
Abstract
Purpose of Review
Skin cancer is the most prevalent human cancer and has presented a considerable world incidence rate increase over the last years. While animals offer highly complex systems to study skin cancer biology, ethical concerns and the differences found between the human and animal skin have promoted the development of alternative methods. In this review, we provide an overview of the most important skin cancer models and describe the advantages of in vitro substitutes over ex vivo and animal models. To facilitate understanding, an insight into the biology of skin cancer is also given.
Recent Findings
Currently, the study of cancer pathology and the development of therapeutic agents are mostly based on animal experiments. The methodologies used are based on mice genetic modifications and the induction of carcinogenesis by UV radiation, chemicals, or viruses. Skin explant models represent one alternative to animal models. These preserve the structure and composition of native skin and are therefore highly suitable. More recent studies have developed in vitro skin models composed of the epidermal and/or the dermal layers, which more faithfully mimic the human skin.
Summary
Various technologies have been studied to create improved skin substitutes. To better understand skin cancer biology, models that more faithfully mimic the human skin are needed. In vitro systems can recreate many of the typical features of both healthy and diseased skin and suggest that complex systems like full-thickness models are highly valuable tools to perform human skin studies.
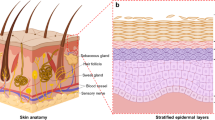
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cancer is characterized by the uncontrolled growth of transformed cells. Mutations in the DNA of these cells lead to an imbalance between proliferation and cell death, resulting in tumor formation. When transformed cells enter the circulation, tumors may arise in other organs (metastasis), invading surrounding tissues and taking away nutrients to survive and grow [1,2,3]. Regarding dermatological cancer, each year between 2 and 3 million new cases are reported, representing around 30% of all newly diagnosed cancers, and making skin cancer by far the most common malignancy [4, 5].
Age is a major risk factor in dermatological cancers and can be a result of life-long sun exposure. Some chemicals, virus infections, and psychological stress are involved in many physiopathological processes and may trigger skin cancer as well. Also, certain genetic features like freckles, moles, red hair, or blue eyes are associated with increased susceptibility to this pathology [3, 6].
There are three main types of skin cancer: melanoma, basal cell carcinoma (BCC), and squamous cell carcinoma (SCC). BCC and SCC belong to the non-melanoma skin cancer (NMSC) group and represent the vast majority of skin cancers. The characteristics of the three types are explained below.
Melanoma
Melanoma is responsible for most skin cancer-related deaths. Each year, more than 130,000 melanoma patients are diagnosed in the USA [1, 7], and, importantly, its incidence has increased more than fourfold in the last 40 years in Caucasian Americans [5, 8]. Early-stage melanoma can be removed through surgical resection and is associated with a survival of 93% [9], whereas survival rates fall to 27% in advanced stages [9].
Melanoma is caused by UV-induced damage to the DNA of the melanocytes residing in the skin, which are responsible for modulating skin tanning upon UV radiation. Transformed melanocytes display dramatically increased proliferation and migration rates, penetrate the basement membrane, and rapidly invade the dermis [7, 8, 10]. This pathology can also alter genotoxic, mitogenic, and immune pathways, allowing tumor growth. For example, melanoma-derived suppressive factors prevent the activation of immature dendritic cells, which results in the suppression of immune responses against tumor cells and thus permit tumor growth. These findings have led to the recent development of immune-focused therapies, among which transfer of tumor-infiltrating lymphocytes and immune checkpoint inhibition stand out [11].
Squamous Cell Carcinoma
SCC is the second most prevalent NMSC [3, 10], grows faster than BCC, and results from the accumulation of genetic alterations. It accounts for most NMSC-related deaths with metastasis rates of 0.3–3.7% [11,12,13].
SCC starts in the squamous cells of the epidermis [12], arising from aberrant, dysplastic keratinocytes that grow to form a tumor. Tumor growth compromises differentiation and then leads to stratum corneum thickening, with cells retaining their nuclei [10]. At the genetic level, SCC displays frequent mutations in the tumor suppressor gene p53, primarily caused by UV radiation [3]. Indeed, over 50% of human SCC present p53 mutations [11, 13]. Another common genetic alteration in SCC involves the overexpression of phosphorylated STAT3 [10], as well as mutations of WNT, Ras, p16INK4, NF-κB, and c-Myc [13].
Basal Cell Carcinoma
BCC is one of the most common human cancers, accounting for about 80% of NMSC, and rarely spread to other parts of the body. It is very rarely fatal despite having a high frequency [3, 11,12,13], but may lead to major deformities and morbidity [10, 12, 13], resulting in a heavy economic burden on patients and healthcare systems [14].
BCC arises from abnormal epidermal basal cells and tends to grow slowly. Unlike melanoma and SCC, it does not present precursor lesions [3]. However, prompt treatment of BCC is vital because as the tumor grows, it becomes more dangerous and may grow into nearby tissue leading to deformity and significant morbidity [10, 12, 13].
The development of BCC can be triggered by genetic mutations. Particularly, mutation of the tumor suppressor PTCH gene, involved in the Hedgehog (Hh) signaling pathway, has been strongly associated with BCC, along with the constant activation of Gli1 and Gli2 transcription factors. Alterations in genes like CYP, GST, and p53 have also been linked to BCC [3, 10, 13].
Various experimental methods exist for recreating the different types of human skin cancer. These facilitate the development of more effective therapies and provide a better understanding of cancer pathology and the mechanisms that mediate invasion and progression [1, 7]. To obtain reliable data, we need to accurately replicate the biology of human skin cancer, a process that largely depends on representative models. The objective of this review is to provide an overview of the main skin cancer models, emphasizing the advantages of in vitro substitutes. We will describe the methodologies used to model the three types of skin cancer, arguing why in vitro models are necessary.
Modeling Human Skin Cancer
Animal Models
Animal models are important tools to better understand different skin processes, as they provide complex systems where several types of cells and tissues are present. In cancer research, animal models are especially useful to investigate the mode of action of new drugs and study cancer progression and metastasis, events that can occur only in vivo [11].
Because of their relatively easy care and biological similarity with humans, mice are the most commonly used animal. They are typically exposed to UV radiation, chemicals, or viruses with the aim of producing carcinogenesis. Many genetically engineered mouse models have been developed through the inactivation of tumor suppressor genes and/or activation of oncogenes. Another approach is the xenograft model in which human cancer cell lines are injected or transplanted into immunodeficient mice [11].
Animal Models of Melanoma
Given its high mortality rate and spread, animal models of melanoma are the most numerous and diverse [5]. In this respect, different mouse models with an innate predisposition to develop melanoma have been created. For example, neonatal mice inappropriately expressing the hepatocyte growth/scatter factor in the skin develop melanoma following UV exposure, with genetic alterations and histopathological characteristics that are similar to the human condition [15]. Other transgenic mice examples include mice with melanocyte overexpression of B-RAFV600E or SV40 T-antigen (an inhibitor of p53), which lead to the development of spontaneous melanomas [10].
Mouse xenograft models are also used to study melanoma growth, spread, and therapeutic response. Metastatic tumors are induced by implanting human melanoma cells into the eyes of immunodeficient mice [5]. However, it has been shown that these xenografts do not faithfully mimic the microenvironment, the initial events of human melanoma, or the invasion of cells to the surrounding tissue [7].
Besides rodents, fish and mammals have been employed as well. In the Xiphophorus fish, melanoma develops after UVA and UVB radiation, affecting the same genes involved in human melanoma [5]. UV radiation is also used in Monodelphis domestica (a South American opossum) to induce melanoma. Given its mammal condition, it results better than the fish to study human diseases. However, unlike human disease, tumors arise from the dermis rather than the epidermis [5], suggesting that differences exist during melanoma development.
Animal Models of SCC
Animal models of SCC are mainly based on the induction of skin tumors in mice by means of chemicals and UV radiation. In the most commonly used method, mice are first treated with a carcinogen that produces mutations in skin cells. Then, a promoter agent is applied to provoke the growth of benign tumors (papillomas) from the mutagenized cells. Over time, such papillomas transform into malignant SCC with the capability of spreading to distant sites [16].
Another approach involves the use of hairless but immunocompetent mice, in which SCC induction is achieved by UV exposure. These SCC usually present p53 mutations, very similar to those seen in human SCC [16], and allow the testing of new therapeutic anti-photocarcinogenic drugs [17]. Nevertheless, even if hair absence is beneficial for UV light experiments, these mice have a non-functional hairless gene, which greatly contribute to skin metabolism and whose absence may influence the outcome of tumor development [16].
In some transgenic mouse models, oncogenes are activated in a subset of the skin cells to investigate tumor formation from a particular cell of origin. For example, expression of mutant H-Ras in the outer root sheath of the hair follicle (where stem cells reside) leads to malignant SCC and spindle carcinomas. However, when H-Ras is targeted to terminally differentiated cells, only benign tumors (papillomas) arise [16].
Animal Models of BCC
Because murine UV-induced BCC do not mimic the human condition very well [10], the development of proper BCC mouse models has been difficult. For example, PTCH heterozygous knockout mice exposed to UV developed BCC-like tumors, which, however, did not recreate completely the different types of the human condition [18]. Despite this, promising results have been achieved when activating other components of the Hh signaling pathway: the first animal model of this kind presented BCC after 4 days of embryonic skin development [19], which were indistinguishable from human tumors. Subsequent studies demonstrated that these types of tumors also arose after the overexpression of mutant SMO [20], and the transcription factors Gli1 and Gli2 [21, 22]. Likewise, other genetic modifications like epidermal Notch1 knockout have led to mice with increased susceptibility to tumor growth that in turn developed BCC-like tumors when subjected to the carcinogen/promoter protocol [23].
As previously mentioned, an alternative method is based on tumor xenografts. However, there have only been a few reproducible xenograft implantations of human BCC in animals [24], since numerous attempts of transplanted human BCC failed to form lesions that closely resembled their human counterpart [24]. For example, when Carlson et al. transplanted 14 BCC tumors into 18 SCID-beige mice (T, B, and natural killer cells deficient), only 3 generated anaplastic tumors [25].
Ideally, a good animal model of cancer should share features with the human disease on various levels [5]. While important progress has been made with various animal models, important limitations need to be considered: (1) the ethical problems raised by the use of laboratory animals; (2) the differences found between the human and animal skin, which makes it difficult to extrapolate the results [1, 10]; and (3) the influence of the mice background on the tumor outcome. Some strains of laboratory mice are known to be tumor-resistant, whereas others are skin tumor-susceptible [16]. Moreover, immune responses of transgenic mice affect tumor growth and the ability of viral vectors to infect cells [24]. The problem with the lack of models that accurately predict human responses is that they may delay the assessment of new treatments against skin tumors.
Are Ex Vivo Models a Suitable Alternative?
Ex vivo models are living tissues obtained from a living organism and, therefore, represent platforms with minimal alteration of the native tissue state. Ex vivo human skin models are therefore highly suitable since they are of a human origin and contain all the cellular components, meaning that these can provide a proper microenvironment [26] leading to more accurate responses.
One of the most outstanding ex vivo models was developed recently by Hochberg. He proposed a creative way of using SCC and BCC biopsies in which skin tumors are sliced and maintained in culture medium. This model has been used to evaluate viruses’ activities on the tumors and the effect of UV radiation. Of note, the three-dimensional (3D) structure is conserved, keeping the original cell types and extracellular matrix products, and positively impacting the results after anti-tumor treatments [24].
Surprisingly, not many ex vivo substitutes have been reported, even when they have shown a high level of similarity to native tumors. Besides inter-donor variability, the reason behind the low popularity of ex vivo platforms is that these require a regular supply of fresh biopsies of proper size [1, 7, 26, 27], which may not be always available. Moreover, upon reception, explants need to be promptly used because cellular viability and metabolic activity rapidly decrease after tissue removal [27]. This strongly limits their use especially when various experiments or tissue replicates are needed (e.g., drug screening).
A Promising Tool: In Vitro Models
As argued above, accurate and more practical model systems are crucial to better understand skin cancer development and progression. While animal models have led to significant progress, they are limited by their accuracy and ethical concerns. Furthermore, the ex vivo alternative is restricted by the availability of living tissue, reason why in vitro substitutes are becoming increasingly more important.
In vitro skin models are advantageous systems since they are of human origin and allow controlling the cellular composition. Researchers can integrate or omit cell components in order to study their importance in the process under investigation [28]. Additionally, these avoid the need to recruit volunteers, do not depend on continuous skin supply, and contribute to the reduction of laboratory animals [29].
To date, two kinds of in vitro skin models are available: bi-dimensional (2D) and 3D cultures. A 2D culture is an anchorage-dependent system with cells growing attached to a surface. Despite its simplicity, cells are restricted by their incapacity to mimic the 3D in vivo structure and behavior. Cell stratification and differentiation are poor and may show hyperproliferative growth, and therefore, this is not the best tool to study cellular responses. In fact, important signaling pathways work only when cells are placed in a 3D structure [30].
In vitro 3D models, on the other hand, provide an in vivo-like structure containing human skin cells and components of extracellular matrix like collagen. Recent advances in tissue engineering have provided novel models to study early neoplastic progression in a more reliable structure [30]. These offer the tools to better understand the biological properties of tumor cells, develop efficient therapies, and allow for the investigation of diverse cellular processes such as proliferation, gene activation, invasion, protein expression, and metastatic behavior [5, 7].
In Vitro Models of Melanoma
The most relevant in vitro melanoma models include spheroids and 3D structures containing both cancer and healthy cells. For example, Genoskin has developed OncoSkin®, an in vitro melanoma model composed of all the human skin layers and cells. Briefly, a melanoma spheroid is generated and embedded into a human skin biopsy, resulting in melanoma cells surrounded by the normal skin environment. This approach enables the study of melanoma cell invasion in situ in a human artificial skin sample, which can maintain viability and integrity for several days [31]. Another option includes the reconstruction of the tumor from de-epidermized dermis and skin cells. Eves et al. developed a 3D model to compare the invasive characteristics of normal melanocytes to a cutaneous melanoma cell line (HBL). To this end, normal fibroblasts were deposited in a de-epidermized human dermis and then HBL cells (or melanocytes) and keratinocytes were seeded on top. In this model, HBL cells were able to invade the dermis, while normal melanocytes were not [32].
Instead of native tissue, Meier et al. used fibroblast-containing collagen gels as a dermal compartment. They studied various stages of melanoma progression by analyzing the growth phases: rapid and vertical growth phases and metastatic melanoma. For this, keratinocytes and melanocytes or melanomas cells (representing different stages of tumor progression) were seeded on top of the dermal equivalent. Interestingly, they identified different growth characteristics associated with the original lesions, which remained stable over time [33], and demonstrated that skin reconstructs are promising tools to model the diverse growth patterns of melanoma.
The reconstruction of the dermal end epidermal compartments also permits the incorporation of different cell types. For instance, Zhang et al. created a novel immune 3D melanoma model composed of 4 different layers: a collagen matrix with embedded fibroblasts, a melanoma cell line (WM35) layer, another collagen-fibroblast layer, and an outer layer of collagen, fibroblasts, and T cells. Interestingly, T cells migrated toward melanoma cells, causing the apoptosis of malignant cells [34] and making this type of model a powerful tool to study the chemokines and the receptors associated with T cell migration.
An alternative method includes the combination of the previous techniques: Vörsmann et al. used both collagen gels and melanoma spheroids containing the melanoma cell line 451-LU. To this end, they generated a collagen I matrix containing fibroblasts with primary keratinocytes seeded on top. By introducing melanoma spheroids into the dermal matrix, they created a model that recapitulates the 3D structure and multicellular complexity of native tumor. They also showed significant differences in therapeutic outcome between regular 2D and complex 3D models, demonstrating the advantages of using 3D structures when studying therapeutic agents responses [35]. Similarly, Leight et al. created a 3D model to study the effect of PLX4032 (a clinically important anticancer drug) treatment on melanoma proteolytic activity. They embedded melanoma cells of different stages in poly(ethylene glycol) (PEG) scaffolds and monitored the matrix metalloproteinases (MMP) activity after drug exposure. They found that treatment with PLX4032 led to an increase in MMP activity in the model, resulting in improved cell migration. These results showed how promising therapeutic agents may lead to higher proteolytic activity and, thus, poor outcomes [36].
The models presented above clearly recapitulate important characteristics of melanoma. However, most of them use human cell lines [5], which can display genetic, physical, and biochemical modifications, and thus, there is a concern that findings in cell lines may not fully represent primary cell behavior [37]. See Table 1 for a summary of the advantages and disadvantages of melanoma models.
In Vitro Models of SCC
Concerning NMSC, current efforts are mainly focused on the development of SCC equivalents [40] (see Table 2), most of which use collagen scaffolds as a dermal compartment. Borchers et al. produced a 3D model of SCC composed of keratinocytes or SCC cells seeded on a type 1 collagen gel, with or without human dermal fibroblasts. The model provided strong evidence that elevated expression of MMP-9 (involved in invasion and metastasis) is dependent upon the interaction of tumor cells with adjacent stromal fibroblasts [41], demonstrating the importance of having more than one cell type in in vitro models. Thereafter, Commandeur et al. generated a three-dimensional model of SCC composed of a collagen matrix with or without normal human fibroblasts and the cell lines SCC-12B2 and SCC-13 seeded on top. The model closely recapitulated important features of SCC such as impaired differentiation and expression of hyperproliferation-related keratins, highlighting the feasibility of creating an in vitro 3D SCC model. The study provided a promising system to test therapeutic agents and better understand SCC-associated events [42]. Using this model, the authors next studied the effect of epidermal growth factor receptor (EGFR) in SCC. Just like the healthy counterpart, the model negatively reacted to increasing concentrations of EGF and inhibition of epidermal growth, making it a suitable platform to investigate the impact of altered EGFR signaling in SCC [43].
Later, Brauchle et al. developed early- and late-stage SCC models containing the SCC cell line SCC-25. For early-stage models, keratinocytes and SCC-25 cells were seeded on top of a type-I collagen matrix. For late-stage models, they used an epidermis composed solely of SCC cells and deposited onto the collagen scaffold. SCC cells were also integrated into the dermal part of the model. As a result, SCC-25 cells were able to generate tumor nests in both models, demonstrating the capability of 3D in vitro models to mimic the different stages of SCC and providing a promising tool to better understand SCC pathology and test new therapeutic agents [44].
An additional alternative relies on using genetically modified cells. An example is that the protein lysyl oxidase (LOX) has been associated with antitumor activity and is downregulated in many carcinomas. Bouez et al. developed a BCC and SCC skin equivalent by seeding fibroblasts on a chitosan-cross-linked collagen-glycosaminoglycan (GAG) matrix, with keratinocytes and the HaCat cell line (wild-type or LOX silenced) seeded on top. Interestingly, they found an invasive phenotype in the skin equivalent where the protein LOX was specifically absent [45], demonstrating the ability of the model to recapitulate the invasive phenotype of NMSC. In the same line, Berning et al. used a fibroblast-derived matrix co-cultured with different human-modified keratinocyte cell lines: HaCaT, benign tumorigenic HaCaT-RAS A-5, malignant tumorigenic HaCaT-RAS II-4, metastatic HaCAT-RAS A5-RT3, and SCC cell lines (SCC-12, SCC-13, SCL-I, and SCL-II). They were able to obtain skin cancer models that mirrored tumor growth, differentiation, and the invasive biology of the specific cancer cells, offering an outstanding in vitro tool for studying skin cancer [46].
Considering the function of macrophages in tumor development, Linde et al. established a 3D organotypic model containing these cells. The dermal compartment was prepared by producing a type I rat collagen matrix with fibroblasts, macrophages, or both, and then the SCC human cell line A-5RT3 was seeded onto the collagen scaffold. They showed that 3D cultures can be generated with macrophages and SCC allowing the study of macrophage activation in the context of skin tumors [47].
More recently, a scaffold-free spheroid model was created by Kochanek et al. For this, they seeded head and neck SCC cell lines in ultra-low attachment plates and cultured them for at least 3 days. Tumor spheroids were formed along with specific microenvironments displaying different growth phenotypes and metabolic activities between cells located in the outer and inner layers of tumor spheroids. This system closely recapitulated in vivo tumors and has the potential of becoming a platform for cancer drug screening [48].
In Vitro Models of BCC
Given their greater propensity for invasion and metastasis, models of cutaneous melanoma and SCC are the most abundant and diverse [5, 40]. Still, a few BCC in vitro models have been described in the literature, most of which are mainly focused on the establishment of a proper 2D culture method. For instance, Grando et al. cultured BCC biopsy fragments obtaining different types of cells: normal fibroblasts, keratinocytes, and epithelioid spindle-shaped cells. Trypsin was used to remove contaminating fibroblasts, and normal keratinocytes were eliminated by cornification after increasing calcium concentration. The remaining cells belonged to nodular BCC, as judged by their biphasic morphology, slow growth, soft agar growing capability, ultrastructural similarities to nodular BCC, and the expression of several BCC-related markers [49]. Likewise, Asada et al. cultured explants from BCC biopsies on mitomycin-treated 3T3 feeder layers. Spindle-shaped cells grew from most of the BCC explants, with differential expression of several antibodies when compared to normal keratinocytes. Furthermore, unlike healthy keratinocytes, BCC-like cells were able to form colonies after being cultured in soft agar [50]. Both works provided protocols for the culture of BCC, which is the first step to obtaining a reliable model of human BCC.
As in vivo studies have revealed keratinization impairments in BCC tumor cells, Flaxman and Van Scott studied the keratinization of BCC cells grown in vitro. Like Grando et al. [49], they used biopsy explants from patients having multiple BCC, which were incubated in growth medium. Contrary to the in vivo situation, biopsy-derived cells presented a highly organized keratinization process, just as exhibited by their healthy counterpart. They concluded that BCC keratinization deficiency may be a reversible defect, and that if cells could keratinize in vivo, it could lead to the generation of keratin cysts instead of skin tumors [51]. However, given the difficulty of growing BCC cells in vitro, it is still possible that biopsy-derived cells belong to normal keratinocytes displaying a normal keratinization process.
In a more recent study, Hehlgans et al. used 2D and 3D cultures of BCC cell lines and examined the effect of the combination of vismodegib (a Hh pathway inhibitor) and radiation therapy. The 2D culture was maintained under standard conditions (37 °C and 5% CO2) while 3D models were created by diluting cells in a laminin-rich extracellular matrix that were then cultured for 24 h before treatment to allow colony formation. They found an increased sensitivity to radiation after vismodegib exposure, suggesting that combined therapy may increase therapeutic efficacy [52] and that spheroid-like models of BCC could be used as platforms for drug testing.
Regarding 3D models, a full-thickness equivalent was developed by Mahmudur to investigate the role of PTCH1 in BCC biology. The dermis component was produced with Matrigel, collagen, and fibroblasts and a keratinocyte cell line (NEB1) with PTCH1 suppression was grown on top of the Matrigel layer. However, there was no recapitulation of BCC characteristics in the 3D model [3], meaning that more than PTCH1 suppression may be needed to properly model BCC at the in vitro level. As previously mentioned, Bouez et al. developed a full-thickness model showing some SCC and BCC typical characteristics, whose dermal part was composed of fibroblasts and chitosan-collagen-GAG, while the epidermal component consisted of LOX-silenced HaCat cells [45].
Table 3 summarizes the characteristics of different in vitro BCC models.
Discussion
Several models have been developed to study normal and diseased skin and provide an important tool to study pathology progression and development. Here, we have presented an overview of the three main types of skin models: in vivo, ex vivo, and in vitro.
In vivo models are useful tools to understand highly complex skin processes, including immune and toxicological responses to external stimuli and tumor progression and spread. Moreover, they allow the evaluation of the efficacy of new therapeutic intervention strategies. Animal skin, however, presents structural and genetic differences from human skin, hampering the correct prediction in the human context.
Ex vivo models are highly suitable to study human skin biology, as they contain all the major cellular and structural components of human skin. Their main disadvantage is the requirement of constant tissue supply and the donor variation they introduce.
In vitro models are a powerful alternative, as they are of human origin, do not require a continuous supply of biopsies, and contribute to the reduction of animal testing. Recent efforts have led to the development of different models of healthy and diseased skin. Many of them are represented by 2D cultures, only involve the use of cell lines, or do not take into account the interaction between the tumor and its micro-environment, even though this interaction is crucial in tumor growth and metastasis [1, 31]. The challenge now is to culture cells with malignant potential in the context of a 3D network of healthy cells. Additionally, models including immunological components could open the avenues for systems in which malignant cell responses are also affected by the immunological human context, allowing a closer representation of reality.
Conclusion
In vitro models are a powerful and promising tool to increase our knowledge of many biological properties of human skin. Still, the methods and technology have to evolve in order to better reflect the in vivo situation. As no model reproduces all the features of the skin, the system of choice should represent the specific condition to be studied, which is easily achievable thanks to the customizable properties of 3D skin models.
References
Bergers LIJC, et al. Immune-competent human skin disease models. Drug Discov Today. 2016;21(9):1479–88. https://doi.org/10.1016/j.drudis.2016.05.008.
WebMD. “Skin cancer”. 2022. https://www.webmd.com/melanoma-skin-cancer/guide/skin-cancer#1-1. Accessed 7 Apr 2022.
Mahmudur M. “Characterisation of a novel in vitro model of Basal cell carcinoma (BCC) through stable PTCH1 suppression in immortalised human keratinocytes,” Queen Mary Unoversity of London, 2013.
World Health Organization. “Radiation: Ultraviolet (UV) radiation and skin cancer,” 2017. https://www.who.int/news-room/q-a-detail/radiation-ultraviolet-(uv)-radiation-and-skin-cancer. Accessed 7 Aug 2021.
Odashiro AN, Pereira PR, Marshall JC, Godeiro K, Burnier MN. Skin cancer models. Drug Discov Today Dis Model. 2005;2(1):71–5. https://doi.org/10.1016/j.ddmod.2005.05.011.
Neagu M, et al. Chemically induced skin carcinogenesis: updates in experimental models (Review). Oncol Rep. 2016;35(5):2516–28. https://doi.org/10.3892/or.2016.4683.
Hill DS, Robinson NDP, Caley MP, Chen M, Toole EAO, Jane L. A novel fully-humanised 3D skin equivalent to model early melanoma invasion. Mol Cancer Ther. 2016;14(11):2665–73. https://doi.org/10.1158/1535-7163.MCT-15-0394.A.
Skin Cancer Foundation. “Melanoma overview,” 2022. https://www.skincancer.org/skin-cancer-information/melanoma. Accessed 7 Apr 2022.
Skin Cancer Foundation. “Skin cancer facts & statistics,” 2022. https://www.skincancer.org/skin-cancer-information/skin-cancer-facts/. Accessed 30 Feb 2021.
Gober MD, Bashir HM, Seykora JT. Reconstructing skin cancers using animal models. Cancer Metastasis Rev. 2013;32(1–2):123–8. https://doi.org/10.1007/s10555-012-9410-8.
Caulin C, et al. An inducible mouse model for skin cancer reveals distinct roles for gain- and loss-of-function p53 mutations. J Clin Invest. 2007;117(7):1893–901. https://doi.org/10.1172/JCI31721.
Canadian Cancer Society. “What is non-melanoma skin cancer?” 2018. http://www.cancer.ca/en/cancer-information/cancer-type/skin-non-melanoma/non-melanoma-skin-cancer/?region=on. Accessed 7 Apr 2018.
Samarasinghe V, Madan V. Nonmelanoma skin cancer. J Cutan Aesthet Surg. 2012;5(1):3–10. https://doi.org/10.4103/0974-2077.94323.
Wu X, Elkin EE, Marghoob AA. Burden of basal cell carcinoma in USA. Futur Oncol. 2015;11(22):2967–74. https://doi.org/10.2217/FON.15.180.
Noonan FP, et al. Neonatal sunburn and melanoma in mice. Nature. 2001;413(6853):271–2. https://doi.org/10.1038/35095108.
Huang P, Balmain A. “Modeling cutaneous squamous carcinoma development in the mouse,” Cold Spring Harb Perspect Med. 2014;1–24. https://doi.org/10.1101/cshperspect.a013623
Huang M-T, et al. Inhibitory effect of topical applications of nondenatured soymilk on the formation and growth of UVB-induced skin tumors. Oncol Res. 2004;14(7–8):387–97. https://doi.org/10.3727/0965040041292350.
Aszterbaum M, et al. Ultraviolet and ionizing radiation enhance the growth of BCCs and trichoblastomas in patched heterozygous knockout mice. Nat Med. 1999;5(11):1285–91. https://doi.org/10.1038/15242.
Oro AE, Higgins KM, Hu Z, Bonifas JM, Epstein EH, Scott MP. Basal cell carcinomas in mice overexpressing sonic hedgehog. Science. 1997;276(5313):817–21. https://doi.org/10.1126/science.276.5313.817.
Xie J, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature. 1998;391(6662):90–2. https://doi.org/10.1038/34201.
Nilsson M, et al. Induction of basal cell carcinomas and trichoepitheliomas in mice overexpressing GLI-1. Proc Natl Acad Sci. 2000;97(7):3438–43. https://doi.org/10.1073/pnas.050467397.
Grachtchouk M, et al. Basal cell carcinomas in mice overexpressing Gli2 in skin. Nat Genet. 2000;24(3):216–7. https://doi.org/10.1038/73417.
Nicolas M, et al. Notch1 functions as a tumor suppressor in mouse skin. Nat Genet. 2003;33(3):416–21. https://doi.org/10.1038/ng1099.
Hochberg M. “Experimental models for BCC,” J Dermatology Clin Res. 2013;1(1).
Carlson JA, Combates NJ, Stenn KS, Prouty SM. Anaplastic neoplasms arising from basal cellcarcinoma xenotransplants into SCID-beige mice. J Cutan Pathol. 2002;29(5):268–78. https://doi.org/10.1034/j.1600-0560.2002.290502.x.
Genoskin. “Ready-to-use ex vivo human skin models for cosmetic testing ,” 2018. https://www.genoskin.com/skin-models-cosmetic-testing/. Accessed 12 Dec 2019.
Abd E, et al. “Skin models for the testing of transdermal drugs,” Clinical Pharmacology: Advances and Applications, vol. 8. Dove Medical Press Ltd. 2016;163–176. https://doi.org/10.2147/CPAA.S64788.
Groeber F, Holeiter M, Hampel M, Hinderer S, Schenke-Layland K. Skin tissue engineering – in vivo and in vitro applications. Adv Drug Deliv Rev. 2011;63(4–5):352–66. https://doi.org/10.1016/j.addr.2011.01.005.
Poumay Y, Coquette A. Modelling the human epidermis in vitro: tools for basic and applied research. Arch Dermatol Res. 2007;298(8):361–9. https://doi.org/10.1007/s00403-006-0709-6.
Garlick JA. “Engineering skin to study human disease--tissue models for cancer biology and wound repair.,” Adv Biochem Eng Biotechnol. 2007;03(207–239). https://doi.org/10.1007/b137206.
Genoskin. “Genoskin presents new skin cancer model at Meet2Win,” 2015. https://www.genoskin.com/en/tissue-samples/skin-cancer-model-oncoskin/. Accessed 7 Apr 2022.
Eves P, et al. Characterization of an in vitro model of human melanoma invasion based on reconstructed human skin. Br J Dermatol. 2000;142(2):210–22. https://doi.org/10.1046/j.1365-2133.2000.03287.x.
Meier F, et al. Human melanoma progression in skin reconstructs : biological significance of bFGF. Am J Pathol. 2000;156(1):193–200. https://doi.org/10.1016/S0002-9440(10)64719-0.
Zhang T, et al. Migration of cytotoxic T lymphocytes toward melanoma cells in three-dimensional organotypic culture is dependent on CCL2 and CCR4. Eur J Immunol. 2006;36(2):457–67. https://doi.org/10.1002/eji.200526208.
Vörsmann H, et al. Development of a human three-dimensional organotypic skin-melanoma spheroid model for in vitro drug testing. Cell Death Dis. 2013;4(7):719–719. https://doi.org/10.1038/cddis.2013.249.
Leight JL, Tokuda EY, Jones CE, Lin AJ, Anseth KS. Multifunctional bioscaffolds for 3D culture of melanoma cells reveal increased MMP activity and migration with BRAF kinase inhibition. Proc Natl Acad Sci U S A. 2015;112(17):5366–71. https://doi.org/10.1073/pnas.1505662112.
Kaur G, Dufour JM. Cell lines: valuable tools or useless artifacts. Spermatogenesis. 2012;2(1):1–5. https://doi.org/10.4161/spmg.19885.
Murekatete B, et al. Targeting insulin-like growth factor-I and extracellular matrix interactions in melanoma progression. Sci Rep. 2018;8(1):583. https://doi.org/10.1038/S41598-017-19073-4.
Duan J, et al. 3D bioprinted GelMA/PEGDA hybrid scaffold for establishing in-vitro model of melanoma. J Microbiol Biotechnol. 2022;32(4):1–10. https://doi.org/10.4014/JMB.2111.11003.
Semlin L, Schäfer-Korting M, Borelli C, Korting HC. In vitro models for human skin disease. Drug Discov Today. 2011;16(3–4):132–9. https://doi.org/10.1016/j.drudis.2010.12.001.
Borchers AH, Steinbauer H, Schafer BS, Kramer M, Bowden GT, Fusenig NE. Fibroblast-directed expression and localization of 92-kDa type IV collagenase along the tumor-stroma interface in an in vitro three-dimensional model of human squamous cell carcinoma. Mol Carcinog. 1997;19(4):258–66. https://doi.org/10.1002/(SICI)1098-2744(199708)19:4%3c258::AID-MC7%3e3.0.CO;2-8.
Commandeur S, de Gruijl FR, Willemze R, Tensen CP, El Ghalbzouri A. An in vitro three-dimensional model of primary human cutaneous squamous cell carcinoma. Exp Dermatol. 2009;18(10):849–56. https://doi.org/10.1111/j.1600-0625.2009.00856.x.
Commandeur S, Van Drongelen V, De Gruijl FR, El Ghalbzouri A. Epidermal growth factor receptor activation and inhibition in 3D in vitro models of normal skin and human cutaneous squamous cell carcinoma. Cancer Sci. 2012;103(12):2120–6. https://doi.org/10.1111/cas.12026.
Brauchle E, Johannsen H, Nolan S, Thude S, Schenke-Layland K. Design and analysis of a squamous cell carcinoma in vitro model system. Biomaterials. 2013;34(30):7401–7. https://doi.org/10.1016/J.BIOMATERIALS.2013.06.016.
Bouez C, et al. The lysyl oxidase LOX is absent in basal and squamous cell carcinomas and its knockdown induces an invading phenotype in a skin equivalent model. Hum Cancer Biol. 2006;12(5):1463–9. https://doi.org/10.1158/1078-0432.CCR-05-1456.
Berning M, Prätzel-Wunder S, Bickenbach JR, Boukamp P. Three-dimensional in vitro skin and skin cancer models based on human fibroblast-derived matrix. Tissue Eng Part C Methods. 2015;21(9):958–70. https://doi.org/10.1089/ten.tec.2014.0698.
Linde N, Gutschalk CM, Hoffmann C, Yilmaz D, Mueller MM. Integrating macrophages into organotypic co-cultures: a 3D in vitro model to study tumor-associated macrophages. PLoS ONE. 2012;7(7): e40058. https://doi.org/10.1371/journal.pone.0040058.
Kochanek SJ, Close DA, Johnston PA. High content screening characterization of head and neck squamous cell carcinoma multicellular tumor spheroid cultures generated in 384-well ultra-low attachment plates to screen for better cancer drug leads. Assay Drug Dev Technol. 2019;17(1):17–36. https://doi.org/10.1089/ADT.2018.896/ASSET/IMAGES/LARGE/FIGURE7.JPEG.
Grando SA, Schofield OM, Skubitz AP, Kist DA, Zelickson BD, Zachary CB. Nodular basal cell carcinoma in vivo vs in vitro. Establishment of pure cell cultures, cytomorphologic characteristics, ultrastructure, immunophenotype, biosynthetic activities, and generation of antisera. Arch Dermatol. 1996;132(10):1185–93. https://doi.org/10.1001/archderm.132.10.1185.
Asada M, et al. Growth characteristics and differentiation of basal cell carcinoma in vitro–immunohistochemical, gel electrophoretic, and ultrastructural analysis. J Invest Dermatol. 1992;99(4):474–81. https://doi.org/10.1111/1523-1747.EP12616161.
Flaxman BA, Van Scott EJ. Keratinizationin vitro of cells from a basal cell carcinoma. JNCI J Natl Cancer Inst. 1968;40(2):411–22. https://doi.org/10.1093/jnci/40.2.411.
Hehlgans S, et al. “Radiation sensitization of basal cell and head and neck squamous cell carcinoma by the hedgehog pathway inhibitor vismodegib,” Int J Mol Sci. 2018;19:(9). https://doi.org/10.3390/IJMS19092485.
Acknowledgements
The authors would like to acknowledge IPS Universitaria (University of Antioquia) for giving the working place facilities. We would like to thank the Corporación de Ciencias Básicas Biomédicas professors and Floris Foijer for critically reading the manuscript and giving helpful feedback. The authors wish to thank Dr. Laura Lema Perez for reviewing their manuscript.
Funding
This work was supported by COLCIENCIAS (Project code 727–2015).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Literature research and writing were performed by Catalina Gaviria Agudelo. Luz Marina Restrepo critically revised the work and provided important feedback. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics Approval
This is a review article and no ethical approval is required.
Consent to Participate
This is a review article and no informed consent is required.
Consent for Publication
This is a review article and no consent to publish is required from participants.
Competing Interests
The authors declare no competing interests.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Gaviria Agudelo, C., Restrepo, L.M. Human Skin Cancer: an Overview Of Animal, Ex Vivo, and In Vitro Models. Curr Derm Rep 11, 168–177 (2022). https://doi.org/10.1007/s13671-022-00361-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13671-022-00361-w