
Abstract
Climatic warming and shifts in precipitation patterns could reduce the growth and reproduction of North American bison (Bison bison). To quantify rates of body mass changes for different classes of bison grazing on native tallgrass prairie, a remote autonomous weighing system was positioned to passively record weights throughout the growing season of radio frequency identification (RFID)-tagged yearling, 2-year-old, and adult female bison. Over a 152-day period, yearling bison gained weight longer into the year than adult females, but adult females had higher intrinsic growth rate. The midsummer asymptote of weight for adult females could not be ascribed entirely to the demands of lactation. Adult females that did not have a calf were heavier than those that did, but individuals of both groups leveled off in body mass at similar times in midsummer. We also assessed dietary quality of adult females approximately every 10 days from fecal samples. Adult female weight gain decreased with decreasing dietary crude protein ([CP]) concentrations suggesting that adult female bison cease gaining weight when [CP] = 54.5 mg g−1 and gained weight less efficiently at moderate dietary quality than expected. Broader adoption of this technique will begin to generate the data necessary to understand the effects of variation in weather and climate on seasonal patterns of large herbivore body mass.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The world’s grasslands are grazed by over 1 billion cattle and over 2 billion sheep, goats, and water buffalo (Ripple et al. 2013). In addition, millions of other non-domesticated ungulates graze grasslands throughout the world. Included in these are approximately 500,000 North American bison (Bison bison), which are distributed in conservation and commercial herds (Gates et al. 2010). Despite their centrality to the ecology and economics of grasslands, we have little ability to understand how short-term and long-term changes in climate are likely to affect bison, no less other grazers. Climatic warming and shifts in precipitation patterns are likely to affect the quantity and quality of forage for bison, but it is unclear how these changes will affect bison weight gain and reproduction.
Recent research that examined interannual and geographic patterns of bison performance is beginning to shed light on how changes in climate are likely to cascade through soils and plants to affect bison performance. In the tallgrass prairie region, bison weight gain and calving is enhanced in years with wet late summers (Craine et al. 2013). In contrast, bison performance is reduced in years with wet early summers, most likely due to reduced nutritional forage quality since grass production is enhanced by precipitation during this time (Craine et al. 2013). The effects of increased early summer precipitation on bison performance appear to be reversed in the more xeric shortgrass prairies of North America (Craine et al. 2010). In tallgrass prairie, hotter weather has little effect on bison behavior (Allred et al. 2013) and does not significantly affect bison performance (Craine et al. 2013). Yet, bison weight gain is lower in grasslands that have hotter climates than those that are cooler (Craine 2013). For example, a typical 3.5-year-old male bison is 13 kg lighter for every 1 ° C increase in mean annual temperature (Craine 2013). In North America, grasslands with hotter temperatures typically have lower forage nutritional quality than grasslands with cooler temperatures (Craine et al. 2010; Tieszen et al. 1998), which would underpin the lower weight gain in hotter regions.
As bison weight gain is sensitive to variation in temperature and precipitation over short- and long-time scales, climatic warming or shifts in the timing of precipitation could reduce the performance of bison. Yet, as these patterns are based on annual weight gain, we have little ability to understand the proximal mechanisms that generate variation in annual performance. If we could understand how weight gain trajectories differ among years and along climate gradients, and whether they are driven by changes in dietary quality, the impact of climate change on bison will be better understood.
Given the importance of quantifying seasonal changes in body mass, the next step to understanding how these differences in weather and climate affect bison performance is to quantify intra-annual patterns of weight gain. Of relevance for bison in more humid grasslands of the Kansas Flint Hills, on which we focus here, weight gain for cattle grazing on native grassland in the Flint Hills of Kansas is highest in spring and then declines as the summer progresses (Anderson et al. 1970). The seasonal pattern of weight gain parallels declining nutritional quality of grass in the Flint Hills as well as other grassland regions (Craine et al. 2010; Rao et al. 1973). Bison in the Flint Hills appear to have similar dietary quality patterns as cattle (Craine et al. 2013), which suggests that bison may only have approximately 100 days to increase their body mass. Yet, it is unknown whether different classes of animals follow similar weight-gain trajectories or whether dietary quality patterns actually correspond to these trajectories.
In order to determine the intra-annual patterns of weight gain in bison and whether it can be predicted by variation in dietary quality, we examined the intra-annual variation in body mass for adult female, 2-year-old, and yearling bison in the Flint Hills tallgrass prairie region of North America. Because the typical handling required for determining bison weights is labor-intensive and stressful on the animals, we utilized a remote autonomous weighing (RAW) system that passively weighs and identifies individuals as they walk between grazing units. Among our hypotheses, if the nutritional quality of plants is the predominant factor influencing patterns of weight gain then short-term weight gain should correlate strongly with dietary quality. This implies that the rate at which bison gain weight should peak in early spring and then gradually decline as the summer progresses. Likewise, if the nutritional quality of plants is the driving factor in the patterns of weight gain, adult females and juvenile bison should show similar patterns of weight gain over the course of the season. Yet, the seasonal demands of lactation and mating could alter the patterns between the classes. For example, since adult female bison that have not had a calf gain more weight at the annual scale than those that did have a calf (Hamel et al. 2010), it is unknown whether adult females that have not calved gain weight over a longer period during the growing season or at a higher rate during the same period.
Methods
Study site
The bison for this study were located at Konza Prairie Biological Station (39° 05′ N and 96° 35′ W), a 34.9-km2 tallgrass prairie preserve in the northern Flint Hills ecoregion in Kansas, USA. Bison have access to 9.6 km2 spread across ten watersheds (Towne 1999). Within the bison unit, two watersheds have been burned in the spring since 1988 at each of three fire frequencies (1, 2, and 20 years), while four watersheds have been alternately burned on a 4-year interval. Vegetation at Konza Prairie is dominated by warm-season perennial grasses, including Andropogon gerardii, Sorghastrum nutans, and Schizachyrium scoparium, but many eudicots are present in grazed areas. Animals graze on natural vegetation available to them and were not supplemented with either protein or energy during the study period.
Herd size is maintained with an annual fall culling program. Bison are collected into a corral in late October, individually weighed on an electronic chute scale, and calves are given a uniquely numbered visual ear tag. In 2013, 90 calves, 9 yearlings, and 21 adult females were also fitted with half-duplex radio frequency identification (RFID) ear tags. Herd size after calving in 2014 was approximately 390 animals. Parturient status of all adult females in 2014 was monitored visually throughout the year, including approximate date that calves were born.
Field methods
A remote autonomous weighing (RAW) system was positioned across a gate that separated the bison unit into two subunits. This was one of five open gates where the bison can freely travel between the two units. The RAW system consisted of components from Tru-Test, Inc. (Mineral Wells, TX, USA): two load bars (MP800), an aluminum platform (86 × 222 cm; AP800), a large EID antenna, an RFID reader (XRP2), and a Walk Over Weighing indicator (WOW-3000). The indicator determines weights when animals pass over the platform. The system was powered by two marine deep-cycle batteries that were charged by a 100-W solar panel. The alley encompassing the platform was approximately 1 m × 3 m with wooden sidewalls and posts.
The scale was installed on 28), but the RFID reader was not installed until day of year (DOY) 118 (Apr 28). The last weight recorded for this study was on DOY 269 after which animals were isolated from the scale in preparation for the annual round-up. Once weight data collection began, the 90 tagged bison calves were yearlings, and the former yearlings were 2 years old. Data were downloaded weekly.
To examine concentrations of dietary crude protein ([CP]) and digestible organic matter ([DOM]), fresh fecal material was collected from an average of 5 adult females and 5 adult males approximately every 10 days over the period of study. For each sampling period, individual fecal samples were pooled by sex. We used near-infrared spectroscopy of dried fecal material to periodically estimate [CP] and [DOM] based on calibrations generated from cattle (Craine et al. 2013). The ratio of forage [DOM] to [CP] is a crude index of ruminal fermentability (Moore et al. 1999), and bison were generally protein limited when weight gain was low. Hence, we examine patterns of [CP] exclusively here.
Data analysis
The RAW system process generates a large number of inaccurate readings. For example, the weights of animals that move too fast across the scale are substantially lower than expected. When multiple animals are present on the scale at the same time, weights will be substantially heavier than expected. In order to remove outlier readings, all weights less than 100 kg were initially excluded. Then, all weights greater than 400, 500, and 600 kg for yearlings, 2-year olds, and cows, respectively, were excluded. These values are far above typical weights for these classes of animals and would indicate an erroneous reading for the individual. For each animal, a spline was fit to the remaining data and residuals calculated. If the coefficient of variation (CV) of the residuals of the remaining data was greater than 6 %, the weight with the greatest residual was removed. This process was iterated until the CV fell below 6 %. Forty-one individuals that did not have sufficient weights distributed over a wide enough period to ensure a valid weight trajectory were excluded from the analyses. The remaining individuals included in this study were 56 yearlings (26 female, 30 male), 8 2-year-old bison (4 female, 4 male), and 15 adult females. After outliers were removed, there was an average of 11 weights per animal throughout the season.
In order to examine the general patterns of weight changes, a linear model was used to standardize weights of each animal in a given class. This approach facilitates determining and visually displaying weight gain trajectories for each class of animal. By modeling animal weight with DOY, animal ID as a categorical factor, and an interaction between the class of the animal and DOY, 97 % of the variation in weights was explained. With results from the model, weights of animals were adjusted by adding a constant to the weights of each animal so that all individuals of a given class had a common mean. Constants were derived from parameter estimates of linear model. This common mean was set to the mean weight of all the animals for the class across all measurements. Parallel procedures were used for male and female yearlings (r 2 = 0.94) as well as adult females that did or did not have a calf in 2014 (r 2 = 0.87).
Using the adjusted weight data, a three-parameter logistic was fit to each class of animal:
where a is the intrinsic growth rate (kg kg−1 day−1), b is the day of year for the inflection point, and c is the asymptotic weight (kg).
The logistic fit was performed for all yearlings, yearlings of each sex, all 2-year-old animals, 2-year olds of each sex, all adult females, and adult females that either had or did not have a calf that year. No 2-year-old females had a calf. A four-parameter logistic is more reasonable over an annual scale since it includes a nonzero lower asymptote, but the timing of when we started weighing animals was typically near or after the inflection point. A four-parameter logistic provided little additional explanatory power, and the parameters became too unconstrained with current data. After fitting a three-parameter logistic to each age class, all individual weights that had a residual whose absolute value was >40 kg were excluded from the dataset. This process was iterated until no additional residuals were removed.
Asymptotic weight and maximum intrinsic weight gain from the logistic curves were compared between categories with equivalence tests on ratios. We also calculated the fraction of asymptotic weight the animals had reached at the end of the study (DOY 269) and the day of year at which animals were at 98 % of asymptotic weight. Given that calculated inflection points were often before the time when weights were measured, weight gain rates was calculated between DOY 120–140, which would be the highest growth rates within study dates.
Only one adult male was fitted with an RFID tag in 2013, the data from which were insufficient to analyze here. Yet, the weight of the largest males at any one point in time could be assessed by examining the maximum weights recorded over a period. To examine the weight trajectories of the largest males, we also examined the upper bound of the weight data envelope over time. To do this, the maximum recorded weights of all individuals were determined every 10 days beginning with DOY 90. A spline (λ = 1000) was then fit to these data, and the general patterns of weight changes over time are described.
All statistics were computed in JMP 11.2 except for the initial outlier analysis, which was conducted in R 3.1.1.
Results
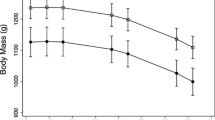
The weight trajectory of yearlings reveals that their predicted asymptotic weights in 2014 were 293.9 ± 4.3 kg with maximum intrinsic growth rates estimated to be 0.015 ± 0.001 day−1 (Fig. 1a). The maximum observed growth rate for the yearlings between DOY 120 and DOY 140 was 1.0 kg day−1. On the last day of measurement (DOY 269, September 26), yearlings weighed 93.3 % of the asymptotic weight as determined by the logistic model. Extrapolating this pattern with the logistic relationship, yearlings would be predicted to reach 98 % of their asymptotic weight on DOY 353 (December 19).

Weight trajectories in 2014 for a 1-year-old bison (yearlings), b 2-year-old bison, and c adult females. The best-fit three-parameter logistic is included for each panel. Horizontal lines represent asymptotic weight. The gray box is the period used to determine maximum weight gain rates (DOY 120–140)
Male and female yearlings did not significantly differ (P > 0.05) in intrinsic growth rates or asymptotic weight (Fig. 2). Yet, male yearlings tended to have a higher asymptotic weight than female yearlings (304.5 ± 5.1 vs. 290.9 ± 6.9 kg; P < 0.1). Between DOY 120 and DOY 140, weight gain was 1.05 kg day−1 for males and 0.95 kg day−1 for females.

Comparison of weight trajectories for male (black line, closed circles) and female (gray line, open circles) yearlings. Best-fit three-parameter logistic is included for each sex class. Horizontal lines represent asymptotic weight (dashed = males, dotted = females). The gray box is the period used to determine maximum weight gain rates (DOY 120–140)
Compared to yearlings, 2-year-old bison had higher growth rates and reached their asymptotic weights earlier (Fig. 1b). The 2-year-old bison had an asymptotic weight of 401.2 ± 4.0 kg and a maximum intrinsic growth rate of 0.021 ± 0.002 day−1, which were both significantly greater than those of yearlings (P < 0.001 for both). Male 2-year-old bison tended to have a higher asymptotic weight than female 2-year-old bison, but it was not significant with the relatively small sample size (417.4 ± 3.7 vs. 398.0 ± 10.7 kg, P < 0.15). For all 2-year-old bison, the maximum observed growth rate was 1.6 kg day−1 between DOY 120 and DOY 140. On DOY 269, their weight was 98.1 % of their asymptotic weight. Two-year old bison reached 98 % of their asymptotic weight on DOY 267 (September 24).
Asymptotic weight for adult females was 452.3 ± 1.7 kg, and their maximum intrinsic growth rate was 0.031 ± 0.004 day−1 (Fig. 1c), which were both significantly greater than yearlings and 2-year-old bison (P < 0.001 for all). Maximum observed growth rates were 1.48 kg day−1. On DOY 269, their weight was 99.8 % of their asymptotic weight. Adult females reached 98 % of their asymptotic weight on DOY 189 (July 8).
The early asymptote of weight for adult females could not be ascribed entirely to the demands of lactation. Adult females that did not have a calf were heavier than those that did not, but both groups of females leveled off in weight at similar times (Fig. 3). Adult females that had a calf had a higher asymptotic weight than those without a calf (467.0 ± 2.3 vs. 437.5 ± 2.3 kg, P < 0.01). Maximum intrinsic growth rates were not significantly higher for adult females without calves (P > 0.1), but maximum weight gain was 65 % higher between DOY 120 and 140 (2.02 vs. 1.23 kg day−1). Adult females with calves reached 98 % of their asymptotic weight 16 days before those without calves (DOY 173 vs. 189).

Comparison of weight trajectories for adult females that produced a calf (gray line, open circles) and those that did not have a calf (black line, closed circles) in 2014. Best-fit three-parameter logistic is included for each class. Horizontal lines represent asymptotic weight (dashed = no calf, dotted = with calf). The gray box is the period used to determine maximum weight gain rates (DOY 120–140)
To infer weight dynamics of the largest males, we examined the upper bounds of the data envelope between DOY 95 and DOY 185 (April 5–July 4) (Fig. 4). During this period, the largest males gained 194.3 kg (726.7 vs. 919.5 kg), which was equivalent to 2.2 kg day−1 weight gain. From DOY 185 to DOY 235 (July 4–August 23), the largest males lost 106 kg (920.9 vs. 804.7 kg). This rate of weight loss was 2.3 kg day−1. The highest weight observed from DOY 260–270 was 836 kg, which suggests that the largest males may have begun to regain mass at a rate of 1.0 kg day−1 after DOY 235.

Recorded weights for all animals in 2014. The heaviest weights in each 10-day increment are shown with black circles. Upper bound of data envelope represents the weight trajectory of the largest males. The curve was generated by determining the heaviest weight recorded for successive 10-day periods and then a spline fit through the data
In 2014, dietary quality for females increased during the spring, peaked on DOY 129 (May 9) at 129.5 mg g−1 and then declined monotonically to a low of 58.8 mg g−1 on DOY 258 (September 15) (Fig. 5). These patterns were largely similar for adult males and females (Fig. 5). For adult females, across all dates where dietary quality was measured, weight gain decreased with decreasing [CP] (n = 11, r 2 = 0.96, P < 0.001 for linear relationship). When weight gain was less than 0.8 kg day−1, the linear relationship between [CP] and weight gain suggests that adult female bison cease gaining weight when [CP] = 54.5 ± 3.4 mg g−1 (r 2 = 0.84, P = 0.001). For yearlings, weight gain also decreased with decreasing adult female [CP] (n = 11, r 2 = 0.90, P < 0.001 for linear relationship). The linear relationship between adult female [CP] and weight gain when weight gain <0.8 kg day−1 suggests that yearlings cease gaining weight when adult female [CP] = 44.8 ± 5.4 mg g−1 (r 2 = 0.92, P = 0.009) (Fig. 6).

Changes in dietary crude protein concentrations ([CP]) over time for adult male (closed symbols) and adult female (open symbols) bison. Dietary quality was determined with fecal near-infrared spectroscopy. Pearson’s correlation between [CP] for males and females in 2014 was 0.97

Relationships between modeled weight gain and dietary crude protein concentrations ([CP]) at different times of year for yearlings (closed symbols, thick line) and adult female (open symbols, thin line) bison. Gray points were not included in the regressions
Discussion
This research successfully demonstrates for the first time the ability of a remote autonomous weighing system to passively generate seasonal patterns of weight gain. Whereas previous approaches only produced changes in weights on annual or monthly time scales and involved stressful handling of the animals, continuous weight trajectories are now possible with minimal effect on the animals beyond the initial tagging. As rates in feeding trials can be off due to handling stress (Koch et al. 1995), this approach may better quantify changes in weight gain, though it cannot account for differences in water or digesta content over time.
As previously observed in the Flint Hills, grass protein concentrations, dietary quality, and weight gain of grazers decline as the growing season progresses (Anderson et al. 1970; Craine et al. 2010; Rao et al. 1973; Raynor et al. 2015). For Konza bison, the declines in dietary quality were associated with declines in weight gain, but the different classes of bison differed in their response to declining dietary quality. For example, adult females with calves reached 98 % of the asymptotic weight on DOY 173 while yearlings continued to gain weight well beyond this, albeit at declining rates. Similar patterns were observed between yearling and adult bighorn sheep (Ovis canadensis) where yearlings continued to gain weight beyond the point where adult weights had leveled off (Festa-Bianchet et al. 1996).
With forage quality declining to near maintenance levels by mid-July, for yearlings to continue to gain weight requires that they either selected higher quality forage than adults, or they received nutritional supplements from their mothers. Smaller grazers are supposed to gain proportionally less, not more, weight on a given quality diet than larger animals (Subcommittee on Beef Cattle Nutrition-Committee on Animal Nutrition-National Research Council 2000). Also, given the broad similarity between adult male and adult female dietary quality (Fig. 5), as well as the fact that yearlings and adult females travel together in herds, yearlings gaining weight over a longer period than adult females is less likely due to selecting higher-quality diets. One hypothesis to test will be maternal nutritional supplementation. Many adult female bison allow yearlings to suckle throughout their second year (Green and Rothstein 1991; Hamel et al. 2012), which could explain their ability to increase in mass despite low forage quality, but more observations, fecal collection, and modeling are required to verify this.
The weight trajectories of the largest males show that patterns of weight gain and loss are not just due to changes in forage quality. Though further monitoring is needed to confirm this, beginning in early July, the largest males apparently lost almost 200 kg during mating season at a time when adult female weights were relatively constant. This is similar in proportion to patterns observed for males and female red deer during rutting season (Yoccoz et al. 2002). These declines appeared to cease after peak mating season was over. Although more tracking of individual males is certainly warranted, this evidence suggests that heightened breeding activity and reduced intake during the mating season must be considered along with nutritional quality for understanding seasonal weight dynamics of adult male bison.
Bison appear to have greater digestive efficiencies than cattle (Hawley et al. 1981; Schaefer et al. 1978), but there is no evidence that they can gain weight better on low-quality forage or worse on high-quality forage (Koch et al. 1995). Based on empirical relationships here, it appears that adult bison can continue to gain weight below what is measured as 60 mg CP g−1, which is generally considered a typical level of [CP] required for maintenance (Van Soest 1994). In addition, bison appear to have less weight gain than expected when [CP] exceed 90 mg CP g−1 based on the relationships between [CP] and weight gain below this threshold. In contrast, cattle are considered to have a largely linear relationship between [CP] and average daily gain between 60 and 160 mg CP g−1 (Subcommittee on Beef Cattle Nutrition-Committee on Animal Nutrition-National Research Council 2000). It is unknown whether weight gain on early season, high-protein forage being less than expected is due to inherent inabilities to process high-quality forage as well as cattle, but changes in body size, metabolic rate, and reproductive status may play roles. The relationship between weight gain and dietary quality may require reexamination of calibration curves between NIRS spectra and dietary quality, which were largely derived from cattle (Roberts et al. 2004; Stuth and Lyons 1995).
Ultimately, one of the main questions that may be answered by utilizing this technique is to how variations in weather and climate affect the weight gain trajectories of bison. This would allow for better prediction of how climate change will affect grazers and better prediction of the effects of weather anomalies and allow for more effective mitigation strategies. For example, in Kansas, bison gain more weight in years when precipitation is high in the late summer, but less weight when precipitation is high in early summer (Craine et al. 2013). Weight trajectories will be necessary across years to understand whether the effect of precipitation at different times of year has an immediate effect on weight gain or whether the effects are delayed as a result of changes in trajectories of forage quality. With this knowledge, any nutritional supplementation strategies, directed animal movements, or timing of prescribed fires could be more effectively managed.
If we are to understand how climate change might affect the growth and reproduction of grazers, no less develop strategies to mitigate effects of climate change on these animals, it will be necessary to compare the weight gain trajectories along climate gradients for different grazers. For example, across the Great Plains, bison gain more weight annually in colder regions than warmer regions (Craine 2013). This would suggest that warming either reduces the length of the period that bison can gain weight, reduces the rate of weight gain during that period, or increases the rate of weight loss outside of the growing season. Examination of cattle diet across these same climate gradients indicates that cattle in warmer locales experience lower maximum and minimum dietary quality compared to colder sites (Craine et al. 2010), suggesting that both a shorter weight gain season and lower rates of weight gain may be in effect. Although this remains to be confirmed, we can expect warmer climates to reduce bison weight by reducing maximum rates of weight gain and the length of time bison have to gain weight. Broader utilization of the remote autonomous weighing systems could test this.
References
Allred BW, Fuhlendorf SD, Hovick TJ, Dwayne Elmore R, Engle DM, Joern A (2013) Conservation implications of native and introduced ungulates in a changing climate. Glob Chang Biol 19:1875–1883. doi:10.1111/gcb.12183
Anderson KL, Smith EF, Owensby CE (1970) Burning bluestem range. J Range Manag 23:81–92
Craine JM (2013) Long-term climate sensitivity of grazer performance: a cross-site study. PLoS ONE 86:e67065. doi:10.1371/journal.pone.0067065
Craine JM, Elmore AJ, Olson KC, Tolleson D (2010) Climate change and nutritional stress in cattle. Glob Chang Biol 16:2901–2911
Craine JM, Towne EG, Tolleson D, Nippert JB (2013) Precipitation timing and grazer performance in a tallgrass prairie. Oikos 122:191–198
Festa-Bianchet M, King WJ, Jorgenson JT, Smith KG, Wishart WD (1996) The development of sexual dimorphism: seasonal and lifetime mass changes in bighorn sheep. Can J Zool 74:330–342
Gates CC, Freese CH, Gogan PJP, Kotzman M (2010) American bison: status survey and conservation guidelines 2010
Green WCH, Rothstein A (1991) Sex bias or equal opportunity? Patterns of maternal investment in bison. Behav Ecol Sociobiol 29:373–384. doi:10.2307/4600634
Hamel S, Cote SD, Festa-Bianchet M (2010) Maternal characteristics and environment affect the costs of reproduction in female mountain goats. Ecology 91:2034–2043
Hamel S, Craine JM, Towne EG (2012) Maternal allocation and offspring characteristics in Bison. Ecol Appl 22:1628–1639
Hawley A, Peden D, Stricklin W (1981) Bison and Hereford steer digestion of sedge hay. Can J Anim Sci 61:165–174
Koch R, Jungt H, Crouset J, Varelt V, Cundifft L (1995) Growth, digestive capability, carcass, and meat characteristics of Bison bison, Bos taurus, and Bos x bison. J Anim Sci 73:1271–1281
Moore JE, Brant MH, Kunkle WE, Hopkins DI (1999) Effects of supplementation on voluntary forage intake, diet digestibility, and animal performance. J Anim Sci 77:122–135
Rao MR, Harbers LH, Smith EF (1973) Seasonal change in nutritive value of bluestem pastures. J Range Manag 26:419–422
Raynor EJ, Joern A, Briggs JM Bison (2015) Foraging responds to fire frequency in nutritionally heterogeneous grassland. Ecology
Ripple WJ, Smith P, Haberl H, Montzka SA, McAlpine C, Boucher DH (2013) Ruminants, climate change and climate policy. Nat Clim Chang 4:2–4
Roberts CA, Stuth J, Flinn P (2004) Analysis of forages and feedstuffs. In: Roberts CA, Workman J, Reeves JB (eds) Near infrared spectroscopy in agriculture. American Society of Agronomy-Crop Science Society of America-Soil Science Society of America, Madison, pp 231–268
Schaefer A, Young B, Chimwano A (1978) Ration digestion and retention times of digesta in domestic cattle (Bos taurus), American bison (Bison bison), and Tibetan yak (Bos grunniens). Can J Zool 56:2355–2358
Stuth JW, Lyons RK (1995) NIRS/NUTBAL nutritional profiling system for grazing land animal management. Ann Zootech 44(Suppl):32
Subcommittee on Beef Cattle Nutrition-Committee on Animal Nutrition-National Research Council (2000) Nutrient requirements of beef cattle: seventh revised edition: update 2000. National Academy Press, Washington, DC
Tieszen LL, Stretch L, Kooi JV (1998) Stable isotopic determination of seasonal dietary patterns in bison at four preserves across the Great Plains. In: Irby L, Knight J (eds) International symposium on bison ecology and management in North America. Montana State University, Bozeman, pp 130–140
Towne EG (1999) Bison performance and productivity on tallgrass prairie. Southwest Nat 44:361–366
Van Soest PJ (1994) Nutritional ecology of the ruminant, 2nd edn. O & B Books, Corvallis
Yoccoz NG, Mysterud A, Langvatn R, Stenseth NC (2002) Age- and density-dependent reproductive effort in male red deer. Proc Biol Sci / R Soc 269:1523–1528. doi:10.1098/rspb.2002.2047
Acknowledgments
This research was supported in part by The National Geographic, The Nature Conservancy, the National Science Foundation (DEB-1342787 to JMC), Konza Prairie Biological Station, and the Konza LTER program (DEB-0823341).
Ethical approval
All procedures performed in studies involving animals were in accordance with the ethical standards of the Kansas State University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Karol Zub
Rights and permissions
About this article

Cite this article
Craine, J.M., Towne, E.G. & Elmore, A. Intra-annual bison body mass trajectories in a tallgrass prairie. Mamm Res 60, 263–270 (2015). https://doi.org/10.1007/s13364-015-0227-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-015-0227-z