D, Jones CP, Klein AM, Krauss J, Letourneau DK, Macfadyen S, Mallinger RE, Martin EA, Martinez E, Memmott J, Morandin L, Neame L, Otieno M, Park MG, Pfiffner L, Pocock MJO, Ponce C, Potts SG, Poveda K, Ramos M, Rosenheim JA, Rundlöf M, Sardiñas H, Saunders Me, Schon NE, Sciligo AR, Sidhu CS, Steffan-Dewenter I, Tscharntke T, Veselý M, Weisser W, Wilson JK, Crowder DW (2017) A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Glob Change Biol 23:4946- 4957
Lira EC, Bolzan A, Nascimento ARB, Amaral FSA, Kanno RH, Kaise IS, Omoto C (2020) Resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to spinetoram: inheritance and cross-resistance to spinosad. Pest Manag Sci 76:2674–2680
Marouelli WA, Medeiros MA, Souza RF, Resende FV (2011) Produção de tomateiro orgânico irrigado por aspersão e gotejamento, em cultivo solteiro e consorciado com coentro. Hortic Bras 29:429–434
Materić D, Bruhn D, Turner C, Morgan G, Mason N, Gauci V (2015) Methods in plant foliar volatile organic compounds research. App Plant Sienc 3:1–10
Mathews CR, Brown MW, Bottrell DG (2007) Leaf extrafloral nectaries enhance biological control of a key economic pest, Grapholita molesta (Lepidoptera : Tortricidae), in peach (Rosales: Rosaceae). Env Entomol 36:383–389
Medeiros MA, Sujii ER, Morais HC (2009a) Effect of plant diversification on abundance of South American tomato pinworm and predators in two cropping systems. Hort Bras 27:300–306
Medeiros MA, Resende FV, Togni PHB, Sujii ER (2009b) Efeito do consórcio cultural no manejo ecológico de insetos em tomateiro. Brasília: Embrapa Hortaliças, 2009 (Embrapa Hortaliças. Comunicado Técnico, 65) 9p
Medeiros MAD, Sujii ER, Rasi GC, Liz RS, Morais HCD (2009c) Padrão de oviposição e tabela de vida da traça-do-tomateiro Tuta absoluta (Meyrick) (Lepidoptera, Gelechiidae). Rev Bras Entomol 53:452–456
Medeiros MA, Ribeiro PA, Morais HC, Castelo Branco M, Sujii ER, Salgado-Laboriau ML (2010) Identification of plant families associated with the predators Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae) and Hippodamia convergens Guérin-Menéville (Coleoptera: Coccinelidae) using pollen grain as a natural marker. Braz J Biol 70:293–300
Medeiros MA, Sujii ER, Morais HC (2011) Fatores de mortalidade na fase de ovo de Tuta absoluta em sistemas de produção orgânica e convencional de tomate. Bragantia 70:72–80
Michereff MFF, Michereff Filho M, Blassioli-Moraes MC, Laumann RA, Diniz IR, Borges M (2015) Effect of resistant and susceptible soybean cultivars on the attraction of egg parasitoids under field conditions. J Appl Entomol 193:207–216
Michereff MFF, Borges M, Aquino MFS, Laumann RA, Mendes Gomes ACM, Blassioli-Moraes MC (2016) The influence of volatile semiochemicals from stink bug eggs and oviposition-damaged plants on the foraging behaviour of the egg parasitoid Telenomus podisi. Bull Entomol Res 106:663–671
Michereff MFF, Magalhães DM, Hassemer MJ, Laumann RA, Zhou JJ, Ribeiro PEA, Viana PA, Guimarães PEO, Schimmelpfeng PHC, Borges M, Pickett JA, Birkett MA, Blassioli-Moraes MC (2019) Variability in herbivore-induced defence signalling across different maize genotypes impacts significantly on natural enemy foraging behaviour. J Pest Sci 92:723–736
Michereff MFF, Grynberg P, Togawa RC. Costa MMC, Laumann RA, Zhou J-J, Schimmelpfegn PHC, Borges M, Picket JÁ, Birkett MA, Blassioli-Moraes, MC. (2021) Priming of indirect defence responses in maize is shown to be genotype-specific. Arthropod-Plant Interactions 15: 313–328. https://doi.org/10.1007/s11829-021-09826-4
Mizell RF, Riddle TC, Blount AS (2008) Trap cropping for management of stink and leaf footed bugs. Proc Fl St Hort Soc 121:377–382
Moreira HJC, Aragão FD. (2009) Manual de Pragas do Milho. Campinas _SP. Disponível em: https://www.agrolink.com.br/downloads/manual%20de%20pragas%20do%20milho.pdf. Acessed 05 Nov 2021
Ngowi AV, Mbise TJ, Ijani AS, London L, Ajayi OC (2007) Pesticides use by smallholder farmers in vegetable production in Northern Tanzania. Crop Potect 26:1617–1624
Nicholls CI, Altieri MA (2004) Agroecological bases of ecological engineering for pest management. In: Gurr GM, Wratten SD, Altieri MA (eds) Ecological engineering for pest management: advances in habitat manipulation for arthropods. CSIRO publishing, Collingwood, pp 33–54
Niederbacher B, Winkler JB, Schnitzler JP (2015) Volatile organic compounds as non-invasive markers for plant phenotyping. J Exp Bot 66:5403–5416
Norris RF, Kogan M (2000) Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sc 48:94–158
Oelofse M, Hogh-Jensenb H, Abreuc LS, Almeidad GF, Huie QY, Sultanf T, Neergarrd A (2010) Certified organic agriculture in China and Brazil: market accessibility and outcomes following adoption. Ecol Econ 69:1785–1793
Orr R, Nelson PN (2018) Impacts of soil abiotic attributes on Fusarium wilt, focusing on bananas. Appl Soil Ecol 132:20–33
Panizzi AR (1997) Wild hosts of pentatomids: ecological significance and role in their pest status on crops. Ann Rev Entomol 42:99–122
Panizzi AR, Slansky F Jr (1991) Suitability of selected legumes and the effect of nymphal and adult nutrition in the southern green stink bug (Hemiptera: Heteroptera:Pentatomidae). J Econ Entomol 84:103–113
Panizzi AR, Saraiva SI (1993) Performance of nymphal and adult southern green stink bug on an overwintering host plant and impact of nymph to adult foodswitch. Entomol Exp Appl 68:109–115
Panizzi AR, Silva FAC (2012) Seed-sucking bugs (Heteroptera). In: Panizzi AR, Parra JRP (eds) Insect Bioecology and Nutrition for Integrated Pest Management. CRC Pres, Boca Raton, pp 295–324
Panizzi AR, Lucini T (2017) Host plant-stink bug (Pentatomidae) relationships. In: Čokl A, Borges M (eds) Stink bugs: biorational control based on communication processes, 1st edn. CRC Press, Boca Raton, pp 31–58
Pantoja GM (2018) Artrópodes predadores da broca-do-café associados ao ingá. Dissertação (Mestrado)-Universidade Federal de Viçosa, Viçosa, MG, 57 p
Patt JM, Hamilton GC, Lashomb JH (1997) Foraging success of parasitoid wasps on flowers: interplay of insect morphology, floral architecture and searching behavior. Entomol Exp Appl 83:21–30
Pattison AB, Wright CL, Kukulies TL, Molina AB (2014) Ground cover management alters development of Fusarium wilt symptoms in Ducasse bananas. Australas Plant Pathol 43:465–476
Pickett JA, Khan ZR (2016) Plant volatile-mediated signalling and its application in agriculture: successes and challenges. New Phytol 212:856–870
Poveda K, Gómez MI, Martínez E (2008) Diversification practices: their effect on pest regulation and production. Rev Col Entomol 34:131–144
Prokopy RJ, Owens ED (1983) Visual detection of plants by herbivorous insects Ann. Rev Entomol. 28:337–364
Quaresma MAL, Oliveira FL, Silva DMN, Coelho RI, Costa EC (2015) Desempenho de bananeiras cultivar “nanicão” sobre cobertura viva de solo no semiárido. Revista Caatinga 28:110–115
Railsback SF, Johnson MD (2014) Effects of land use on bird populations and pest control services on coffee farms. PNAS 111:6109–6114
Ranum P, Pena-Rosas JP, Garcia-Casal MN (2014) Global maize production, utilization, and consuption. Ann N Y Acad Sci 1312:105–112
Rand TA, Tylianakis JM, Tscharntke T (2006) Spillover edge effects: the dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecol Lett 9:603–614
Ratnadass A, Fernandes P, Avelino J, Habibi R (2012) Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: a review. Agron Sustain Dev 32:273–303
Rea J, Wratten S, Sedcole R, Cameron P, Davis S (2002) Trap cropping to manage green vegetable bug Nezara viridula (L.) (Heteroptera: Pentatomidae) in sweet corn in New Zealand. Agric for Entomol 4:101–107
Resende ALS, Haro MM, Silva VF, Souza B, Silveira LCP (2012) Diversidade de predadores em coentro, endro e funcho sob manejo orgânico. Arq Inst Biol 79:193–199
Resende ALS, Ferreira RB, Silveira LCP, Pereira LPS, Landim DV, Carvalho CF (2015) Desenvolvimento e reprodução de Eriopisconnexa (Germar, 1824) (ColeopteraCoccinellidae) alimentada com recursos florais de coentro (Coriandrumsativum L.) Entomotropica. 30:12–19
Resende ALS, Souza B, Ferreira RB, Aguir-Menezes EL (2017) Flowers of Apiaceous species as sources of pollen for adults of Chrysoperla externa (Hagen) (Neuroptera). Biol Control 106:40–44
Rezende MQ, Venzon M, Perez AL, Cardoso IM, Janssen A (2014) Extrafloral nectaries of associated trees can enhance natural pest control. Agr Ecos Env 188:198–203
Rezende MQ, Venzon M, Santos PS, Cardoso IM, Janssen A (2021) Extrafloral nectary-bearing leguminous trees enhance pest control and increase fruit weight in associated coffee plants. Agr Ecosy Environ 319:107538
Ribeiro AEL, Castellani MA, Pérez-Maluf R, Moreira AA, Leite AS, Costa DR (2014) Ocurrence of green lacewings (Neuroptera: Chrysopidae) in two coffee cropping systems. Afr J Agrl Res 9:1597–1603
Ribeiro AL, Gontijo LM (2017) Alyssum flowers promote the biological control of collard pests. Biocontrol 62:185–196
Robinson KA, Jonsson M, Wratten SD, Wade MR, Buckley HL (2008) Implications of floral resources for predation by an omnivorous lacewing. Basic Appl Ecol 9:172–181
Root RB (1973) Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecol Mon 43:95–124
Rosado, MC Plantas favoráveis a agentes de controle biológico (2007) Dissertação (Mestrado em Entomologia) - Universidade Federal de Viçosa, Viçosa, MG. 59 p
Rosado MC, de Araújo GJ, Pallini A, Venzon M (2021) Cover crop intercropping increases biological control in coffee crops. Biol Control 160:104675
Rosenheim JA, Rusch A, Schellhorn N, Wanger TC, Wratten S, Zhang W (2016) When natural habitat fails to enhance biological pest control – five hypotheses. Biol Cons 204:449–458
Rusch A, Bommarco R, Jonsson M, Smith HG, Ekbom, (2016) Flow and stability of natural pest control services depend on complexity and crop rotation at the landscape scale. J Appl Ecol 50:345–354
Saldanha AV, Gontijo LM, Carvalho RMR, Vasconcelos CJ, Corrêa AS, Gandra RLR (2019) Companion planting enhances pest suppression despite reducing parasitoid emergence. Bas Appl Ecol 41:45–55
Sarthou JP, Badoz A, Vaissière B, Chevallier A, Rusch A (2014) Local more than landscape parameters structure natural enemy communities during their overwintering in semi-natural habitats. Agr Ecosys Environ 194:17–28
Savary S, Willocquet L, Pethybridge SJ, Esker P, McRoberts N, Nelson A (2019) The global burden of pathogens and pests on major food crops. Nat Ecol Evol 3:430–439
Sheehan W (1986) Response by specialist and generalist natural enemies to agroecosystem diversification: a selective review. Env Entomol 15:456–461
Shields MW, Johnson AC, Pandey S, Cullen R, González-Chang M, Wratten SD, Gurr GM (2019) History, current situation and challenges for conservation biological control. Biol Control 131:25–35
Silva CCA, Blassioli-Moraes MC, Borges M, Laumann RA (2018) Food diversification with associated plants increases the performance of the Neotropical stink bug, Chinavia impicticornis (Hemiptera: Pentatomidae). Arth Pl Int 13:423–429
Silva JHC, Saldanha AV, Carvalho RMR, Machado CFM, Flausino BF, Antonio AC, Gontijo LM (2021) The interspecific variation of plant traits in brassicas engenders stronger aphid suppression than the intraspecific variation of single plant trait. J Pest Sci. https://doi.org/10.1007/s10340-021-01421-z
Silva VF, Silveira LCP, Santos A, Santos AJN, Tomazella VB (2016) Companion plants associated with kale increase the abundance and species richness of the natural-enemies of Lipaphis erysimi (Kaltenbach) (Hemiptera:Aphididae). Afr J Agr Res 11:2630–2639
Silveira LCP, Berti Filho E, Pierre LSR, Peres FSC, Louzada JNC (2009) Marigold (Tagetes erecta L.) as an attractive crop to natural enemies in onion fields. Sci Agr 66:780–787
Simpson M, Gurr GM, Simmons AT, Wratten SD, James DG, Leeson G, Nicol HI, Orre-Gordon G (2011) Attract and reward: combining chemical ecology and habitat manipulation to enhance biological control in field crops. J Appl Ecol 48:580–590
Soto-Pinto L, Romero-Alvarado Y, Caballero-Nieto J, Segura Warnholtz G (2001) Woody plant diversity and structure of shade-grown-coffee plantations in Northern Chiapas, Mexico. Rev Biol Trop 49:977–987
Souza HN, Cardoso IM, Fernandes JM, Garcia FCP, Bonfim VR, Santos AC, Carvalho AF, Mendonca ES (2010) Selection of native trees for intercropping with coffee in the Atlantic Rainforest biome. Agrofor Syst 80:1–16
Souza HN, Goede RGM, Brussaard L, Cardoso IM, Duarte EMG, Fernandes RBA, Gomes LC, Pulleman MM (2012) Protective shade, tree diversity and soil properties in coffee agroforestry systems in the Atlantic Rainforest biome. Agr Ecos Env 146:0179–0196
Souza IL, Tomazella VB, Santos AJN, Moraes T, Silveira LCP (2019) Parasitoids diversity in Organic Sweet Pepper (Capsicum annuum) associated with Basil (Ocimum basilicum) and Marigold (Tagetes erecta). Braz J Biol 79:603–611
Straub CS, Simasek NP, Dohm R, Gapinski MR, Aikens EO, Nagy C (2014) Plant diversity increases herbivore movement and vulnerability to predation. Bas Appl Ecol 15:50–58
Thiery D, Visser JH (1986) Masking of host plant odour in the olfactory orientation of the Colorado potato beetle. Entomol Exp Appl 41:165–172
Tamiru A, Bruce TJA, Woodcock CM, Caulfield JC, Midega CAO, Ogol CKPO, Mayon P, Birkett MA, Pickett JA, Khan ZR (2011) Maize landraces recruit egg and larval parasitoids in response to egg deposition by a herbivore. Ecol Lett 14:1075–1083
Tillman PG (2006) Sorghum as a trap crop for Nezara viridula (L.) (Heteroptera: Pentatomidae) in cotton. Environ Entomol 35:771–783
Tillman PG, Cottrell TE (2012) Case study: Trap crop with pheromone capture traps for managing Euschistus servus (Heteroptera: Pentatomidae) in cotton. Psyche ID 401703 10 pp. | https://doi.org/10.1155/2012/401703
Togni PHB, Frizzas MR, Medeiros MA, Nakasu EYT, Pires CSS, Sujii ERS (2009) Dinâmica populacional de Bemisia tabaci biótipo B em tomate monocultivo e consorciado com coentro sob cultivo organico e convencional. Hort Bras 27:183–188
Togni PHB, Laumann RA, Medeiros MA, Sujii ER (2010a) Odour masking of tomato volatiles by coriander volatiles in host plant selection of Bemisia tabaci biotype B. Entomol Exp Appl 136:164–173
Togni PHB, Cavalcante KR, Langer LF, Gravina CS, Medeiros MA, Pires CSS, Fontes EMG, Sujii ER (2010b) Conservação de inimigos naturais (Insecta) em tomateiro orgânico. Arq Inst Biol 77:669–679
Togni PHB, Venzon M, Muniz CA, Martins EF, Pallini A, Sujii ER (2016) Mechanisms underlying the innate attraction of an aphidophagous coccinellid to coriander plants: implications for conservation biological control. Biol Control 92:77–84
Togni PHB, Marouelli WA, Inoue-Nagata AK, Pires CSS, Sujii ER (2018) Integrated cultural practices for whitefly management in organic tomato. J Appl Entomol 142:998–1007
Togni PHB, Venzon M, Souza LM, Santos JPCR, Sujii ER (2019a) Biodiversity provides whitefly biological control based on farm management. J Pest Sci 92:393–403
Togni PHB, Venzon M, Souza LM, Sousa AATC, Harterreiten-Souza ES, Pires CSS, Sujii ER (2019b) Dynamics of predatory and herbivorous insects at the farm scale: the role of cropped and noncropped habitats. Agr for Entomol 21:351–362
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005a) Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol Lett 8:857–874
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist TO, Dormann CF, Ewers RM, Fründ J, Holt RD, Holzschuh A, Klein AM, Kleijn D, Kremen C, Landis DA, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, van der Putten WH, Westphal C (2012) Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol Rev 87:661–685
Tscharntke T, Karp DS, Chaplin-Kramer R, Batáry P, DeClerck F, Gratton C, Hunt L, Ives A, Jonsson M, Larsen A, Martin EA, Martínez-Salinas A, Meehan TD, O’Rourke M, Poveda K, Tylianakis JM, Klein AM, Tscharntke T (2005b) Spatiotemporal variation in the diversity of Hymenoptera across a tropical habitat gradient. Ecology 86:3296–3302
Velasco LRI, Walter GH (1993) Potential of host- switching in Nezara viridula (Hemiptera: Pentatomidae) to enhance survival and reproduction. Environ Entomol 22:326–333
Venzon M, Janssen A, Sabelis MW (2001) Prey preference, intraguild predation and population dynamics of an arthropod food web on plants. Exp Appl Acarol 25:785–808
Venzon M, Rosado MC, Euzébio DE, Souza B, Schoereder JH (2006) Suitability of leguminous cover crop pollens as food source for the green lacewing Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop Entomol 35:371–376
Venzon M, Togni PHB, Amaral DSSL, Rezende MQ, Batista MC, Chiguachi JAM, Martins EF, Perez A (2018) Mobilisation des mécanismes de régulation naturelle des ravageurs via desplantes à multiples services écosystémiques. Inn Agronom 64:83–95
Venzon M, Togni PHB, Chiguachi JAM, Pantoja GM, Silva Brito EA, Sujii ER (2019a) Agrobiodiversidade como estratégia de manejo de pragas. Informe Agropecuário (belo Horizonte) 40:21–29
Venzon M, Togni PHB, Chiguachi JAM, Pantoja GM, da Silva Brito EA, Sujii ER (2019b) Agrobiodiversidade como estratégia de manejo de pragas. Inf Agropec 40:21–29
Vieira CR, Blassioli-Moraes MC, Borges M, Pires CSS, Sujii ER, Laumann RA (2013) cis-Jasmone indirect action on egg parasitoids (Hymenoptera: Scelionidae) and its application in biological control of soybean stink bugs (Hemiptera: Pentatomidae). Biol Control 64:75–82
Vieira CR, Blassioli-Moraes MC, Borges M, Pires CSS, Sujii ER, Laumann RA (2014) Field evaluation of (E)-2-hexenal efficacy for behavioral manipulation of egg parasitoids in soybean. Biocontrol 59:525–537
Volkov A, Ranatunga DR (2006) Plants as Environmental biosensors. Plant Sign Behav 1:105–115
Wäckers FL (2005) Suitability of (extra-) floral nectar, pollen, and honeydew as insect food sources. In: Wäckers FL, Van Rijn PCJ, Bruin J (eds) Plant-provided food for carnivorous insects: a protective mutualism and its applications. Cambridge University Press, Cambridge p, pp 17–74
Zaché B (2009) Manejo de biodiversidade de insetos-praga e inimigos naturais em cultivo de alface (Lactuca sativa) orgânica através do uso de cravo-de-defunto (Tagetes erecta) como planta atrativa. Lavras: UFLA,60 p. Dissertação (Mestrado em Entomologia)
Funding
This work received financial support from the National Council of Technological and Scientific Development (CNPq), the Federal District Research Foundation (FAP-DF) and the Brazilian Corporation of Agricultural Research (EMBRAPA) (20.19.00.182.00.00). The work at Rothamsted forms part of the Smart Crop Protection (SCP) strategic programme (BBS/OS/CP/000001) funded through Biotechnology and Biological Sciences Research Council’s GCRF Translation Awards BBSRC Reference: BB/R020000/1. Reported studies on coffee had support from CNPq and from Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG). Reports studies on diversification from Santa Catarina State Agricultural Research and Rural Extension Agency (Epagri) had support from CNPq (429226/2018-7) and Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC - 2021TR001441).
Author information
Authors and Affiliations
Contributions
This work was conceived during the workshop “Stakeholders engagement workshop: Companion plants and their semiochemicals for pest control in agriculture”. All authors participate in the elaboration of the first draft and revised versions of the ms. All authors approved the version considered for publication.
Corresponding author
Ethics declarations
Ethics Approval
Not applicable.
Conflict of Interest
The authors declare no competing interests.
Additional information
Edited by Juliano Morimoto
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Blassioli-Moraes, M.C., Venzon, M., Silveira, L.C.P. et al. Companion and Smart Plants: Scientific Background to Promote Conservation Biological Control. Neotrop Entomol 51, 171–187 (2022). https://doi.org/10.1007/s13744-021-00939-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-021-00939-2