
Abstract
Background
Sparc/osteonectin, cwcv and kazal-like domain proteoglycan 1 (SPOCK1) has been reported to function as an oncogene in a variety of cancer types. Increasing evidence suggests that SPOCK1 contributes to the metastatic cascade, including invasion, epithelial-mesenchymal transition (EMT) and micro-metastasis formation. As yet, however, the underlying mechanism is not clearly understood. Here, we evaluated the expression and clinicopathological significance of SPOCK1 in primary pancreatic cancer (PC) specimens and explored the mechanisms underlying SPOCK1-mediated PC cell growth and metastasis.
Methods
The clinical relevance of SPOCK1 was evaluated in 81 patients with PC. The effect of SPOCK1 on proliferation, cell cycle progression, EMT and metastasis was examined in vitro and in vivo. The molecular mechanisms involved in SPOCK1-mediated regulation of NF-κB-dependent EMT were assessed in PC cell lines.
Results
We found that SPOCK1 expression was increased in PC tissues and was associated with lymph node metastasis. Silencing or exogenous overexpression of SPOCK1 markedly altered the proliferation of PC cells through cell cycle transition. Overexpression of SPOCK1 promoted PC cell migration and invasion by regulating EMT progression. Moreover, we found that SPOCK1 contributes to EMT and metastasis by activating the NF-κB signalling pathway via direct interaction with IκBα. After NF-κB pathway inhibition by BAY11–7082, we found that PC cell motility and EMT induced by SPOCK1 were reversed.
Conclusion
From our data we conclude that SPOCK1 promotes PC metastasis via NF-κB-dependent EMT by interacting with IκBα. This newly identified mechanism may provide novel clues for the (targeted) treatment of PC patients.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Pancreatic cancer (PC) is a devastating disease with a 5-year overall survival rate of approximately 9%. It is the fourth leading cause of cancer-related death in the United States [1]. The low survival rate is attributed to the fact that this extremely malignant type of tumour frequently exhibits rapid progression with less visible symptoms in early disease stages [2]. Once PC is clinically apparent, the primary tumour may already have invaded adjacent tissues and/or metastasized to distant organs to become unresectable [3]. Aproximately 90% of PC cases exhibit distant metastases [4]. One of the molecular mechanisms that promote the metastatic spread of PC is epithelial-mesenchymal transition (EMT) [5]. Shin et al. observed a significantly better overall survival in PC patients with a high E-cadherin expression, and Javle et al. identified EMT as a significant marker for a poor prognosis in PC patients who underwent surgical resection [6]. EMT may be involved in the initial steps of the metastatic cascade, including invasion, intravasation and micro-metastasis formation [7]. Therefore, it is of great importance to investigate the molecular mechanism underlying EMT in PC.
Sparc/osteonectin, cwcv, and kazal-like domains proteoglycan 1 (SPOCK1; also known as testican-1), for which the gene is located on chromosome 5q31.2, was first defined as an unnamed chondroitin/heparan sulfate proteoglycan in seminal plasma [8]. SPOCK1, along with testican-2 and testican-3, belongs to a matricellular protein family named secreted protein, acidic, cysteine-rich (SPARC). Members of the SPARC family share an extracellular calcium-binding motif, regulate extracellular matrix assembly and deposition and extracellular protease activity, and modulate growth factor/cytokine signalling pathways [9, 10]. SPOCK1 has been found to be overexpressed in gallbladder and prostate cancer [11, 12], whereas in lung and breast cancer SPOCK1 has been reported to serve as a target of transforming growth factor-β (TGF-β) that regulates cell migration and invasion by inducing EMT [13, 14]. Moreover, SPOCK1-mediated EMT signaling has been found to confer resistance to lapatinib in HER2-positive gastric cancer [15]. Yang et al. found that SPOCK1 can promote the proliferation, migration and invasion of glioma cells by activating the PI3K/AKT and Wnt/β-catenin pathways [16]. Others reported that SPOCK1 can promote the growth of osteosarcoma cells through the mTOR-S6K signalling pathway [17]. All these studies suggest a previously unappreciated role of SPOCK1 in cancer formation and development. However, to date, the detailed role of SPOCK1 in PC remains unclear.
Nuclear factor kappa B (NF-κB) is a critical transcription factor that regulates many cellular and organismal processes, including cell survival and invasion, and enhances angiogenesis via vascular endothelial growth factor (VEGF) expression [18]. Aberrant NF-κB activity is considered one of the hallmarks of cancer and chronic inflammatory diseases. In resting cells, NF-κB resides as an inactive form within the cytoplasm through the inhibitory actions of IκB proteins [19]. Under specific circumstances, inflammatory stimuli may induce IκB kinase β (IKKβ) activation via phosphorylation of two critical serine residues, Ser177 and Ser181. IKKβ in turn can phosphorylate IκBα at the N-terminal Ser32 and Ser36, which allows IκBα degradation by the ubiquitin-proteasome system [20]. These reactions result in nuclear translocation of NF-κB (p65 phosphorylation) and subsequent binding to its cognate κB sites in the promoters of target genes [21]. More importantly, NF-κB activation has been reported by Chiao et al. in 70% of PC specimens and 80% of PC cell lines [22]. Recently, accumulating evidence indicated that NF-κB activation and its target genes are required for advanced and metastatic PC progression [23,24,25]. Thus, understanding how the NF-κB pathway is regulated during PC development is considered important for finding reliable anti-metastatic strategies to treat PC.
In this study, we provide evidence that SPOCK1 acts as an oncogene in PC and that its oncogenic role is correlated with its regulation of the cell cycle and EMT characteristics. In particular, using human PC models, we observed a positive association between SPOCK1 and the NF-κB signalling pathway. Mechanistically, we found that SPOCK1 activates the NF-κB signalling pathway by regulating IκBα stabilization to subsequently allow NF-κB translocation from the cytoplasm to the nucleus. Consequently, this study links SPOCK1-IκBα-NF-κB signalling to PC progression and identifies SPOCK1 as a valuable prognostic biomarker and therapeutic target.
2 Materials and methods
2.1 Cell lines and culture conditions
The human PC cell lines BxPC-3, PANC-1, MIA PaCa-2 and Capan-1 were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA). BxPC-3, MIA PaCa-2 and PANC-1 cells were cultured in Dulbecco’s modified essential medium (DMEM) and Capan-1 cells were cultured in RPMI-1640 medium. The media were supplemented with 10% fetal bovine serum (FBS). The cells were incubated at 37 °C in a humidified chamber with 5% CO2 and were used within 10 passages after thawing of the stock cultures. MG-132 proteasome inhibitor (A2585) and BAY11–7082 IKK inhibitor (A4210) were purchased from APExBIO (Houston, TX, USA) and used at the indicated concentrations.
2.2 Plasmid construction and transfection
Lentiviral vector constructs containing SPOCK1 shRNA (shSPOCK1) and negative control shRNA (shRNA-NC) were designed and synthesized by GeneChem (GeneChem Co. Ltd., Shanghai, China). The target sequences of SPOCK1 shRNA are: 5′-TTTCGAGACGATGATTATT-3′ (sh#1), 5′-GCTGGATGACCTAGAATAT-′ (sh#2), 5′-TCTCCTGGAGCAATATAT-3′ (sh#3). The corresponding vector was hU6-MCS-Ubiquitin-EGFP-IRES-puromycin. For exogenous SPOCK1 overexpression, full-length SPOCK1 or control sequences were cloned into a GV143 vector containing EGFP. Cells were treated with puromycin (2 μg/ml) to produce stably transfected cell lines for further experiments. The infection efficiencies were based on GFP expression.
2.3 Flow cytometry-based cell cycle analysis
For cell cycle analysis, PC cells were seeded and grown in 10 cm cell culture dishes for up to 48 h. Then, approximately 1 × 106 cells were collected and fixed in 70% ice-cold ethanol at −20 °C overnight. After washing with cold phosphate-buffered saline (PBS), the cells were stained with PI/RNase staining buffer (550,825, BD, CA, USA) for 30 min at room temperature in the dark. Finally, the cells were run on a flow cytometer (BD Accuri C6, CA, USA) after which cell cycle distributions were analysed using Modfit LT4.1 Software (Verity Software House, Inc., Topsham, ME, USA).
2.4 Immunofluorescence assay
For immunostaining, PC cells were seeded on glass coverslips and allowed to grow for 48 h. Then, the cells were fixed with 4% paraformaldehyde for 10 min and permeabilized with 0.5% Triton X-100 for 10 min. After blocking with 3% bovine serum albumin fraction V (A8020, Solarbio, Beijing, China), the cells were incubated with primary antibody at 4 °C overnight. Next, the cells were washed twice with PBS and incubated with Alexa Fluor 488 goat anti-rabbit IgG (A-11008, 1:400, Invitrogen, CA, USA) or anti-mouse IgG (A-11001, 1:400, Invitrogen, CA, USA) for 2 h at room temperature (RT). Finally, the coverslips were mounted with Anti-fade Mounting Medium (P0126; Beyotime, Shanghai, China). Images of immunostained cells were captured using a Leica SP5II confocal microscope (Leica, Germany).
2.5 Migration and invasion assays
Cell migration and invasion assays were conducted using 24-well plates and 8-μm Transwell inserts (3422, Corning Life Science, MA, USA) with or without coated Matrigel (356,234, Corning Life Science, MA, USA). 4 × 104 cells were seeded in the upper chambers in serum-free media, and the lower chambers were filled with 600 μl medium supplemented with 10% FBS. After 48 h of culture, non-migrated or non-invaded cells remaining in the upper chamber were removed with cotton swabs, whereas those that had migrated or invaded to the lower membrane surfaces were fixed in methanol and stained with Giemsa Stain Solution for 1 h. Migrated or invaded cells were photographed and counted using an Olympus BX53 microscope (Olympus, Japan).
2.6 Dual-luciferase reporter assay
Cells were seeded in 24-well plates at 60–80% confluence on the day of transfection. The cells were co-transfected with a mixture of 1 μg pGL4.32 [lucNF-κB-RE/Hygro] (firefly luciferase) reporter vector (E8491, Promega, WI, USA) and 0.1 μg [pRL-TK] (Renilla luciferase) vector (E2241, Promega, WI, USA) using Lipofectamine™ 3000 Reagent (L3000015, Thermo Fisher Scientific, CA, USA) according to the manufacturer’s instructions. 24 h after transfection, the cells were lysed with 100 μl 1× passive lysis buffer (16,185, Promega, WI, USA) after which the cell lysates were analysed for luciferase expression using a dual luciferase assay kit (16,185, Promega, WI, USA) in a Tecan Spark™ 10 M luminometer. Firefly luciferase data were obtained by normalizing luciferase activity to Renilla luciferase activity. The average values of the tested constructs were calculated as fold change of the activity of the empty vectors. All transfection experiments were performed in triplicate for each group and repeated independently three times.
2.7 Co-immunoprecipitation and ubiquitination assays
Total lysates from BxPC-3 cells (500 μg) were harvested using RIPA buffer (CW2334, CWBIO, Beijing, China) supplemented with protease inhibitors. For preclearing of the cell lysates, 0.5 μg normal IgG rabbit antibody (sc-2027, Santa Cruz Biotechnology, TX, USA) and 25 μl Protein A/G PLUS-Agarose beads (sc-2003, Santa Cruz Biotechnology, TX, USA) were incubated with the lysates for 1 h at 4 °C. Then, 2 μg anti-SPOCK1 or normal IgG rabbit antibodies were added to the precleared lysates, after which the mixtures were incubated overnight at 4 °C under rotation. The following day, the protein complexes were cross-linked with 25 μl agarose beads at 4 °C with rotation for an additional 4 h. After three washes with PBS, the immunoprecipitates were eluted with 2x loading buffer and then subjected to SDS-PAGE and Western blot analysis. For ubiquitinated protein identification, the cells were treated with 20 μM MG132 for 6 h and lysed in an appropriate volume of lysis buffer. The lysates of the control group and SPOCK1-knockdown group were subjected to immunoprecipitation as described above. Thereafter, Western blots were probed using an anti-ubiquitin antibody.
2.8 Antibodies
The following primary antibodies were used: anti-β-actin (1:5000, 60,008–1-Ig, Proteintech, IL, USA), anti-SPOCK1 (1:1000, NBPI-30603, Novusbio, CO, USA), anti-Histone-H3 (1:1000, 17,168–1-AP, Proteintech, IL, USA), anti-IKKα (1:1000, 11,930, CST, MA, USA), anti-IKKβ (1:1000, 8943 s, CST, MA, USA), anti-phospho-IKKα/β Ser176/180 (1:1000, 2697 s, CST, MA, USA), anti-NF-κB p65 (1:1000, 8242 s, CST, MA, USA), anti-phospho-NF-κB p65 Ser536 (1:1000, 3033 s, CST, MA, USA), anti-IκBα (1:1000, 4814 s, CST, MA, USA), anti-phospho-IκB-α Ser32 (1:1000, 2859 s, CST, MA, USA), anti-phospho-NF-κBp65 Ser536 (1:200, ab86299, abcam, MA, USA), anti-p27 [Kip1] (1:2000, ab610242, abcam, MA, USA), anti-p21 [CDKN1A] (1:1000, 10,355–1-AP, Proteintech, IL, USA), anti-CDK1 (1:1000, 19,532–1-AP, Proteintech, IL, USA), anti-phospho-Cdc2 (1:1000, ENP0054, Elabscience, Wuhan, China), anti-Cdc25c (1:1000. ESAP11964, Elabscience, Wuhan, China), anti-Cyclin B1 (1:1000, 4138 s, CST, MA, USA), anti-Vimentin (1:1000, CBL202-K, Millipore, MA, USA), anti-E-cadherin (1:10000, ab610182, abcam, MA, USA), anti-Slug (1:1000, 3879 s, CST, MA, USA), anti-ZEB1 (1:1000, 21,544–1-AP, Proteintech, IL, USA), anti-ZEB2 (1:1000, 14,026–1-AP, Proteintech, IL, USA) and anti-Ubiquitin (1:1000, 10,201–2-AP, Proteintech, IL, USA).
2.9 Mouse xenograft model
Animal care and experimental procedures used in the present study were approved by the animal ethics committee of the Medical School of Yanbian University and performed in accordance with the institutional guidelines. For subcutaneous tumour models, cell suspensions with a 1:1 solution of PBS/Matrigel (2 × 106 cells) in a total volume of 100 μl (five mice per group) were subcutaneously inoculated into the left flanks of 6-week-old male BALB/c nude mice (Beijing SLC Inc., China). Tumour sizes were measured every 4 days from 7 days to 27 days after injection, after which the tumour volumes were calculated using the following formula: volume (mm3) = 1/2 × (width)2 × length. For in vivo lung metastasis models, 1 × 106 cells in 100 μl PBS (five mice per group) were intravenously injected via the tail vein into six-week-old male BALB/c nude mice. All mice were sacrificed 8 weeks after injection, after which the lungs were removed and fixed in 10% buffered formalin for further haematoxylin and eosin staining.
2.10 Patients and tissue specimens
To determine the clinical and prognostic significance of SPOCK1 expression in PC, tissue microarrays containing 81 PC tissues and 72 paired non-tumour tissues, along with complete clinicopathological data (including sex, age, tumour location and size, pathological grade, tumour stage, perineural invasion, lymph node metastasis, and follow-up data) were purchased from Shanghai Outdo Biotech (Shanghai, China). To compare the expression levels of SPOCK1 and phospho-p65 in PC tissues, microarrays comprising 193 PC tissues were purchased from Xian Alenabio (Xi’an, China). All samples were derived from formalin-fixed, paraffin embedded blocks, which were sliced into 4-μm-thick sections. This study was approved by the Ethical Committee of Yanbian University Medical College in China and was conducted in compliance with the tenets of the Declaration of Helsinki.
2.11 Immunohistochemistry assay
Cytoplasmic staining of tumour cells was considered indicative of anti-SPOCK1 staining, and nuclear staining of anti-phospho-p65 staining. The total immunohistochemical staining scores were estimated by calculating the staining intensity (graded as no obvious staining, 0; weak staining, 1; moderate staining, 2; and strong staining, 3) and extent of staining (percentage of positive cells: none or < 5% positive cells, 0; 5%–25% positive cells, 1; 26%–50% positive cells, 2; 51%–75% positive cells, 3; > 75% positive cells, 4). The sections were divided into low or high expression groups by multiplying the intensity and staining degree scores (range 0 to 12). Sections with a total score of 0 (negative) and 1–4 were classified as low expression (+), and sections with a total score of > 4 were classified as high expression (5 ~ 8: ++, 9 ~ 12: +++).
2.12 Statistical analysis
All data are expressed as the mean ± standard deviation of experiments performed at least in triplicate. For group comparisons, one-way analysis of variance (ANOVA) followed by Dunnett’s or Bonferroni’s post-test or Student’s t test was applied. The protein expression levels and clinicopathological parameters were compared by χ2 test. Correlations between gene expression levels were calculated using Spearman’s rank correlation coefficients. A two-sided p < 0.05 was considered to be statistically significant. All statistical analyses were carried out using the SPSS 17.0 statistical software package (SPSS Inc., Chicago, IL, USA) and GraphPad Prism version 6.0 (GraphPad, San Diego, CA, USA).
3 Results
3.1 SPOCK1 expression is increased and associated with lymph node metastasis in PC
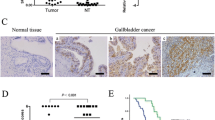
To gain more insight into the specific roles of SPOCK1 in carcinogenesis, we analysed the expression of SPOCK1 across seven different types of cancer according to the cBioProtal/TCGA datasets. Bioinformatic analysis revealed that the mRNA expression levels of SPOCK1 showed similar patterns in multiple sets of cancer, indicating the importance of SPOCK1 in driving malignant cancer cell behaviour (Fig. 1A). Next, we analysed the mRNA expression levels of SPOCK1 in ONCOMINE validation data sets, including two different sets of PC tissues and normal pancreatic tissues. We found that the mRNA expression levels of SPOCK1 were upregulated in PC tissues compared to their corresponding normal tissues (Fig. 1B). Subsequent survival analysis based on the KM plotter database showed that PC patients with a high SPOCK1 mRNA expression exhibited a shorter survival than those with a low expression (Fig. 1C).

SPOCK1 is overexpressed in PC tissues. (A) Multiple levels of SPOCK1 mRNA expression were observed in seven types of cancer using cBioPortal data analysis. (B) Significantly differential SPOCK1 mRNA expression levels in PC tissues compared with normal tissues using ONCOMINE data analysis. (C) SPOCK1 expression and overall survival of PC patients. (D) Immunohistochemical staining and statistical analysis of SPOCK1 protein expression in PC tissue samples. Positive rate: percentage of positive cases with +, ++, and +++ staining scores. Strongly positive rate: percentage of positive cases with ++ and +++ staining scores. ** p < 0.01 compared with normal pancreatic tissues. (E) Significant differential SPOCK1 expression levels in tumours with and without lymph node metastasis (** p < 0.01)
Subsequently, we examined SPOCK1 protein expression in a series of clinical PC samples using IHC staining. We found that SPOCK1 was expressed both in epithelia and stroma, and that SPOCK1 was highly expressed in the cytoplasm of the PC cells (Fig. 1D). The percentage of positive SPOCK1 staining was markedly higher in PC tissues (71.60%; 58/81) than in normal pancreatic tissues (31.94%; 23/72) (p < 0.01). Also, the percentage of strong positive staining for SPOCK1 was higher in PC tissues (55.56%; 45/81) than in normal pancreatic tissues (4.17%; 3/72) (p < 0.01; Fig. 1D). To explore the clinical significance of SPOCK1 expression in PC, we analysed the relationship between patient clinicopathological features and SPOCK1 status. We found that high SPOCK1 expression was significantly associated with lymph node metastasis (p < 0.01; Fig. 1E). These findings suggest that a high SPOCK1 expression correlates with an aggressive behaviour in certain types of cancer, including PC.
3.2 SPOCK1 accelerates the proliferation of PC cells
To study the potential oncogenic role of SPOCK1 in PC cells, we first surveyed the level of endogenous SPOCK1 expression in a series of PC cell lines (PANC-1, MIA PaCa-2, Capan-1, SW1990 and BxPC-3) by Western blotting. We found that BxPC-3 and PANC-1 cells showed a high SPOCK1 expression, while MIA PaCa-2, SW1990 and Capan-1 cells showed a low expression (Fig. 2A). Therefore, we chose BxPC-3 and PANC-1 cells for SPOCK1 knockdown and MIA PaCa-2 and Capan-1 cells for stable SPOCK1 overexpression. As described in Materials and methods, we used three shRNA sequences specific for SPOCK1 (designated sh1, sh2 and sh3), as well as scrambled shRNA. The efficacy and specificity of each shRNA in BxPC-3 and PANC-1 cells were verified by Western blotting. Similarly, successful transfection of MIA PaCa-2 and Capan-1 cells with stable vector overexpression was verified (Fig. 2B).

SPOCK1 regulates PC cell proliferation and cell cycle progression in vitro. (A) SPOCK1 protein expression levels in PC cell lines determined by Western blotting. β-Actin was used as internal control. (B) SPOCK1 knockdown and overexpression confirmed by Western blotting. β-Actin was used as internal control. (C) Effect of SPOCK1 on PC cell proliferation determined by MTT assay. (D) Effect of SPOCK1 on DNA replication determined by EdU incorporation assay. Original magnification 100×. (E) Effects of SPOCK1 knockdown or overexpression on colony growth determined by colony formation assay. (F) Effects of SPOCK1 knockdown or overexpression on cell cycle distribution determined by flow cytometry. (G-H) Western blot analysis of the expression of G2/M-related molecules in SPOCK1 knockdown, overexpression and control groups. β-Actin was used as internal control
Next, we examined the role of SPOCK1 in PC cell proliferation. Using MTT assays we found that SPOCK1 knockdown significantly suppressed the growth of BxPC-3 and PANC-1 cells. Conversely, we found that SPOCK1 overexpression slightly promoted the growth in MIA PaCa-2 and Capan-1 cells (Fig. 2C). In addition, we found by using an EdU incorporation assay that SPOCK1 knockdown inhibited DNA replication in BxPC-3 cells and that SPOCK1 overexpression increased DNA replication in Capan-1 cells (Fig. 2D). Using colony formation assays we found that the capacities of SPOCK1-knockdown cells to form foci were notably impaired compared with those of the controls, while SPOCK1 overexpression enhanced colony formation (Fig. 2E). These results indicate that elevated expression of SPOCK1 accelerates the proliferation of PC cells.
3.3 SPOCK1 regulates the PC cell cycle by modulating G2/M phase transition
Analysis of cell cycle progression by flow cytometry revealed that SPOCK1 knockdown in BxPC-3 cells induced an increase in the proportion of cells in G2/M phase and a decrease in the proportion of cells in G0/G1 phase, with no significant change in the number of S phase cells. In contrast, SPOCK1 overexpression in MIA PaCa-2 cells resulted in a significant decrease in the proportion of cells in G2/M phase and an increase in the proportion of cells in S phase (Fig. 2F). Given the effects of SPOCK1 on the G2/M phase, we further investigated the expression of proteins regulating cell cycle progression at the G2/M transition. We found that the levels of CDK1, Cyclin B1 and Cdc25c were decreased, which reduced the number of PC cells entering mitosis (Fig. 2G). Conversely, we found that the level of phospho-Cdc2 (p-Cdc2), i.e., negative phosphorylation of residues Tyr15 on CDK1 that maintains the CDK1-Cyclin B1 complex in an inactive form, was increased in SPOCK1 knockdown BxPC-3 and PANC-1 cells. These data suggest that reduced CDK1-Cyclin B1 activity is responsible for SPOCK1 knockdown-induced G2/M arrest in our model system. Moreover, the levels of p21WAF1/CIP1 and p27KIP1 (inhibitors of the CDK family) were found to be increased in SPOCK1 knockdown BxPC-3 and PANC-1 cells and inhibited further progression of DNA replication and the cell cycle. In contrast, we found that when SPOCK1 was overexpressed in MIA PaCa-2 and Capan-1 cells, the levels of CDK1 and Cyclin B1 were upregulated, while the levels of p-Cdc2, p21WAF1/CIP1 and p27KIP1 were downregulated (Fig. 2H). Additionally, we analysed correlations between SPOCK1 transcript levels and cell cycle related molecules using the GEPIA database (Supplement Fig.1A-C). Collectively, these data indicate that SPOCK1 promotes PC cell proliferation by modulating cell cycle progression at the G2/M transition phase.
3.4 SPOCK1 regulates migration, invasion and EMT of PC cells
To study the functional relevance of SPOCK1 in the metastasis of PC cells, we assessed the migratory capacity of PC cells using wound healing assays. We found that the wounds healed slower in SPOCK1 knockdown PANC-1 cells than in control cells. In contrast, we found that the wounds healed better and faster in SPOCK1 overexpressing MIA PaCa-2 cells than in control cells (Fig. 3A). We further tested the behaviour of PC cells using transwell migration and invasion assays. We found that SPOCK1 knockdown significantly abrogated the migrative and invasive abilities of PC cells, whereas SPOCK1 overexpression strongly promoted these abilities (Fig. 3B). Collectively, these results suggest that SPOCK1 regulates PC cell migration and invasion.

SPOCK1 induces PC cell migration and invasion. (A) Impact of SPOCK1 on the migration ability of PC cells determined by scratch wound-healing assay. Images of the wound areas were obtained at 0, 24 and 48 h after scratching. * p < 0.05, ** p < 0.01. (B) Effects of SPOCK1 on the migration and invasion abilities of PC cells measured by Transwell assays. Invaded cells were quantified by counting five randomly selected fields in each chamber and calculating the average number. * p < 0.05, ** p < 0.01. Original magnification 100×. (C-D) Aberrant expression levels of E-cadherin and Vimentin after SPOCK1 knockdown and overexpression determined by immunofluorescence. Original magnification 630×. (E-F) Representative Western blot images indicating positive regulation of EMT markers by SPOCK1. β-Actin was used as internal control
Since EMT plays an important role in cellular migration and invasion, we next questioned whether there is a link between SPOCK1 overexpression and EMT progression. Using confocal laser scanning microscopy of SPOCK1 knockdown BxPC-3 cells we observed a higher expression of the epithelial cell marker E-cadherin and a lower expression of the mesenchymal cell marker Vimentin compared to those in the corresponding control groups. Opposite results were obtained in SPOCK1 overexpressing Capan-1 cells (Fig. 3C-D). Moreover, we found that SPOCK1 knockdown not only significantly increased the expression of E-cadherin but also decreased the expression of Vimentin and EMT transcription factors such as Snail, Slug, ZEB1 and ZEB2 in BxPC-3 and PANC-1 cells (Fig. 3E). Conversely, overexpression of SPOCK1 in MIA PaCa-2 and Capan-1 cells reduced the expression of E-cadherin and induced the expression of Vimentin, Snail, Slug, ZEB1 and ZEB2 (Fig. 3F). Taken together, these data suggest that SPOCK1 promotes PC cell migration and invasion via EMT.
3.5 The oncogenic activity of SPOCK1 correlates with activation of the NF-κB pathway
Since a profound impact of the NF-κB signalling pathway on EMT progression has been reported in various cancers, we sought to investigate a putative association between this pathway and SPOCK1 expression. Activation of NF-κB is known to require translocation of its p65 subunit from the cytoplasm to the nucleus. We first assessed the expression levels of SPOCK1 and the activated nuclear form p-p65 (Ser536 phosphorylation of p65) in 193 pairs of PC and normal samples and found a positive correlation between these proteins (r = 0.392, p < 0.001) (Fig. 4A-B), suggesting that the NF-κB signalling pathway may be involved in cancer-promoting effects induced by SPOCK1. In addition, we surveyed nuclear localization of the p65 protein by immunofluorescence in SPOCK1 knockdown and SPOCK1 overexpressing cells. We found that SPOCK1 knockdown significantly reduced the expression of p-p65 and the nuclear accumulation of p65 in both BxPC-3 and PANC-1 cells, while SPOCK1 overexpression markedly elevated p-p65 expression and the nuclear accumulation of p65 in both MIA PaCa-2 and Capan-1 cells (Fig. 4C).

High levels of SPOCK1 expression activate the NF-κB pathway in PC cells. (A) Representative micrographs of SPOCK1 and phospho-p65 immunohistochemical staining in PC tissues (original magnification: left images 40×, right images 400×). (B) Correlation between SPOCK1 expression levels and phospho-p65 in 193 PC tissues analysed by Spearman rank correlation. (C) Immunofluorescence staining showing sub-cellular localization of phospho-p65 in different PC cells. Original magnification 630×. (D) NF-κB transcriptional activity in different PC cells measured by dual-luciferase reporter assay. * p < 0.05, ** p < 0.01. (E-F) Nuclear NF-κB, cytosolic NF-κB, IKKα, IKKβ, p-IKKα, p-IKKβ, IκBα, p-IκBα and SPOCK1 protein levels in SPOCK1 knockdown or overexpression groups compared to their corresponding control groups detected by Western blot analysis. Histone H3 or β-actin were used as internal controls. * p < 0.05, ** p < 0.01
Simultaneously, we found using a luciferase reporter assay that SPOCK1 knockdown reduced NF-κB transcriptional activity, while SPOCK1 overexpression effectively elevated the transcriptional activity of NF-κB (p < 0.01) (Fig. 4D). We also examined the phosphorylation of IKKα/β (Ser176/180) and IκBα (Ser32), which leads to the release of bound NF-κB for subsequent nuclear translocation. We found that SPOCK1 knockdown markedly suppressed the levels of phosphorylated IKKα/β (p-IKKα/β) and phosphorylated IκBα (p-IκBα) while inhibiting the levels of phosphorylated p65 (p-p65) (Fig. 4E and F). Conversely, we found that SPOCK1 overexpression markedly induced IKKα/β and IκBα phosphorylation with substantial nuclear accumulation of p65. Additionally, we analysed the correlation between SPOCK1 expression and molecules belonging to the NF-κB signalling pathway using the GEPIA database (Supplemental Fig. 1D and F). Overall, these results indicate that SPOCK1 may exert its oncogenic role via activation of the NF-κB pathway.
3.6 The SPOCK1/IκBα/NF-κB axis plays a role in the metastatic ability of PC cells
It is widely acknowledged that IκBα acts as an inhibitory protein involved in the NF-κB pathway. We hypothesized that SPOCK1 might activate the NF-κB pathway by interfering with IκBα protein stability. To better understand how SPOCK1 influences IκBα function, confocal scanning microscopy analysis was performed to determine the localization of SPOCK1 and IκBα in PC cells. We found that SPOCK1 was mainly localized in the cell membrane and that IκBα was localized both in the cell membrane and the cytoplasm of BxPC-3 and Capan-1 cells. Notably, SPOCK1 and IκBα were colocalized in the cell membrane (Fig. 5A). We also observed co-immunoprecipitation of IκBα with endogenous SPOCK1 in BxPC-3 cells using Western blotting (Fig. 5B). Decreased co-immunoprecipitation between SPOCK1 and IκBα was observed in SPOCK1 overexpressing Capan-1 cells (Fig. 5C), whereas the amount of IκBα co-immunoprecipitated with SPOCK1 was decreased in SPOCK1 knockdown cells (Fig. 5D). A major element of labile IκBα expression is ubiquitination and proteasomal degradation. This notion led us to examine whether SPOCK1 affects IκBα stabilization via ubiquitination-mediated degradation. As expected, we found that SPOCK1 knockdown successfully reduced IκBα ubiquitination under MG132 treatment (Fig. 5E). These results indicate that SPOCK1 regulates IκBα stabilization by interacting with IκBα in overlapping subcellular compartments.

High levels of SPOCK1 contribute to EMT and metastasis of PC cells by targeting IκBα/NF-κB/ZEB2 signalling. (A) Sub-cellular localization of SPOCK1 and IκBα in BxPC-3 and Capan-1 cells determined by immunofluorescence staining. Original magnification 630×. (B-C) Interaction of SPOCK1 and IκBα in BxPC-3 and SPOCK1 overexpressing Capan-1 cells determined by endogenous co-immunoprecipitation. Protein lysates were immunoprecipitated with an anti-SPOCK1 antibody or control IgG followed by Western blotting with an anti-IκBα antibody. (D) Effects of SPOCK1 knockdown on the binding affinities of SPOCK1 and IκBα in BxPC-3 cells. Protein extracts were immunoprecipitated with an anti-SPOCK1 antibody or control IgG followed by Western blotting with an anti-IκBα antibody. (E) Effects of SPOCK1 knockdown on the ubiquitination of IκBα. Cells in each group were treated with the proteasomal inhibitor MG132 (10 μM). Protein extracts were immunoprecipitated with an anti-IκBα antibody or control IgG followed by Western blotting with an anti-ubiquitin antibody. (F) The migration and invasion abilities of SPOCK1-overexpressing BxPC-3 and Capan-1 cells were effectively inhibited by BAY11–7082 (20 μM) as determined by Transwell assays. The invaded cells were quantified by counting five randomly selected fields in each chamber and calculating the average number, ** p < 0.01. Original magnification 100×. (G) Protein levels of EMT markers in BxPC-3 and Capan-1-SPOCK1 cells treated with BAY11–7082 (20 μM) compared to DMSO and negative control groups detected by Western blot analysis. * p < 0.05, ** p < 0.01. (H) Schematic model of the potential mechanism of SPOCK1-mediated PC cell proliferation and metastasis. On the one hand, overexpression of SPOCK1 promoted PC cell proliferation by activating the CDK1-Cyclin B1 complex and regulating the expression levels of p21WAF1/CIP1 and p27KIP1, which in turn influenced cell cycle progression at the G2/M transition point; on the other hand, overexpression of SPOCK1 contributed to EMT and metastasis of PC cells by targeting IκBα/NF-κB/ZEB2 signalling
To further confirm that SPOCK1 triggers the NF-κB pathway via IKKα/β and IκBα phosphorylation to promote EMT and PC cell metastasis, we used the IKK inhibitor BAY11–7082 to block NF-κB activity. We found that BAY11–7082 markedly suppressed the migration and invasion of BxPC-3 and Capan-1 cells (Fig. 5F). In addition, we found that BxPC-3 and SPOCK1 overexpressing Capan-1 cells underwent mesenchymal-epithelial transition (MET) in response to BAY11–7082 treatment. Unexpectedly, among the several transcription factors monitored, ZEB2 showed a markedly decreased expression in both BxPC-3 cells and SPOCK1 overexpressing Capan-1 cells. However, BAY11–7082 did not affect the expression level of SPOCK1 in PC cells (Fig. 5G). These data indicate that SPOCK1 contributes to EMT and metastasis of PC cells by targeting IκBα/NF-κB signalling and that ZEB2 may act as a downstream gene of NF-κB involved in SPOCK1-mediated tumorigenesis.
3.7 SPOCK1 facilitates PC cell growth and metastasis in vivo
We further assessed whether SPOCK1 can influence tumour progression in vivo. To this end, stably transduced BxPC-3-shRNA, BxPC-3-nc, Capan-1-Vector and Capan-1-SPOCK1 cells were subcutaneously injected into the left flanks of BALB/c nude mice. We found that the tumour volumes and sizes in the SPOCK1 knockdown group (BxPC-3-shRNA) were significantly smaller than those in the negative control group (BxPC-3-nc) (Fig. 6A). Conversely, we found that tumour growth was enhanced in the SPOCK1 overexpressing group (Capan-1-SPOCK1) compared to the control group (Capan-1-Vector). Lung colonization models were applied to investigate the role of SPOCK1 in the promotion of metastasis. To this end, BxPC-3-shRNA, BxPC-3-nc, Capan-1-Vector and Capan-1-SPOCK1 cells were inoculated into BALB/c nude mice via lateral tail vein injection. Remarkably, we found that the tumour growth inhibitory effect paralleled that of the decreased metastatic potential in SPOCK1 knockdown cells. Compared to the mice in the control group, the mice in the SPOCK1 knockdown group showed a marked reduction in the incidence of lung metastasis. No significant difference in the number of lung nodules was found between the SPOCK1 overexpressing group and its control group, but the sizes of the lung nodules formed by SPOCK1 overexpressing cells were markedly larger than those of control cells (Fig. 6B). To corroborate the regulatory functions of SPOCK1 on PC cell proliferation, EMT and the NF-κB pathway in vivo, the expression of related proteins was analysed by IHC. We found that the expression levels of Ki67, SPOCK1 and p-p65 were markedly decreased in the SPOCK1 knockdown group compared to those in the control group, but were increased in the SPOCK1 overexpressing group compared with the control group. In addition, we found that the expression of E-cadherin was increased by SPOCK1 depletion but reduced by SPOCK1 overexpression (Fig. 6C). Collectively, these findings underscore the in vitro results, indicating that SPOCK1 facilitates pancreatic tumour growth and metastasis in vivo.

High levels of SPOCK1 promote PC cell growth and metastasis in vivo. (A) Photographs showing xenograft tumours dissected from nude mice after subcutaneous inoculation. Tumour growth curves and tumour weights of SPOCK1 knockdown and overexpression groups compared to their parallel control groups are shown (each group, n = 5). * p < 0.05, ** p < 0.01. (B) Left, upper: whole-lung images with metastatic foci, as indicated by the arrows. Left, lower: Haematoxylin and eosin staining of lung sections showing metastatic tumours. Right: Graphs representing the number of lung metastatic foci in the SPOCK1 knockdown and overexpression groups compared to their corresponding control groups (each group, n = 5), * p < 0.05. (C) Representative immunohistochemical staining images of Ki-67, SPOCK1, E-cadherin and phospho-NF-κB expression in xenograft tumours of the SPOCK1 knockdown and overexpression groups. Original magnification 200 ×
4 Discussion
The poor clinical outcome of PC is attributed to the notion that most PCs present with extremely aggressive properties, desmoplastic reactions, high metastatic potentials and refractoriness to traditional therapy [26]. Hence, further knowledge on the molecular mechanisms regulating the progression of PC cell growth and metastasis may help to uncover novel therapeutic targets and strategies. In the present study, we systematically investigated the biological role of SPOCK1 in PC. We found that SPOCK1 overexpression is significantly associated with the aggressive behaviour of PC cells and that SPOCK1 manifests its tumorigenic activities by activating the NF-κB pathway.
The rapid spread of cancer is a dynamic multistep process that mainly arises from activation of oncogenes, inactivation of tumour suppressor genes and other alterations caused by both internal and external factors. An increasing number of studies focuses on SPOCK1 in various malignancies, as its structure is similar to that of SPARC, which is well-known as an important candidate oncogene [11]. By using publicly available gene expression data, Veenstra et al. found that SPOCK1 was predominantly expressed in stroma, and that a high SPOCK1 mRNA expression correlated with a poor prognosis of pancreatic adenocarcinoma patients [27]. In contrast to Veenstra’s study, we found that SPOCK1 protein levels were more abundant in epithelia. This discrepancy may be due to differences in post-translational regulation of SPOCK1 mRNA and protein levels in stroma and tumour cells. In lung and gastric cancers, a high expression of SPOCK1 protein has also been found to be correlated with a poor prognosis [28, 29].
In this study, we found by comparing data between normal and cancerous tissues from the ONCOMINE database that the SPOCK1 mRNA levels were upregulated in PC tissues. In addition, through IHC examination, we found that the rates of positive and strongly positive SPOCK1 protein expressing cells were higher in PC tissues than in corresponding normal tissues (p < 0.01). More intriguingly, we found that SPOCK1 overexpression was significantly associated with lymph node metastasis (p < 0.01). Similar results have been reported for urothelial carcinoma [30]. These findings suggest that increased SPOCK1 expression contributes to the development and progression of PC.
It is well known that uncontrolled proliferation is a key hallmark of cancer cells, depending on how the cell cycle is dysregulated. Recently, it has been reported that SPOCK1 may accelerate cellular growth through cell cycle regulation. In prostate cancer, for example, SPOCK1-mediated promotion of cellular proliferation has been correlated with its modulation of cell cycle progression [31]. In colorectal cancer, it has been reported that SPOCK1 knockdown significantly inhibited cell growth and induced cell cycle arrest at the G2/M transition phase [32]. Similarly, we found that SPOCK1 downregulation suppressed PC cancer cell growth and that, conversely, SPOCK1 upregulation enhanced PC cancer cell growth. Subsequent cell cycle analysis revealed G2/M arrest in SPOCK1 knockdown BxPC-3 cells, and a decrease in the proportion of cells in G2/M phase in SPOCK1 overexpressing MIA PaCa-2 cells compared with their respective controls. In line with these results, Western blotting showed that SPOCK1 regulates the expression of G2/M-associated proteins such as CDK1, Cyclin B1, Cdc25c, p-Cdc2, p21WAF1/CIPI and p27KIPI in PC cells. Taken together, these results underscore the notion that SPOCK1 promotes PC cell proliferation by regulating cell cycle progression.
PC is renowned for its aggressive biological behaviour, which is characterized by early local invasion and metastasis. EMT is a complex process involved in tumour invasion and metastasis that is influenced by a variety of factors during which cancer cells usually lose cell-cell adhesions and engage in enhanced cell-matrix adhesions [33]. SPOCK1 belongs to the Ca2+-binding proteoglycan family, which links cellular interactions with growth factors, chemokines and matrix architecture. Some of these interactions endow cancer cells with survival advantage and invasive characteristics [34]. Thus, we speculate that SPOCK1 expression is closely related to invasion and metastasis by regulating EMT in PC cells. We found that the migration and invasion abilities of PC cells were inhibited by SPOCK1 knockdown and enhanced by SPOCK1 overexpression. Upon further Western blot analysis, we found that SPOCK1 induced expression of the mesenchymal marker Vimentin and reduced the expression of the epithelial marker E-cadherin. Different SPOCK1 expression levels also affected the expression of several transcriptional suppressors of E-cadherin, such as Snail, Slug, ZEB1 and ZEB2. In support of these results, it has been reported that SPOCK1 induces EMT-like morphological changes and actin cytoskeleton rearrangements in oesophageal squamous cell carcinoma [35]. An additional in vivo study of prostate cancer reported that apigenin could improve the survival of animals by targeting SPOCK1-Snail/Slug axis-mediated EMT [29]. Similar results were obtained with our xenograft nude mouse tumour models, suggesting a crucial role of SPOCK1 in the migration, invasion and induction of EMT in PC cells.
Recent work has shown that aberrant activation of NF-κB signalling is required for promoting and maintaining PC cell invasion by controlling EMT [36, 37]. As yet, however, the association between SPOCK1 activity and the NF-κB signalling pathway in PC has not been examined. Therefore, we set out to find a connection between NF-κB pathway activation and SPOCK1 expression in PC. We found that phosphorylated NF-κB, which is located in the nucleus, showed an expression pattern similar to that of SPOCK1 in both PC tissues and PC cell lines, suggesting a potential link between these two. Since translocation of the p65 subunit from the cytoplasm to the nucleus is a hallmark of NF-κB pathway activation, we monitored the nuclear accumulation of p65 following SPOCK1 overexpression and observed an increase. This observation was further supported by an increased transcriptional activity of NF-κB in response to SPOCK1 overexpression. Phosphorylation of IKKβ and IκBα is a crucial step facilitating translocation of the p65 subunit from the cytoplasm to the nucleus. Our analysis of the nuclear/cytoplasmic compartmentalization of IKKα/β-IκBα-NF-κB pathway proteins clearly showed that the overexpression of SPOCK1 induced IKKα/β, IκBα and NF-κB phosphorylation in PC cells. These findings indicate that the oncogenic activity of SPOCK1 is associated with activation of the NF-κB pathway.
Degradation of IκBα by the ubiquitination-proteasome system resulting in activation of the NF-κB pathway has been well investigated. Therefore, we speculated that SPOCK1 may facilitate IκBα degradation via the ubiquitination-proteasome system. We observed colocalization of SPOCK1 and IκBα in PC cells including an endogenous interaction between SPOCK1 and IκBα, and the interaction was reduced following SPOCK1 knockdown in PC cells. Importantly, we found that the level of ubiquitinated IκBα was decreased after SPOCK1 knockdown. Therefore, our data suggest that SPOCK1 controls IκBα levels via ubiquitin-mediated degradation. To further validate the requirement of the NF-κB pathway in SPOCK1-mediated PC tumorigenesis, we adopted a pharmacological approach. We found that treatment of SPOCK1 overexpressing PC cells with the IKK inhibitor BAY11–7082 functionally reversed the expression of EMT markers and altered PC cell motilities induced by SPOCK1 overexpression. Most interestingly we found that, among the several transcription factors examined, ZEB2 showed a remarkably abrogated expression in both SPOCK1 overexpressing cell lines tested. It has been reported that NF-κB acts as a central mediator of EMT, and induces several EMT-associated transcription factors, such as snail and ZEB1 [38, 39]. Our data indicate that NF-κB may target ZEB2 in SPOCK1-induced migration and invasion of PC cells. Further experiments are needed to investigate the potential mechanism underlying this response.
In conclusion, we found that SPOCK1 is overexpressed in PC tissues and is associated with the aggressive behaviour of PC cells. Overexpression of SPOCK1 promotes PC cell proliferation by regulating cell cycle transition at the G2/M phase. Overexpression of SPOCK1 contributes to EMT and metastasis of PC cells by targeting IκBα/NF-κB/ZEB2 signalling (Fig. 5H). We conclude that SPOCK1 may serve as a novel diagnostic and therapeutic target for managing PC patients.
Data availability
The data presented in this study are available in this article.
Abbreviations
- SPOCK1:
-
Sparc/osteonectin, cwcv, and kazal-like domains proteoglycan 1
- PC:
-
Pancreatic cancer
- EMT:
-
Epithelial-mesenchymal transition
- NF-κB:
-
Nuclear factor kappa B
- SPARC:
-
Secreted protein, acidic, cysteine-rich
References
R.L. Siegel, K.D. Miller, A. Jemal, Cancer statistics, 2019. CA Cancer J. Clin. 69, 7–34 (2019)
T. Kamisawa, L.D. Wood, T. Itoi, K. Takaori, Pancreatic cancer. Lancet 388, 73–85 (2016)
A. Vincent, J. Herman, R. Schulick, R.H. Hruban, M. Goggins, Pancreatic cancer. Lancet 378, 607–620 (2011)
G. Patman, Pancreatic cancer: From normal to metastases--a whole gamut of pancreatic organoids. Nat. Rev. Gastroenterol. Hepatol. 12, 61 (2015)
H. Ying, P. Dey, W. Yao, A.C. Kimmelman, G.F. Draetta, A. Maitra, R.A. DePinho, Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 30, 355–385 (2016)
S.J. Shin, K.O. Kim, M.K. Kim, K.H. Lee, M.S. Hyun, K.J. Kim, J.H. Choi, H.S. Song, Expression of E-cadherin and uPA and their association with the prognosis of pancreatic cancer. Jpn. J. Clin. Oncol. 35, 342–348 (2005)
K.R. Fischer, A. Durrans, S. Lee, J. Sheng, F. Li, S.T. Wong, H. Choi, T. El Rayes, S. Ryu, J. Troeger, R.F. Schwabe, L.T. Vahdat, N.K. Altorki, V. Mittal, D. Gao, Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 527, 472–476 (2015)
C.J. Edgell, M.A. BaSalamah, H.S. Marr, Testican-1: A differentially expressed proteoglycan with protease inhibiting activities. Int. Rev. Cytol. 236, 101–122 (2004)
A.D. Bradshaw, Diverse biological functions of the SPARC family of proteins. Int. J. Biochem. Cell Biol. 44, 480–488 (2012)
U. Hartmann, H. Hulsmann, J. Seul, S. Roll, H. Midani, I. Breloy, D. Hechler, R. Muller, M. Paulsson, Testican-3: A brain-specific proteoglycan member of the BM-40/SPARC/osteonectin family. J. Neurochem. 125, 399–409 (2013)
Y.J. Shu, H. Weng, Y.Y. Ye, Y.P. Hu, R.F. Bao, Y. Cao, X.A. Wang, F. Zhang, S.S. Xiang, H.F. Li, X.S. Wu, M.L. Li, L. Jiang, W. Lu, B.S. Han, Z.G. Jie, Y.B. Liu, SPOCK1 as a potential cancer prognostic marker promotes the proliferation and metastasis of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol. Cancer 14, 12 (2015)
A. Wlazlinski, R. Engers, M.J. Hoffmann, C. Hader, V. Jung, M. Muller, W.A. Schulz, Downregulation of several fibulin genes in prostate cancer. Prostate 67, 1770–1780 (2007)
L. Miao, Y. Wang, H. Xia, C. Yao, H. Cai, Y. Song, SPOCK1 is a novel transforming growth factor-beta target gene that regulates lung cancer cell epithelial-mesenchymal transition. Biochem. Biophys. Res. Commun. 440, 792–797 (2013)
R. Vishnubalaji, H. Shaath, E. Elkord, N.M. Alajez, Long non-coding RNA (lncRNA) transcriptional landscape in breast cancer identifies LINC01614 as non-favorable prognostic biomarker regulated by TGFbeta and focal adhesion kinase (FAK) signaling. Cell Death Dis. 5, 109 (2019)
H.P. Kim, S.W. Han, S.H. Song, E.G. Jeong, M.Y. Lee, D. Hwang, S.A. Im, Y.J. Bang, T.Y. Kim, Testican-1-mediated epithelial-mesenchymal transition signaling confers acquired resistance to lapatinib in HER2-positive gastric cancer. Oncogene 33, 3334–3341 (2014)
J. Yang, Q. Yang, J. Yu, X. Li, S. Yu, X. Zhang, SPOCK1 promotes the proliferation, migration and invasion of glioma cells through PI3K/AKT and Wnt/beta-catenin signaling pathways. Oncol. Rep. 35, 3566–3576 (2016)
Y. Wang, W. Wang, E. Qiu, SPOCK1 promotes the growth of osteosarcoma cells through mTOR-S6K signaling pathway. Biomed. Pharmacother. 95, 564–570 (2017)
P. Viatour, M.P. Merville, V. Bours, A. Chariot, Phosphorylation of NF-kappaB and IkappaB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 30, 43–52 (2005)
D.M. Rothwarf, E. Zandi, G. Natoli, M. Karin, IKK-gamma is an essential regulatory subunit of the IkappaB kinase complex. Nature 395, 297–300 (1998)
J.A. DiDonato, M. Hayakawa, D.M. Rothwarf, E. Zandi, M. Karin, A cytokine-responsive IkappaB kinase that activates the transcription factor NF-kappaB. Nature 388, 548–554 (1997)
H. Sakurai, H. Chiba, H. Miyoshi, T. Sugita, W. Toriumi, IkappaB kinases phosphorylate NF-kappaB p65 subunit on serine 536 in the transactivation domain. J. Biol. Chem. 274, 30353–30356 (1999)
W. Wang, J.L. Abbruzzese, D.B. Evans, L. Larry, K.R. Cleary, P.J. Chiao, The nuclear factor-kappa B RelA transcription factor is constitutively activated in human pancreatic adenocarcinoma cells. Clin. Cancer Res. 5, 119–127 (1999)
P. Yuan, X.H. He, Y.F. Rong, J. Cao, Y. Li, Y.P. Hu, Y. Liu, D. Li, W. Lou, M.F. Liu, KRAS/NF-kappaB/YY1/miR-489 signaling Axis controls pancreatic Cancer metastasis. Cancer Res. 77, 100–111 (2017)
A. Nomura, V.K. Gupta, P. Dauer, N.S. Sharma, V. Dudeja, N. Merchant, A.K. Saluja, S. Banerjee, NFkappaB-mediated invasiveness in CD133(+) pancreatic TICs is regulated by autocrine and paracrine activation of IL1 signaling. Mol. Cancer Res. 16, 162–172 (2018)
W. Wang, J.L. Abbruzzese, D.B. Evans, P.J. Chiao, Overexpression of urokinase-type plasminogen activator in pancreatic adenocarcinoma is regulated by constitutively activated RelA. Oncogene 18, 4554–4563 (1999)
A.A. Ashour, N. Gurbuz, S.N. Alpay, A.A. Abdel-Aziz, A.M. Mansour, L. Huo, B. Ozpolat, Elongation factor-2 kinase regulates TG2/beta1 integrin/Src/uPAR pathway and epithelial-mesenchymal transition mediating pancreatic cancer cells invasion. J. Cell. Mol. Med. 18, 2235–2251 (2014)
V.L. Veenstra, H. Damhofer, C. Waasdorp, A. Steins, H.M. Kocher, J.P. Medema, H.W. van Laarhoven, M.F. Bijlsma, Stromal SPOCK1 supports invasive pancreatic cancer growth. Mol. Oncol. 11, 1050–1064 (2017)
Y. Gao, M. Yu, M. Ma, Y. Zhuang, X. Qiu, Q. Zhao, J. Dai, H. Cai, X. Yan, SPOCK1 contributes to the third-generation EGFR tyrosine kinase inhibitors resistance in lung cancer. J. Cell. Biochem. 120, 12566–12573 (2019)
D. Chen, H. Zhou, G. Liu, Y. Zhao, G. Cao, Q. Liu, SPOCK1 promotes the invasion and metastasis of gastric cancer through slug-induced epithelial-mesenchymal transition. J. Cell. Mol. Med. 22, 797–807 (2018)
L.J. Ma, W.J. Wu, Y.H. Wang, T.F. Wu, P.I. Liang, I.W. Chang, H.L. He, C.F. Li, SPOCK1 overexpression confers a poor prognosis in urothelial carcinoma. J. Cancer 7, 467–476 (2016)
Q. Chen, Y.T. Yao, H. Xu, Y.B. Chen, M. Gu, Z.K. Cai, Z. Wang, SPOCK1 promotes tumor growth and metastasis in human prostate cancer. Drug Des. Devel. Ther. 10, 2311–2321 (2016)
P. Zhao, H.T. Guan, Z.J. Dai, Y.G. Ma, X.X. Liu, X.J. Wang, Knockdown of SPOCK1 inhibits the proliferation and invasion in colorectal Cancer cells by suppressing the PI3K/Akt pathway. Oncol. Res. 24, 437–445 (2016)
A.D. Rhim, E.T. Mirek, N.M. Aiello, A. Maitra, J.M. Bailey, F. McAllister, M. Reichert, G.L. Beatty, A.K. Rustgi, R.H. Vonderheide, S.D. Leach, B.Z. Stanger, EMT and dissemination precede pancreatic tumor formation. Cell 148, 349–361 (2012)
Y. Li, L. Chen, T.H. Chan, M. Liu, K.L. Kong, J.L. Qiu, Y. Li, Y.F. Yuan, X.Y. Guan, SPOCK1 is regulated by CHD1L and blocks apoptosis and promotes HCC cell invasiveness and metastasis in mice. Gastroenterology 144, 179–191 e174 (2013)
X. Song, P. Han, J. Liu, Y. Wang, D. Li, J. He, J. Gong, M. Li, W. Tu, W. Yan, M. Liu, H. Huang, D. Tian, J. Liao, Up-regulation of SPOCK1 induces epithelial-mesenchymal transition and promotes migration and invasion in esophageal squamous cell carcinoma. J. Mol. Histol. 46, 347–356 (2015)
Y. Wang, H. Xiong, D. Liu, C. Hill, A. Ertay, J. Li, Y. Zou, P. Miller, E. White, J. Downward, R.D. Goldin, X. Yuan, X. Lu, Autophagy inhibition specifically promotes epithelial-mesenchymal transition and invasion in RAS-mutated cancer cells. Autophagy 15, 886–899 (2019)
E. Karamitopoulou, I. Zlobec, B. Gloor, A. Kondi-Pafiti, A. Lugli, A. Perren, Loss of Raf-1 kinase inhibitor protein (RKIP) is strongly associated with high-grade tumor budding and correlates with an aggressive phenotype in pancreatic ductal adenocarcinoma (PDAC). J. Transl. Med. 11, 311 (2013)
H.L. Chua, P. Bhat-Nakshatri, S.E. Clare, A. Morimiya, S. Badve, H. Nakshatri, NF-kappaB represses E-cadherin expression and enhances epithelial to mesenchymal transition of mammary epithelial cells: Potential involvement of ZEB-1 and ZEB-2. Oncogene 26, 711–724 (2007)
Y. Wu, J. Deng, P.G. Rychahou, S. Qiu, B.M. Evers, B.P. Zhou, Stabilization of snail by NF-kappaB is required for inflammation-induced cell migration and invasion. Cancer Cell 15, 416–428 (2009)
Funding
This research was funded by grants from the National Natural Science Foundation of China (no. 82060484).
Author information
Authors and Affiliations
Contributions
Conceptualization, J.J.P.; Data curation and formal analysis, X.L.C., W.Y.X.; Funding acquisition, J.J.P.; Methodology, W.Y.X., S.H.W., Y.C; Project administration, J.J.P.; Software, W.Y.X.; Supervision, Z.H.L., X.L.Z.; Validation, Z.H. L., X.L.Z.; Visualization, Z.H.L.; Writing original draft, X.L.C.; Writing, review and editing, J.J.P. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All the patients were informed of sample collection and usage. The tissue samples were collected and used in accordance with approval by the Clinical Research Ethics Committee of Yanbian University.
Consent for publication
The authors declare that they agree to submit the article for publication.
Competing interests
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(PDF 183 kb)
Rights and permissions
About this article

Cite this article
Cui, X., Wang, Y., Lan, W. et al. SPOCK1 promotes metastasis in pancreatic cancer via NF-κB-dependent epithelial-mesenchymal transition by interacting with IκB-α. Cell Oncol. 45, 69–84 (2022). https://doi.org/10.1007/s13402-021-00652-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13402-021-00652-7