
Abstract
Throughout the distribution of the bottlenose dolphin Tursiops truncatus, the occurrence, abundance, local distribution, and residential patterns are highly variable according to the pressure of the habitat where sighting occurs. In Venezuela, a study has been developed for the first time describing these aspects for the western of Aragua State (central coast of the country). From 2004 to 2008, 100 field surveys (30 km long) were conducted; the daily encounter ratio ranged from 0.79 to 1.11 dolphins/survey (Permanova, P < 0.01); the abundance was 18.70 (± 9.95) dolphins/transect (relative abundance of 0.62 dolphins/km), registering an increase since April 2007 by income of individuals (pseudo-t, P < 0.05). The sightings covered 56.63 km2, 90% of study area. Eighty-six dolphins were photo-identified (identification of new dolphins was constant). Residency analysis was limited to September 2006–August 2007 (continuous sampling); 7 (20.59%) residents (part of a group of 12), 9 (26.47%) occasional visitors (“neighboring group” and “outsiders”), and 18 (52.94%) transients were found. The study area is an open coast with shelter (bay) and prey that houses a resident group (sighted throughout the study) that was occasionally visited by a “neighbor” and occasional groups between October and February, and by transients throughout the year. This resident group inhabits the main bay and the coastal axis, and was associated (~ 75% of sightings) with Stenella frontalis (sighted throughout the year). We point out the importance of the area for both research and conservation due to the presence of dolphins throughout the year.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The bottlenose dolphin Tursiops truncatus is widely distributed along the coast of tropical and temperate oceans, occupying habitats from inland coastal areas to pelagic areas (Shane et al. 1986; Wells and Scott 1999), considering the coastal and oceanic ecotypes (Leatherwood and Reeves 1982). As with all species, their occurrence, abundance, distribution, and residence patterns depend on factors such as habitat type (in this case, protected coast, open coast, and neritic), prey distribution, predation risk, and thermal and saline tolerances, among other factors (Shane et al. 1986; Wells and Scott 1999; Gowans et al. 2008). Therefore, throughout the distribution of the bottlenose dolphin in the coast, there is a wide variability of these ecological aspects in response to the environment of each locality.
The occurrence of T. truncatus can be from 5.32 dolphins/survey at the Turneffe Atoll (Belize) (Campbell et al. 2002) or 3–15 dolphins/survey at the Atol das Rocas (Brazil) (Baracho et al. 2007), whereas Defran and Weller (1999) in an open coast in San Diego (USA) found an occurrence/distance of 1.15 dolphins/km. On the other hand, the variability in abundance can be seen in the following findings: in open coasts of Kvarnerić (Greece), Bearzi et al. (1997) found a minimum of 0.13 dolphins/km2. Another example in open coast was in the Asinara National Park (Italy), where Lauriano et al. (2003) found 0.10 dolphins/km2. In the west Florida continental shelf (neritic) (USA), Griffin and Griffin (2004) found an abundance of 0.086 dolphins/km2. Whereas, in a protected coast of Sardinia (Italy), Díaz-López and Bernal Shirai (2007) found 0.27 dolphins/km2. Another protected coast as the Gulf of Amvrakikos (Greece), Bearzi et al. (2008) found 0.37 dolphins/km2. Finally, in open coasts of Mississippi Sound (USA), Pitchford et al. (2016) showed temporal variation ranging of 1.12 dolphins/km2 in spring 2012 to 0.27 dolphins/km2 in spring 2013; however, following the Deepwater Horizon oil spill, Mullin et al. (2017) showed an inshore abundance variation between 0.77 and 1.61 dolphins/km2.
In terms of range, an extension of 46 km2 along the open coasts in Plettenberg Bay have been determined (South Africa) (Saayman and Tayler 1973); 85 km2 have also been identified along closed coasts such as Sarasota (USA) (Wells et al. 1990) and New Zealand (Williams et al. 1993), and a larger extension (100 km2) was estimated along the open coast of Moray Bay (Scotland) (Wilson 1995; Wilson et al. 1999). Some of these ranges may be limited by depth-enhancing physiographic factors, such as underwater canals or canyons (Wells and Scott 2002).
The high variability in these ecological aspects also occurs in their residential pattern. The first report of residency (14 months) in San José (Argentina) was by Würsig and Würsig (1977). Then, later studies have shown that in the same locality, there may be dolphins that stay days, seasons, or years (Irvine et al. 1981; Hansen 1990; Wells 1991; Williams et al. 1993; Barco et al. 1999; Campbell et al. 2002; Zolman 2002; Díaz and Bernal 2013). For example, in the Stono River estuary (USA) from October 1994 to January 1996, Zolman (2002) found a 20.5% of resident individuals, 8.82% seasonal residents, and 70.58% transients. In open coasts of Veracruz (Mexico), Martínez-Serrano et al. (2011) found 12% of residents, 14% of occasional visitors, and 73% of transients. While in the wide Bay of St. George (Northern Gulf of California, México), Mellink and Orozco-Meyer (2006) found a 3% of the individuals were during all surveys, 7% in three surveys, 23% in two, and 67% in one.
In Venezuela, T. truncatus is found along the coast (Romero et al. 2001). In the central coast, from the western end of the State of Carabobo to Cabo Codera in Miranda State (~ 200 km), there is a predominance of rocky cliffs with a pronounced continental slope. This oceanic coast has weather and oceanographic dynamics that experience little changes (Novoa et al. 1998). As such, the coast of the state of Aragua is located to the west end of the state and has a constant presence of T. truncatus and Stenella frontalis (Acevedo 2007; Cobarrubia-Russo 2010). The objective of this study was to describe the occurrence, abundance, distribution, and residence patterns of T. truncatus within this study area, which may have an importance for preservation and researching abundance.
Materials and methods
Study area
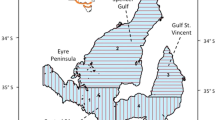
The study area is a coastal strip of ~ 30 km long and ~ 3 km wide, from Bahía de Turiamo (10° 28′ N, 67° 50′ W) to Puerto Colombia (10° 30′ N, 67° 36′ W). It has a sea surface temperature of 25 to 27 °C, a salinity of 34–36 ppm, a tidal regime of ± 24 cm, and two climatic seasons (dry: November–April, rainy: May–October) (Novoa et al. 1998). This coast has rocky cliffs and some sandy beaches with three types of habitats (Gowans et al. 2008): internal and external coastal habitats and neritic habitats. The first habitat involves semi-enclosed bays such as Bahía de Turiamo, La Ciénaga, and ElPlayón. The second habitat is from the coastline to the 100-m isobath, approximately from Cuyagua beach to the outside of the Bahía de Turiamo, progressively moving away from the coastline as it moves westward. The third habitat occupies an area that is limited to the south by the coastal axis and to the north by the 200-m isobath, and its predominant area is located towards the western end of the study area (Fig. 1).

Map of the study area (thin black line) and the predefined transect (thick black arrow) (IGVSB 2003)
Field surveys, sightings, and photo-ID
Due to logistical and fund constraints, the field surveys were opportunistic. These field surveys were carried out on a predefined sampling transect of 30 km, parallel to the coast to ~ 1.5 km. It started at the western end (Bahía de Turiamo), and it ended to the eastern end (Puerto Colombia) (Fig. 1). The mean speed of the vessel was approximately seven knots in a 9-m-long 45 HP outboard-powered boat with three observers on board (one on bow, one on port, and one on starboard).
When a dolphin group was encountered, we approached to a distance of approximately 10 m and recorded the following information: time of sighting, group size, and location (using a handheld global positioning system). A group was defined as all dolphins within a 100-m radius and that appeared to be coordinating their activities (Irvine et al. 1981). Group size was estimated between observers by visual counts during each sighting. After the initial encounter, an observer made the photo-identification. Dolphins were photographed individually with a DSLR camera fitted with an autofocus 18–200 or 18–250-mm zoom lens using large/fine resolution. Individual dolphins were identified by natural marks (permanent marks suitable for reliable long-term identification) on the body and the trailing edge of their dorsal fins (Würsig and Würsig 1977; Wilson et al. 1999). The photo-identification effort time depended on the size of the group and its behavior towards the vessel; at the end of the photo-identification, the boat returned to the transect to complete it in search of other groups.
Only high-quality photographs with clear identification of individuals were used for analysis (i.e., those in focus, with the dorsal fin perpendicular to the plane of the photograph, or close enough to identify small notches) (Díaz-López and Bernal Shirai 2007, 2008). A marked individual is one that is recognized not by a single feature, but by several features. Repeated sightings of individuals on the same day were treated as a single sighting. The photographs were analyzed and chosen by the eye of the three observers and then with support of the Finscan 1.6.1 program in order to minimize the identification errors (Hillman et al. 2003). Once identified, it was assigned the alpha-numeric code (D ##). Finally, the identified groups were assigned a letter from A to Z according to the chronological order of their identification.
Daily encounter ratio and abundance
To have an estimate of the occurrence of the species, the daily encounter ratio was calculated [daily encounter ratio (DER) = No. of sightings / Search effort (h)] (Díaz-López 2006). This process was also applied to determine the annual rate to assess possible differences. The abundance (mean ± SD) was estimated by the number of dolphins sighted in the transect (Abundance = No. of dolphins / distance). Then, DER and abundance were compared separately between years and between climatic seasons, in order to detect variations.
The data from each sampling were ordered by the years and the climatic season (dry, rainy) to which the samplings corresponded, and based on the different months, the samplings were assigned to one of the seasons. From these tables, Bray-Curtis matrices were created for the relative abundance and DER variables. To determine the differences and interactions between years and seasons, two-way variance analysis based on permutations (PERMANOVA) was used. The Permanova a posteriori comparisons (between pairs of combinations of year and season) were obtained from a pairwise test which uses the t statistic permuted (pseudo-t). In addition, the homogeneity test of dispersion also based on permutations (PERMDISP) was obtained, because the PERMANOVA test is sensitive to dispersion heterogeneity. The P values of all analyses were obtained from 9999 permutations, using PRIMER 6.1.13 and PERMANOVA1.0.3 software (Anderson et al. 2008).
Range (distribution of sightings)
The geographical location records of T. truncatus and the calculation of the area were made using a geographic information system (Arcview 3.3), drawn from the Ocumare map sheet of La Costa, No. 6647 (1:100,000) of the Simón Bolívar Geographical Institute and nautical chart no. 24450 (1:300,000) of the Hydrographic Agency of Defense and Mapping and the US Topographic Center.
Residence
Due to the restrictions mentioned, there were months without sampling. Nevertheless, from September 2006 to August 2007, continuous field surveys were conducted for 12 months and the analysis of residence was restricted to this period. The residence criterion considered two non-exclusive characteristics of sightings per dolphin: (1) the frequency of sighting [high (H) > 25% of surveys, medium (M) 10–25%, low (L) < 10%] (modified from Möller et al. 2002) and (2) the distribution of sightings throughout the year [annual (A): sightings in 7 (separated at least by 2 months minimum without sightings) to 12 months; periodic (P): sightings in 2–6 months mostly spaced throughout the year and/or clustered seasonally; sporadic (S): sightings in 1 month] (Table 1). A resident dolphin has a high frequency with an annual to periodic sighting distribution, or a medium frequency with annual distribution (H + A-P or M + A). An occasional dolphin has a medium frequency with periodic to sporadic distribution (M + P-S), or a low frequency with periodic distribution, and a transient dolphin has a low frequency with sporadic distribution (L + S) (Table 1).
Results
Field surveys, effort, and sightings
One hundred field surveys were conducted between May 2004 and October 2008 (total effort = 377.38 h), 6 in 2004 (4.99 ± 1.89 h), 15 in 2005 (3.77 ± 0.72 h), 19 in 2006 (4.32 ± 1.16 h), 42 in 2007 (3.38 ± 1.09 h), and 18 in 2008 (3.71 ± 1.59 h). Every field survey was conducted in 3.77 ± 1.28 h. Eighty-four sightings were recorded, 5 in 2004, 11 in 2005, 15 in 2006, 33 in 2007, and 20 in 2008 (Table 2).
Sightings by group and species
In 56 field surveys, T. truncatus sightings were recorded and 3360 photos were taken, of which 602 were photographs suitable for analysis. Of these sightings, 55 (65.48%) corresponded to a group (named A) of 11.95 (± 1.12) dolphins, 6 (7.14%) to an “outsider” group (B) of 4.5 (± 6.13) dolphins, 6 (7.14%) to a possible neighboring group (C) of 16.5 (± 0.5) dolphins, 4 (4.76%) to an outsider group (D) of 4.5 (± 3.07), and the rest corresponded to mixed sightings: 1 of A + B + Stenella frontalis, 5 of A + C, and 2 of A + C + S. frontalis. It should be noted that of the 84 sightings of T. truncatus, this other species was present in 61 (72.6%) (Cobarrubia-Russo 2010) (Fig. 2).

Sighting records of Tursiops truncatus
Daily encounter ratio and abundance
The DER was 0.47 ± 0.37 sightings/survey for 2004 (min), 1.37 ± 0.35 for 2005 (max) (Permanova, P < 0.01), 0.49 ± 0.27 for 2006, 0.72 ± 0.35 for 2007, and 0.64 ± 0.29 for 2008 (Fig. 3a). The maximum registered in 2005 that doubles the rest of the years, it was due to the few field surveys in which one recorded the highest number of sightings (04); however, this peak was not significant (pseudo-t, P > 0.05).

Annual mean of dolphins sighted by survey during the rainy and dry seasons (± SD)
Abundance had a stable trend from May 2004 to April 2007, which remained below 5.00. In this trend, small fluctuations were observed between periods, with a minimum between the 2006 (rainy season) and the 2007 (dry season), but these were not significant (pseudo-t, P > 0.05). However, as of the April 2007 (rainy), abundance increased abruptly (9.00) until it reached 11.00 (pseudo-t, P < 0.05) during the 2008 (dry) to decrease to 6.00 during the 2008 (rainy) (pseudo-t, P < 0.05) (Fig. 3). Dispersion homogeneity was found for both the interaction and the factors evaluated in each of the variables (Permdisp, P > 0.05), except for the year factor for the abundance variable (Permdisp, P < 0.01). This abrupt variation was due to incursions of groups (named B and C) during 2007 and 2008, and during 2008 (rainy), the number of transient individuals increased; in fact, in the accumulation curve of photo-identified individuals, there was an increase abrupt of 45 individuals to 78 between June and September (rainy) (Fig. 4).

Accumulative number of new dolphins during the study time
Range
The sightings of T. truncatus (n = 84) were distributed within 56.63 km2 encompassing the internal coastal, external coastal, and neritic habitats; this extension was 61.47% of the study area (~ 92.12 km2). Of these records, 76 (84%) were found in areas where the depth did not exceed the 100-m isobaths (Fig. 2).
Residence patterns
Along the study, 86 dolphins were identified. The curve of the accumulative number of new dolphins vs. time did not reach the asymptote (Fig. 4). The new dolphin identification rate was 0.66/survey. The maximum number of sighting per dolphin was 31, and the minimum was 1 (Fig. 5).

Frequencies distribution of sightings vs. number of dolphins
Since the analysis of residence was limited to 12 months of continuous and uniform sampling (September 2006–August 2007), of the 86 dolphins, 34 were identified in this period, of which 7 (20.59%) were categorized as residents (4 with H + A, 17–18 sightings on 10–11 months; 3 with M + A, 8–9 sightings on 7–11 months), 9 (26.47%) as occasional visitors (7 with M + P, 5–8 sightings on 4–6 months; 2 with L + P, 3–4 sightings on 3–2 months), and 18 (52.94%) as transients (L + S, 1–3 sightings on 2–1 months) (Table 3). These resident dolphins were part of the group (named A) of 11.95 ± 1.12 individuals (mother calves and subadults) sighted along the study, with individuals that reached between 16 and 31 sightings (Table 3). The rest of the non-photo-identified dolphins of this group were due to lack of marks or evasive behavior (Personal communication). All resident individuals had a similar sighting frequency between the rainy and drought seasons, except for individuals D5 (three sightings in rain, six in drought) and D7 (two in rain, six in drought) (Table 3).
Of the occasional dolphins, D8, D9, D10, D11, and D13 were part of a group (B) of five adults who visited the study area during the rainy season and eventually joined group A (between 2006 and 2008). Individuals D12 and D15 were often seen always with group A mainly during the dry season. The individual D16 was part of a “neighbor” group (named C) of 16.5 ± 0.5 individuals (mother calves and subadults) that during the dry season (2007 and 2008) entered the study area with possible mergers with group A (Personal communication). Finally, of the transient dolphins, the individuals D17, D18, and D19 also were members of the group C; D18 was sighted individually once during the rainy season. The rest of individuals were sighted one to two times in one season or another.
Discussion
Bottlenose dolphins have a wide distribution, occupying different habitats, where occurrence, abundance, range, and residence patterns can vary considerably (Díaz-López and Bernal Shirai 2007; Fruet et al. 2011; Defran and Weller 1999; Defran et al. 1999; Zhou and Qian 1985). This study confirms this high variability reported elsewhere; therefore, this species could modulate its behavior according to local environmental circumstances (Martínez-Serrano et al. 2011). This species was sighted in more than half of the field surveys, which was due to the presence of a group (A) and the constant income of other groups and individuals, which could be seen both separately and together, a characteristic of their social dynamics (Wells and Scott 2002). Therefore, the occurrence and abundance of T. truncatus showed a relative trend (by group A) that could be sensitive to abrupt changes due to seasonality and the opening of the coast (other groups), resembling patterns already observed off the coasts of California (USA) (Hansen 1990; Wells et al. 1990).
T. truncatus occupied more than 60% of this study area, being sighted more frequently in localities with less than 100-m depths which is relatively consistent with observations in other localities where the depth was a critical factor in the local distribution of the coastal form of this species where acoustics for navigation, prey detection, and predator avoidance seem to be advantageous (Shane et al. 1986; Martínez-Serrano et al. 2011). In the western coast of Aragua, this species seems to tolerate deeper areas, and perhaps because of this, it is usually associated with other species. This study could not cover the inner part of the Bahía de Turiamo because it is a military zone; however, in two sightings, we tracked the group to the entrance of the Bahía de Turiamo and we knew from the personnel of the Army Base that they used to sight the species at the end of the day and in the morning hours around the submarine dock. It is noteworthy that in the coastal axis, the continental slope is pronounced, with an exceptional presence of T. truncatus in the open and neritic habitats with depths of 30 to 100 m and with extraordinary sightings in deeper. This scenario resembles observations by Sayman and Tayler (1973) in open and neritic coastal habitats in South Africa, where dolphins frequented deep areas but in groups larger than 100 individuals; however, despite this similarity, it is necessary to emphasize that these individuals could be the oceanic ecotype (Leatherwood and Reeves 1982). In our case, although group size off the coast of Aragua matched the reported group size range of the coastal ecotype (Leatherwood and Reeves 1982; Shane et al. 1986), frequent associations were detected with groups or large aggregations of Stenella frontalis (whose group size was superior to that of T. truncatus) between El Playón y Cuyagua (Fig. 1) (Cobarrubia-Russo 2010). This association may be exerting an effect similar to that reported for the oceanic bottlenose dolphin that associates with other species to increase the probability of detection of prey and predators due to the considerable openness of the neritic habitat (Gowans et al. 2008).
The study area is an open coast with a large bay to the west, which houses a resident group, occasional visitors, and constant transients. The cumulative curve of identification of new dolphins never reached the asymptote; this was because the residents only represented the 20.59% of the photo-identified population. This scenario where residents represented a fifth of the sampled population was similar to the one reported by Zolman (2002) in the Stono River estuary (USA) (despite being a protected coast) and Quintana-Rizzo and Wells (2001) (estuarine semi-protected habitat). We expected results like those obtained by Martínez-Serrano et al. (2011) who found 12% of residents on an open-neritic coast of Veracruz (Mexico). Bouveroux et al. (2014) also found a 12% of residents although in a closed-open coastal habitat in Panama City (USA) while Simões-Lopes and Fabian (1999) in the Laguna channel and the lagoon system of Imarui-Santo Antonio (Brazil) found a 90% of resident population, and Fury and Harrison (2008) even though they studied T. aduncus in two subtropical estuaries of Australia found that residents accounted for 60 and 37% of the population. This portion of residents that make up group A seems to define a residence area between Bahía de Turiamo (refuge) and the coastal axis to the Cuyagua beach (prey) with relative boundaries according to how it looks in the density of sightings, as suggested by Shane (1986). The resident dolphins were represented by the group A that covered a large part of the study area, the occasional dolphins that were members of the group C (neighbor), and adults (B and D) who visited the study area between October and January (northern winter). The analysis underestimated the number of dolphins in groups A and C because not all the individuals were marked or get sufficiently close to the boat even though we tried to approach these evasive individuals which in response kept always distance. On the other hand, transitory dolphins formed the predominant residence pattern, as observed by Hubard et al. (2004) in the Mississippi Straits (USA). Finally, these results resemble studies on reproductive groups reporting that they remain in certain areas and are visited by dolphins from neighboring groups and breeding males (Shane et al. 1986; Félix 1997; Wells and Scott 2002). Based on our residency results, we propose that all dolphins in the western coast of Aragua State can be considered as a single resident community, according to the description made by Rossbach and Herzing (1999): “A resident community is a group of dolphins that includes both genders, shows long-term site-fidelity, relatively high association between members, and low association with neighboring individuals, and shares similar habitats and feeding habits.”
We have shown a coastal-oceanic landscape that is inhabited throughout the year by dolphins aggregated in group A and an additional group (herein called group C) that may be overlapping intermittently its range and that are subject to annual visits by adults during the year and by constant visits throughout the year by transient dolphins. This annual presence of groups of T. truncatus (and Stenella frontalis) makes this area important for conservation and a suitable field for developing prolonged studies on life histories, social organization, sympatry, and global change.
References
Acevedo R (2007) Potential geographical distribution of seven species of marine cetaceans reported in Venezuela, Southeast Caribbean. Acta Zool Sin 53(5):853–864
Anderson M, Gorley R, Clarke K (2008) PERMANOVA+for PRIMER: guide to software and statistical methods. PRIMER-E, Plymouth, p 214
Baracho C, Cipolotti S, Marcovaldi E, Apolinario M, Brito-Silva M (2007) The occurrence of bottlenose dolphins (Tursiops truncatus) in the biological reserve of Atol das Rocas in North-Eastern Brazil. Mar Biodivers Rec 1(75):1
Barco SG, Swingle WM, McLellan WA, Harris RN, Pabst DA (1999) Local abundance and distribution of bottlenose dolphins (Tursiops truncatus) in the nearshorewaters of Virginia Beach, Virginia. Mar Mamm Sci 15:394–408
Bearzi G, Agazzi S, Bonizzoni S, Costa M, Azzelino A (2008) Dolphins in a bottle: abundance, residency patterns and conservation of bottlenose dolphins Tursiops truncatus in the semi-closed eutrophic Amvrakikos Gulf Greece. Aq Cons Mar Fresh Ecosyst 18(2):130–146
Bearzi G, Notoarbatolo-di-Sciara G, Politi E (1997) Social ecology of bottlenose dolphins in Kvarneric (northern Adriatic Sea). Mar Mamm Sci 13:650–668
Bouveroux T, Tyson RB, Nowacek DP (2014) Abundance and site fidelity of bottlenose dolphins in coastal waters near Panama City, Florida. J Cet Manag 14:37–42
Campbell GS, Bilgre BA, Defran RH (2002) Bottlenose dolphins (Tursiops truncatus) in Turneffe Atoll, Belize: occurrence, site fidelity, group size, and abundance. Aquat Mamm 28(2):170–180
Cobarrubia-Russo S (2010) Ecología y comportamiento del delfín nariz de botella Tursiops truncatus en la costa del Estado Aragua. Master's Thesis. Simon Bolivar University. Caracas, p 30–32
Defran RH, Weller DW (1999) Occurrence, distribution, site fidelity, and school size of bottlenose dolphins (Tursiops truncatus) off San Diego, California. Mar Mamm Sci 15(2):366–380
Defran RH, Weller DW, KellyDL EMA (1999) Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California bight. Mar Mamm Sci 15:381–393
Díaz-López B (2006) Interactions between Mediterranean bottlenose dolphins (Tursiops truncatus) and gillnets off Sardinia, Italy. ICES J Mar Sci 63:946–951
Díaz-López B, Bernal Shirai JA (2007) Bottlenose dolphin (Tursiops truncatus) presence and incidental capture in a marine fish farm on the north-eastern coast of Sardinia (Italy). J Mar Biol Assoc UK 87:113–117. https://doi.org/10.1017/S0025315407054215
Díaz-López B, Bernal Shirai JA (2008) Marine aquaculture and bottlenose dolphins’ (Tursiopstruncatus) social structure Shirai. Behav Ecol Sociobiol 62:887–894. https://doi.org/10.1007/s00265-007-0512-1
Félix F (1997) Organization and social structure of the coastal bottlenose dolphin Tursiops truncatus in the Gulf de Guayaquil, Ecuador. Aquat Mamm 23(1):1–16
Fruet P, Secchi E, Di Tullio J, Kinas P (2011) Abundance of bottlenose dolphins, Tursiops truncatus (Cetacea: Delphinidae), inhabiting the Patos lagoon estuary, southern Brazil: implications for conservation. Zool 28(1):23–30
Fury CA, Harrison PL (2008) Abundance, site fidelity and range patterns of indo-Pacific bottlenose dolphins (Tursiops aduncus) in two Australian subtropical estuaries. Mar Freshw Res 59(11):1015–1027
Gowans S, Würsig B, Karczmarski L (2008) The social structure and strategies of delphinids: predictions based on an ecological framework. Adv Mar Biol 53:197–267
Griffin RB, Griffin NJ (2004) Temporal variation in Atlantic spotted dolphin (Stenella frontalis) and bottlenose dolphin (Tursiops truncatus) densities on the West Florida continental shelf. Aquat Mamm 30(3):380–390
Hansen LH (1990) California coastal bottlenose dolphins. In: Leatherwood S, Reeves R (eds) The bottlenose dolphin. Academic Press, San Diego, pp 403–420
Hillman GR, Würsig B, Gailey GA, Kehtarnavaz N, Drobyshevsky A, Araabi BN, Tagare HD, Weller DW (2003) Computer-assisted photo-identification of dolphin marine vertebrates: a multi-species system. Aquat Mamm 29:117–123
Hubard CW, Maze-Foley K, Mullin KD, Schroeder WW (2004) Seasonal abundance and site fidelity of bottlenose dolphins (Tursiops truncatus) in Mississippi Sound. Aquat Mamm 30:299–310
Instituto Geográfico Venezolano Simón Bolívar. (2003) Aragua, Ocumare de la Costa 6647. 1:100.000
Irvine AB, Scott MD, Wells RS, Kaufman JH (1981) Movements and activities of the Atlantic bottlenose dolphin, Tursiops truncatus in Sarasota, Florida. Fish Bull (US) 79:671–688
Lauriano G, Mackelworth P, Fortuna CM, Moltedo G, Notarbartolo Di Sciara G (2003) Densitá e abbondanza del Tursiope (Tursiops truncatus) nel Parco nazionaledell’Asinara, Sardegna. Biol Mar Mediterr 10(2):717–720
Leatherwood S, Reeves R (1982) Bottlenose dolphins (Tursiops truncatus) and other toothed cetaceans. In: Chapman J, Feldhamer GA. (eds). Wild mammals of North America. Maryland, p 369–414
Martínez-Serrano I, Serrano A, Heckel G, Schramm Y (2011) Distribution and home range of bottlenose dolphins (Tursiops truncatus) off Veracruz, Mexico. Cienc Mar 37(4A):379–392
Mellink E, Orozco-Meyer A (2006) Abundance, distribution, and residence of bottlenose dolphins (Tursiops truncatus) in the Bahía san Jorge area, northern gulf of California, México. Aquat Mamm 32(2):133–139
Möller LM, Allen SJ, Harcourt RG (2002) Group characteristics, site fidelity and abundance of bottlenose dolphins (Tursiops aduncus) in Jervis Bay and Port Stephens, southeastern Australia. Aust Mamm 24:11–21
Mullin KD, McDonald T, Wells RS, Balmer BC, Speakman T, Sinclair C, Zolman ES, Hornsby F, McBride SM, Wilkinson KA, Schwacke LH (2017) Density, abundance, survival, and ranging patterns of common bottlenose dolphins (Tursiops truncatus) in Mississippi sound following the Deepwater horizon oil spill. PLoS One 12(10):e0186265
Novoa D, Mendoza J, Marcano L, Cárdenas J (1998) In: Mac Sarpa Vencep (ed) El Atlas Pesquero Marítimo de Venezuela. Caracas, p 5
Pitchford JL, Pulis EE, Evans K, Shelley JK, Serafin BJS (2016) Seasonal density estimates of Tursiops truncatus (bottlenose dolphin) in the Mississippi sound from 2011 to 2013. Southeast Nat 15(2):188–206
Quintana-Rizzo E, Wells RS (2001) Resighting and association patterns of bottlenose dolphins (Tursiops truncatus) in the cedar keys, Florida: insights into social organization. Can J Zool 79:447–456
Romero A, Agudo I, Green SM, Notarbartolo di Sciara G. (2001) Cetaceans of Venezuela: distribution and conservation status. NOAA technical report NMFS 151. Rep Fish Bull 36–37
Rossbach KA, Herzing DL (1999) Inshore and offshore bottlenose dolphin (Tursiops truncatus) communities distinguished by association patterns near Grand Bahama Island, Bahamas. Can J Zool 77:581–592
Saayman GS, Tayler CK (1973) Social organization of inshore dolphins (Tursiops truncates and Sousa) in the Indian Ocean. J Mamm 54(4):993–996
Shane SH, Wells RS, Würsig B (1986) Ecology, behavior and social organization of the bottlenose dolphin: a review. Mar Mamm Sci 2(1):34–63
Simões-Lopes PC, Fabian ME (1999) Residence patterns and site fidelity in bottlenose dolphins, Tursiops truncatus (Montagu) (Cetacea, Delphinidae) off southern Brazil. Rev Bras Zootec 16(4):1017–1024
Wells RS (1991) The role of long-term study in understanding the social structure of a bottlenose dolphin community. In: Pryor K, Norris KS (eds) Dolphin societies: discoveries and puzzles. University of California Press, Berkeley, pp 199–225
Wells RS, Hansen, LJ, Baldridge A, Dohl TP, Kelly DL, Defran RH (1990) Northward extension of the range of bottlenose dolphins along the California coast. In: Leatherwood S, Reeves RR (eds) TheBottlenose Dolphin, San Diego, p 421–431
Wells RS, Scott MD (1999) Bottlenose dolphin Tursiops truncatus. In: Ridgeway SH, Harrison RJ (eds) Handbook of marine mammals. Academic Press, San Diego, pp 137–182
Wells RS, Scott MD (2002) Bottlenose dolphin. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of marine mammals. Academic Press, San Diego, pp 122–127
Williams JA, Dawson SM, Slooten E. (1993). The abundance and distribution of bottlenosed dolphin (Tursiops truncatus) in Doubtful Sound, New Zealand. Can J Zool 71(10):2080–2088
Wilson B. (1995) The ecology of bottlenose dolphins in the Moray firth, Scotland: a population at the northern extreme of the species’ range. Ph. D. Thesis. University of Aberdeen. Scotland, p 201
Wilson B, Hammond PS, Thompson PM (1999) Estimating size and assessing trends in a coastal bottlenose dolphin population. Ecol Appl 9:288–300
Würsig B, Würsig M (1977) The photographic determination of group size, composition, and stabilityof coastal porpoises (Tursiops truncatus). Sci 198:755–756
Zhou K, Qian W (1985) Distribution of the dolphins of the genus Tursiops in the China Sea. Aquat Mamm 1:16–19
Zolman ES (2002) Residence patterns of bottlenose dolphins (Tursiops truncatus) in the Stono River estuary, Charleston County, South Carolina, U.S.a. Mar Mamm Sci 18:879–892
Acknowledgments
We are grateful to the fisherman Nelson Barrios for his great help at sea. We are grateful to Mr. William Rossiter and the Cetacean Society International.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Thales Renato Ochotorena de Freitas
Rights and permissions
About this article

Cite this article
Cobarrubia-Russo, S., Barreto, G., Quintero-Torres, E. et al. Occurrence, abundance, range, and residence patterns of Tursiops truncatus on the coast of Aragua, Venezuela. Mamm Res 64, 289–297 (2019). https://doi.org/10.1007/s13364-018-0401-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-018-0401-1