
Abstract
The raccoon dog (Nyctereutes procyonoides) is an East Asian Canid that has been introduced in Europe. Introduction of alien species is an increasing conservation issue. We examined the diet of a recently established raccoon dog population in Denmark by analysing stomach content in 249 carcasses collected in 2008–2016. Raccoon dog diet was compared to the diet of native badger (Meles meles) and red fox (Vulpes vulpes) in Denmark. The most common food for raccoon dogs were invertebrates (frequency of occurrence, FO 69%), small mammals (FO 68%), birds (FO 41%), fruits (FO 38%), amphibians (FO 36%) and carrions (FO 34%). The occurrence of invertebrates was highest during spring and summer, while fruits, cereals and carrions were eaten most often during autumn and winter. As expected, raccoon dog shared the major food categories with badger and red fox, but generally, it had a wider dietary niche. Overall, dietary overlap between raccoon dog and badger was 0.74 (Pianka index, Ojk). The dietary overlap with red fox was relatively high in all seasons, peaking in summer (Ojk 0.87) and dropping in winter (Ojk 0.79). Despite the dietary overlap between the alien racoon dog and native red fox and badger, the species may coexist due to partitioning of feeding habitats and/or because the red fox is limited by other factors, e.g. diseases and anthropogenic activities. The introduced raccoon dog seems to fit a dietary niche between badger and red foxes in human-dominated landscapes in north-western Europe.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Introduction of species outside their natural distribution range may pose a risk to native species and ecosystems, and is regarded as an increasing conservation issue globally (Allendorf and Lundkvist 2003; Simberloff et al. 2013). Alien species may cause population declines of native species through displacement and competition (Maran et al. 1998; Ikeda et al. 2004), predation (Ferreras and Macdonald 1999; Nordström et al. 2003; Salo et al. 2010), behavioural interference and introduction of new diseases (Sainsbury et al. 2000; Holdich et al. 2009), and introgression (Bowman et al. 2017).
The raccoon dog (Nyctereutes procyonoides) is an East-Asian carnivore that was introduced in large numbers in Eastern Europe as a fur game from the late 1920s and is bred in fur farms (Kauhala and Kowalczyk 2011). The species rapidly established a viable population in Europe that has colonised most of Eastern and Northern Europe. Escapees from fur farms may have supplemented the populations and enhanced dispersal and establishment of the species (Nowak 1984; Mulder 2012). Fur farming of raccoon dog was introduced in Denmark in the 1970s. Since 1980, free ranging raccoon dogs have been recorded sporadically (Asferg 1991; Baagøe and Ujvári 2007). These individuals were assumed to be escapees from fur farms and other domestic stocks. In 2008–2009, the number of raccoon dog observations increased substantially in a small area in the western parts of Denmark indicating that a population had established (Sunde and Elmeros 2016). The Danish population was founded by escapees from farms and domestic stocks, but supplementation by animals from the expanding population in Germany occurs (Drygala et al. 2016; Nørgaard et al. 2017).
The raccoon dog has an omnivorous diet that typically includes small mammals, birds, invertebrates, fruits and cereals, amphibians and carrions (Sasaki and Kawabata 1994; Hirasawa et al. 2006; Sutor et al. 2010; Kauhala and Kowalczyk 2011; Koike et al. 2012; Takatsuki et al. 2018). The species’ feeding behaviour is very opportunistic, and the diet varies temporarily and spatially according to the seasonal availability of food items and habitat diversity. In its native distribution range in eastern Asia, the raccoon dog coexists with red fox (Vulpes vulpes), Asian badger (Meles leucurus) and Japanese badger (Meles anakuma) (Wilson and Mittermeier 2009). Compared to the naturally sympatric carnivores in eastern Asia, the raccoon dog fed on invertebrates, fruits and other edible plants more frequently than the red fox which has a more carnivorous diet, while badgers are more reliant on earthworms than sympatric raccoon dog (Yamamoto 1994; Kaneko et al. 2006; Wilson and Mittermeier 2009; Koike et al. 2012).
The opportunistic and adaptive feeding behaviour of raccoon dog has been assumed to be an important factor for the species’ success following the introduction in Europe (Kauhala and Kowalczyk 2011). The colonisation of Europe by raccoon dog has raised concern that it may impact the status of native carnivore as the European badger (Meles meles, hereafter badger) and the red fox negatively through food competition (Skov- og Naturstyrelsen 2010; European Environmental Agency 2012; Kowalczyk 2014). The raccoon dog is also feared to have a negative impact on the population status of prey species, e.g. amphibians and wetland birds as has been documented for American mink (Neovison vison) (Ferreras and Macdonald 1999; Nordström et al. 2003; Salo et al. 2010). Furthermore, the raccoon dog can serve as a reservoir and vector for parasites and diseases, e.g. Echinococcus multilocularis, Sarcoptes scabeii, Trichinella spp. and rabies (Kauhala and Kowalczyk 2011; Al-Sabi et al. 2013; Süld et al. 2014). Raccoon dog has been deemed as invasive in Europe although significant impacts on native species or ecosystems have yet to be documented (Kauhala and Kowalczyk 2011; European Environmental Agency 2012; Mulder 2012; European Commission 2014).
Knowledge on introduced species’ ecology is essential to understand their interactions with native species and effects on ecosystems and to decide on efficient management and conservation actions. The diet of introduced raccoon dog and potential dietary overlap with native generalists carnivores, e.g. badger and red fox, has primarily been studied in wetlands and forested areas in Eastern and Central Europe (e.g. Barbu 1972; Kauhala et al. 1998; Baltrunaite 2002; Sidorovich et al. 2008; Drygala et al. 2013). Landscape composition may influence dietary overlap and potential food competition between species, increasing in managed areas with low habitat variability (Pianka 1974; Kauhala and Ihalainen 2014). The objectives for this study were to (1) analyse the diet of the newly established raccoon dog population and (2) to compare its diet and estimate dietary overlap with native badger and red fox (Laursen 2002; Madsen et al. 2002) intensively human-modified landscapes in Denmark in the temperate reaches of north-western Europe. We assume that the raccoon dog forage on a broad spectrum of foods and that it has a wider dietary niche than the native badger and red fox in Denmark (Laursen 2002; Madsen et al. 2002). Furthermore, we assume higher dietary niche overlap and potential food competition with red fox, particularly during winter as raccoon dog is not expected to be dormant in winter in the temperate climates (Kauhala et al. 1998; Zoller and Drygala 2013).
Materials and methods
Samples
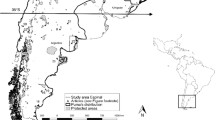
The diet of raccoon dogs was analysed in carcasses collected in the period 2008–2016 in agricultural landscapes and wetlands on the Jutland peninsular, Denmark (55°–57° N, 8°–10° E). The sampling area comprises intensively human-dominated landscapes, consisting of 62% agriculture, 13% urban areas and infrastructures, 12% forest and 11% uncultivated areas (wetlands, heathers, fens, etc.) (Normander et al. 2009). Winter climate is mild with average temperatures between 0 and 1 °C (www.dmi.dk). Raccoon dog carcasses were submitted to the National Veterinary Institute by game managers, hunters and naturalists to monitor the occurrence of zoonotic parasites. The stomach was sampled and stored at − 20 °C. Location, date, gender, season and cause of death were recorded upon reception (Table 1). Seasons were defined as spring, March–May; summer, June–August; autumn, September–November; and winter, December–February. The raccoon dogs were mainly road kills or culled as part of a National Raccoon Dog Action Plan (Skov- og Naturstyrelsen 2010). Individuals that were culled using bait stations and traps were excluded in the diet analysis as the stomach content would represent wildlife managers bait selection, rather than the raccoon dogs food selection. The individuals included in the diet analyses were assessed as adult and subadult based on their weight (> 2 kg).
Diet analysis
The stomach contents of the raccoon dogs were emptied into a 20-cm diameter 1-mm sieve and gently rinsed with running water. The total weight and volume of the content was gauged before rinsing. Different types of food were separated and categorised macroscopically as rodents, insectivores, ungulates, other mammals (e.g. mustelids, lagomorphs and unidentified mammals), birds, amphibians, fish, invertebrates, cereals and fruits. The percentage volume of each category was estimated for each stomach. Food items were stored in 70% ethanol for later identification. Mammalian prey was identified to species, genus or order from the cuticular, cell structure and cross section of guard hairs (Teerink 1991) and teeth if present (Jensen 1993). Feathers were determined to order following Day (1966). Amphibians, reptiles, insects, vegetation and larger undigested body parts of mammals and birds were identified from field guides and reference collections (Jensen 1993; Fog et al. 1997; Toft 2010). Fruits and cereals were classified as food items, while other plant materials, e.g. fragments of grass, leaves and coniferous needles, were classified as non-food (Drygala et al. 2013). Carrions and unidentified matters include skin without hair, feathers or other characteristics, large bones and tissues. Unidentified tissues were characterised as carrions, if maggots were present. Anthropogenic materials (e.g. plastic and rope) and a few raccoon dog hairs occurred in some stomachs (4.4 and 15.3%, respectively). Raccoon dog hair, anthropogenic materials and non-food plants (present in 72.3% of stomachs) were presumed to have been ingested incidentally during grooming or along with the actual food items, and hence not classified as diet.
Statistical analyses
The diet was estimated as frequency (FF: number of stomachs with a food item), frequency of occurrence (FO: percentage of occurrence of food items in non-empty stomachs) and percentage volume (vol%) of different food categories and species in non-empty stomachs. The statistical error of the frequency of occurrence for each food category and species was estimated by binary logistic regression (Proc logistic in SAS 9.4 with logit link function). Means and 95 percentile range confidence intervals of percentage volume of non-empty stomachs were calculated from 10,000 replicate bootstrap estimates (Reynolds and Aebischer 1991; Krebs 1999). Dietary differences between gender and season were tested with χ2 tests of frequency of occurrence and non-parametric Wilcoxon (Wc) and Kruskal-Wallis (K-W) tests of percentage volume of the different food categories.
The dietary niche width was estimated by the Levin’s index (B) and the standardised Levin’s index (Bstd):
where pi is the proportion of each food category i and Bmax is the maximum food categories (Krebs 1999).
Niche overlap between the species was estimated as the Pianka index (Ojk);
where pij og pik is the proportion of food category i in the species j and k, and n is the total number of food categories. Sex and seasonal variances in raccoon dog dietary niche width were compared with Wilcoxon and Kruskal-Wallis tests.
Badger and red fox diets were estimated from undigested food items in the gastro-intestine tract from carcasses collected in rural Denmark, primarily from Jutland (Laursen 2002; Madsen et al. 2002). The food categories ‘rodents’ and insectivores’ were combined as small mammals (rodents and insectivores < 500 g), and ‘fish’ was included in the carrions/unidentified category for comparison of the diet with badger and red fox and estimation of dietary niche widths and niche overlap. Laursen (2002) presented seasonal diet on red foxes enabling seasonal estimates of dietary niche overlap between raccoon dog and red fox. The dietary niche overlap between badger and raccoon dog was estimated for the overall annual food composition, as seasonal data were not presented by Madsen et al. (2002). The overall seasonal distribution of the analysed badgers was similar to the seasonal distribution of analysed raccoon dogs. Dietary differences between raccoon dog, badger and red fox were compared with χ2 tests of frequency of occurrence of the major food categories. Statistical analyses were performed in SAS 9.4 and SAS Enterprise (SAS Institute Inc., Cary, USA).
Results
General food composition
A total of 249 raccoon dog stomachs were examined. Forty-seven of the stomachs were empty. Overall, rodents and carrions/unidentified tissues, birds, fruits, invertebrates and amphibians made up the major food categories, all comprising more than 10% of the volume of the diet (Table 2). Voles (Cricetidae) were the most important small mammalian prey group (FO 32.7%, 12.3 vol%) followed by shrews (FO 14.4%, 3.3 vol%). Passeriformes and Anseriformes had the highest occurrence among the birds (FO 12.4% and FO 9.4%, respectively). One stomach contained feet from a minimum of nine young gulls indicating predation on flightless chickens, and another stomach contained egg shells. This latter individual was killed in January. Frogs and toads made up most of the amphibian prey (FO 35.6%). However, a stomach from one raccoon dog killed in September contained 33 smooth newts (Lissotriton vulgaris). Reptiles were only recorded sporadically (FO 2.0%). Invertebrates were the most commonly recorded food category (FO 69.3%), but it made up a small volume ( 12.8 vol%). Beetles (Coleoptera) and earthworms (Lumbricidae) were the most regularly ingested invertebrates: FO 47.5 and 34.7%, respectively. Fifty-four percent of the stomachs contained fruits or cereals. These were mainly Malus fruits (FO 25.7%), Prunus berries (FO 9.9%), cereal grains (FO 17.8%) and maize (FO 13.9%).
Seasonal variation and dietary niche width
The frequency of occurrence and percentage of volume of some of the raccoon dog major food categories varied seasonally (Fig. 1). The occurrence and percentage volume of invertebrates were highest during spring and summer (χ2 test, FO \( {\chi}_3^2 \) = 15.63, P < 0.01; K-W, vol% \( {\chi}_3^2 \) = 30.39, P < 0.001), while cereals and fruits were most important during summer and autumn (cereals FO \( {\chi}_3^2 \) = 9.28, P < 0.05, vol% \( {\chi}_3^2 \) = 10.68, P < 0.05; fruits FO \( {\chi}_3^2 \) = 17.66, P < 0.001, vol% \( {\chi}_3^2 \) = 19.97, P < 0.001). The occurrence and percentage volume of carrions/unidentified tissues increased during autumn and winter (FO \( {\chi}_3^2 \) = 8.08, P < 0.05; vol% \( {\chi}_3^2 \) = 8.08, P < 0.05), while small rodents were stable food item during the whole year. There was a tendency for wider dietary niche for individual raccoon dog during autumn and winter than during spring and summer (Bstd (mean ± SE), spring 0.21 ± 0.17; summer 0.25 ± 0.18; autumn 0.32 ± 0.19; winter 0.29 ± 0.23, K-W, \( {\chi}_3^2 \) = 6.89, P = 0.075).

Seasonal variations in the percentage frequency of occurrence (a) and volume (b) of food categories for Danish raccoon dogs (mean and 95% confidence intervals). The category ‘Others’ include carrions, fish, and unidentified matters
The seasonal distribution of examined males and females was similar (Table 1, χ2test, \( {\chi}_3^2 \) = 1.93, P = 0.59), and there were no differences in the frequency of occurrence or percentage volume of any of the individual food categories in the males and females. The dietary niche widths of the two genders were similar (Bstd mean ± SE, males 0.26 ± 0.19; females 0.29 ± 0.19, Wc, Z = 0.90, P = 0.37).
Dietary niche overlap with badger and red fox
Raccoon dog shares major food categories with badger (small mammals, invertebrates and cereals) and red fox (small mammals, birds and fruits) (Tables 3 and 4). However, raccoon dogs ate small mammals, birds, amphibians and reptiles, and fruits more frequently than badgers (χ2test FO small mammals \( {\chi}_1^2 \) = 17.40, P < 0.001; birds \( {\chi}_1^2 \) = 28.28, P < 0.001; amphibians and reptiles \( {\chi}_1^2 \) = 7.10, P < 0.01; fruits \( {\chi}_1^2 \) = 20.87, P < 0.001), while badgers ate invertebrates more frequently than raccoon dogs (arthropods and gastropods \( {\chi}_1^2 \) = 38.81, P < 0.001; earthworms \( {\chi}_1^2 \) = 38.81, P < 0.001). Invertebrates also made up a larger volume in badgers than in raccoon dogs (arthropods and gastropods 16.0 vol%; earthworms 29.5 vol%). Raccoon dogs regularly consumed amphibians and reptiles and cereals, which was not recorded in red foxes. Raccoon dogs fed on invertebrates more frequently than red fox in all seasons (spring \( {\chi}_1^2 \) = 9.65, P < 0.01; summer \( {\chi}_1^2 \) = 12.93, P < 0.001; autumn \( {\chi}_1^2 \) = 17.93, P < 0.001; winter \( {\chi}_1^2 \) = 37.91, P < 0.001). During autumn, raccoon dogs also fed on birds and fruits more often than red fox (birds \( {\chi}_1^2 \) = 5.76, P < 0.05; fruits \( {\chi}_1^2 \) = 5.06, P < 0.05).
Overall, raccoon dogs had a wider annual dietary niche (Bstd 0.91) than badger (Bstd 0.52) and a wider dietary niche width than the red fox in all seasons (Table 4). The dietary niche overlap between raccoon dog and badger was 0.74. Seasonal dietary overlap between red fox and raccoon dog varied from 0.79 to 0.89, peaking during summer.
Discussion
Raccoon dogs had a broad omnivorous diet comprising mainly small rodents, birds, amphibians, carrions and fruits in the intensively human-modified landscape in Denmark. The recorded dietary niche and the major food categories of the species parallel the results from dietary studies in other European countries and in the species native range (e.g. Barbu 1972; Sasaki and Kawabata 1994; Kauhala et al. 1998; Hirasawa et al. 2006; Sutor et al. 2010; Koike et al. 2012; Drygala et al. 2013; Süld et al. 2014; Takatsuki et al. 2018). Small rodents and shrews were stable food resources for raccoon dogs during most seasons in the present study. Raccoon dogs predate on other prey groups, e.g. birds and amphibians, when small mammals are scarce and the alternative food items are easily available (Sutor et al. 2010; Kauhala and Ihalainen 2014). Ungulate carrions were primarily an important food source in autumn and winter. Raccoon dogs are probably not capable of killing cervids but may scavenge on carcasses. Scavenging on ungulate killed by large carnivores is an important resource for raccoon dogs during winter in regions with harsh winter climates (Jędrzejewska and Jędrzejewski 1998; Sutor et al. 2010). In Denmark, large predators are essentially absent, but game hunting is widespread during autumn and winter (Asferg et al. 2016), and offal from the game hunts are available for raccoon dogs. Furthermore, raccoon dogs may feed on ungulate carcasses from road kills, which are easily available throughout the year, but peaks during the late autumn (Elmeros et al. 2014).
Birds were important food category for raccoon dogs in Denmark. Predation by raccoon dog is often promoted as a major threat to ground nesting waterfowls (Skov- og Naturstyrelsen 2010; European Environmental Agency 2012). However, we only found evidence of predation on wild breeding birds and nestlings in one stomach, while the observation of eggshells in a raccoon dog from January indicated predation on domestic birds. Less than 1% waterfowl nest predation in a wetland area of Latvia was caused by raccoon dogs (Opermanis et al. 2001). Water fowl generally has a low occurrence in raccoon dog diet (0–5%), but higher predation rates by raccoon dog have been recorded on islands (Sutor et al. 2010; Kauhala and Kowalczyk 2011). As observed in other studies from Central Europe, the major group of birds in the diet of raccoon dogs was passerines (Sutor et al. 2010; Drygala et al. 2013).
Amphibians can be an important prey category for raccoon dogs during spring and summer in some landscapes (Barbu 1972; Kauhala et al. 1998; Baltrunaite 2002; Sidorovich et al. 2008; Sutor et al. 2010). The high number of smooth newts in one stomach in the present study suggests predation on amphibians at hibernation sites as described for other medium-sized predators, e.g. otter (Jędrzejewska and Jędrzejewski 1998). Road-killed amphibians may represent an abundant and easily accessible food source for amphibians, especially during the amphibian breeding migration period. Kauhala et al. (1998) suggested that many reptiles are primarily eaten as carcasses from road kills, since the reptiles typically are inactive during night when raccoon dogs are foraging. Predation on reptiles by raccoon dog was more frequent in Finland and Belarus than recorded in the present study (Kauhala et al. 1998; Sidorovich et al. 2000; Kauhala and Ihalainen 2014). Thus, a low occurrence of reptiles in the raccoon dog diet in the heavily modified landscapes with dense road networks in Denmark may suggest a general unfavourable status of reptile populations.
Raccoon dog often consumed fruits and cereals, especially during autumn and winter. Fruits are important food items throughout the introduced range and in species native Asian range (Sasaki and Kawabata 1994; Sutor et al. 2010; Takatsuki et al. 2018). Cereals are also recorded in most raccoon dog dietary studies, but the present study and studies in Germany found a relatively high occurrence in all seasons than other dietary studies from Europe (Sutor et al. 2010; Drygala et al. 2013; Süld et al. 2014). In Denmark, cereals are widely available for the fauna during most of the year at game feeding stations, which evidently also benefits the raccoon dog.
The raccoon dog had a more diverse diet compared to badger and red fox in our study area as expected. The major food categories, dietary differences and niche width parallels result from studies in Central and Northern Europe, where raccoon dog populations have thrived for decades (Kauhala et al. 1998; Baltrunaite 2002; Sidorovich et al. 2008; Drygala et al. 2013; Süld et al. 2014). The seasonal dietary overlap was highest during summer and lowest during winter, which mirrors observations on the species’ diet and niche overlap in eastern Germany (Drygala et al. 2013). This contrast findings in Eastern and Northern Europe where the seasonal dietary niche overlaps between red fox and raccoon dog were low in summer but peaked in winter, when both species may rely heavily on carrions (Sidorovich et al. 2000; Baltrunaite 2002).
Species with similar ecological requirements may coexist because niche partitioning according to diet, temporal or spatial habitat use may be allowed and/or because the populations are limited by other factors than food resources (Wiens 1993; Remonti et al. 2012; Lovari et al. 2013). Despite the diet overlap between feral raccoon dog and native red fox and badger, there is little evidence for competitive interaction among the species (Kauhala et al. 1998; Drygala et al. 2013). Declines in all native predator populations were reported to coincide with an increase in the raccoon dog population in Belarus (Sidorovich et al. 2000), but no significant negative impacts on the populations of red fox and badger have been detected in e.g. Finland, Poland and Germany, where the raccoon dog populations are well established (Kowalczyk et al. 2008; Kauhala and Kowalczyk 2011; Drygala et al. 2013; www.jagdverband.de, http://stat.luke.fi). Potential resource competition is reduced by habitat partitioning between raccoon dog, red fox and badger and by winter dormancy by badger and raccoon dog in regions with severe winter climate (Kauhala and Auttila 2010, Zoller and Drygala 2013). Furthermore, the populations of the native species might be limited by other factors than food resources in heavily human-dominated landscapes, e.g. diseases and human activities such as road kills, hunting and culling (Asferg et al. 2016). Indeed, red fox and the badger species coexist with racoon dog across Eastern Europe and in its natural range in East Asia. In conclusion, the alien raccoon dog may fit a trophic niche between badger and red fox in north-western Europe.
References
Al-Sabi MNS, Chriél M, Jensen TH, Enemark HL (2013) Endoparasites of the raccoon dog (Nyctereutes procyonoides) and the red fox (Vulpes vulpes) in Denmark (2009-2013) A comparative study. Int J Parasitol 2:144–251
Allendorf FW, Lundkvist LL (2003) Introduction: population biology, evolution, and control of invasive species. Conserv Biol 17:24–30
Asferg T (1991) Rovdyr. In: Muus B (ed) Danmarks pattedyr 2. Gyldendal, København, pp 7–103
Asferg T, Clausen P, Christensen TK, Bregnballe T, Clausen KK, Elmeros M, Fox AD, Haugaard L, Holm TE, Laursen K, Madsen AB, Madsen J, Nielsen RD, Sunde P, Therkildsen OR (2016) Vildtbestande og jagttider i Danmark. Det biologiske grundlag for jagttidsrevisionen 2018. Aarhus University, Scientific report no. 195, Aarhus
Baltrunaite L (2002) Diet composition of the red fox (Vulpes vulpes), pine marten (Martes martes) and raccoon dog (Nyctereutes procyonoides Gray) in clay plain landscape, Lithuania. Acta Zool Lituan 12:1392–1657
Barbu P (1972) Beiträge zum Studium des Marderhundes, Nyctereutes procyonoides ussuriensis aus dem Donaudelta. Säugetierkd Mitt 20:375–405
Baagøe HJ, Ujvári M (2007) Mårhund Nyctereutes procyonoides (Gray, 1834). In: Baagøe HJ, Jensen TS (eds) Dansk Pattedyr Atlas. Gyldendal, København, pp 182–183
Bowman J, Beauclerc K, Farid AH, Fenton H, Klütsch CFC, Schulte-Hostedde AI (2017) Hybridization of domestic mink with wild American mink (Neovison vison) in eastern Canada. Can J Zool 9:443–451
Day MG (1966) Identification of hair and feather remains in the gut and faeces of stoats and weasels. J Zool, Lond 148:201–217
Drygala F, Werner U, Zoller H (2013) Diet composition of the invasive raccoon dog (Nyctereutes procynoides) and the native red fox (Vulpes vulpes) in north-east Germany. Hysterix 24:190–194
Drygala F, Korablev N, Ansorge H, Fickel J, Isomursu M, Elmeros M, Kowalczyk R, Baltrunaite L, Balciauskas L, Saarma U, Schulze C, Borkenhagen P, Frantz AC (2016) Homogenous population genetic structure of the non-native raccoon dog (Nyctereutes procyonoides) in Europe as a result of rapid population expansion. PLoS ONE 11:e0153098. https://doi.org/10.1371/journal.pone.0153098
Elmeros M, Andersen PN, Sunde P, Haugaard L, Skov F, Madsen AB (2014) Påkørte større vilde dyr i Danmark (2003-2012). Aarhus University. Scientific report no. 91. In: Aarhus
European Commission (2014) EU Regulation 1143/2014 on Invasive Alien Species. http://ec.europa.eu/environment/nature/invasivealien/index_en.htm. Assessed 10/09/2017
European Environmental Agency (2012) The impacts of invasive alien species in Europe. EEA Technical report no. 16/2012
Ferreras P, Macdonald DW (1999) The impact of American mink Mustela vison on water birds in the upper Thames. J Appl Ecol 36:701–708
Fog K, Schmedes A, de Lasson DR (1997) Nordens pattedyr og krybdyr. GAD, København
Hirasawa M, Kanda E, Takatsuki S (2006) Seasonal food habits of the raccoon dog at a western suburb of Tokyo. Mamm Study 31:9–14
Holdich DM, Reynolds JD, Souty-Grosset C, Sibley PJ (2009) A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl Manag Aquat Ecosyst 349–395. https://doi.org/10.1051/kmaa/2009025
Ikeda T, Asano M, Matoba Y, Abe G (2004) Present status of invasive alien raccoon and its impact in Japan. Global. Environ Res 8:125–131
Jędrzejewska B, Jędrzejewski W (1998) Predation in vertebrate communities. The Białowieźa Primeval Forest as a Case Study. Springer Verlag, Berlin
Jensen TS (1993) Mus, rotter og spidsmus. Natur og Museum, Naturhistorisk Museum, Århus
Kaneko Y, Maruyama N, Macdonald DW (2006) Food habits and habitat selection of suburban badgers (Meles meles) in Japan. J Zool 270:78–89
Kauhala K, Auttila M (2010) Habitat preferences of the native badger and the invasive raccoon dog in southern Finland. Acta Theriol 55:231–240
Kauhala K, Ihalainen A (2014) Impact of landscape and habitat diversity on the diversity of diets of two omnivorous carnivores. Acta Theriol 59:1–12
Kauhala K, Kowalczyk R (2011) Invasion of the raccoon dog Nyctereutes procynoides in Europe: History of colonization, features behind its success, and threats to native fauna. Curr Zool 5:584–598
Kauhala K, Laukkanen P, Rége I (1998) Sommer food composition and food niche overlap of the raccoon dog, red fox and badger. Ecography 21:457–463
Koike S, Morimoto H, Goto Y, Kozakai Y (2012) Insectivory by five sympatric carnivores in cool-temperate deciduous forest. Mamm Study 37:73–83
Kowalczyk R (2014) NOBANIS invasive alien species fact sheet Nyctereutes procyonoides. European Network on Invasive Alien Species, http://nobanis.org. Accessed 27/09/2017
Kowalczyk R, Jedrzejewska B, Zalewski A, Jedrzejewski W (2008) Facilitative interactions between the Eurasian badger Meles meles, the red fox Vulpes vulpes and the invasive raccoon dog Nyctereutes procyonoides in Białowieza Primeval Forest, Poland. Can J Zool 86:1389–1396
Krebs CJ (1999) Ecological methodology. Addison Welsey Longman, New York
Laursen LS (2002) Identifikation, kvantificering og sammenligning af føden hos landræve og byræve i Danmark. Unpublished M.Sc. thesis from Copenhagen University and the National Environmental Research Institute, Denmark
Lovari S, Minder I, Ferretti F, Mucci N, Randi E, Pellizzi E (2013) Common and snow leopards share prey, but not habitats: competition avoidance by large predators? J Zool 291:127–135
Madsen SA, Madsen AB, Elmeros M (2002) Seasonal food of badgers (Meles meles) in Denmark. Mammalia 3:341–352
Maran T, Kruuk H, Macdonald DW, Polma M (1998) Diet of two species of mink in Estonia: displacement of Mustela lutreola by Mustela vison. J Zool, Lon 245:218–222
Mulder JL (2012) A review of the ecology of the raccoon dog (Nyctereutes procyonoides) in Europe. Lutra 55:101–127
Nordström M, Högmander J, Laine J, Nummelin J, Laanetu N, Korpimäki E (2003) Effects of feral mink removal on seabirds, waders and passerines on small islands of the Baltic Sea. Biol Conserv 109:359–368
Normander B, Jensen TS, Henrichs T, Sanderson H, Pedersen AB (eds) (2009) Natur og Miljø 2009 – Del A: Danmarks miljø under globale udfordringer. Aarhus University, National Environmental Research Institute
Nowak E (1984) Verbreitungs- und Bestandsentwicklung des Mardelhundes Nyctereutes procyonoides (Gray 1834) in Europa. Z Jagdwiss 301:37–154
Nørgaard LS, Mikkelsen DMG, Elmeros M, Chriél M, Madsen AB, Nielsen JL, Pertoldi C, Randi E, Fickel J, Brygida S, Ruiz-Gonzáles A (2017) Population genomics of the raccoon dog (Nyctereutes procyonoides) in Denmark: insights into invasion history and population development. Biolo Invasions 19:1637–1652. https://doi.org/10.1007/s10530-017-1385-5
Opermanis O, Mednis A, Bauga I (2001) Duck nests and predators: interaction, specialisation and possible management. Wildl Biol 7:87–96
Pianka ER (1974) Niche Overlap and Diffuse Competition. Proc Nat Acad Sci 71:2141–2145
Remonti L, Balestrieri A, Ruiz-González A, Gómez-Moliner BJ, Capelli E, Prigioni C (2012) Intraguild dietary overlap and its possible relationship to the coexistence of mesocarnivores in intensive agricultural habitats. Popul Ecol 54:521–532
Reynolds JC, Aebischer NJ (1991) Comparison and quantification of carnivore diet by faecal analysis: a critique, with recommendations, based on a study of the Fox Vulpes vulpes. Mamm Rev 3:97–122
Sainsbury AW, Nettleton P, Gilray J, Gurnell J (2000) Grey squirrels have high seroprevalence to a parapoxvirus associated with deaths in red squirrels. Anim Conserv 3:229–233
Salo P, Ahola MP, Korpimäki E (2010) Habitat-mediated impact of alien mink predation on common frog densities in the outer archipelago of the Baltic Sea. Oecol 163:405–413
Sasaki H, Kawabata M (1994) Food habits of the raccoon dog Nyctereutes procyonoides viverrinus in a mountainous area in Japan. J Mamm Soc Japan 19:1–8
Sidorovich AG, Polozow AG, Lauzhel GO, Krasko DA (2000) Dietary overlap among generalist carnivores in relation to the impact of the introduced raccoon dog Nyctereutes procyonoides on native predators in northern Belarus. Z Säugetierkd 65:271–285
Sidorovich VE, Solovej IA, Sidorovich AA, Dyman AA (2008) Seasonal and annual variation in the diet of the raccoon dog Nyctereutes procynoides in northern Belarus: the role of habitat type and family group. Acta Theriol. 53:27–38
Simberloff D, Martin JL, Genovesi P, Maris V, Wardle DA, Aronson J, Courchamp F, Galli B, García-Berthou E, Parcal M, Pyšek P, Sousa R, Tabacchi E, Vilà M (2013) Impacts of biological invasions: what’s what and the way forward. Trends Ecol Evol 28:58–66
Skov- og Naturstyrelsen (2010) Indsatsplan mod mårhund. Skov- og Naturstyrelsen, København
Sunde P, Elmeros M (2016) Bestandsudvikling af mårhund i Danmark 2009-2016 i relation til nutidig og fremtidig bekæmpelsesindsats. Consultancy note to National Nature Agency. Aarhus University, Aarhus
Sutor A, Kauhala K, Ansorge H (2010) Diet of the raccoon dog Nyctereutes procynoides—a canid with an opportunistic foraging strategy. Acta Theriol 55:65–176
Süld K, Valdmann H, Laurimaa L, Soe E, Davison J, Saarma U (2014) An invasive vector of zoonotic disease sustained by anthropogenic resources: the raccoon dog in northern Europe. PLoS ONE 9:e96358. https://doi.org/10.1371/journal.pone.0096358
Takatsuki S, Miyaoka R, Sugaya K (2018) A comparison of food habits between Japanese marten and raccoon dog in Western Tokyo with reference to fruit use. Zool Sci 35:68–74
Teerink B (1991) Atlas and identification key—hair of West European mammals. Cambridge University Press, Cambridge
Toft S (2010) Leddyrenes biologi. Aarhus University, Biological Institute
Wiens JA (1993) Fat times, lean times and competition among predators. TREE 8:348–349
Wilson DE, Mittermeier RA (eds) (2009) Handbook of the mammals of the world, vol 1 Carnivores. Lynx Edicions, Barcelona
Yamamoto Y (1994) Comparative analyses on food habits of Japanese marten, red fox, badger and raccoon dog in the Mt. Nyugasa, Nagano Prefecture, Japan. Nat. Environ Sci Res 7:45–52
Zoller H, Drygala F (2013) Spatial use and interaction of the invasive raccoon dog and the native red fox in Central Europe: competition or coexistence? Eu J Wildl Res 59:683–691
Acknowledgements
We thank the game consultants on the Nature Agency, hunters and other naturalists for collecting and submitting raccoon dog carcasses for analysis. Thanks to the Aalborg Zoo Conservation Foundation (AZCF) and Jægernes Naturfond for providing financial support for the project, and Søren Østergaard Gertsen for identifying some of the insect species. We are grateful to two anonymous referees for their comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Rafał Kowalczyk
Rights and permissions
About this article

Cite this article
Elmeros, M., Mikkelsen, D.M.G., Nørgaard, L.S. et al. The diet of feral raccoon dog (Nyctereutes procyonoides) and native badger (Meles meles) and red fox (Vulpes vulpes) in Denmark. Mamm Res 63, 405–413 (2018). https://doi.org/10.1007/s13364-018-0372-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-018-0372-2