
Abstract
We characterized dung beetles food preference and diel activity and examined the way such characteristics may structure a Scarabaeinae community in a dry forest. We sampled a fragment of Arboreal Caatinga in Milagres, Bahia, Brazil, during the dry and wet seasons, using baited pitfall (bovine spleen, human feces, cow dung, and rotten banana). Species were classified by activity (nocturnal and diurnal) and food preference (coprophagous, necrophagous, saprophagous, copro-necrophagous, and generalist). In total, 1,581 individuals belonging to 16 morphospecies were sampled, with six new records for Caatinga. The dung beetles were mainly from generalist and coprophagous species; seven species presented nocturnal activity, and five were diurnal. There was higher species richness during the day and greater abundance during the night. Species composition differences were influenced by functional guilds and beetle size according to temporal segregation. These factors may be related to physiological, morphological, and behavioral differences.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Dung beetles belong to the Scarabaeinae subfamily (Coleoptera, Scarabaeidae), which presents rich variation, consisting of about 7,000 species (Philips 2011). The Scarabaeinae comprise several detritivorous species that feed on fungi, animal carcasses, or decomposing fruit, although the diet of most species is composed principally of feces (Halffter & Matthews 1966).
The Scarabaeinae niche can be reduced to three dimensions: space, time, and food resource (Halffter 1991). Two types of temporal variations may be discriminated: on a smaller scale, variations in daily activity patterns can be simplified into diurnal and nocturnal guilds (Gill 1991), and on a larger scale, the seasonal variations commonly associated with rainfall in arid regions (Hernández 2007). The processes that determine temporal variations may be related to abiotic factors (insolation, temperature, humidity, and soil) and to behavioral and morphological factors such as size, color, and type of resource allocation (Vulinec 1997, Lobo et al 1998, Hernández 2002, Verdú et al 2007).
Specialization in food resources can be a way of avoiding competition, although few species present extreme specialization: most Neotropical Scarabaeinae tend to consume all sources of dung (Howden & Nealis 1975, Larsen et al 2006). In South America, most species are coprophages; however, due to the extinction of certain large mammals, it is quite common to find species in tropical forests that have developed other types of food preference, such as necrophagy and saprophagy (Cambefort 1991, Halffter & Halffter 2009).
Studies have examined dung beetle temporal partition and resource utilization in tropical rainforests (e.g., Davis 1999, Silva et al 2012), demonstrating that these two factors can help to reduce interspecific competition (Davis 1999, Feer & Pincebourde 2005). Caatinga ecosystems, like other seasonally dry tropical forests, have undergone a series of adaptations, leading to species survival in areas with low rain concentration throughout the year. Factors related to arid environments can induce changes to dung beetle community structure, mainly through a reduction in species richness and changes in species composition (Hernández 2005, Lopes & Louzada 2005, Lopes et al 2006, Hernández 2007, Liberal 2008, Neves et al 2010, Liberal et al 2011). However, there are no studies focusing on Scarabaeinae resource utilization and temporal segregation in seasonally dry tropical forests in Brazil. This study therefore aims to extend knowledge about Caatinga dung beetles species, with a focus on resource utilization and the relationship between temporal segregation and body size, in order to understand how these characteristics influence the dung beetle community.
Material and Methods
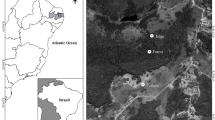
The Caatinga fragment we investigated had Arboreal Caatinga physiognomy, a seasonality dry tropical forest endemic to Brazil. This fragment covered an area of approximately 15,000 m2 and was located on a hilltop surrounded by shrub Caatinga and pastures, both used by livestock. This fragment is located in the municipality of Milagres, in the state of Bahia, Brazil (12°53′ S, 39°51′ W), situated within the eastern part of the ecosystem in the ecoregion known as the “Depressão Sertaneja Meridional” (Velloso et al 2002).
The climate is tropical semiarid (BSh on the Köppen classification) with a mean temperature of 24.3°C, a mean maximum temperature of 29.9°C, and a minimum of 20.6°C. Mean annual precipitation is 551 mm, varying between 142 mm in drier years and 1,206 mm in rainier ones (Aguiar & Zanella 2005). Over the sampling period, the average temperature and precipitation were 24.8°C and 152.65 mm, respectively (Ingá 2011).
Samples were collected in July and October 2010 (dry season) and in January and April 2011 (wet season). Dung beetles were collected using baited pitfalls consisting of a plastic container with an 18-cm opening and 10 cm deep, buried in the ground with its rim at the surface level and partially filled with a detergent solution (2%). Baits were attached to the center of the container, which was then covered with canvas to prevent animals from stealing the bait. A rubber cover was positioned above the pitfall to reduce the effects of weather (Lopes et al 2006).
Every month, 16 sampling points (25 m apart) were arranged evenly in a square grid of 75 m × 75 m, and four pitfalls (2 m apart) were placed at each sampling point, each containing a different type of bait: carrion (bovine spleen), human feces, cow dung, and rotten banana. Eight sampling points were placed during the early morning and eight in the late afternoon, providing fresh resources for each period.
All pitfalls were checked in the early morning (5–6:00 a.m.) and late afternoon (5–6:00 p.m.), and sampled specimens were retrieved, and the detergent solution was renewed. Traps were exposed for 72 h in the field (except during April 2011, when the pitfalls were only exposed for 48 h). The specimens were placed in 70% ethanol, mounted, identified, and deposited in the Johann Becker entomological collection of the university’s Zoology Museum (Museu de Zoologia da Universidade Estadual de Feira de Santana: MZFS).
Species mean length was obtained using a digital caliper to measure the distance between the clypeus and abdomen of 20 individuals from each species whenever possible. From mean length, we defined two size classes: large species (size >1 cm) and small species (≤1 cm) (Arellano et al 2005).
Species were classified as nocturnal or diurnal according to individual predominance during these periods (Hernández 2002). Species were classified as specialist when 80% of the individuals were attracted by one type of bait, and were then categorized as coprophagous (feces), necrophagous (carrion), and saprophagous (rotten banana). Species caught in only feces and carrion pitfalls were considered copro-necrophagous, and those that did not display a marked predominance for baits (<80%) were defined as generalists.
The dung beetle community was analyzed to identify patterns of abundance and richness, according to temporal segregation (day and night) and use of resources, through a generalized linear model (GLM) with Poisson distribution and log link function; sampling points were used for temporal segregation comparisons and pitfalls for resource utilization. Chi-square (χ2) was used to test dung beetle size (small and large) and functional group (tunnellers and rollers) effects on temporal segregation. These analyses were performed on R (R Development Core Team 2006).
Composition changes for temporal segregation and resource utilization were compared using Non-metric dimensional scaling (NMDS) based on a Bray-Curtis similarity matrix with the abundance square root transformed and standardized—sample units were the same as those used for the GLM. The clusters presented in the NMDS were verified using an ANOSIM with 50,000 repetitions, and tests were conducted using Primer® 6.0 software (Clarke & Gorley 2006).
Results
Sampling resulted in 1,581 dung beetle specimens belonging to 16 different morphospecies (Table 1), of which only seven were identified at species level. This study extends the number of species recorded in Caatinga from 34 to 40. Five of the sampled species were diurnal and seven nocturnal. Species of Ateuchus Weber, Dichotomius Hope, Deltochilum Eschscholtz, Ontherus Erichson, and Uroxys Westwood presented nocturnal activity, whereas those of Canthidium Erichson and Canthon Hoffmannsegg presented diurnal activity.
Species richness by point sampling was slightly higher during the day (Daymean = 2.75, Nightmean = 2.09; χ2 = 5.71, p = 0.05), while abundance was higher during the night (Daymean = 9.62, Nightmean = 15.14; χ2 = 77.68, p = 0.01). There were marked differences in species composition according to temporal use of resources (R = 0.994, p = 0.01) (Fig 1), revealing a division in community structure between species. This may reflect the influence of functional guilds, since 88.8% of diurnal individuals were rollers and 73.0% of nocturnal individuals were tunnellers (χ2 = 266.24, df = 1, p = 0.01), or of size, since 55.2% of the diurnal individuals were small and 93.1% of nocturnal individuals were large (χ2 = 368.17, df = 1, p = 0.01).

Non-metric multidimensional scaling (NMDS) based on the Bray-Curtis similarity matrix, comparing species composition between diurnal and nocturnal assemblages in Milagres, Bahia, Brazil.
Four species were exclusively found on feces, three on carrion and one on rotten banana baited pitfalls. Different degrees of richness were found for the baited pitfalls containing carrion and human feces, attracting three times more species than cow dung and rotten banana (χ2 = 155.35, p = 0.01) and seven times more individuals (χ2 = 1145.14, p = 0.01). When using confidence intervals for comparison, the results for overall richness and abundance presented the same pattern (Figs 2 and 3), revealing no differences between human feces and carrion, or between cow dung and banana. However, human feces and carrion attracted more species and individuals than cow dung and banana.

Species richness (means, 99% confidence interval) of dung beetles caught on resources in Milagres, Bahia, Brazil (B banana, C carrion, HF human feces, CD cow dung).

Abundance (means, 99% confidence interval) for of dung beetles caught on resources in Milagres, Bahia, Brazil (B banana, C carrion, HF human feces, CD cow dung).
Non-metric dimensional scaling presented differences in species composition according to resource utilization (baits) (Fig 4) (R = 0.57, p = 0.01). There were minor differences between rotten banana and cow dung (R = 0.197) and intermediate differences between carrion and human feces (R = 0.416), and banana and carrion (R = 0.465). Other pairwise comparisons measured over 67.5%.

Non-metric multidimensional scaling (NMDS) based on the Bray-Curtis similarity matrix, comparing species composition between resources in Milagres, Bahia, Brazil.
Discussion
The number of species in the Caatinga is underestimated because of the high number of morphospecies in each study (43.5 ± 11.9%). Due to the lack of access to type specimens, species listed as affinis were compared to the literature, potential differences were disregarded, and they were evaluated as a single species. We only found three species common to the Caatinga: Dichotomius puncticollis (Luederwaldt), Coprophanaeus pertyi (d’Olsoufieff), and Deltochilum verruciferum Felsche (Lopes et al 2006, Neves et al 2010).
Saprophagous species were represented by Ateuchus sp. 5 and Canthidium sp., the latter has also been collected in Atlantic forest where it demonstrated a preference for traps with rotten banana (A.M.M., personal observations). In a recent review of saprophagy in dung beetles, only three species of Ateuchus were described as saprophagous in the Neotropical region (Halffter & Halffter 2009). This is the first study to record fruit preference by Scarabaeinae in the Caatinga, as well as to register two species with a saprophagous preference. Singleton species were not classified according to feeding habits, although they were captured in baits consistent with genera expectations (Halffter & Matthews 1966), the only exception being Ateuchus sp. 4, which was found in carrion baited traps.
Canthon aff. nigripennis displayed diurnal activity and a preference for carrion. The species has also been recorded in the Atlantic Forest (Endres et al 2007, Filgueiras et al 2009, Silva et al 2010), where it was more frequently found in traps containing carnivore feces than herbivore or omnivore feces—carrion was not offered as bait (Filgueiras et al 2009). These species could therefore be using carnivore feces because it is the food resource most similar to carrion.
Canthon staigi (Pereira) was diurnal and generalist, the latter was supported by the fact that it has even been recorded in cut liana (Pereira & Martinez 1956). This species has been found in semi-deciduous forest (Lopes & Louzada 2005), Atlantic forest (Schiffler et al 2003, Endres et al 2007), and Restinga (Schiffler et al 2003, Vieira et al 2008) and has been described as apparently intolerant to changes in vegetation physiognomy (Vieira 2008).
Coprophanaeus pertyi was a singleton species but was recorded as necrophagous (Hernández 2002); however, it was also classified as generalist (preferably necrophagous) (Silva et al 2007). This may result from local variations or differences in food preference classification. This species is characteristically found in open Caatinga areas at altitudes below 200 m. Although considered endemic to this biome (Gillet et al 2010), it has been found in tree and shrub Caatinga vegetation at an altitude of 650 m (Hernández 2007) and in “Brejo de Altitude” (Atlantic Forest enclaves in Caatinga areas) surrounded by open Caatinga (Silva et al 2007), as well as in Atlantic Forest (Costa et al 2009, Edmonds & Zidek 2010). In Atlantic Forest, the species was collected with flight intercept traps (Costa et al 2009), while in open areas, this was undertaken through baited pitfalls (Silva et al 2010).
Deltochilum aff. irroratum, which was nocturnal and necrophagous, has also been collected in “Brejo de Altitude,” with a food preference similar to that reported here (Silva et al 2007). Deltochilum verruciferum was nocturnal and copro-necrophagous, as it was in another study (Hernández 2007); this is a representative Caatinga species, since it has been found in all studies to date (Hernández 2005, Hernández 2007, Lopes & Louzada, 2005, Lopes et al 2006, Liberal 2008, Neves et al 2010, Liberal et al 2011), as well as in “Brejo de Altitude” (Silva et al 2007).
Dichotomius aff. laevicollis was nocturnal and generalist and belongs to a complex of species widely distributed along the northeast coast of Brazil (Atlantic Forest and Restinga). It is often the most abundant species (Silva et al 2010) and has also been found in Caatinga (Liberal et al 2011). Dichotomius nisus (Olivier) was a singleton species and is classified as nocturnal and coprophagous (Flechtmann et al 1995, Hernández 2007). This species has been found in open Caatinga (Liberal et al 2011) and in pastures and open areas (Aidar et al 2000, Schiffler et al 2003, Mendes & Linhares 2006, Endres et al 2007, Koller et al 2007, Louzada & Silva 2009, Silva et al 2010) and is widely distributed across Brazil (Louzada et al 2007). It is sometimes found with low abundance in closed areas, such as Atlantic Forest, Pantanal, Restinga, and Amazon (Schiffler et al 2003, Scheffler 2005, Vieira et al 2008, Fletchmann et al 2009), although in “Brejo de Altitude” it demonstrated high abundance (Silva et al 2007).
Ontherus ulcopygus Génier was nocturnal and coprophagous. It has been recorded in Cerrado (“Estação Florestal Cabeça de Veado” and “Chapada dos Veadeiros”) (Génier 1996) but potentially has a wider distribution, since the biomes from which it was collected are not clearly defined in the literature describing this species but were found in several Brazilian states with diversified coverage (Génier 1996), corresponding to the first Caatinga record.
Small species, less than 2 g, in line with the limit indicated by Verdú et al (2006), tend to be thermo-conformers and are forced to be active at hotter times of the day, particularly in the middle of the day, given the importance of temperature and body size for biological clock regulation (Verdú et al 2006). Moreover, larger dung beetle species have to be thermo-regulators, in order to deal with the lower temperatures of the Caatinga night, which are similar to those recorded in mountain forest in Mexico (Arellano et al 2005).
A study of mountain forest in Mexico demonstrated that small tropical rollers require a relatively high environmental temperature (22.3–26.0°C) in order to be active (Verdú et al 2007), mainly because of the problem of expending energy to maintain thoracic warmth, due to loss of heat to the environment (Cavenay et al 1995). Moreover, the behavior of rolling portions of resource away from the pile, a feature of telecoprids, is energetically expensive, and performing it at high temperatures may be a way of saving the energy required during the lower temperature hours (Krell-Westerwalbesloh et al 2004).
Thus, we would expect small-sized Canthonini species, for example, to limit their activity to the daytime. This pattern has been found in other studies (Feer & Pincebourde 2005, Hernández 2007). Small roller activity is restricted to the daytime because of the difficulty of increasing body temperature. On the other hand, large tunneller activity is limited to the nighttime hours because of the risk of overheating.
Three generalist species were among the five most abundant species, indicating the possible advantage of generalized diets in hostile environments, such as the Caatinga. The generalist preference of the dominant species, Dichotomius aff. laevicollis, may drive other species to resource specialization and, therefore, limit their population growth. Alternatively, the low abundance of other species may be due to the limited availability of other resources they could use (McNaughton & Wolf 1970).
In contrast to other studies (Liberal 2008, Milhomen et al 2003), human feces did not attract more species or individuals than carrion, and there were small composition differences, since most species in the Neotropical region can utilize both feces and carrion. Under drier conditions, the attractiveness of carrion for dung beetles tends to lessen (Louzada & Lopes 1997, Silva et al 2007), but the drier conditions in Caatinga may also have reduced the attractiveness of human feces, preventing the emergence of this expected pattern. Alternatively, species in this environment are able to utilize carrion under drier conditions.
The low abundance and low richness found in the rotten banana and cow dung indicated that these resources are not very significant in the Caatinga, but demonstrated the possibility of using other available resources, especially for generalist species. Four generalist species were similarly attracted by rotten banana and cow dung, which could be due to the high concentration of plant material.
The greatest difference in composition was between human feces and cow dung, since cow dung was restricted to generalist species. Although both are excrement, differences exist in the volatile compounds they emit (Dormont et al 2010), and dung beetles are more attracted to omnivore dung (Filgueiras et al 2009, Hernández et al 2012). Similar to our findings, cow dung was unattractive in a dry forest in Colombia, where it was pointed out that most Neotropical dung beetles have little capacity to use this resource, given that cattle have only recently been introduced to the region (Bustos-Gómez & Lopera-Toro 2003). Since dung beetles feed almost entirely off the fluid present in feces (Aschenborn et al 1989) and feces tends to desiccate more quickly in drier conditions, it may become unattractive for these species in arid environments.
Despite their rather low number, this study extends the number of species recorded in Caatinga and our knowledge on the ecology of some of the species it carries. Although not conclusive, due to lack of replication in other Caatinga areas, some of the community level aspects addressed here demonstrate that, compared to other habitats, differences exist in the use of resources. Although most species can be sampled with human feces, other resources, such as banana and carrion, add valuable information regarding food preference and increase the number of recorded species. Differences in temporal segregation in this Caatinga area were related to physiological constraints, probably arising from differences in morphology and guild group.
References
Aguiar CML, Zanella FCV (2005) Estrutura da comunidade de abelhas (Hymenoptera: Apoidea: Apiformis) de uma área na margem do domínio da Caatinga (Itatim, BA). Neotrop Entomol 34:15–24
Aidar T, Koller WW, Rodrigues SR, Corrêa AM, Silva JCC, Balta OS, Oliveira JM, Oliveira VL (2000) Besouros coprófagos (Coleoptera: Scarabaeidae) coletados em Aquidauana, MS, Brasil. An Soc Entomol Brasil 29:817–820
Arellano L, Favila ME, Huerta C (2005) Diversity of dung and carrion beetles in a disturbed Mexican tropical montane cloud forest and on shade coffee plantations. Biodivers Conserv 14:601–615
Aschenborn HH, Loughnan ML, Edwards PB (1989) A simple assay to determine the nutritional suitability of cattle dung for coprophagous beetles. Entomol Exp Appl 53:73–79
Bustos-Gómez LF, Lopera-Toro A (2003) Preferencia por cebo de los Escarabajos Coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de un remanente de Bosque Seco Tropical al Norte del Tolima (Colombia), p.59–65. In: Onore G, Reyes-Castillo P, Zunino M (orgs) Escarabeidos de Latinoamérica: Estado del conocimiento. Zaragoza, m3m: Monografías Tercer Milenio, Sociedad Entomológica Aragonesa, SEA, 83 p
Cambefort Y (1991) Biogeography and evolution. In: Hanski I, Cambefort Y (eds) Dung beetle ecology. Princeton University Press, Princeton, pp 198–210
Caveney S, Scholtz CH, McIntyre P (1995) Patterns of daily flight activity in onitine dung beetles (Scarabaeinae: Onitini). Oecologia 103:444–452
Clarke KR, Gorley RN (2006) PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth
Costa CMQ, Silva FAB, Farias ÂI, Moura RC (2009) Diversidade de Scarabaeinae (Coleoptera, Scarabaeidae) coletados com armadilha de interceptação de vôo no Refúgio Ecológico Charles Darwin, Igarassu-PE, Brasil. Rev Bras Entomol 53:88–94
Davis AJ (1999) Species packing in tropical forests: diel flight activity of rainforest dung-feeding beetles (Coleoptera: Aphodiidae, Scarabaeidae, Hybosoridae) in Borneo. Raffles B Zool 47:473–486
Dormont L, Jay-Robert P, Bessière J, Rapior S, Lumaret J (2010) Innate olfactory preferences in dung beetles. J Exp Biol 213:3177–3186
Edmonds WD, Zidek J (2010) A taxonomic review of the neotropical genus Coprophanaeus Olsoufieff, 1924 (Coleoptera: Scarabaeidae, Scarabaeinae). Insecta Mundi 0129:1–111
Endres AA, Creão-Duarte AJ, Hernández MIM (2007) Diversidade de Scarabaeidae s. str. (Coleoptera) da Reserva Biológica Guaribas, Mamanguape, Paraíba, Brasil: uma comparação entre Mata Atlântica e Tabuleiro Nordestino. Rev Bras Entomol 51:67–71
Feer F, Pincebourde S (2005) Diel flight activity and ecological segregation within an assemblage of tropical forest dung and carrion beetles. J Trop Ecol 21:21–30
Filgueiras BKC, Liberal CN, Aguiar CDM, Hernández MIM, Iannuzzi L (2009) Attractivity of omnivore, carnivore and herbivore mammalian dung to Scarabaeinae (Coleoptera, Scarabaeidae) in a tropical Atlantic rainforest remnant. Rev Bras Entomol 53:422–427
Flechtmann CAH, Rodrigues SR, Gaspareto CL (1995) Controle biológico da mosca-dos-chifres (Haematobia irritans) em Selvíria, Mato grosso do Sul. 5. Seleção de besouros coprófagos. Rev Bras Entomol 39:277–286
Flechtmann CAH, Tabet VG, Quintero I (2009) Influence of carrion smell and rebaiting time on the efficiency of pitfall traps to dung beetle sampling. Entomol Exp Appl 132:211–217
Génier F (1996) A revision of the Neotropical genus Ontherus Erichson (Coleoptera: Scarabaeidae, Scarabaeinae). Mem Entomol Soc Can 170:1–169
Gill BD (1991) Dung beetles in American tropical forest. In: Hanski I, Cambefort Y (eds) Dung beetle ecology. Princeton University Press, Princeton, pp 211–229
Gillett CPDT, Gillett MPT, Gillett JEDT, Vaz-de-Mello FZ (2010) Diversity and distribution of the scarab beetle tribe Phanaeini in the northern states of the Brazilian Northeast (Coleoptera: Scarabaeidae: Scarabaeinae). Insecta Mundi 118:1–19
Halffter G (1991) Historical and ecological factors determining the geographical distribution of beetles (Coleoptera Scarabaeidae: Scarabaeinae). Folia Entomol Mex 82:194–238
Halffter G, Halffter V (2009) Why and where coprophagous beetles (Coleoptera: Scarabaeinae) eat seeds, fruits or vegetable detritus. Bol Soc Entomol Aragon 45:1–22
Halffter G, Matthews E (1966) The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera, Scarabaeidae). Folia Entomol Mex 12–14:1–312
Hernández MIM (2002) The night and day of dung beetles (Coleoptera, Scarabaeidae) in the Serra do Japi, Brazil: elytra colour related to daily activity. Rev Bras Entomol 46:597–600
Hernández MIM (2005) Besouros Scarabaeidae (Coleoptera) da área do Curimataú, Paraíba, p.369–380. In: de Araújo FS, Rodal MJN, Barbosa MRV (orgs) Análise das Variações da Biodiversidade do Bioma Caatinga: Suporte a Estratégias Regionais de Conservação. Brasília. Ministério do Meio Ambiente, 445 p
Hernández MIM (2007) Besouros escarabeíneos (Coleoptera: Scarabaeidae) da Caatinga Paraibana, Brasil. Oecol Bras 11:356–364
Hernandez NM, Muñoz GS, Quintero KS, Méndez JB (2012) Escarabajos coprófagos (Coleoptera: Scarabaeinae) asociados a excrementos de mamíferos en un fragmento de bosque seco tropical en el Departamento del Atlántico, Colombia. Ecol Austral 22:203–210
Howden HF, Nealis VG (1975) Effects of clearing in a tropical rain forest on the composition of the coprophagous scarab beetle fauna Coleoptera. Biotropica 7:77–83
Ingá (2011) Pluviometria diária de Milagres. http://www.inga.ba.gov.br. Accessed 06 August 2011
Koller WW, Gomes A, Rodrigues SR, Goiozo PFI (2007) Scarabaeidae e Aphodiidae coprófagos em pastagens cultivadas no cerrado sul-mato-grossense. Rev Bras Zooc 9:81–93
Krell-Westerwalbesloh S, Krell F-T, Linsenmair KE (2004) Diel separation of Afrotropical dung beetle guilds avoiding competition and neglecting resources (Coleoptera: Scarabaeoidea). J Nat Hist 38:2225–2249
Larsen TH, Lopera A, Forsyth A (2006) Extreme trophic and habitat specialization by Peruvian dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Coleop Bull 60:315–324
Liberal CN (2008) Diversidade de Scarabaeinae (Coleoptera: Scarabaeidae) em uma Área de Caatinga na Região de Parnamirim, PE, p. 79–93. In: Leal IR, Almeida-Cortez J, Santos JC (orgs) Ecologia da Caatinga: Curso De Campo. Recife, Editora Universitária, 179 p
Liberal CN, Isidro de Farias ÂM, Meiado MV, Filgueiras BKC, Iannuzzi L (2011) How habitat change and rainfall affect dung beetle diversity in Caatinga, a Brazilian semi-arid ecosystem. J Insect Sci 11:1–11
Lobo JM, Lumaret JP, Jay-Robert P (1998) Sampling dung beetles in the French Mediterranean area: effects of abiotic factors and farm practices. Pedobiol 42:252–266
Lopes PP, Louzada JNC (2005) Besouros (Scarabaeidae e Histeridae), p.284–298. In: Juncá FA, Funch, L, Rocha W (orgs) Biodiversidade e Conservação da Chapada Diamantina. Brasília, Ministério do Meio Ambiente, 435 p
Lopes PP, Louzada JNC, Vaz-de-Mello FZ (2006) Organization of dung beetle communities (Coleoptera, Scarabaeidae) in areas of vegetation re-establishment in Feira de Santana, Bahia, Brazil. Sitientibus - Ser Ciênc Biolog 6:261–266
Louzada JNC, Lopes FS (1997) A comunidade de Scarabaeidae copro-necrófagos (Coleoptera) de um fragmento de mata atlântica. Rev Bras Entomol 41:117–121
Louzada JNC, Silva P (2009) Utilization of introduced Brazilian pastures ecosystems by native dung beetles: diversity patterns and resource use. Insect Conserv Diver 2:45–52
Louzada JNC, Lopes FS, Vaz-de-Mello FZ (2007) Structure and composition of a dung beetle community (Coleoptera, Scarabaeinae) in a small forest patch from Brazilian Pantanal. Rev Bras Zooc 9:199–203
McNaughton SJ, Wolf LL (1970) Dominance and the niche in ecological systems. Science 167:131–139
Mendes J, Linhares AX (2006) Coleoptera associated with undisturbed cow pats in pastures in Southeastern Brazil. Neotrop Entomol 35:715–723
Milhomem MS, Vaz-de-Mello FZ, Diniz IR (2003) Técnicas de coleta de besouros copronecrófagos no Cerrado. Pesq Agropec Bras 38:1249–1256
Neves FS, Oliveira VHF, Espírito-Santo MM, Vaz-de-Mello FZ, Louzada JNC, Sanchez-Azofeifa A, Fernandes GW (2010) Successional and seasonal changes in a community of dung beetles (Coleoptera: Scarabaeinae) in a Brazilian tropical dry forest. Nat Conserv 8:160–164
Pereira FS, Martínez A (1956) Os gêneros de Canthonini americanos (Col. Scarabaeidae). Rev Bras Entomol 6:91–192
Philips TK (2011) The evolutionary history and diversification of dung beetles. In: Simmons LW, Ridsdill-Smith TJ (eds) Ecology and evolution of dung beetles. Blackwell, Chichester, pp 21–26
R Development Core Team (2006) R.2.14.1 A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, http://www.R-project.org
Scheffler PY (2005) Dung beetle (Coleoptera: Scarabaeidae) diversity and community structure across three disturbance regimes in eastern Amazonia. J Trop Ecol 21:9–19
Schiffler G, Vaz-de-Mello FZ, Azevedo CO (2003) Scarabaeidae s. str. (Coleoptera) do Delta do Rio Doce e Vale do Suruaca no Município de Linhares, Estado do Espírito Santo, Brasil. Rev Bras Zooc 5:205–211
Silva FAB, Hernández MIM, Ide S, Moura RC (2007) Comunidade de Escarabeíneos (Coleoptera, Scarabaeidae) Copro-necrófagos da Região de Brejo Novo, Caruaru, Pernambuco, Brasil. Rev Bras Entomol 51:228–233
Silva FAB, Costa CMQ, Moura RC, Farias I (2010) Study of the dung beetle (Coleoptera: Scarabaeidae) community at two sites: Atlantic forest and clear-cut, Pernambuco, Brazil. Environ Entomol 39:359–367
Silva P, Vaz-de-Mello FZ, Di Mare RA (2012) Attractiveness of different bait to the Scarabaeinae (Coleoptera: Scarabaeidae) in Forest Fragments in Extreme Southern Brazil. Zool Stud 51:429–441
Velloso AL, Sampaio EVSB, Pareyn FGC (2002) Ecorregiões propostas para o Bioma Caatinga. Recife, Associação Plantas do Nordeste, Instituto de Conservação Ambiental, The Nature Conservancy do Brasil, 76 p
Verdú JR, Arellano L, Numa C (2006) Thermoregulation in endothermic dung beetles (Coleoptera: Scarabaeidae): effect of body size and ecophysiological constraints in flight. J Insect Physiol 52:854–60
Verdú JR, Arellano L, Numa C, Micó E (2007) Roles of endothermy in niche differentiation for ball-rolling dung beetles (Coleoptera: Scarabaeidae) along an altitudinal gradient. Ecol Entomol 32:544–551
Vieira L, Louzada JNC, Spector S (2008) Effects of degradation and replacement of Southern Brazilian coastal sandy vegetation on the dung beetles (Coleoptera: Scarabaeidae). Biotropica 40:719–727
Vulinec K (1997) Iridescent dung beetles: a different angle. Fla Entomol 80:132–141
Acknowledgments
We would like to thank our field team, Monica Ulysséa, Elkiaer Campos, Felipe Camurugi, and Ivan. We also thank Fernando Zagury Vaz-de-Mello of the Universidade Federal de Mato Grosso (UFMT) for his help in dung beetle identification and Evandro Silva and Carlos Bichara for their ongoing support, from project design to the early versions of this manuscript. During the study period, Anderson Medina received a graduate grant from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by Kleber Del Claro – UFU
Rights and permissions
About this article
Cite this article
Medina, A.M., Lopes, P.P. Resource Utilization and Temporal Segregation of Scarabaeinae (Coleoptera, Scarabaeidae) Community in a Caatinga Fragment. Neotrop Entomol 43, 127–133 (2014). https://doi.org/10.1007/s13744-014-0198-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-014-0198-9