
Abstract
Many insects find resources by means of the olfactory cues of general odors after learning. To evaluate behavioral responses to the odor of a particular chemical after learning with reward or punishment quantitatively, we developed a standardized odor-training method in the German cockroach, Blattella germanica (Linnaeus), an important urban pest species. A classical olfactory conditioning procedure for a preference test was modified to become applicable to a single odor, by which a (−)-menthol or vanillin odor was independently associated with sucrose (reward) or sodium chloride solution (punishment). The strength of the association with the odor was evaluated with the increase or decrease in visit frequencies to the odor source after olfactory conditioning. The frequency increased after (−)-menthol was presented with a reward, while it did not change with the rewarded vanillin odor. With both odors, the frequency decreased significantly after training with a punishment. These results indicate that cockroaches learn a single compound odor presented as a conditioned stimulus, although the association of the odor with a reward or punishment depends on the chemical. This olfactory conditioning method can not only facilitate the analysis of cockroach behavior elicited by a learned single chemical odor, but also quantify the potential attractiveness or repellency of the chemical after learning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many insects find resources, such as food, a mate and habitat, by using olfactory cues in the environment. Both attractant pheromones and learned general odors evoke searching behavior in the insects; however, it is still unclear whether both odors induce identical behavior in the orientation toward resources. To evaluate the behavioral responses after learning the odor of a particular chemical quantitatively, the olfactory conditioning procedure, as well as the behavioral test, should be standardized. We adopted the cockroach as a model animal for the study, and modified and developed an olfactory conditioning method applicable to this model.
Since insect pheromones were identified widely in moth species, the distant olfactory behavior of insects has been mainly studied for these specific semiochemicals (e.g., Cardé and Willis 2008). This is also true in cockroaches. Chemo-orientation behavior in the American cockroach, Periplaneta americana (Linnaeus), for its sex attractant pheromone was intensively studied in arenas (Bell and Tobin 1981; Hawkins 1978; Seelinger and Gagel 1985; Takahashi and Kitamura 1972), wind tunnels (Seelinger and Schuderer 1985; Tobin 1981; Willis and Avondet 2005), on a Y-maze lobe (Rust et al. 1976) and a servosphere locomotion compensator (Bell and Kramer 1980). In the German cockroach, Blattella germanica (Linnaeus), the olfactory behavior released by its volatile sex attractant pheromone (Nojima et al. 2005) and aggregation attractant pheromone (Ishii and Kuwahara 1968; Sakuma and Fukami 1985, 1990) has been studied with a choice chamber and olfactometer.
Olfactory behavior in the cockroach has also been studied with respect to its food attractants. Cockroaches are economically important pests, and various bait recipes have been formulated using general odors (e.g., Nalyanya et al. 2001). The German cockroach was shown to be attracted by a food odor (Tsuji 1966; Wileyto and Boush 1983), and even more after learning the odor (Durier and Rivault 2001). Although olfactory learning has been studied in a wide variety of insect species including cockroaches, we chose this species because of its known odor-learning performance, the various pheromones it uses and its importance in urban entomology. A standardized odor-training method is required to evaluate the post-learning attractiveness of a general odor.
Odor learning in the cockroach has been investigated in the American cockroach by operant conditioning procedures, which have revealed the ability of olfactory learning and memory in this species (Balderrama 1980; Sakura and Mizunami 2001). In operant conditioning, cockroaches have free access to a pair of different odors with a respective sugar reward or appetitive US (unconditioned stimulus) and a saline punishment or aversive US; however, this caused intrinsic difficulties in standardizing the learning process. An alternative method was a carefully designed classical conditioning procedure in which the timing of odor-exposure and feeding could be manually controlled by the researcher (Watanabe et al. 2003).
This classical conditioning procedure included differential conditioning trials in which peppermint odor was associated with sucrose solution and vanilla odor was associated with saline solution. Each of the reward and punishment trials was conducted alternately on the same test animal. The learning performance of the animal was then examined by an odor preference test, which allowed test animals to choose between peppermint and vanilla sources, and the percentage of visits to the peppermint source was scored as a preference index.
The classical conditioning procedure, using differential conditioning trials followed by the preference test, indeed provides adequate results for the cognitive study, but it does not specify the absolute potency of the odor in modifying the behavior of the animal after learning. To evaluate the olfactory responses induced after conditioning by the odor, we simplified the procedure to elementary conditioning by associating only one odor with a reward or punishment. Accordingly, the preference test between two odors was replaced with a test of visit frequencies to one odor source before and after training. This study dealt with the simplified version of a classical olfactory conditioning paradigm to establish the learning of a single compound odor in the cockroach.
Materials and methods
Insects
Adult male cockroaches, B. germanica, were obtained from a laboratory colony reared under a 16-h light:8-h dark photocycle at 25–27 °C and 50–70 % relative humidity.
A group of 20–30 animals was used in a series of experiments. Two days before the start of experiments, animals were isolated from the colony and maintained in groups in plastic containers (150 × 105 × 45 mm) without feeding for 2 days to improve their appetitive motivation for food and water. The wall of the container was smeared with liquid paraffin to prevent a test animal from escaping. The bottom of the container was lined with a sheet of filter paper.
Training
The training method was modified from classical olfactory conditioning used for the American cockroach, P. americana (Watanabe et al. 2003). Training (Tr) and testing (T) were all carried out in a light period under a dim red light (Fig. 1). A test animal was transferred into a glass cup (52 i.d. × 45 mm) and acclimatized for 30 min. Subsequently, the animal received training to associate an odor with a reward or punishment. (−)-Menthol and vanillin were used as the olfactory conditioned stimuli (CS) or odors. Ten % sucrose and 20 % sodium chloride (saline) solutions were used as appetitive (US+) and aversive (−US) unconditioned stimuli or as a reward and punishment, respectively (US as whole unconditioned stimuli). All chemicals were purchased from Wako Pure Chemical Industries, Ltd., Tokyo, Japan.

Stimulus arrangement for classical conditioning in the German cockroach. A test animal received training in a glass cup to associate an odor (CS) with a reward or punishment (US). A 1-ml syringe with a filter paper disc on its needle, scented with vanillin or (−)-menthol CS (a), was placed within 1 cm of the animal’s head, and after 2 s from the onset of odor presentation, a droplet of sucrose or saline solution US was presented from the needle tip to the mouth part of the animal for 2 s (b). The cup was then closed with a lid after odorous air had been removed with a desktop ventilation hood (Fig. 3). The animal was maintained in the cup until the next behavioral test. Also refer to the text
A 1-ml hypodermic syringe (Terumo Corp., Tokyo, Japan) with a scented filter paper disc (7 mm o.d., no. 1, Advantec Toyo Kaisha, Ltd., Tokyo, Japan) on its needle presented CS and US to the animals. The filter paper disc was soaked with 10 μl solution of either vanillin (150 mg/ml) or menthol (150 mg/ml) and then air-dried before use. Syringes used for appetitive conditioning and those for aversive conditioning were filled with sucrose and saline solutions, respectively. For the conditioning trial, the scented filter paper disc attached to the needle was placed within 1 cm of the cockroach’s head, and after 2 s from the onset of odor presentation, a droplet of sucrose or saline solution was presented to the mouth part of the cockroach for 2 s. Then the cup was closed with a lid after odorous air had been removed with a desktop ventilation hood (320 × 260 × 235-mm opening). The animal was maintained in the cup until the next behavioral test.
In the differential conditioning procedure, appetitive and aversive conditioning trials were carried out alternately, in which menthol CS was associated with sucrose US + and vanillin CS was associated with saline −US (Fig. 2a). In elementary conditioning procedures, appetitive or aversive conditioning trials were individually replicated several times (Fig. 2b). In the control experiment for CS, we presented a menthol or vanillin odor to cockroaches without pairing with US, which we referred to as unpaired CS presentation (Fig. 2c). The possible participation of non-associative learning, such as sensitization and habituation, was also examined by presenting sucrose or saline solution to animals without pairing with CS (Fig. 2c). The inter-trial interval (ITI) was fixed at 5 min for all experiments.

Timetables for training and tests in the olfactory conditioning experiments. a Differential conditioning. Preference tests (open columns) were conducted twice: 30 min before (T-0) and then 30 min (T-30 min), 1 day (T-1d), 2 days (T-2d) or 3 days (T-3d) after training (Tr: solid column). The menthol preference index (MPI) was obtained in each test from the number of visits to a pair of odor sources. The bottom array shows that appetitive- and aversive-conditioning trials were conducted alternately three times in which menthol CS (horizontally striped) was associated with sucrose US+ (open square), and vanillin CS (oblique striped) was associated with saline −US (solid square). The inter-trial interval (ITI) was fixed at 5 min in all experiments. The top array shows photo- (open) and scoto-phases (solid) of 16 L:8 D photoperiods. Tests were conducted from 3 h after light-on. b Elemental conditioning. In both elemental conditioning and unpaired presentation experiments, the number of visits of a test animal to the odor source was compared 30 min before (T-0) and 30 min after (T-30 min) training (Tr). Appetitive and aversive conditioning trials were independently conducted in which a CS odor, menthol or vanillin, was associated with sucrose US+ or saline −US. Trials were repeated various times for each of the four combinations (examples of the presentation trial six times are shown). c Unpaired presentation of CS or US. Only menthol or vanillin odor, or sucrose or saline solution was presented to a test animal in an unpaired presentation experiment. Odor presentation trials were repeated six times, whereas sucrose or saline solution was presented once, twice and four times to examine the effect of the number of trials (examples of the presentation trial, four times, are shown)
Symbols for stimulus categories and those for other terms in the classical conditioning procedure are listed together with stimulus components used in the present study (Table 1).
Behavioral test
The searching behavior of a test animal for an odor source(s) was observed 30 min before and after training. A rectangular plastic container (150 × 105 × 45 mm) was used as a test chamber (Fig. 3). Before testing, the test animal was introduced into the test chamber to become familiarized for 5 min. A glass dish (15 o.d. × 5 mm) with a scented filter paper disc in it was used as the odor source. The same paper disc as that used in training was impregnated with an odorant chemical (1.5 mg) and then placed in the bottom center of the dish. The dish was then covered with perforated aluminum foil and placed in the chamber to start the test. During the test, the chamber was placed in the ventilation hood, which drew the air over the chamber at ca. 25 cm/s.

Schematic drawing of the odor source and test chamber. A plastic letter case was used as a test chamber (center). After familiarizing the animal for 5 min in the chamber, an odor source or a pair of sources was placed in its center to start the test. The odor source was a glass dish covered with perforated aluminum foil in which a scented filter paper disc was set on the bottom (right). During the test, the chamber was placed in the ventilation hood (left), which gently drew air over the chamber. The visit frequencies of the test animal to the odor source or a pair of sources were counted during a 10-min test period. Also refer to the text for details
The number of visits of the test animal to an odor source or a pair of sources was counted during a 10-min period. After each visit, the odor source(s) was randomly rearranged. An odor source was considered to have been visited when the cockroach probed the odor source with its antennae, mouthparts and foreleg tarsi. A single odor source was placed in the test chamber for a visit-frequency test, whereas two sources of different odors were used for a preference test between two odors. The odor source and the filter paper floor lining were changed at every test to avoid odor contamination. All experiments were carried out at 25 ± 2 °C, 50–70 % RH, from 3 h after light-on.
Data analysis
Preference data
In the preference test, the number of visits to a pair of odor sources was transformed to a preference index. The menthol preference index (MPI) denoted the odor preference for menthol over vanillin by
where F m and F v are the frequencies of visits to menthol and vanillin sources during the odor preference test, respectively. MPI was defined after the peppermint preference index in a previous study (Watanabe et al. 2003). We treated MPI as quantitative data by following the preceding data analysis. The initial odor preference of untrained cockroaches was evaluated by the Z-test. Wilcoxon’s signed-rank test (WSR test) was used to compare MPI values before and after training in a given cockroach group.
Visit-frequency data
The frequency of visits to an odor source was compared before and after training for the same animals in a given group by the WSR test. The Kruskal-Wallis (KW) test and Scheffé post hoc test were also used to compare the frequency of visits between different groups of animals.
Statistical tests were conducted with JMP 8.0 software (SAS Institute, Cary, NC, USA), except for the Z-test, with Microsoft Excel 2007 software (Microsoft Corp., WA, USA).
Results
Behavior of test animals
After being introduced into the test chamber, a cockroach continued walking along the wall interspersed with short stops. During these stops, it waved its antennae to catch the airflow, then turned and walked toward the odor source after detecting an odor. The animal may have accidentally touched an odor-source dish sometimes; however, a visit to the odor source was distinguishable by its probing behavior. When it visited the odor source, it often climbed with its forelegs onto the top lid of the dish and probe holes for odor with its antennae and palpi. This was typically observed in test animals after rewarded conditioning for the odor. Test animals occasionally became immobile, particularly in a test after training with sucrose alone as well as with saline. A decrease in the frequency of visits measured may have been the result of a long halt.
Odor preference of untrained German cockroaches
Test animals were made to choose between menthol and vanillin odor sources to obtain menthol preference indices (MPI) from the number of visits. The distribution of the MPI of untrained animals is shown in a histogram (Fig. 4). The average MPI was less than 50 % (T-0, Z-test, T = 7.6986, df = 128, p < 0.001), which indicates that untrained German cockroaches exhibited a significant preference for vanillin over menthol.

The odor preference of untrained German cockroaches. The histogram shows the distribution of the menthol preference index for each individual in an untrained group of cockroaches. Number of cockroaches is indicated in parentheses
Retention of olfactory memory in cockroaches after differential conditioning
Animals were subjected to three sets of a differential associative conditioning trials, each consisting of an appetitive conditioning trial followed by an aversive conditioning trial with an inter-trial interval (ITI) of 5 min. Menthol CS was associated with a 10 % sucrose solution US + and vanillin CS was associated with a 20 % saline solution −US in the appetitive and aversive conditioning trials, respectively (Fig. 2a). Test animals were subjected to three differential conditioning trials consecutively with a 5-min ITI.
Four groups of animals were examined with different delay periods after the trial (Fig. 2a). One was subjected to the odor preference test 30 min after the trial. Others were kept in groups for 1, 2 and 3 days and were fed only with water daily for 1 h until the day before the test. We did not examine animals for longer than 3 days because moribund males were almost immobile in a test after being dehydrated for 4 days. The retention of memory was examined by comparing the MPI of the same animal before and after the conditioning trial. The frequencies of the MPI of trained animals are shown in histograms (Fig. 5). Thirty min and 1 day after conditioning, animals exhibited a significantly greater preference for menthol than before training (WSR, T-30 min: W = 926, p < 0.0001; T-1d: W = 193, p < 0.01), whereas the preference for menthol did not differ from that before conditioning at 2 and 3 days (WSR, T-2d: W = 14, p = 0.7905; T-3d: W = 91, p = 0.1992). These results show that the retention of olfactory memory conditioned with paired odor lasted for 1 day in the German cockroach.

Retention of olfactory memory in German cockroaches. Four groups of cockroaches were subjected to three sets of differential conditioning trials in which menthol and vanillin odors were associated with sucrose and saline solutions, respectively (Fig. 2a). Odor preference was tested 30 min (T-30 min), 1 day (T-1d), 2 days (T-2d) and 3 days (T-3d) after conditioning, respectively. The distribution of the menthol preference index (MPI) for each individual is shown. Number of test animals of each test is indicated in parentheses
Visits of untrained cockroaches to an odor source
The visit frequency of an untrained test animal to an odor source was compared with that to an odorless object, i.e., a glass dish with a filter paper disk treated only with the solvent methanol. Animals visited the odorless object 1.5 times on average in a 10-min observation period (Fig. 6). The odor applied to the source significantly influenced the frequency of visits (KW, H = 65.5146, df = 2, p < 0.01). Cockroaches visited the vanillin odor source more than the odorless object (Scheffé, p < 0.01), whereas no significant difference was observed between the frequency of visits to the menthol odor source and to the odorless object (Scheffé, p = 0.2125).

Visit frequency of untrained cockroaches to an odor source. Menthol and vanillin were used as the odorous materials. The visit frequency to an object without odorous material in it is also shown. The bar indicates the mean ± SE visit frequency. The number of animals of each test is indicated in parentheses. Different letters indicate significant pairwise differences (Scheffé test after the KW test)
Effect of unpaired presentations of CS or US
The olfactory and gustatory stimuli used as CS and US in the present experiments may modify the behavior of animals by habituation, sensitization or other non-associative learning processes. We conducted the same training as in the following associative conditioning process, except we replaced either the CS or US presentation with a sham operation.
Groups of animals were presented with a menthol or vanillin odor six times with an interval of 5 min. They were examined for their visit frequency to the source of the same odor 30 min before and after training. Figure 7a shows that animals trained with menthol visited the odor source as frequently as before (WSR, W = 9, p = 0.8383), whereas those trained with vanillin visited the odor source significantly less than before (WSR, W = 747, p < 0.01).

Effect of unpaired presentation of CS or US. a Visit frequencies to a menthol or vanillin odor source before (open column) and after training (filled column) with the same odor alone. CS presentations with odor were conducted six times with a 5-min ITI. b Visit frequencies before and after training with the saline solution alone. US presentations were conducted once, twice and four times in the different groups of animals. All the groups were tested with menthol. c Visit frequencies before and after training with sucrose solution alone. Refer to notes in b for details. The bar indicates the mean ± SE visit frequency. The number of animals of each test is indicated in parentheses. The stimuli are indicated at the top of the figure. The results of statistical comparison before and after conditioning (WSR test) are shown as asterisks (*p < 0.05; **p < 0.01; NS p > 0.05)
Other groups of animals were trained with either sucrose or saline solution without the presentation of an odor and were tested for the frequency of visits to the menthol odor source 30 min before and after training. They were subjected to one, two and four presentations of US with an interval of 5 min. Animals fed with saline visited the odor source as frequently as before irrespective of the repetition of US presentations (Fig. 7b, WSR, 1-trial: W = 53, p = 0.1066; 2-trials: W = 7, p = 0.8395; 4-trials: W = 51, p = 0.1272), whereas they visited less after more than two US presentations fed with sucrose solution (Fig. 7c, WSR, 1-trial: W = 3, p = 0.9470; 2-trials: W = 95, p < 0.01; 4-trials: W = 78, p < 0.05). It was also observed that when sucrose was presented more than twice, 58 % of the cockroaches halted and remained still for more than 5 min in the 10-min test period.
Effect of the number of trials on elemental conditioning with a single chemical
Groups of cockroaches were subjected to elemental conditioning trials in which a single odor was associated with a reward or punishment. We used the visit frequency of the animal to a trained odor source to evaluate olfactory memory. The frequency of visits 30 min after training was compared with that before.
We conducted experiments for all combinations between the two US and two CS independently: US+ (sucrose reward) with menthol CS, −US (saline punishment) with menthol CS, US+ with vanillin CS and −US with vanillin CS. In order to examine the effect of the number of conditioning trials, groups of cockroaches were subjected to various sets of elemental conditioning trials with a 5-min ITI.
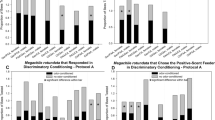
Cockroaches visited the menthol odor source more than before after four and six trials of conditioning of the odor with a sucrose reward, although no significant difference was observed in the number of visits before and after training in the 1-, 2- and 3-trial groups (Fig. 8a; WSR, 1-trial: W = 123, p = 0.0564; 2-trials: W = 12, p = 0.822; 3-trials: W = 63, p = 0.3457; 4-trials: W = 193, p < 0.01; 6-trials: W = 204, p < 0.01). After aversive conditioning with saline, they visited the menthol odor source less than before, except for the 1-trial group (Fig. 8b; WSR, 1-trial: W = 110, p = 0.0709; 2-trials: W = 154, p < 0.05; 3-trials: W = 214, p < 0.05; 6-trials: W = 297, p < 0.05).

Effect of the number of trials for conditioning with menthol. a Appetitive conditioning, in which menthol odor was associated with a sucrose reward. Conditioning trials were conducted once, twice, three, four and six times in the different groups of animals. The stimuli are indicated at the top of the figure. All the groups were tested with menthol. The visit frequencies of animals to the odor source 30 min before (open column) and after (filled column) training are shown as mean ± SE. The numbers of animals are indicated in parentheses. b Aversive conditioning, in which the menthol odor was associated with a saline punishment. Conditioning trials were conducted once, twice, three and six times in the different groups of animals. Refer to notes in a for details. The results of statistical comparison before and after conditioning (WSR test) are shown as asterisks (*p < 0.05; **p < 0.01; NS p > 0.05)
They did not visit the vanillin odor source more than before after appetitive conditioning of the odor with sucrose in all groups including the adult female group (Fig. 9a; WSR, 1-trial: W = 87, p = 0.0768; 2-trials: W = 136, p = 0.0783; 4-trials: W = 22, p = 0.7713; 6-trials: W = 9, p = 0.9102; 8-trials: W = 150, p = 0.1027; female, 6-trials: W = 72, p = 0.2744). In contrast, they visited the vanillin odor source much less than before after more than three trials of aversive conditioning with saline (Fig. 9b; WSR, 2-trials: W = 123, p = 0.0557; 3-trials: W = 141, p < 0.01; 4-trials: W = 333, p < 0.01; 6-trials: W = 269, p < 0.01).

Effect of the number of trials for conditioning with vanillin. a Appetitive conditioning, in which the vanillin odor was associated with a sucrose reward. Conditioning trials were conducted once, twice, four, six and eight times in the different groups of animals. The stimuli are indicated at the top of the figure. All groups were tested with vanillin. The visit frequencies of animals to the odor source 30 min before (open column) and after (filled column) training are shown as mean ± SE together with those of females subjected to a trial six times. The numbers of animals are indicated in parentheses. b Aversive conditioning, in which the vanillin odor was associated with a saline punishment. Conditioning trials were conducted twice, three, four and six times in the different groups of animals. Refer to notes in a for details. The results of statistical comparison before and after conditioning (WSR test) are shown as asterisks (**p < 0.01; NS p > 0.05)
Discussion
Studies of olfactory learning require manageable experimental conditions. A classical conditioning procedure allows the experimenter to control manually the sensory experience of the test animal, in particular the contiguity of conditioned stimulus and unconditioned stimulus presentation to regulate the responsiveness of the animal to the learned conditioned stimulus (Rescorla 1988). In the present study, we reconfirmed the classical olfactory conditioning procedure that associates an odor with a taste in the cockroach and modified it to allow an evaluation of attractiveness demonstrated by a single compound odor after olfactory conditioning.
The procedure used in a previous study was developed for neuronal research into learning in the American cockroach and was designed for that purpose (Watanabe et al. 2003). First, we re-examined whether this olfactory conditioning procedure was applicable to the German cockroach, an important urban pest whose odor-learning ability has been demonstrated (Durier and Rivault 2001). By differential conditioning trials, in which sucrose US+ (reward) with menthol odor CS and saline −US (punishment) with vanillin odor CS were presented alternately, male German cockroaches demonstrated a high capability of olfactory learning 30 min after three trials of differential conditioning with a 5-min ITI and retained this memory for at least 24 h (Fig. 5). These results are almost consistent with those obtained in American cockroaches and two-spotted crickets trained by similar procedures (Matsumoto and Mizunami 2000, 2002; Watanabe et al. 2003). We concluded that the German cockroach formed olfactory memory by a classical conditioning procedure and could be used in studies of learning and memory.
In order to determine behavioral changes after training, we modified both the conditioning procedure and test used in a previous study. Instead of differential conditioning associating two odors with both a reward and punishment, we presented one odor associated with either a reward or punishment. We used pure chemicals, instead of mixtures, which have been used in the past, to simplify the chemical composition of the odorous material. In previous studies, elemental conditioning using only one odor with a reward or punishment formed olfactory memory showing a preference between two odors in two-spotted crickets (Unoki et al. 2005). In the present study, we counted the number of visits of the test animal to the odor source to conform to previous studies, although we did not use a preference test by pairing the odor with an additional odor. The preference test is beneficial for extracting odor-preference elements by canceling irrelevant factors. Instead, we compared the number of visits including various elements before and after training, which enabled us to analyze the elemental behavior of test animals modified after olfactory conditioning.
We can expect a specific behavioral response from an animal for a pheromone odor according to the message it carries, whereas the response to a general odor from a single chemical without training is unpredictable and mostly limited. In practice, of 30 substances, only one showed significant attractiveness to the German cockroach by competing well with a control food material (Wileyto and Boush 1983). Untrained males exhibited menthol preference indices (MPI) significantly lower than 50 % (Z-test, p < 0.05), indicating an innate preference for vanillin over menthol in German cockroaches (Fig. 4). This is consistent with the response observed in the American cockroach to vanilla and menthol (peppermint) essences (Balderrama 1980; Sakura and Mizunami 2001). We then examined the behavioral response to a single odor source. Before training, males visited the menthol odor source no more frequently than the non-odored control source, whereas they visited the vanillin odor source 2.3 times more than the control source (Fig. 6). These results indicate that unconditioned German cockroaches did not behaviorally respond to menthol, but they did to vanillin.
The materials used as CS or US could have primarily altered animal behavior through non-associative learning processes, such as sensory adaptation, habituation or sensitization after training and even testing (Thompson and Spencer 1966). We compared the number of visits before and after the presentation of an odor without a sucrose reward and saline punishment. Cockroaches presented with the menthol odor almost equally visited the menthol odor source before and after training (Fig. 7a). However, after being trained with vanillin, they visited the vanillin odor source significantly less than before. It seems possible that the innate response of cockroaches to the vanillin odor decreased after olfactory training by habituation. It may also be possible that cockroaches, which have been conditioned to the vanillin odor in the laboratory colony, lost responsiveness to the odor by extinction after unrewarded conditioning trials. Latent or conditioned inhibition (blocking phenomenon) could have been involved in this result (Mackintosh 1983; Papini and Bitterman 1993; Rescorla 1988). In contrast, cockroaches visited the menthol odor source as frequently as before. We can use (−)-menthol as a relatively neutral odor in a cockroach learning study.
The local search activity of the test animal could have been influenced by imbibing sucrose or saline solutions, i.e., the number of visits could be enhanced by sensitization or reduced by habituation and/or suppression. Figure 7c shows that the number of visits to the menthol odor source significantly decreased after odorless training with sucrose solutions. We could not observe the sensitization effect observed in a hungry fly continuing a local search after a single touch of sugar (Dethier 1957). Instead, the cockroach was slightly more immobile after imbibing sucrose solution, which caused a decrease in the number of visits. Odorless training with saline solution did not alter the mobility of test animals, and the number still remained at the same level (Fig. 7b).
We presented a single compound to animals as an olfactory conditioning stimulus, which enabled us to specify the compound responsible for memory formation. Once test animals had been conditioned by the menthol odor with a sucrose reward, they became responsive to the odor (Fig. 8a), whereas they did not visit the vanillin odor source more than before after appetitive conditioning with vanillin (Fig. 9a). The increased response after learning may have been cancelled out with non-associative elements, such as decreased visits by presenting sucrose (Fig. 7c) and/or by repeating vanillin odor exposure (Fig. 7a). It is suggested that cockroaches are capable of forming olfactory memory with a general odor by appetitive conditioning, whereas learning potential may be influenced by hidden non-associative learning elements depending on the chemical. Nevertheless, we chose an elemental conditioning procedure rather than a differential one because our interest was olfactory behavior after learning, which is also useful for this application.
Conditioning trials with saline punishment apparently established aversive memory by both menthol and vanillin odor presentations (Figs. 8b, 9b). The numbers of visits to the odor source after six trials were almost comparable to the number of accidental visits counted in the odorless sources (KW, p = 0.217). It is still possible that animals simply neglect or could not access the odor source instead of forming aversive memory to the odor. After training with the saline punishment, animals sometimes stopped for a long time irrespective of odor presentation. Further pharmacological studies by blocking the aversive conditioning system (Unoki et al. 2005) may discriminate each contribution to the lowered visit frequency. The number of visits to the vanillin odor source was further decreased, possibly because of the additional decrease observed after unrewarded training in the second test (Fig. 7a).
In the present study we quantified olfactory memory formation by counting the number of visits of test animals to an odor source. Although the number of visits depended on the chemo-orientation or searching behavior of animals modified by olfactory memory, what behavioral components are responsible for the increase and decrease in the number of visits is still unknown. After training, test animals may simply modulate ortho- and klino-kinesis components to change mobility and diffusion, or they may direct themselves toward the odor source by employing taxis components of various modalities (Bell and Tobin 1982; Cardé and Willis 2008; Kennedy 1978). In more complicated systems, cockroaches may learn odors in combination with other cues. They seem to learn specific food odors in combination with visual cues and locate themselves to the food resources (Durier and Rivault 2001). Including multiple cue systems, the combined use of classical conditioning of test animals and quantitative analysis of locomotion measures under controlled cue presentation (e.g. Sakuma 2002) may clarify the causation of olfactory behavior evoked by a learned general odor. The German cockroach is a suitable animal for behavioral studies.
References
Balderrama N (1980) One trial learning in the American cockroach, Periplaneta americana. J Insect Physiol 26:499–504
Bell WJ, Kramer E (1980) Sex pheromone-stimulated orientation of the American cockroach on a servosphere apparatus. J Chem Ecol 6:287–295
Bell WJ, Tobin TR (1981) Orientation to sex pheromone in the American cockroach: analysis of chemo-orientation mechanisms. J Insect Physiol 27:501–508
Bell WJ, Tobin TR (1982) Chemo-orientation. Biol Rev 57:219–260
Cardé RT, Willis MA (2008) Navigational strategies used by insects to find distant, wind-borne source of odor. J Chem Ecol 34:854–866
Dethier VG (1957) Communication by insects: physiology of dancing. Science 125:331–336
Durier V, Rivault C (2001) Effects of spatial knowledge and feeding experience on foraging choices in German cockroaches. Anim Behav 62:681–688
Hawkins WA (1978) Effects of sex pheromone on locomotion in the male American cockroach, Periplaneta americana. J Chem Ecol 4:149–160
Ishii S, Kuwahara Y (1968) Aggregation of German cockroach (Blattella germanica) nymphs. Experientia 24:88–89
Kennedy JS (1978) The concepts of olfactory ‘arrestment’ and ‘attraction’. Physiol Entomol 3:91–98
Mackintosh NJ (1983) Conditioning and associative learning. Oxford University Press, London
Matsumoto Y, Mizunami M (2000) Olfactory learning in the cricket Gryllus bimaculatus. J Exp Biol 203:2581–2588
Matsumoto Y, Mizunami M (2002) Temporal determinants of long-term retention of olfactory memory in the cricket Gryllus bimaculatus. J Exp Biol 205:1429–1437
Nalyanya G, Liang D, Kopanic RJ, Schal C (2001) Attractiveness of insecticide baits for cockroach control (Dictyoptera: Blattellidae) laboratory and field studies. J Econ Entomol 94:686–693
Nojima S, Schal C, Webster FX, Santangelo RG, Roelofs WL (2005) Identification of the sex pheromone of the German cockroach, Blattella germanica. Science 307:1104–1106
Papini MR, Bitterman ME (1993) The two test strategy in the study of inhibitory conditioning. J Exp Psychol 19:342–352
Rescorla RA (1988) Behavioral studies of Pavlovian conditioning. Ann Rev Neurosci 11:329–352
Rust MK, Burk T, Bell WJ (1976) Pheromone-stimulated locomotory and orientation responses in the American cockroach. Anim Behav 24:52–67
Sakuma M (2002) Virtual reality experiments on a digital servo sphere: guiding male silkworm moths to a virtual odour source. Comput Electron Agric 35:243–254
Sakuma M, Fukami H (1985) The linear track olfactometer: an assay device for taxes of the German cockroach, Blattella germanica (L.) (Dictyoptera: Blattellidae) toward their aggregation pheromone. Appl Entomol Zool 20:387–402
Sakuma M, Fukami H (1990) The aggregation pheromone of the German cockroach, Blattella germanica (L.) (Dictyoptera: Blattellidae): isolation and identification of the attractant components of the pheromone. Appl Entomol Zool 25:355–368
Sakura M, Mizunami M (2001) Olfactory learning and memory in the cockroach, Periplaneta americana. Zool Sci 18:21–28
Seelinger G, Gagel S (1985) On the function of sex pheromone components in Periplaneta americana: improved odour source localization with periplanone-A. Physiol Entomol 10:221–234
Seelinger G, Schuderer B (1985) Release of male courtship display in Periplaneta americana: evidence for female contact sex pheromone. Anim Behav 33:599–607
Takahashi S, Kitamura C (1972) Bioassay procedure of the sex stimulant of the American cockroach, Periplaneta americana (L.) (Orthoptera: Blattidae). Appl Ent Zool 7:133–141
Thompson RF, Spencer WA (1966) Habituation: a model phenomenon for the study of neuronal substrates of behavior. Psychol Rev 73:16–43
Tobin TR (1981) Pheromone orientation: role of internal control mechanisms. Science 214:1147–1149
Tsuji H (1966) Attractive and feeding stimulative effect of some fatty acids and related compounds on three species of cockroaches. Jpn J Sanit Zool 17:89–97
Unoki S, Matsumoto Y, Mizunami M (2005) Participation of octopaminergic reward system and dopaminergic punishment system in insect olfactory learning revealed by pharmacological study. Eur J Neurosci 22:1409–1416
Watanabe H, Kobayashi Y, Sakura M, Matsumoto Y, Mizunami M (2003) Classical olfactory conditioning in the cockroach, Periplaneta americana. Zool Sci 20:1447–1454
Wileyto PE, Boush MG (1983) Attraction of the German cockroach, Blattella germanica (Orthoptera: Blattellidae), to some volatile food components. J Econ Entomol 76:752–756
Willis MA, Avondet JL (2005) Odor-modulated orientation in walking male cockroaches Periplaneta americana, and the effects of odor plumes of different structure. J Exp Biol 208:721–735
Acknowledgments
We express our sincere gratitude to Prof. Mizunami of the Graduate School of Life Science, Hokkaido University, Japan, for the invaluable comments and critical reading of this manuscript. We also thank two anonymous reviewers for their helpful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Liu, JL., Sakuma, M. Olfactory conditioning with single chemicals in the German Cockroach, Blattella germanica (Dictyoptera: Blattellidae). Appl Entomol Zool 48, 387–396 (2013). https://doi.org/10.1007/s13355-013-0199-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13355-013-0199-x